
Abstract
The submerged and floating plant communities in floodplain wetlands of the Upper Columbia River have never been described. To explore mechanisms behind the influence of the annual flood pulse on vegetation, we investigated how species group into flood response guilds whose distributions vary along a connectivity gradient between 44 floodplain wetlands and the river, how connectivity influences water and sediment, and to what degree the effect of connectivity on vegetation is mediated by its effects on these environmental variables. We characterised assemblages with cluster and indicator species analysis, as well as non-metric scaling ordination and tested a structural equation model, which defined the relationship between assemblage composition, sediment and water quality, and connectivity to the river. We found four assemblages, each associated with different water and sediment conditions, and positioned at differing degrees of connectivity. The model provided a good fit to the data. We conclude that in highly connected floodplain wetlands the direct effect of flooding supersedes the influence of water and sediment quality in structuring vegetation assemblages. Yet where flooding is less intense, these environmental variables resume their structuring role.
Similar content being viewed by others

Introduction
The Upper Columbia River floodplain is characterized by a natural flood pulse that structures the ecology of the floodplain and is a rare example of a North American river system with relatively little disturbance from human infrastructure. The floodplain consists of open water wetlands, which constitute a critical component of the Pacific Flyway, and is recognized by RAMSAR as a wetland of international importance. The open-water portions of these floodplain wetlands are characterized by assemblages of floating and submerged aquatic vegetation, which provide critical habitat and other resources to fish, invertebrates, amphibians, and birds. However, the dynamics governing these aquatic plant assemblages are not well understood. In fact, they have never before been researched. Floodplain plant communities are known to respond to the frequency, duration, timing, and depth of flooding as well as the period between flood events (Nielsen and Chick 1997; Blanch et al. 1999; Casanova and Brock 2000; Robertson et al. 2001; Greet et al. 2011), such that species can assemble into response guilds characterized by their adaptations to tolerate or avoid flooding (Merritt et al. 2010). Flood-related factors may vary based on the manner by which wetlands are connected to the river. Some wetlands are isolated except during the river’s flood pulse (sensu Junk et al. 1989), whereas others remain continually connected through breaks in the river channel’s levees (Filgueira-Rivera et al. 2007). This creates a gradient in connectivity to the river and water residence time (Hermoso et al. 2012), which we predict will correspond with the degree to which the river’s flood pulse influences wetland vegetation (Bornette et al. 2008) and the spatial distribution of flood response guilds.
River restoration is growing in importance given that human efforts to regulate flow and stabilize river channels during the 20th century have impacted most large North American rivers (Bayley 1995), with consequent disruption to natural disturbance regimes, truncation of environmental gradients, and severing of connections between rivers and their floodplains (Ward 1998). To restore degraded floodplains and mitigate the effects of human infrastructure, it is critical that we understand the mechanisms by which the natural flood pulse structures floodplain wetland plant communities.
The relationship between large rivers and the vegetation in their floodplain wetlands is complex and nonlinear (e.g., Amoros and Bornette 1999). In floodplain wetlands of the Athabasca River in Jasper National Park, semi-isolated wetlands supported less productive (Bayley and Guimond 2009) but more diverse (Bayley and Guimond 2008) plant life than either highly connected or isolated wetlands. Non-linear responses are common in floodplain wetland studies because flooding acts as a disturbance (sensu Grime 1973) that also influences the pattern and intensity of biological interactions (Riis and Biggs 2001; Bornette et al. 2008; Hermoso et al. 2012). Connectivity may be confounded with successional dynamics (Hodgson et al. 2009), e.g., a highly connected site may be regularly reverted to an early successional stage if accumulated organic sediments, ions, and nutrients as well as established vegetation are scoured out by the annual flood pulse. These changes constitute stresses acting on plant communities that are likely to influence competition among species (e.g., McCreary 1991; Bornette et al. 1998; Lenssen et al. 1999). In contrast to the associated physical disturbance, flooding may also supply inorganic nutrients, silt, and plant propagules (Heiler et al. 1995) which are beneficial to plant communities. For example, working on the Elbe River, Leyer (2006) notes the importance of hydrochory as a dispersal mechanism for floodplain vegetation, with the consequence that isolated wetlands faced dispersal limitation. Thus, river water may have both beneficial and detrimental effects on plants (Amoros and Bornette 1999). The sum of these contradictory effects may appear much less influential than if they were considered independently.
Researchers have tested theoretical relationships between connectivity and plant diversity in numerous studies (Ward and Tockner 2001), but the theoretical relationship between connectivity and plant community composition is less well developed (Riis and Biggs 2001; Barrett et al. 2010). Furthermore, the mechanisms behind the influence of connectivity on floating and submerged aquatic vegetation remain unclear because the direct and indirect effects of connectivity on plant communities are often confounded. Some relationships could be predicted based on the life history strategies of different species: e.g., rapid colonists should persist where physical disturbance is common and competitively dominant species should succeed where nutrients are plentiful (Grime 1973). Species adapted to flooding can form a flood tolerant response guild (sensu Merritt et al. 2010), whereas those sensitive to high rates of flow or intolerant of turbid, cold flood waters could be flood sensitive. The autecology of species will determine the environmental conditions under which they survive. Variables including salinity, alkalinity, water clarity, and water and sediment nutrient levels have been shown in prior studies to influence the distribution of aquatic vegetation (Chambers and Kalff 1987; Koch 2001; Lacoul and Freedman 2006). We hypothesize that increased connectivity to the Upper Columbia River will result in reduced nutrients, salinity, alkalinity, and sediment organic matter because of the river’s scouring action, but increased turbidity because of the river’s high sediment load. The degree to which these environmental conditions are affected by river connectivity will constitute some portion of the net effect of river connectivity on aquatic plant communities. Thus, the net effect of connectivity can be decomposed into an indirect portion, mediated by its influence on these specified environmental conditions, and a direct portion, including the physical disturbance imposed by the flood pulse and associated scouring as well as any influence of connectivity on plant dispersal (Leyer 2006) or seed bank composition (Abernethy and Willby 1999). Presumably, both indirect and direct components will combine to affect which flood response guild survives best in which wetland. The relative contribution of these two components (direct and indirect) to the influence of river connectivity on aquatic plant community composition has never before been addressed.
Our general objective was to explore the role of connectivity in shaping floating and submerged aquatic vegetation communities in floodplain wetlands of the Upper Columbia River. Specifically, we sought to: 1) characterize the floating and submerged vegetation communities across a gradient in connectivity to the river and explore their assembly into flood response guilds; 2) confirm whether connectivity to the river reduces water clarity through inputs of turbid water, and reduces salinity, alkalinity, nitrogen, and sediment carbon levels by scouring; and 3) compare the indirect effects of connectivity as mediated by its effects on these water and sediment variables with any direct (residual) effects on floating and submerged aquatic vegetation.
Methods
Study Location
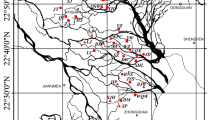
The Columbia River begins in Columbia Lake and flows northwest between the Rocky and Purcell Mountains (Fig. 1). Discharge in the Upper Columbia River fluctuates seasonally in response to rainfall and snowmelt. Glacial melt-water constitutes only a minor component of the total discharge (Makaske et al. 2009); snowpack melt from the mountain ranges constitutes the major source of water to the river, and has recently been in decline due to a combination of anthropogenic warming and decadal variability (Pederson et al. 2011). Above the Nicholson, British Columbia gauging station the river’s drainage basin covers 6,660 km2, with a mean annual discharge of 107 m3.s−1 and an average peak discharge of 436.1 m3.s−1 during the June-July flood pulse.

Map of the study region, indicating the location of the 44 sampled wetlands between the towns of Invermere and Golden along the Upper Columbia River. Grey rectangle in inset depicts location of study region within British Columbia
Between the towns of Invermere and Golden, British Columbia, the river is relatively unaffected by human infrastructure and flows through a floodplain about 1.5 km wide (Fig. 1). The Upper Columbia River is anastomosing, with a gentle slope (average = 11.5 cm.km−1) and is comprised of several laterally stable channels (Makaske et al. 2009). The channels are restricted by naturally occurring, silty, cohesive levees and by a railway embankment on the eastern side. The levees are controlled by the balance of aggradation and erosion, and although they are generally stable, avlusions can occur (Filgueira-Rivera et al. 2007). These avlusions create gaps in the levees that connect the river directly to floodplain wetlands, even when the river water levels are not high enough to overtop the levees. The floodplain wetlands in this study retain open water year round, although water level drawdown occurs.
Study Design
Because of the size and uniqueness of large rivers and their floodplains, manipulative experiments designed to tease apart the mechanisms by which connectivity to large rivers influences floodplain plant communities present immense logistical challenges (Sparks et al. 1990). We are left with mensurative experiments and correlations that preclude causal interpretations. Structural equation modeling (SEM) is a multivariate statistical tool that is well suited to teasing apart complex multivariate relationships that may provide insight into mechanisms by indicating support for causal interpretations of data collected in mensurative experiments (Grace 2008). Therefore, we designed this study as a mensurative experiment that compares the floating and submerged aquatic vegetation in 44 floodplain wetlands that span a gradient of connectivity to the Upper Columbia River. Ecological researchers have been using SEM for over two decades (e.g., Mitchell 1992; Johnson et al. 1993) and Grace (2006) provides a good introductory text. In brief, SEM combines elements of path and factor analysis, allowing researchers to test a hypothesized model of relationships between variables based on a covariance matrix. Further, SEM enables parameterization of those relationships with standardized regression coefficients for an assessment of the relative role of different ecological processes represented in the model (e.g., Arkema et al. 2009; Rooney and Bayley 2011a). Most relevant to our purposes, SEM allows us to model a variable believed to have mechanistic importance, but which cannot be directly measured, i.e. a latent variable. We suspect that connectivity to the river influences the submerged aquatic plant community, both directly and indirectly, yet it has no obvious measure. Instead, we measured three variables we believe indicate the level of connectivity (see below), and use these indicators to include the concept of connectivity as a latent variable in our model.
We first characterized the composition of floating and submerged aquatic vegetation in each wetland using hierarchical cluster analysis and indicator species analysis to identify community groupings. We then used non-metric multidimensional scaling ordination (NMS) to confirm cluster and indicator species analysis results and to summarize variance in community composition along three orthogonal axes. Next, we tested a SEM representing our hypothesis of how community composition is related to environmental variables known to be important to aquatic plants (Chambers and Kalff 1987; Koch 2001; Lacoul and Freedman 2006) and likely to be influenced by river connectivity: total suspended solids (TSS) represents the influence of light availability; total nitrogen (TN) indicates the role of nutrients; calcium carbonate (CaCO3) reflects alkalinity; sulfate (SO4 2−), as the dominant anion, represents the role of salinity; and sediment carbon content represents the balance of organic and mineral material in the sediment (Fig. 2).

Structural equation model indicating the pathways connecting the latent variable Connectivity to the measured variables. See the text for greater description of the model. Numbers associated with pathways are standardized regression coefficients, whereas numbers associated with variables are multiple r 2 values. Note that each measured variable has an associated error term, but these were left out of the figure for simplicity. The regression coefficients between connectivity and CaCO3 and connectivity and Sediment C are the same (−0.82). CaCO3 and Sediment C are correlated (r = 0.69), likely because of their similar response to connectivity
Connectivity, as a concept, cannot be measured directly. As mentioned, one advantage of SEM is that it allows us to quantify connectivity as a latent variable, indicated by three measurable variables: 1) amplitude of wetland water depth, 2) the sinuosity ratio for channels connecting the wetland to the river through a gap in the levees, and 3) the width of any gaps in the levees (Fig. 2). Connectivity is found in the agreement among these indicators. Gap width and gap depth are correlated (unpublished data), reflecting a gradient in planar area of gaps. We used gap width as a proxy for gap planar area because it can be measured from air photos.
As mentioned, another advantage of SEM is that it allows us to partition out the role of connectivity in shaping aquatic plant community composition that is due to its effects on particular environmental variables from any residual (or direct) effects that are attributable to factors like the physical disturbance of flooding (Bornette and Arens 2002; Riis and Hawes 2003) and the role of flood waters in dispersing plant propagules (Boedeltje et al. 2004; Combroux and Bornette 2004). For example, depending on velocity, floods may be depositional and cause burial or they may be erosional and cause uprooting and plant breakage (Sparks et al. 1990; Bornette et al. 2008).
Sampling
Water and sediment samples were collected from each wetland in July 2010, shortly after high water is traditionally observed. Samples were kept refrigerated prior to analysis. Water samples were collected using an integrated-depth sampling tube at the centre of each wetland. For TN, bulk water samples were digested by potassium persulfate analysed by flow injection analysis with a Lachat QuikChem 8500 FIA automated ion analyzer following standard methods for nitrate-nitrite (AWWA 1999). Alkalinity was determined by titration at pH 4.5 with a Man-Tech PC-Titrate with a conductivity probe, and SO4 2− was measured using a Dionex DX600 Ion Chromatograph and following U.S. EPA method 300.1 (Pfaff et al. 1997). To measure total suspended solids, a known volume was filtered on a pre-weighed 0.7 μm pore-sized Whatman GF/F filter paper, which was then oven dried at 105 °C to a constant weight before weighing total suspended solids.
Sediment samples comprised of three replicate 10 cm deep cores were collected at random locations within the wetland where water >20 cm deep, using a 5.7 cm diameter suction corer. Replicates were composited, homogenized, and dried before a 4–6 mg sub-sample was taken and analysed for total carbon by combustion in an Exeter Analytical CE40 Elemental Analyser.
Floating and submerged aquatic vegetation was sampled during peak biomass, following high water in July. Methods followed Rooney and Bayley (2011b). In brief, ten 1 m2 quadrats were deployed at random locations within the open water zone of each of the 44 floodplain wetlands. All floating and submerged vegetation within the column of water delineated by the quadrat was collected and sorted by species, with taxonomy following Klinkenberg (2012). Taxa that could not be resolved to the species-level included the marcroalgae Chara sp. and Nitella sp. The relative biovolume of each taxon was estimated using a modified Braun-Blanquet scale.
Connectivity to the river was estimated based on three indicators. Amplitude was calculated as the difference between maximum and minimum water levels based on observations of water depth taken weekly between May and August 2010. In more strongly connected wetlands, the amplitude tends to mirror that of the river proper, whereas in more isolated wetlands, the amplitude is reduced and the flood pulse is delayed. The levee gap width and the sinuosity ratio of channels connecting the wetlands to the river were measured from a 2004 black and white orthophotograph with 1 m pixels, provided by the British Colombia Ministry of Environment. This orthophoto was a mosaic of 1:20,000 air photos taken during late summer and early fall. Gap width and sinuosity were ground-truthed during plant sampling in 2010 and adequate agreement was found between the orthophoto and field measures (unpublished data). Several basins had multiple gaps connecting them to the river. Thus, gap width was the sum of all such gaps. The sinuosity ratio is calculated as the length of any connections between wetlands and the main river channel divided by the straight-line distance (i.e. the shortest distance between the river channel and the wetland across the levee). The width of channels connecting the wetlands to the river is representative of the size of the gap and will influence the degree to which river water impacts the wetland. Similarly, the sinuosity of any such drainage channels has been shown to directly affect the morphology of floodbasins and their biota (Bornette et al. 1998).
Community Characterisation
We adopted a riparian response guild approach to characterize the species present in the floodplain wetlands in terms of their adaptations to tolerate or avoid stress related to river flooding (sensu Merritt et al. 2010). This assisted us in our interpretation of the results of multivariate community analyses described below.
First, rare species (those present at <4 of the 44 wetlands) were removed and the relative biovolume of remaining species was arcsine-square root transformed, as recommended by McCune and Grace (2002). Next, Bray-Curtis distances (Bray and Curtis 1957) were calculated and the resulting dissimilarity matrix was used for multivariate analyses. All multivariate analyses were performed with PC-ORD (version 6.0, MjM Software, Glenden Beach, OR, US).
To identify consistent assemblages of floating and submerged species, we carried out hierarchical clustering analysis with the flexible beta linkage method, using a flexible beta value of −0.25, as recommended by McCune and Grace (2002). We used indicator species analysis (Dufrene and Legendre 1997) to help determine where to prune the resulting dendrogram and to identify which species were both faithful and exclusive to the resulting clusters or assemblages. Indicators were used to help characterize assemblages of submerged and floating vegetation following the riparian response guild approach. Their significance was evaluated with a Monte Carlo test involving 4,999 permutations.
The same dissimilarity matrix was then used to carry out non-metric scaling ordination (NMS). The optimal NMS solution was determined by an iterative process, comparing optimal solutions with one to four dimensions based on 50 runs with real data and 50 runs with randomized data. The optimal solution was then re-run to determine the final stress and instability values. Site scores from this solution were used to characterize the floating and submerged aquatic vegetation community composition in the SEM.
Correlations
We measured the Pearson correlation coefficients between NMS scores and the three indicators of connectivity: water amplitude, the width of any gaps in the levees separating wetlands from the river, and the sinuosity ratio of channels connecting wetlands to the river. We contrasted the strength of these correlations with the Pearson correlation coefficients between species important on NMS axes and the three measures of connectivity. Correlation coefficients and associated p-values were measured using SYSTAT software (version 13.0, SYSTAT Software, Inc., Chicago, IL, US).
Connectivity Model
The model (Fig. 2) was developed based on a review of pertinent literature and our first-hand experience in these wetlands (Carli 2011). To improve multivariate normality, some variables were log-transformed prior to analysis: gap width, TN, TSS, CaCO3, SO4 2−, and sediment carbon. The model parameters were estimated using AMOS software (version 19.0.0, SPSS Inc., IBM Corp., Somers, NY, US). Model-data agreement was evaluated based on a χ 2 test, which tests the null hypothesis that the model provides an adequate fit to the data, and the calculated root mean square error of approximation (RMSEA), which tests the null hypothesis that the model provides a close fit to the data in relation to the degrees of freedom in the model. Finally, the relative influence of connectivity via environmental intermediaries was compared to its direct influence by comparing standardized regression coefficients (regression weights transformed into units of standard deviations). Standardization allows us to compare the relative strength of different pathways in the model which are otherwise expressed in differing units of measure (Grace 2006).
Results
Community Characterisation
We produced a dendrogram indicating the hierarchical clustering of sites on the basis of dissimilarity in the relative biovolumes of their floating and submerged aquatic species, with 2.15 % chaining (Fig. S1). Pruning where about 25 % of the information about species distributions is remaining yields four distinct assemblages. We selected to prune at four rather than five clusters because this yielded a smaller average p-value among assemblage indicators following indicator species analysis (mean p-value was 0.18 vs. 0.21). Indicator species analysis revealed that each assemblage possesses diagnostic species with significant indicator value (Tables 1 and 2). Indicator values may range from 0 to 100 and denote the degree to which each species is faithfully and exclusively present in a given assemblage (Dufrene and Legendre 1997).
The results of clustering and indicator species analysis are strongly mirrored in our optimal NMS solution: both approaches identify four assemblages with consistently dominant or diagnostic species. The optimal NMS ordination possessed three dimensions, with reasonable final stress (14.17) and low final instability (<0.00001) after 69 iterations. This solution explained 83.6 % of the variance in community composition: 35.5 % on axis 1, 34.9 % on axis 2, and 13.2 % on axis 3. Joint plots that depict the wetland ordination scores and the species reasonably (r2 > 0.25) correlated with each NMS axis are presented in Fig. 3.

Joint plot of NMS ordination of plant community data. Location of points indicates the relative similarity in community composition of the 44 floodplain wetlands. Vectors indicate correlation coefficients between NMS axes and the relative abundance of indicated species. Note, only species with r 2 values > 0.25 on at least one axis are presented. Panel a) compares NMS axes 1 and 2, whereas panel b) compares NMS axes 1 and 3
Essentially, NMS 1 represents a gradient between two assemblages: assemblage one comprised of Stuckenia pectinata (L.) Börner, Zannichellia palustris L., Potamogeton richardsonii (A. Benn.) Rydb., and Potamogeton gramineus L.; and assemblage two comprised of Nuphar lutea (L.) Sm., Potamogeton natans L., Sparganium angustifolium Michx., Myriophyllum sibiricum Kom., and Utricularia macrorhiza Leconte. NMS 2 represents a gradient between assemblage one and assemblage three, dominated by Chara sp. NMS 3 represents variation in the relative biovolume of Potamogeton pusillus L. and Najas flexilis (Willd.) Rostk. & W.L.E. Schmidt, which constitute assemblage 4. In other words, as our cluster analysis revealed, we have four main plant assemblages: 1) S. pectinata and Z. palustris, usually in combination with P. richardsonii and P. gramineus; 2) N. lutea, P. natans, S. angustifolium, M. sibiricum, and U. macrorhiza; 3) Chara sp.; and 4) P. pusillus and N. flexilis. Characteristics of the important species were listed in Table 1.
Correlations with Connectivity
Not surprisingly, the three measures of connectivity are all positively correlated with one another (Pearson r > 0.4, p-values < 0.01). Amplitude was particularly strongly correlated with the relative biovolume of many of the species identified as characteristic of the different plant assemblages identified by clustering analysis and NMS ordination (Table 3). The species reasonably correlated with NMS 1 were correlated with measures of connectivity; however, the NMS1 scores were not (Table 3).
Connectivity Model
The model provided a close fit to the data (Table 4), and was able to explain 34 % of the variance in NMS1 scores, 49 % of the variance in NMS2 scores, and 62 % of the variance in NMS3 scores (Fig. 2). The five water and sediment chemistry variables were influenced by connectivity (Table 5). As anticipated in our second objective, connectivity to the river leads to increased TSS, but reduced TN, SO4, CaCO3, and sediment carbon. These sediment and water chemistry variables, in turn, influence the composition of floating and submerged aquatic vegetation. The standardized regression weights of sediment and water quality on NMS scores are depicted in Fig. 2 and in the electronic appendix in Supplementary Table 1. These results indicate that NMS 1 is primarily influenced negatively by SO4 2− and positively by TSS; NMS 2 is primarily negatively influenced by TSS and CaCO3; and NMS3 is positively influenced by TN and sediment carbon, but negatively influenced by CaCO3. Connectivity has both direct and indirect effects on floating and submerged aquatic vegetation community composition (Table 6). Net effects are the sum of these direct and indirect effects, and relate to the correlations observed between indicators of connectivity and the NMS scores (Table 6).
Discussion
We found four distinct assemblages of floating and submerged aquatic vegetation growing in the Upper Columbia floodplain wetlands, each comprised of co-occurring species with similar or complementary adaptations to the direct (residual) and indirect effects of flooding (Table 1) (i.e., flood response guilds). Furthermore, we found that connectivity to the river reduces water clarity through inputs of turbid water, as well as reducing salinity, alkalinity, nitrogen, and sediment carbon levels by scouring (Table 6), and that these variables are related to the distribution of our four assemblages. Traditionally, the confounded nature of direct and indirect effects of connectivity on plant community composition, combined with the logistical difficulties in carrying out whole basin experiments, obscures the mechanisms behind the influence of the annual flood pulse on vegetation in floodplain wetlands. We have succeeded in teasing apart the effect of these specific environmental variables from the residual effects of connectivity on plant community composition. By using a flood response guild approach, we have connected the life history traits of individual species to the composition of plant assemblages and their spatial distribution in relation to connectivity, sediment, and water quality.
Plants have evolved a variety of life history, behavioral, and morphological adaptations to flooding (Lytle and Poff 2004). Yet where floods are less frequent or intense, forces like nutrient limitation, propagule fluxes, competition, and herbivory become the dominant driving forces behind community composition and the costs of adaptations to frequent or intense flooding begin to outweigh the benefits. Bornette et al. (2008) proposed a model whereby habitats exposed to high frequency or intensity flooding (i.e. connected wetlands) would select opportunistic species or those tolerant of substrate instability, low light levels, and scouring while less connected habitats would favor competitive dominants. Merritt et al. (2010) built on this model by describing different riparian response guilds that vary in terms of morphology, life history, reproductive strategy, and other adaptations to flood disturbance. Using this concept, we can characterize the four assemblages identified in our ordination and understand their distributions in relation to wetland connectivity (Table 1).
The strong agreement between our different statistical approaches to characterizing the floating and submerged assemblages gives us confidence when using the ordination scores in our structural equation model, and so we will restrict our discussion to the NMS results. With the exception of NMS 1 scores (where indirect effects are equally important), the influence of connectivity on plant community composition is predominantly direct (residual) (Table 6). We interpret the direct effect of connectivity primarily as the physical disturbance and scouring related to flooding. This is in agreement with the Bornette et al. (2008) model, suggesting that adaptations that minimize the damage plants receive during floods and the energy required to persist in floodplain wetlands are favoured in highly connected wetlands. This is also in agreement with the experimental findings of Drinkard et al. (2011), who found that marsh plants were most stressed in the intermittently flooded zone, where flood disturbance is most intense.
We note that additional mechanisms may also contribute to the observed direct effect of connectivity on the SAV community composition, such as effects on dispersal and seed bank composition (Abernethy and Willby 1999; Leyer 2006). Furthermore, any variable influencing the floating or submerged community composition that also responds to connectivity but that is not included in our model will have its influence lumped together with the residual effects of connectivity, so there is a risk of ascribing unmeasured indirect effects to the direct effect of connectivity. However, we find evidence that physical disturbance is an important contributor to plant community composition in the plant traits common to different flood response guilds.
There are three main strategies that plants employ to persist in floodplain wetlands. They may avoid flood disturbance by reducing hydraulic resistance, tolerate damage to their shoots by rapidly regenerating from extensive rhizomes, or succumb to physical disturbance, but re-colonize frequently (Table 1). Species that are streamlined and have thin, flexible shoots will present little hydraulic resistance to flood waters, whereas bushier species present greater hydraulic resistance and thus require greater anchoring strength to avoid uprooting (Haslam 1978). In our first assemblage, Z. palustris and S. pectinata are streamlined submerged plants that present little hydraulic resistance. P. richardsonii and P. gramineus, also members of assemblage 1, have broader submerged leaves that present slightly greater resistance to flow, but when shoots are damaged they are able to regenerate from rhizomes (Combroux and Bornette 2004). In heterophyllous P. gramineus, the more streamlined submerged leaves may compensate for damage to broader floating leaves incurred during the annual flood pulse, while floating leaves overcome light limitation when floodwaters recede. This may explain why the direct effect of Connectivity favours assemblage 1 (Fig. 2). P. pusillus, and N. flexilis (assemblage four) and S. angustifolium (assemblage 2) are also streamlined in morphology, but they are more prone to tangling and are easily uprooted unless they are able to root deeply in organic or silty sediments (Haslam 1978), which seldom accumulate in well-connected wetlands. This may explain why the direct effect of connectivity on assemblages two and four is negative. Chara sp. (assemblage three) escapes some hydraulic resistance because of its low profile, but it has a bushy, brittle growth form and has only holdfasts rather than proper roots to anchor it to the sediment (Bornette and Arens 2002). Thus, it is clear why the direct effect of connectivity on assemblage three is negative (Fig. 2), despite Chara sp. being a pioneering taxon (Wade 1990). In assemblage two, the broad floating leaves of N. lutea, and P. natans and the bushy shoots of U. macrorhiza and M. sibiricum all present greater hydraulic resistance (Haslam 1978). Floating-leaved species are particularly susceptible to damage from surface currents because of the risk of tearing and immersion (Sculthorpe 1967), even the narrow floating leaves of S. angustifolium. The extensive rhizomes of N. lutea, P. natans, and M. sibiricum may permit regeneration following the flood pulse, but U. macrorhiza is not even rooted. Instead, it recolonizes from unspecialized vegetative fragments (Combroux and Bornette 2004) or floating turions. The energy used to regenerate from rhizomes or unspecialized fragments may explain why the direct influence of connectivity on assemblage two is also negative (Fig. 2).
A response-guild approach can also be employed to interpret the indirect effects of flooding on the four plant assemblages (Fig. 2), based on the kinds of adaptations described in Table 1. For example, although the range in salinity among the floodplain wetlands was slight (162 to 1,380 μS.cm−1), species belonging to assemblage one prefer the alkaline (high CaCO3) and saline (high SO4 2−) end of this range, whereas species belonging to assemblage two prefer fresh water (Stewart and Kantrud 1972). The four species of Chara (assemblage three) found in British Columbia collectively span a broad range of salinity preferences (Klinkenberg 2012) but all thrive in alkaline waters (Wade 1990), explaining why assemblage three responded differently to CaCO3 and SO4 2− (Fig. 2).
Relationships to TSS are most easily interpreted based on light limitation. The floating leaves of N. lutea, S.angustifolium, and P. natans (assemblage two) should be robust against turbidity compared to submerged P. richardsonii, S. pectinata, and Z. palustris (assemblage one). Although it has a submerged growth form, Chara sp. (assemblage three) is also known to tolerate low light (Wade 1990; Kufel and Kufel 2002; de Winton et al. 2004).
In terms of nutrients (TN and Sediment C), Chara sp. adheres to the sediment with holdfasts that attach best to fine grained mineral substrates, whereas S. pectinata, Z. palustris (assemblage one) both prefer organic substrates and nutrient rich environments (Klinkenberg 2012). Similarly, P. pusillus and N. flexilis (assemblage four) are both nutrient loving and anchor better in organic substrates. However, the relationship between nutrients and NMS 1 scores is less easily explained. TN appears unrelated to assemblages one and two, perhaps because several of these species take up nutrients from the sediment (Barko et al. 1991a). There is also evidence that nutrient requirements are lessened when light is limited (Barko et al. 1991b). In which case, requirements would be lower in highly connected wetlands where nutrients are scarcer, potentially obfuscating any relationship between TN and NMS 1 scores. U. macrorhiza (assemblage two) is not rooted and as a carnivorous species it is advantaged under low nutrient levels (Adamec 2010); however, other members of assemblage two (N. lutea, P. natans, and M. sibiricum) are known to prefer organic substrates (Haslam 1978; Klinkenberg 2012). In contrast, Sediment C is positively associated with S. pectinata, Z. palustris, P. gramineus, and P. richardsonii (assemblage one), which span a range in nutrient preferences from very rich to poor (Klinkenberg 2012). NMS 1 was the axis our model was least able to explain (r 2 = 0.34), and it seems that nutrient levels in water and sediment were not the most influential factors in the distribution of vegetation assemblages one and two. Perhaps biotic interactions such as facilitation or niche partitioning play a role in the composition of these assemblages that supersedes the individual nutrient level preferences of constituent species.
Amplitude is the strongest indicator of connectivity in our model and the measure most strongly correlated with NMS scores and species relative biovolumes. Likely, this is because it is the most direct measure of connectivity, indicating the magnitude of the annual flood pulse in each wetland. In contrast, gap width and channel sinuosity are indicators that apply to the subset of wetlands directly connected to the river. They do not apply to wetlands that are only connected to the river when the levees are overtopped.
If amplitude is strongly correlated with the relative biovolumes of all the species that covary with NMS 1 scores (except for N. lutea), why are NMS 1 scores not correlated with amplitude (Table 3)? This is particularly curious as our anecdotal field observations suggest that assemblage one (S. pectinata and Z. palustris) was associated with the wetlands we suspected of being more connected to the river based on the presence of gaps in the natural levees. This apparent contradiction is explained by comparing the direct and indirect standardized regression coefficients between Connectivity and NMS 1 scores (Table 6). Floating-leaved species in assemblage two will be far less subject to light limitation by elevated TSS and U. macrorhiza may be better able to compete in nutrient limited environments. In contrast, S. pectinata and Z. palustris (assemblage one) are more tolerant of salts and alkaline conditions. Because of the influence of connectivity on these water and sediment quality variables, the indirect effect of connectivity favors assemblage two (N. lutea, P. natans, U. macrorhiza, S. angustifolium, and M. sibiricum). However, the direct effect of connectivity, which we interpret as the physical disturbance associated with scouring by flood waters, favors assemblage one (S. pectinata, Z. palustris, P. richardsonii, and P. gramineus), which better tolerates flood-related scouring through reduced hydraulic resistance and the capacity to quickly regenerate from rhizomes. Consequently, the net effect of connectivity on NMS 1 scores is negligible. Without the use of SEM to tease apart these direct and indirect effects, the apparent absence of any relationship between NMS1 scores and connectivity could not be explained.
In conclusion, we observed four assemblages of floating and submerged aquatic vegetation in 44 floodplain wetlands of varying connectivity to the river. Connectivity, as a concept, was primarily indicated by amplitude, but the width and sinuosity of channels connecting the wetlands to the river via gaps in the levees were also indicators of connectivity. As anticipated, plant community composition was related to the degree of connectivity to the river directly, most likely through the effect of physical disturbance, erosion, and scour from the annual flood pulse. Plant community composition was also affected by indicators of turbidity, salinity, alkalinity, nutrient levels, and sediment organic content, and these, in turn, were influenced by connectivity. However, these indirect effects were generally less influential than the direct effect, and based on our analysis of plant traits this seems to be largely attributable to physical disturbance associated with the annual flood pulse. Our study therefore suggests that the direct effect of the flood pulse can override the influence of sediment and water quality in structuring plant assemblages in floodplain wetlands, but that in wetlands where the intensity of flooding is reduced, these water and sediment conditions resume their structuring role. Finally, the correlations between indicators of connectivity and ordination scores defining the four assemblages were inconsistent with field observations, but by breaking the net effects of connectivity into its direct (residual) and indirect (mediated by water and sediment quality) components with SEM, we were able to explain these inconsistencies and better understand the community dynamics in the Upper Columbia River floodplain wetlands.
References
Abernethy VJ, Willby NJ (1999) Changes along a disturbance gradient in the density and composition of propagule banks in floodplain aquatic habitats. Plant Ecology 140:177–190
Adamec L (2010) Mineral cost of carnivory in aquatic carnivorous plants. Flora 205:618–621
Amoros C, Bornette G (1999) Antagonistic and cumulative effects of connectivity: a predictive model based on aquatic vegetation in riverine wetlands. Archiv Fur Hydrobiologie 115:311–327
Arkema KK, Reed DC, Schroeter SC (2009) Direct and indirect effects of giant kelp determine benthic community structure and dynamics. Ecology 90:3126–3137
AWWA (1999) 4500-NO3 I. In: Rice EW, Baird RB, Eaton AD, Clesceri LS (eds) Standard methods for the examination of water and wastewater, American Public Health Association, American Water Works Association, and the Water Environment Federation, Denver, CO
Barko JW, Gunnison D, Carpenter SR (1991a) Sediment interactions with submersed macrophyte growth and community dynamics. Aquatic Botany 41:41–65
Barko JW, Smart RM, McFarland DG (1991b) Interactive effects of environmental conditions on the growth of submersed aquatic macrophytes. Journal of Freshwater Ecology 6:199–207
Barrett R, Nielsen DL, Croome R (2010) Associations between the plant communities of floodplain wetlands, water regime and wetland type. River Research and Applications 26:866–876
Bayley PB (1995) Understanding large river floodplain ecosystems. Bioscience 45:153–158
Bayley SE, Guimond JK (2009) Aboveground biomass and nutrient limitation in relation to river connectivity in montane floodplain marshes. Wetlands 29:1243–1254
Bayley SE, Guimond JK (2008) Effects of river connectivity on marsh vegetation community structure and species richness in montane floodplain wetlands in Jasper National Park, Alberta, Canada. Ecoscience 15:377–388
Blanch SJ, Ganf GG, Walker KF (1999) Tolerance of riverine plants to flooding and exposure indicated by water regime. Regulated Rivers-Research & Management 15:43–62
Boedeltje G, Bakker JP, Ten Brinke A, Van Groenendael JM, Soesbergen M (2004) Dispersal phenology of hydrochorous plants in relation to discharge, seed release time and buoyancy of seeds: the flood pulse concept supported. Journal of Ecology 92:786–796
Bornette G, Amoros C, Lamouroux NL (1998) Aquatic plant diversity in riverine wetlands: the role of connectivity. Freshwater Biology 39:267–283
Bornette G, Arens MF (2002) Charophyte communities in cut-off river channels—The role of connectivity. Aquatic Botany 73:149–162
Bornette G, Tabacchi E, Hupp C, Puijalon S, Rostan JC (2008) A model of plant strategies in fluvial hydrosystems. Freshwater Biology 53:1692–1705
Bray JR, Curtis JT (1957) An ordination of the upland forest communities of southern Wisconsin. Ecological Monographs 27:326–349
Carli CM (2011) Floodplain ecology of the Upper Columbia River. Dissertation, University of Alberta
Casanova MT, Brock MA (2000) How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecology 147:237–250
Chambers PA, Kalff J (1987) Light and nutrients in the control of aquatic plant community structure: in situ experiments. Journal of Ecology 75:611–619
Combroux ICS, Bornette G (2004) Propagule banks and regenerative strategies of aquatic plants. Journal of Vegetation Science 15:13–20
de Winton MD, Casanova MT, Clayton JS (2004) Charophyte germination and establishment under low irradiance. Aquatic Botany 79:175–187
Drinkard MK, Kershner MW, Romito A, Nieset J, de Szalay FA (2011) Responses of plants and invertebrate assemblages to water-level fluctuation in headwater wetlands. Journal of the North American Benthological Society 30:981–996
Dufrene M, Legendre P (1997) Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs 67:345–366
Filgueira-Rivera M, Smith ND, Slingerland RL (2007) Controls on natural levee development in the Columbia river, British columbia, Canada. Sedimentology 54:905–919
Grace JB (2006) Structural equation modeling and natural systems. Cambridge University Press, Cambridge
Grace JB (2008) Structural equation modeling for observational studies. Journal of Wildlife Management 72:14–22
Greet J, Webb JA, Cousens RD (2011) The importance of seasonal flow timing for riparian vegetation dynamics: a systematic review using causal criteria analysis. Freshwater Biology 56:1231–1247
Grime JP (1973) Competitive exclusion in herbaceous vegetation. Nature 242:344–347
Haslam SM (1978) River plants. Cambridge University Press, London
Heiler G, Hein T, Schiemer F, Bornette G (1995) Hydrological connectivity and flood pulses as the central aspects for the integrity of a river-floodplain system. Regulated Rivers-Research & Management 11:351–361
Hermoso V, Ward DP, Kennard MJ (2012) Using water residency time to enhance spatio-temporal connectivity for conservation planning in seasonally dynamic freshwater ecosystems. Journal of Applied Ecology 49:1028–1035
Hodgson J, Moilanen A, Thomas CD (2009) Metapopoulation responsees to patch connectivity and quality are masked by successional habitat dynamics. Ecology 90:1608–1619
Johnson ML, Huggins DG, Denoyelles F Jr. (1993) Structural equation modeling and ecosystem analysis. In: Graney RL (ed) SETAC Special Publications Series; Aquatic mesocosm studies in ecological risk assessment. CRC Press, London, UK. pp 627–652
Junk WJ, Bayley PB, Sparks RE (1989) The flood pulse concept in river-floodplain systems. In: Dodge DP (ed) Canadian special publication of fisheries and aquatic sciences: Proceedings of the International Large River Symposium (LARS). Department of Fisheries and Oceans, Ontario, pp 110–127
Klinkenberg B (2012) E-Flora BC: Electronic atlas of the flora of British Columbia. Lab for advanced spatial analysis. Department of Geography, University of British Columbia, Vancouver, B.C. www.eflora.bc.ca. Accessed 19 July 2012
Koch EM (2001) Beyond light: physical, geological, and geochemical parameters as possible submersed aquatic vegetation habitat requirements. Estuaries 24:1–17
Kufel L, Kufel I (2002) Chara beds acting as nutrient sinks in shallow lakes—a review. Aquatic Botany 72:249–260
Lacoul P, Freedman B (2006) Environmental influences on aquatic plants in freshwater ecosystems. Environmental Reviews 14:89–136
Lenssen J, Menting F, van der Putten WH, Blom K (1999) Control of plant species richness and zonation of functional groups along a freshwater flooding gradient. Oikos 86:523–534
Leyer I (2006) Dispersal, diversity and distribution patterns in pioneer vegetation: the role of river-floodplain connectivity. Journal of Vegetation Science 17:407–416
Lytle DA, Poff NL (2004) Adaptation to natural flow regimes. Trends in Ecology & Evolution 19:94–100
Makaske B, Smith DG, Berendsen HJA, Boer AG, van Nielen-Kiezebrink MF, Locking T (2009) Hydraulic and sedimentary processes causing anastomosing morphology of the upper Columbia River, British Columbia, Canada. Geomorphology 111:194–205
McCreary NJ (1991) Competition as a mechanism of submersed macrophyte community structure. Aquatic Botany 41:177–193
McCune B, Grace JB (2002) Analysis of ecological communities. MjM Software Design, Gleneden Beach
Merritt DM, Scott ML, Poff NL, Auble GT, Lytle DA (2010) Theory, methods and tools for determining environmental flows for riparian vegetation: riparian vegetation-flow response guilds. Freshwater Biology 55:206–225
Mitchell RJ (1992) Testing evolutionary and ecological hypotheses using path-analysis and structural equation modeling. Functional Ecology 6:123–129
Nielsen DL, Chick AJ (1997) Flood-mediated changes in aquatic macrophyte community structure. Marine and Freshwater Research 48:153–157
Pederson GT, Gray ST, Woodhouse CA, Betancourt JL, Fagre DB, Littell JS, Watson E, Luckman BH, Graumlich LJ (2011) The unusual nature of recent snowpack declines in the North American Cordillera. Science 333:332–335
Pfaff JD, Hautman DP, Munch DJ (1997) In: U.S.EPA (ed) EPA method 300.1: Determination of inorganic anions in drinking water by ion chromatography. National Exposure Research Laboratory Office of Research and Development, U.S. Environmental Protection Agency, Cincinnati, p 39
Riis R, Biggs BJF (2001) Distribution of macrophytes in New Zealand streams and lakes in relation to disturbance frequency and resource supply—a synthesis and conceptual model. New Zealand Journal of Marine and Freshwater Research 35:255–267
Riis T, Hawes I (2003) Effect of wave exposure on vegetation abundance, richness and depth distribution of shallow water plants in a New Zealand lake. Freshwater Biology 48:75–87
Robertson AI, Bacon P, Heagney G (2001) The responses of floodplain primary production to flood frequency and timing. Journal of Applied Ecology 38:126–136
Rooney RC, Bayley SE (2011a) Relative influence of local- and landscape-level habitat quality on aquatic plant diversity in shallow open-water wetlands in Alberta’s boreal zone: direct and indirect effects. Landscape Ecology 26:1023–1034
Rooney RC, Bayley SE (2011b) Setting reclamation targets and evaluating progress: submersed aquatic vegetation in natural and post-oil sands mining wetlands in Alberta, Canada. Ecological Engineering 37:569–579
Sculthorpe CD (1967) The biology of aquatic vascular plants. Edward Arnold Ltd, London
Sparks RE, Bayley PB, Kohler SL, Osborne LL (1990) Disturbance and recovery of large floodplain rivers. Environmental Management 14:699–709
Stewart RE, Kantrud HA (1972) Vegetation of prairie potholes, North Dakota, in relation to quality of water and other environmental factors. Geologic Survey Professional Paper 585-D, U.S. Bureau of Sport Fisheries and Wildlife. Washington, D.C. p 35
Wade PM (1990) The colonization of disturbed fresh-water habitats by Caraceae. Folia Geobotanica & Phytotaxonomica 25:275–278
Ward JV (1998) Riverine landscapes: biodiversity patterns, disturbance regimes, and aquatic conservation. Biological Conservation 83:269–278
Ward JV, Tockner K (2001) Biodiversity: towards a unifying theme for river ecology. Freshwater Biology 46:807–819
Acknowledgments
We thank area landowners for advice, and Christina Buelow for assistance in the field. Funding for this project came in part from scholarships to R. C. Rooney from the Killam Trusts and to C. Carli from the Queen Elizabeth II Graduate Scholarship, administered by the University of Alberta. Additional support came from the Columbia Wetlands Stewardship Partners.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOC 112 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Rooney, R.C., Carli, C. & Bayley, S.E. River Connectivity Affects Submerged and Floating Aquatic Vegetation in Floodplain Wetlands. Wetlands 33, 1165–1177 (2013). https://doi.org/10.1007/s13157-013-0471-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-013-0471-4