
Abstract
The libellulid dragonfly genus Sympetrum has been recognized since 1833, but lacks any morphological synapomorphies to unite the taxon. Previous researchers have disagreed over which species belong in Sympetrum, bringing the monophyly of the genus into question. We use DNA sequence data from 6 genetic loci (16S, tRNA-valine, 12S, elongation factor 1 alpha, cytochrome oxidase subunit I, and the second internal transcribed spacer region) and 25 morphological characters (mainly genitalic) to test the monophyly of Sympetrum with Bayesian inference and maximum likelihood analyses. Under Bayesian inference, all Sympetrum species included in this study form a clade, which also contains the Hawaiian monotypic genus Nesogonia, often considered a close relative of Sympetrum. Phylogenetic analyses also reveal at least six strongly supported clades (treated as species groups) within Sympetrum, but relationships between these species groups remain unresolved or unsupported. Although the relationships between Sympetrum species groups remain unresolved, several species groups include taxa from multiple biogeographic regions/continents, and the species group sister to the rest of Sympetrum contains migratory species from the New World and Africa. This pattern suggests a complex biogeographic history in Sympetrum shaped by vicariance and dispersal. Preliminary estimates of the divergence dates of Sympetrum species groups outline a rapid radiation of the groups approximately 32-38 million years ago, possibly influenced by cooling and drying climates of the late Eocene and early Oligocene.



Similar content being viewed by others



References
Allioni, C. (1766). Manipulus Insectorum Taurinensium. Melanges de la Societe de Turin, 3(7), 185–198.
Bartenev, A. K. (1913). Contributions à la connaissance des Odonates de l’Asie paléarctique du Musée Zoologique de l’Académie Impériale des Sciences de St. Pétersbourg, 2. Annuaire du Musee Zoologique de l’Academie des Sciences de St. Petersbourg, 17(3/4), 289–310.
Bartenev, A. K. (1914). Matériaux pour l’étude de la faune des Libellules de la Sibérie. 16. Odonata de la province d’Oussouri. Horae Societatis Entomological Rossica, 41(2), 1–21.
Borror, D. J. (1945). A key to the New World genera of Libellulidae (Odonata). Annals of the Entomological Society of America, 38, 168–194.
Calvert, P. P. (1890). In Hagen, H.A: A synopsis of the odonata genus Leucorrhinia Britt. Transactions of the American Entomological Society, 17, 229–236.
Cannings, R. A. (1981). The larva of Sympetrum madidum (Hagen) (Odonata: Libellulidae). Pan-Pacific Entomologist, 57, 341–346.
Cannings, R. A., & Garrison, R. W. (1991). Sympetrum signiferum, a new species of dragonfly (Odonata: Libellulidae) from western Mexico and Arizona. Annals of the Entomological Society of America, 84, 474–479.
Carle, F. L. (1993). Sympetrum janeae spec. nov. from eastern North America, with a key to Nearctic Sympetrum (Anisoptera: Libellulidae). Odonatologica, 22, 1–16.
Charpentier, T. (1840). Libellinae Europaeae descriptae ac depictae. Voss: Lipsiae.
DeMarmels, J. (1994). Sympetrum chaconi spec. nov. from Auyan-Tepui, Venezuela, with notes on a pantepuyan form of Tramea binotata (Rambur) (Anisoptera: Libellulidae). Odonatologica, 23, 405–412.
DeMarmels, J. (2001). Sympetrum paramo sp. n. (Odonata: Libellulidae) from the Venezuelan high Andes, with a key to the species of Sympetrum Newman, 1833 found in Venezuela. Entomotropica, 16, 15–19.
Denk, T., Grimsson, F., & Zetter, R. (2010). Episodic migration of oaks to Iceland: Evidence for a North Atlantic "land bridge" in the latest Miocene. American Journal of Botany, 97, 276–287.
Drummond, A. J., & Rambaut, A. (2007). BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evolutionary Biology, 7, 214.
Drury, D. (1773). Illustrations of Natural History. London, England: Simpkin.
Dunkle, S. D. (2000). Dragonflies through Binoculars. New York: Oxford University Press.
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research, 32(5), 1792–1797.
Garrison, R. W., von Ellenrieder, N., & Louton, J. A. (2006). Dragonfly genera of the New World. Baltimore, Maryland: Johns Hopkins University Press.
Gentilini, G. (1988). The Upper Miocene dragonflies of Monte Castellaro (Marche, Central Italy) (Odonata: Libellulidae). Memoire della Societa Entomologica Italiana, 67, 251–272.
Gloyd, L. K., & Wright, M. (1959). Odonata. In W. T. Edmondson (Ed.), Freshwater biology (pp. 917–940). New York: J. Wiley.
Hagen, H. A. (1861). Synopsis of the Neuroptera of North America. Washington, D.C.: Smithsonian Institute.
Hagen, H. A. (1867). Revision der von Herrn Uhler beschriebenen Odonaten. Stettiner Entomologische Zeitung, 28, 87–95.
Hagen, H. A. (1874). Report on the Pseudo-Neuroptera and Neuroptera collected by Lieut. W. L. Carpenter in 1873 in Colorado. Report for the U.S. Geological Survey, 7, 571–606.
Hagen, H. A. (1890). A synopsis of the odonata genus Leucorrhinia Britt. Transactions of the American Entomological Society, 17, 229–236.
Han, F.-Y., & Zhu, H.-Q. (1997). Sympetrum xiaoi spec. nov., a new dragonfly from Shanxi, China (Anisoptera: Libellulidae). Odonatologica, 26, 343–345.
Kiauta, B. (1969). The chromosomes of the Hawaiian endemic dragonflies Megalagrion oahuense (Coenagrionidae: Pseudoagrioninae) and Nesogonia blackburni (Libellulidae: Sympetrinae) with a note on the cytotaxonomic affinities between the genera Nesogonia and Sympetrum (Odonata). Proceedings of the Hawaiian Entomology Society, 20, 429–433.
Kormondy, E. J. (1958). Catalogue of the Odonata of Michigan. Miscellaneous Publications of the Museum of Zoology of the University of Michigan, 104, 43.
Kormondy, E. J. (1960). New North American records of anisopterous Odonata. Entomology News, 71, 121–130.
Linnaeus, C. (1758). Systema naturae (10th ed.). Laurentii Salvii: Holmiae.
Linnaeus, C. (1763). Centuria Insectorum. Proposuit Boas Johansson. Amoenitates Acadamicae, 6, 384–415.
Lomolino, M. V., Riddle, B. R., & Brown, J. H. (2005). Biogeography (3rd ed.). Sinauer Associates: Sunderland, Massachusetts.
McLachlan, R. (1883). Neuroptera of the Hawaiian Islands. Part I. Pseudo-Neuroptera. Annals and Magazine of Natural History, 5(12), 226–240.
Miller, K. G., Browning, J. V., Aubry, M.-P., Wade, B. S., Katz, M. E., Kulpecz, A. A., et al. (2008). Eocene-Oligocene global climate and sea-level changes: St, Stephens Quarry, Alabama. Geological Society of America Bulletin, 120(1/2), 34–53.
Misof, B., Anderson, C. T., & Hadrys, H. (2000). A phylogeny of the damselfly genus Calopteryx (Odonata) using mitochondrial 16S rDNA markers. Molecular Phylogenetics and Evolution, 15(1), 5–14.
Montgomery, B. E. (1943). Sympetrum internum, new name for Sympetrum decisum Auct., nec Hagen (Odonata, Libellulidae). Canadian Entomologist, 75(3), 57–58.
Müller, R. A. (1764). Fauna Insectorum Fridrichsdalina. Gleditsch: Hafniae et Lipsiae.
Needham, J. G., & Fisher, E. (1936). The nymphs of North American Libelluline dragonflies (Odonata). Transactions of the American Entomological Society, 62, 107–116.
Needham, J. G., Westfall, M. J., Jr., & May, M. L. (2000). Dragonflies of North America. Gainesville, Florida: Scientific Publishers.
Newman, E. (1833). Entomological Notes. Entomological Magazine, 1, 505–514.
Nylander, J. A. A. (2004). MrModeltest v2. Program distributed by the author. Uppsala: Evolutionary Biology Centre, Uppsala University.
Oguma, K. (1915). A list of dragonflies collected by Mr. Oguma from Kiushiu and Loo-Choo. Entomology Magazine Kyoto, 1, 141–148.
Oguma, K. (1922). The Japanese Dragonfly-Fauna of the Family Libellulidae. Deutsche Entomologische Zeitschrift, 1922, 96–112.
Paulson, D. (2009). Dragonflies and Damselflies of the West. Princeton, New Jersey: Princeton University Press.
Pilgrim, E. M., & von Dohlen, C. D. (2007). Molecular and morphological study of species-level questions within the dragonfly genus Sympetrum (Odonata: Libellulidae). Annals of the Entomological Society of America, 100(5), 688–702.
Pilgrim, E. M., & von Dohlen, C. D. (2008). Phylogeny of the Sympetrinae (Odonata: Libellulidae): further evidence of the homoplasious nature of wing venation. Systematic Entomology, 33(1), 159–174.
Rambaut, A., & Drummond, A. J. (2007). Tracer v1.4. Published by authors: http://beast.bio.ed.ac.uk/Tracer.
Rambur, J. P. (1842). Histoire naturelle des insectes. Névroptères. Paris, France: Roret.
Ris, F. (1911). Ueber einige Gomphinen von Südbraisilien und Argentina. Mémoires: Société entomologique de Belgique, 19, 101–119.
Ronquist, F., & Huelsenbeck, J. P. (2003). MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics, 19, 1572–1574.
Sanmartin, I., Enghoff, H., & Ronquist, F. (2001). Patterns of animal dispersal, vicariance and diversification in the Holarctic. Biological Journal of the Linnean Society, 73, 345–390.
Say, T. (1839). Descriptions of new North American Neuropterous insects, and observations on some already described. Journal of the Academy of Natural Science of Philadelphia, 8(1), 9–46.
Schmidt, E. (1987). Generic reclassification of some West Palearctic Odonata taxa in view of their Nearctic affinities (Anisoptera: Gomphidae, Libellulidae). Advances in Odonatology, 3, 135–145.
Selys, E. (1840). Monographie des Libellulidées d’Europe. Paris, France: Roret.
Selys, E. (1841). Nouvelles Libellulidées d’Europe. Revue Zoologique, 4, 243–246.
Selys, E. (1850). Revue des odonates; ou libellules d’Europe. Paris, France: Chez Roret.
Selys, E. (1883). Les Odonates du Japon. Annales de la Société entomologique belge, 27, 82–143.
Selys, E. (1884). Révision des Diplax paléarctiques. Annales de la Société entomologique belge, 28, 29–45.
Shapiro, B., Rambaut, A., & Drummond, A. J. (2006). Choosing appropriate substitution models for the phylogenetic analysis of protein-coding sequences. Molecular Biology and Evolution, 23, 7–9.
Stamatakis, A. (2006). RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics, 22, 2688–2690.
Sulzer, J. H. (1776). Abgekürtze Geschichte der Insekten nach dem Linnaeischen System. 1. Germany: Winterthur, H. Steiner & County.
Walker, E. M., & Corbet, P. S. (1975). The Odonata of Canada and Alaska vol. 3. Toronto: Toronto University Press.
Ware, J. L., Simon, Y. W., Ho, & Kjer, K. (2008). Divergence dates of libelluloid dragonflies (Odonata: Anisoptera) estimated from rRNA using paired-site substitution models. Molecular Phylogenetics and Evolution, 47(1), 426–432.
Whitfield, J. B., & Kjer, K. M. (2008). Ancient rapid radiations of insects: Challenges for phylogenetic analysis. Annual Review of Entomology, 53, 449–472.
Acknowledgements
We wish to thank all the individuals that provided specimens for this study: K.D. Dijkstra, Kiyoshi Inoue, Margi Chriscinske, Jerrell Diagle, Jim Johnson, Mike May, Blair Nikula, Heath Ogden, Bill Radke, Graham Reels, Todd Sformo, and Steve Valley. We would also like to thank James Pitts, Paul Wolf, Terry Griswold, and Carol Dehler for reviews of previous versions of this research, and Carrie Drake and Usha Spaulding for laboratory assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
This is a contribution to the Festschrift for Michael L. May
Appendix
Appendix
Descriptions of the morphological characters used in the phylogenetic analyses.
-
1.
Cornua of penis: (0) absent; (1) two cornua present; (2) four cornua present.
Cornua are structures that are believed to aid in the removal of sperm from previous matings. They do not occur on all male odonate genitalia, but are found in pairs when they do occur. All Sympetrum species had at least one pair of cornua. As the cornua were not found in the outgroup species, these taxa were coded as missing for characters 2, 3, 4, 9, 10, and 17.
-
2.
Cornua curve: (0) anterior pointing; (1) posterior pointing.
Only members of the flaveolum group had posterior-directed cornua (Fig. 4a). All other clades of Sympetrum exhibited anterior pointing cornua (Fig. 4b, c).
Fig. 4 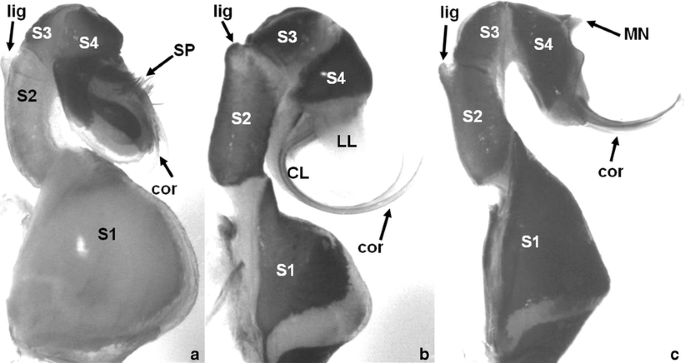
Lateral views of representative Sympetrum male genitalia to identify coded morphological characters: a S. rubicundulum; b S. danae; c S. gracile. Labeled structures: CL central lobe; cor cornua; LL lateral lobe; lig ligula notch; MN median nodule; S1-S4 penis segments 1 to 4; SP setal patch
-
3.
Cornua thickness: (0) flattened; (1) thin/cylindrical.
The S. flaveolum and S. infuscatum clades had flattened cornua (Fig. 4a), along with S. illotum and N. blackburni, whose affiliations to species-clades were unclear. The basal clade (fonscolombii group) had cylindrical, but shortened cornua. The other species clades (danae, pedemontanum, and vulgatum groups) all exhibited long cylindrical cornua (Fig. 4b, c). Sympetrum signiferum, a member of the danae group, had flattened cornua.
-
4.
Cornua tips: (0) tapering smoothly to point; (1) hooked at point; (2) rounded tips.
The distal ends of the cornua tapered smoothly to a point for all the members of the danae, flaveolum, and vulgatum species groups. Members of the fonscolombii and pedemontanum clades along with N. blackburni had cornua that were distinctly hooked at their distal ends (Fig. 4). The exception for this second state was S. pedemontanum. Sympetrum illotum, only, had cornua that were rounded into paddle-like shapes at the tip.
-
5.
Setal patches of S4 of penis: (0) absent; (1) present.
These tufts of wide setae (Fig. 4a) were found only in members of the flaveolum group, but not in S. flaveolum or S. madidum.
-
6.
Basal medial nodule on S4 sclerite of penis: (0) absent; (1) present.
For most Sympetrum, the distal edge of the S4 sclerite was without ornamentation. The flaveolum and infuscatum species groups, however, had a distinct median nodule along this distal margin (Fig. 4). Sympetrum sanguineum of the vulgatum group also exhibited this nodule.
-
7.
Width of penile S4: (0) widest at base; (1) widest apically where cornua originate; (2) subequal width throughout.
For Sympetrum species, the fourth segment of the penis was either widest at its base at the distal margin of the third segment, widest at its distal margin, or nearly the same width throughout its length. Members of the pedemontanum group were the only taxa to exhibit a wider base than distal margin for S4. The flaveolum and vulgatum groups along with S. illotum and N. blackburni had S4 sclerites that were widest at their distal margins. Members of the danae, fonscolombii, and infuscatum groups had S4 sclerites of nearly equal widths throughout. Within these clades, however, several species (S. danae, S. darwinianum, S. depressiusculum, S. frequens, and S. maculatum) had S4 sclerites that were widest at their distal margins. This character certainly varied in its utility for distinguishing species groups.
-
8.
Ligula notch of S2 of penis: (0) prominent/hooked but less than 1/3 width of S2; (1) larger than 1/3 width of S2.
The ligula is a hooked structure responsible for helping hold the penis internally when not in use. All Sympetrum species, except S. villosum, had ligulae that were less than a third of the width of the second penis segment. The outgroup Leucorrhinia species had larger ligulae than those found in Sympetrum.
-
9.
Cornua origin: (0) in plane of lateral lobes of S4; (1) above plane of lateral/central lobes of S4.
For most Sympetrum species, the cornua originated from the same area of S4 as the lateral lobes. For N. blackburni, S. illotum, and the flaveolum group, the cornua arose from the S4 margin above where the lateral and central lobes originated.
-
10.
Distance between cornua: (0) subparallel along length; (1) diverging apically; (2) converging apically.
For the flaveolum and infuscatum groups, and also for S. illotum, the cornua were basically parallel along their length. When viewed ventrally, the cornua of the danae, fonscolombii, pedemontanum, and vulgatum groups along with N. blackburni were distinctly divergent along their length. Converging cornua was an autapomorphy for S. vicinum.
-
11.
Penis vesicle (S1): (0) smooth at base; (1) basal margin projecting outward.
-
12.
Lateral margin of S1 of penis: (0) smoothly curved; (1) angular projection at widest point.
Characters 11 and 12 showed variability within the Sympetrum clades and therefore were not phylogenetically informative with respect to relationships among species groups. These characters could be useful as a key character, but not as a synapomorphy for a species group.
-
13.
Lateral margin of S4 sclerite of penis: (0) weak/absent; (1) expanded ventrally beyond cornua; (2) expanded dorsally behind cornua; (3) expanded dorsally and ventrally beyond cornua.
The lateral margin of the S4 sclerite had multiple states. Sympetrum gracile, S. infuscatum, and S. risi had S4 sclerites that were not expanded on their lateral margins. Members of the flaveolum, fonscolombii, and pedemontanum groups had S4 sclerites that were expanded only ventrally beyond the cornua. The S4 sclerites were expanded dorsally for N. blackburni, S. darwinianum, S. illotum, S. maculatum, and the danae group. The vulgatum group had S4 sclerites that were expanded dorsally and ventrally, except for S. signiferum and S. vicinum, which had sclerites that were only ventrally expanded.
-
14.
S4 sclerite of penis surface: (0) smooth; (1) heavily contoured with bumps and ridges.
A heavily contoured S4 sclerite was a synapomorphy for the flaveolum group and did not occur in any other taxa.
-
15.
Central unexpanded lobe of S4 of penis: (0) in line w/ lateral lobes; (1) above plane of lateral lobes; (2) below plane of lateral lobes.
For many taxa, the lateral and central lobes of the last segment of the penis were small and arranged in the small plane (state 0) as in the infuscatum group. Some species of the pedemontanum and vulgatum groups had a central lobe that was above the plane of the lateral lobes (state 1). Many taxa, including the outgroups Celithemis and Leucorrhinia, S. illotum, N. blackburni, and the fonscolombii and danae groups, had a central lobe that was well below the plane of the lateral lobes (state 2).
-
16.
Width of S1 of penis: (0) more than ½ S1 length; (1) less than ½ S1 length.
For most species, including outgroups, the width of the 1st penal segment was much greater than half its length. Sympetrum fonscolombii and S. villosum of the fonscolombii group were the only species that had a thin 1st penal segment.
-
17.
Visibility of cornua: (0) obscured by central/lateral lobes of S4; (1) visible along nearly entire length.
For species in the fonscolombii group, the cornua are so short that their visibility is obscured by the central and lateral lobes of the last penis segment. This character state was a synapomorphy for this species clade.

-
18.
Ventral tooth of cerci: (0) prominent; (1) weak; (2) absent.
In many libellulid taxa, the cerci (superior anal appendages) of the male have a prominent tooth-like projection on the ventral edge prior to the cerci tapering to a point. This tooth was found in outgroups, and all species of the flaveolum, infuscatum, and pedemontanum groups. This tooth was completely absent in the fonscolombii group, but varied between weak and prominent within the danae and vulgatum groups.
-
19.
Curve of dorsal side of cerci: (0) straight; (1) curved dorsally; (2) curved ventrally.
In libellulids, the cerci can remain uncurved along their length, or can curve dorsally or ventrally as they taper to a point. In this study, ventral curving was rare and only occurred in the outgroup taxa Celithemis, and Leucorrhinia hudsonica, and in the ingroup taxa N. blackburni and S. illotum. All the species of the danae, fonscolombii, infuscatum, and vulgatum groups had uncurved cerci. The species of the pedemontanum group consistently had dorsal curving cerci, and the flaveolum group varied between uncurved and dorsally curved cerci. The utility of this character may, however, be somewhat doubtful due to individual variation. As the adult emerges from the exuviae, the cerci may be distorted and would then harden, possibly with curving that may not occur for all members of that species.
-
20.
Epiproct length: (0) subequal to ventral projection of cerci; (1) longer than the ventral projection of cerci; (2) shorter than the ventral projection of cerci.
In Anisoptera, the epiproct is the inferior anal appendage of the male and is typically shorter than the full length of the cerci. The epiproct may extend beyond the ventral projection of the cerci (see character 19), be subequal in length to this projection, or not reach the projection. The danae and fonscolombii groups were consistent with all species having an epiproct subequal to the ventral projection of the cerci. The species of the infuscatum and pedemontanum groups had epiprocts that extended beyond the ventral projection. Members of the flaveolum and vulgatum groups were variable with some species having a subequal length, and some species having a much shorter epiproct.
-
21.
Dorsal apical teeth of epiproct: (0) absent; (1) present.
The distal edge of the dorsum of the epiproct often had several tooth-shaped projections. After examining all the taxa here, only Nesogonia blackburni was found to be without these tooth-like projections.
-
22.
Genital lobe shape: (0) pointed; (1) rounded; (2) quadrate.
The genital lobe of libellulids projects from the posterior margin of the genital fossa in males. This lobe may taper to a point, have a rounded margin, or have a quadrate shape. The species of the danae group all had rounded genital lobes. The only taxon to have a quadrate lobe was S. corruptum. The other species clades varied between pointed and rounded genital lobes. The utility of this character was doubtful, partly due to within-clade variation, but also because of difficulty coding individuals as either pointed or rounded.
-
23.
Genital lobe angle: (0) ventral; (1) ventro-posterior; (2) posterior.
The genital lobe may point directly ventral, ventro-posterior, or directly posterior. All species of the infuscatum group had ventro-posterior angled genital lobes, but the other species clades varied in the states within these clades. Coding this character was difficult, because consistently determining the angle of the lobe was problematic. Intraspecific variation occurs for some species.
-
24.
Carina of abdominal segment 4: (0) present; (1) absent.
The presence of a transverse carina on the 4th abdominal segment was used as a key character in Needham et al. (2000), and was described as a possible character for validating Tarnetrum. Further examination of this character showed that it may vary intraspecifically, and that it was found in many other taxa that were never considered to belong in Tarnetrum.
-
25.
Lateral ridges of occiput: (0) absent; (1) present.
Lateral ridges were found only on species of the flaveolum group, except for S. flaveolum and S. madidum.

Rights and permissions
About this article
Cite this article
Pilgrim, E.M., von Dohlen, C.D. Phylogeny of the dragonfly genus Sympetrum (Odonata: Libellulidae). Org Divers Evol 12, 281–295 (2012). https://doi.org/10.1007/s13127-012-0081-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13127-012-0081-7