
Abstract
The common Japanese conger Conger myriaster is an important commercial coastal fisheries species in East Asia, but its spawning area has not been determined. A larval sampling survey was conducted in September 2008 along 136°E between 13°N and 22°N, which roughly followed the Kyushu-Palau Ridge in the western North Pacific. Twenty larval specimens were confirmed to be C. myriaster using DNA analysis. Two were newly hatched larvae (preleptocephali) 5.8 and 7.8 mm in total length (TL), which were caught at 17°N. The 5.8 mm TL larva was estimated to be 3–4 days after hatching, the youngest preleptocephalus (i.e., the earliest stage) of this species ever collected. Eighteen other leptocephali were caught at 18°N and 21°N, and these ranged from 18.6 to 40.0 mm TL. Based on these collections, we discerned that there is a spawning area of C. myriaster in the area along the Kyushu-Palau Ridge approximately 380 km south of Okinotorishima Island. Similar to the Japanese eel spawning area along the West Mariana Ridge, the Kyushu-Palau Ridge may play an important role as a landmark of the spawning area. The discovery of this offshore spawning area should lead us to a better understanding of the recruitment mechanisms of C. myriaster, and help to facilitate future international management efforts.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The common Japanese conger Conger myriaster (also called the whitespotted conger) is an important commercial fish species in the seas around Japan, Korea [1, 2] and the East China Sea [3]. It mainly inhabits shallow coastal waters to the edge of the continental shelf (e.g., the East China Sea) in temperate regions [4–6], and it is absent in subtropical areas such as the Ryukyu Islands in southern Japan, where other Conger species are present. Because mature individuals have not been collected from inshore waters to the continental margin, where many adults are caught commercially, the spawning area of this species had been presumed to occur somewhere in deeper offshore waters [5–7]. However, until recently, C. myriaster larvae (leptocephali) had been only caught in relatively shallow coastal areas of East Asia (Japan and Korea) [7–15], including brackish waters of the upper estuaries of rivers [16] and over the continental shelf in the East China Sea [17], and most of them were large [80–130 mm in total length (TL)] in the late leptocephalus or metamorphic stages.
Recently, the distribution of larvae of C. myriaster was studied using molecular genetic analysis of collections in the East China Sea along the shelf break near the Kuroshio mainstream [18, 19] and south of the Ryukyu Islands [19]. In addition, a likely spawning area in the North Equatorial Current region was discovered, based on the collection of six genetically identified preleptocephali at one station and by the historical collection of possible C. myriaster leptocephali in this region that were not genetically identified [20]. This was considered to be in an analogous location to the offshore spawning area of the American conger eel, C. oceanicus, in the western North Atlantic [20, 21]. The offshore distribution of these larvae showed that C. myriaster spawned in an offshore area far south of its growth habitats in East Asia, and that its adults and larvae perform long-distance migrations similar to catadromous eel species such as the Japanese eel Anguilla japonica, which has been found to spawn in a narrow area along the West Mariana Ridge [22–25]. However, further confirmation beyond those larvae caught at one station is needed to help determine the location of the spawning area of C. myriaster and identify the possible landmarks used to find the spawning area.
Understanding the precise location of the spawning area of this species is of critical importance, because landings of C. myriaster in Japanese waters declined from 13,000 to 6,300 metric tons during 1995–2008 (source: Ministry of Agriculture, Forestry and Fisheries of Japan, Tokyo), despite the fact that various stock management approaches were implemented during this period, such as a reduction in fishing efforts targeting smaller individuals in major local fishing grounds in Japanese coastal areas [26, 27]. The landings in Korea declined from 20,000 to 8,000 tons during 1985–2001, and then rapidly increased to nearly 20,000 tons (source: FAO, Rome). These resource fluctuations and the lack of statistical information about the East China Sea landings of C. myriaster in China are triggering concern about the decline of this important fisheries species. In order to implement an efficient and stable stock management plan, the spawning area, recruitment mechanisms, and resource structure of C. myriaster throughout East Asia needs to be much better understood.
In the present study, we describe the collection of genetically identified larvae of C. myriaster in relation to seafloor structure and ocean current flow in the survey area which indicate the presence of a spawning area of this species in this particular area along the Kyushu-Palau Ridge, and we discuss its larval migration toward East Asia, which could lead to further research on the C. myriaster recruitment mechanism.
Materials and methods
Research cruise and collection of C. myriaster larvae
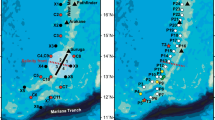
The C. myriaster collections were made as a part of the expedition to study the spawning ecology of the Japanese eel that was conducted by the R/V Kaiyo Maru of the Fisheries Agency of Japan (KY-08-4, 20 August to 15 September 2008) [24] along a 12°N, 135.5°E to 13°N, 136°E line and a 136°E line between 13°N and 22°N that roughly followed the Kyushu-Palau Ridge in the North Pacific (Fig. 1) from 5 to 8 September 2008. Larval collections were conducted at ten stations at 1°N latitudinal intervals (Fig. 1) by a 3 m Isaacs–Kidd midwater trawl (IKMT), which had a 0.5 mm mesh and a mouth opening of 8.7 m2. Tows of the IKMT fished from a depth of 300 m to the surface with step towing ranging from 150 to 50 m in depth, and took approximately 60 min (including the 45 min step towing) during both day and night. Hydrographic observations consisting of conductivity, temperature, and depth measurements (CTD) up to 1000 m were carried out at the ten stations of the 136°E line using a SBE-911plus sensor (Sea-Bird Electronics, USA). Multilevel current velocities were observed along the ship track by a shipboard acoustic Doppler current profiler (ADCP); 38 kHz narrow band (RD Instruments, USA). The velocity data were horizontally gridded into 0.05° boxes, where the velocities included were ensemble averaged.

Survey area along the Kyushu-Palau ridge on the 136°E line between 13°N and 22°N in September 2008. Closed circles indicate the sampling stations at which an Isaacs-Kidd midwater trawl (IKMT) net survey and conductivity–temperature–depth (CTD) observations were conducted. Stars with circles indicate stations where Conger myriaster larvae were collected. The black square shows the station where preleptocephali were collected in June 2008 [20]
Identification of Conger myriaster larvae
Anguilliform larvae (leptocephali and preleptocephali) were sorted out of the plankton samples on board and measured to the nearest 0.1 mm in total length (TL). The total number of myomeres (TM) and the position of the last vertical blood vessel on the myomeres (LVBV) were recorded using a binocular microscope unless the specimen was too damaged. Conger leptocephali were morphologically identified to the genus level based on body shape, pigmentation pattern, and/or number of TM (i.e., 138–149) and LVBV (i.e., 50–59) [28, 29], and then preserved in 99% ethanol. In the laboratory, the samples of Conger leptocephali and preleptocephali were identified to species by analyzing nucleotide sequences of approximately 550 base pairs (bp) containing the mitochondrial DNA 16S rRNA region using the primers 5′-GGTCCWRCCTGCCCAGTGA-3′ and 5′-CCGGTCTGRACYAGATCACGT-3′. There were sequence differences of 3–15% in the fragments among each of the four Conger species present in the western north Pacific (C. myriaster, C. japonicus, C. erebennus, C. cinereus [4]), and a maximum difference of 1% within the same Conger species [18], that were compared to the known sequences of C. myriaster [18, 19, 30].
Results
Collection of Conger myriaster leptocephali and preleptocephali
A total of 20 Conger myriaster larvae were caught at three stations at 17°N, 18°N, and 21°N along the 136°E line (Fig. 1; Table 1) from 6 to 7 September 2008, and these were confirmed to be C. myriaster by analyzing mitochondrial DNA fragments of approximately 550 bps containing the 16S ribosomal RNA region in the laboratory. Comparisons of the sequences of these 20 specimens (DDBJ/EMBL/GenBank accession numbers AB617683–AB617702) with those of known C. myriaster specimens found sequence identities of 99.2–100%. The two that were collected at 17°N (Figs. 1, 2) were 5.8 and 7.8 mm TL (Table 1) and were newly hatched larvae (preleptocephali). The 5.8 mm TL larva had no teeth or jaws, and had an early pigmented eye with an oil globule (Fig. 2a), which was the youngest preleptocephalus (i.e., earliest stage) of this species ever collected. The other 18 larvae collected at 18°N and 21°N ranging from 18.6 to 40.0 mm TL had myomere counts that ranged from 141 to 149 TM and 51 to 57 LVBV (Table 1), and had an elongate body, pigment spots on the gut, and a crescentic patch of pigment beneath the eye, which are typical morphological features of all known species of Conger leptocephali [18, 19, 28, 29].

Newly hatched larvae of Conger myriaster collected at 17°N, 136°E. a Early-stage larvae: 5.8 mm in total length with no teeth or jaws, and early eye pigmentation with an oil globule; b 7.8 mm in total length with teeth, jaws, and eye pigmentation. Scale bars are 1 mm
Oceanographic features
Current flow along the 136°E line
The shipboard ADCP observations near the surface at 40 m (Fig. 3a) and in the subsurface at 112 m (Fig. 3b) along the ship track of the transect from 12°N, 135.5°E to 13°N, 136°E and the 136°E line between 13 and 22°N (Fig. 3) showed alternating east- and west-flowing currents and countercurrents or eddies, which indicated a mostly continuous steady westward current (the North Equatorial Current, NEC) between 12°N and 14°N, a strong eastward current at around 17°N, and an eastward current (possibly the Subtropical Countercurrent, STCC) between 21°N and 22°N (Fig. 3). The current at 17°N, 136°E, where newly hatched larvae were caught (Fig. 1), was flowing eastward or northeastward at about 0.45–0.24 m/s near the surface and in the subsurface layer (Fig. 3).

Acoustic Doppler current profiler (ADCP) current vectors at depths of 40 m (a) and 112 m (b) along the line from 12°N, 135.5°E to 13°N, 136°E and the 136°E line between 13°N and 22°N, which were obtained from 4 to 7 September 2008
Hydrographic structure at 17°N, 136°E
Figure 4 shows the CTD profile down to a mean depth of 1000 m at 17°N, 136°E, where newly hatched preleptocephali were caught. Water temperature at the net deployment depths (0–300 m) ranged from approximately 16 to 30°C, and within the 50–150 m depth layer where the step tow was conducted, the temperature ranged from 23 to 29°C. The salinity profile indicated that the survey areas were in the North Pacific Tropical Water (NPTW), which showed a salinity maximum (>35.0 PSU) in the subsurface layer at a depth of around 175 m. The density drastically increased from σt = 21–24.5 in the 50–200 m depth layer associated with pycnocline.

Vertical profiles of water temperature (gray line), salinity (black line), and density (dashed line) (σt) at 17°N, 136°E, where newly hatched larvae of C. myriaster were caught
Discussion
Spawning area of Conger myriaster along the Kyushu-Palau Ridge
The collection of newly hatched larvae of Conger myriaster as well as larger specimens in the present study indicates that this species had spawned in the open ocean near the survey area around 17°N, 136°E, approximately 205 nautical miles (380 km) south of Okinotorishima Island (20°25′N, 136°04′E), in early September 2008 and in previous months. The location at which these preleptocephali were collected in September (17°N, 136°E) was remarkably close to the location (16°N, 137°E) at which six preleptocephali had been collected several months earlier, in June 2008, and these were also genetically confirmed to be C. myriaster [20]. The presence of larger genetically identified larvae to the north of this area in the present study and the inferred presence of larger leptocephali of this species in July of 1991 and 1995 that could not be genetically identified [20] are strong evidence that C. myriaster spawns in this particular area of the western North Pacific. Combining these data, it can also be concluded that C. myriaster spawns in this area between at least June and September, which is several months earlier than previous estimations of the spawning season obtained by back-calculation based on otolith daily increments of large leptocephali of this species, which peak from about September to February [11, 17].
In the present study, the preleptocephali had only recently hatched, so they must have been collected relatively close to their spawning location. The smallest larva (5.8 mm TL), which had no teeth or jaws, and early pigmented eyes with an oil globule, was similar to the Anguilla japonica preleptocephali estimated to be 3–4 days after hatching based on their eye pigmentation state and otolith analyses [22]. They were also similar in shape and size to the preleptocephali collected in this area in June 2008 (5.6–6.9 mm), which mostly had pigmented eyes and early teeth [20]. Reared larvae of Conger myriaster from artificial fertilization were reported to develop such that they reached about 5.8 mm TL at 4–5 days after hatching and mouth opening was observed at 7 days after hatching [31]; thus, the smallest larva (5.8 mm TL) in the present collection was equivalent to 4–6 days after hatching of the reared larvae of C. myriaster. However, their rearing temperature was significantly lower (12–14°C) than the temperature profile of the survey area (approximately 16–30°C in the depth range of net deployment—the upper 300 m). Within the 50–150 m depth layer, where the step tow was conducted and the larvae were likely captured, the temperatures ranged from 23 to 29°C. Therefore, it is possible that the age in days of the smallest larva (5.8 mm TL) was about 3–4 days after hatching and that it had experienced rapid growth caused by the warmer water temperature environment.
The easterly current in which the newly hatched larvae were found (17°N, 136°E) was flowing at about 0.24–0.45 m/s (Fig. 3), so the smallest larva (estimated to be 3–4 days after hatching) could have been transported, depending on its depth, 62–156 km after hatching in the western area near the longitudinal axis of the Kyushu-Palau Ridge around 135°E (Fig. 3). This suggests that C. myriaster had spawned in September and probably also in June quite near to or just to the east of the Kyushu-Palau Ridge. This oceanic ridge could play an important role as a landmark for the spawning area, as appears to be the case for the Japanese eel that spawns just to the west of the West Mariana Ridge [22–25]. Since the oceanic ridge crest within the present survey area (13–22°N) is quite deep (approximately 2,500–3,500 m deep), except for Okinotorishima Island, C. myriaster spawning appears to occur in a pelagic environment similar to that of the Japanese eel, which has been studied through the collection of mature adults [23–25], naturally spawned eggs, and newly hatched preleptocephali [25] along the West Mariana Ridge. The Kyushu-Palau Ridge is at the boundary between the Philippine Basin (5,000–6,000 m deep) and the West Mariana Basin (approximately 4,000–5,000 m deep), and may provide longitudinal cues for C. myriaster adults to aggregate and spawn due to possible geomagnetic anomalies if they have a magnetic sense like anguillid eels [25, 32]. Other cues such as olfactory cues could also be possible if they migrate to the spawning area at deeper depths than anguillid eels. The Japanese eel and other anguillid eels migrate at depths of less than about 800 m and show vertical migrations to shallower depths at night [33]. However, the maturation of C. myriaster appears to occur at much colder temperatures of about 6°C [34] than the maturation temperatures of about 19–22°C seen for the Japanese eel [35, 36]. This suggests the possibility that C. myriaster migrates at deeper depths than the Japanese eel, and that the Kyushu-Palau Ridge could be a landmark used by this species to locate the area of aggregation and spawning. Based on the molecular phylogenetic relationships of anguilliform fishes, conger eels and freshwater eels evolved through different lineages, even though their general body morphologies are quite similar, so conger eels appear to be most related to the shelf and slope eels of the Nettastomatidae and may have retained characteristics of deep-benthic eels such as the cutthroat eels of the Synahobranchidae, while freshwater eels are most related to the midwater pelagic deep-sea eels such as the gulper eels of the Eurypharyngidae and the saw-tooth eels of the Serrivomeridae [37].
However, further studies are needed to determine the larger-scale distribution of spawning and to learn about possible latitudinal cues for spawning, such as fronts [38] or current flows, which have been suggested to be important in determining the locations of Japanese eel spawning [25, 39].
Laval distribution and transport
The collections of C. myriaster larvae were limited to the northern part of our survey area (17–21°N), which belongs to the marginal area adjacent to the northernmost part of the NEC (south of approximately 15°N [40]) and covers the STCC region (north of 19–20°N [40–43]). In contrast, the larvae were absent from the southern region (south of 16°N), which belongs to the mainstream of the NEC as a stable westward current [40].
Since the spawning area of the Japanese eel was found to occur along the West Mariana Ridge within the latitudes of the main part of the NEC (12–14°N) [22–25], with their larvae being transported westward by the NEC and then transferring to the Kuroshio to recruit to East Asia [39, 44–46], it appears that C. myriaster has a slightly different spawning and recruitment strategy. It has also been estimated that C. myriaster larvae are transported westerly toward their nursery areas along the coasts of East Asia [18–20], but their transport may be more affected by eddies and countercurrents at more northern latitudes. C. myriaster larvae (22.3, 46.5 mm TL) were reportedly collected near the western boundary of the western North Pacific, south of the Ryukyu Islands (approximately 21°N, 125°E) [19]. However, it is unlikely that westerly larval transport occurs via the simple, stable, westward flow of the NEC because of the complex current structure, including the likely influence of the eastward current of the STCC within the area of 17–21°N. This area is along the southern edge of a region with much greater sea surface height (SSH) variability due to the occurrence of many eddies [43, 47].
The shipboard ADCP observations along the 136°E transect showed alternating east- and west-flowing currents and countercurrents (Fig. 3), which may have been associated with mesoscale eddies. Similar alternating bands of eastward and westward water flow were also seen in the Doppler current profile along 137°E in July and August in a previous study [41]. Because both small and large C. myriaster larvae appear to have been collected mostly north of 16°N in this area previously [20], it appears that this spawning area of C. myriaster may be typically located just to the north of the main part of the NEC, within a band of westerly flow that likely exists between two subtropical fronts associated with eastward countercurrents in this region [38]. Various sizes of C. myriaster larvae (5.8–40.0 mm TL) were collected in the range of 17–21°N along 136°E in the present study (Table 1), indicating larval retention in the eddies in this region. However, SSH observations show that mesoscale eddies propagate westward through this area [48], which is just along the typical southern edge of the STCC [43], through the influence of the baroclinic Rossby waves that propagate westward across the Pacific basin in 2.5–3 years [43]. Therefore, the C. myriaster larvae trapped within the mesoscale eddies could be transported to the western boundary in several months, even if they were temporarily entrained into eastward currents such as the STCC. Mesoscale eddies are also considered to have an influence on the larval migration route of the Japanese eel [49], and the possible role of the directional swimming of these migrating Conger and Anguilla leptocephali to increase their westward transport is not yet known. Further studies on larval transport simulations using physical oceanographic models for the western North Pacific subtropical current system are required to elucidate the recruitment mechanism of C. myriaster, which should lead to international control of the stock management of this important fisheries resource.
References
Mochioka N, Tokai T (2001) Ecology and fisheries of Conger myriaster. Kaiyo Monthly 33:525–528 (in Japanese)
Park CD (2001) Conger-eel fisheries in Korea. Nippon Suisan Gakkaishi 67:127–128 (in Japanese)
Tokimura M (2001) Conger-eel fisheries and fisheries resources in the East China Sea. Nippon Suisan Gakkaishi 67:125–126 (in Japanese)
Hatooka K (2002) Congridae. In: Nakabo T (ed) Fishes of Japan with pictorial keys to the species, English edn. Tokai University Press, Tokyo, pp 227–234
Ochiai A, Tanaka M (1986) Maanago Conger myriaster. In: Ichthyology, 2nd edn. Koseisha-Kosei-kaku, Tokyo, pp 576–580 (in Japanese)
Yamada U (1986) Conger myriaster. In: Okamura O (ed) Fishes of the East China Sea and the Yellow Sea. Seikai Reg Fish Res Lab, Nagasaki, pp 68–69 (in Japanese)
Takai T (1959) Studies on the morphology, ecology and culture of the important apodal fishes, Muraenesox cinereus (Forsskål) and Conger myriaster (Brevoort). J Shimonoseki Coll Fish 8:209–555 (in Japanese with English summary)
Kubota S (1961) Studies on the ecology, growth and metamorphosis of a conger eel, Conger myriaster (Brevoort). J Fac Fish Pref Univ Mie 5:90–370 (in Japanese with English summary)
Tanaka K, Tabeta O, Mochioka N, Yamada J, Kakuda S (1987) Otolith microstructure and ecology of the conger eel (Conger myriaster) larvae collected in the Seto Inland Sea, Japan. Nippon Suisan Gakkaishi 53:543–549
Mochioka N (1988) Congridae. In: Okiyama M (ed) An atlas of the early stage fishes in Japan. Tokai University Press, Tokyo, pp 44–54 (in Japanese)
Lee TW, Byun JS (1996) Microstructural growth in otoliths of conger eel (Conger myriaster) leptocephali during the metamorphic stage. Mar Biol 125:259–268
Onuma Y (1996) Metamorphosing location of Conger myriaster leptocephali. Bull Fish Exp St Ibaraki-ken 33:103–107 (in Japanese)
Otake T, Ishii T, Ishii T, Nakahara M, Nakamura R (1997) Changes in otolith strontium:calcium ratios in metamorphosing Conger myriaster leptocephali. Mar Biol 128:565–572
Mochioka N (2001) Early life history of Conger myriaster. Nippon Suisan Gakkaishi 67:111–112 (in Japanese)
Katayama S, Shimizu Y (2006) Occurrence pattern of conger eel larva, Conger myriaster in south Tohoku area. Bull Jpn Soc Fish Oceanogr 70:10–15
Yagi Y, Kodono N, Kinoshita I, Fujita S (2010) Late-stage metamorphosing Conger myriaster leptocephali collected in a river estuary of Ariake Bay, Japan. Ichthyol Res 57:310–313
Kurogi H, Mochioka N, Takai Y, Tabeta O (2002) First offshore record of Conger myriaster leptocephali in the East China Sea. Fish Sci 68:1155–1157
Kurogi H (2006) Study on the inshore migration mechanism of Conger myriaster leptocephali (Ph.D. thesis). Kyushu University, Fukuoka (in Japanese)
Ma T, Miller MJ, Aoyama J, Tsukamoto K (2007) Genetic identification of Conger myriaster leptocephali in the East China Sea region. Fish Sci 73:989–994
Miller MJ, Yoshinaga T, Aoyama J, Otake T, Mochioka N, Kurogi H, Tsukamoto K (2011) Offshore spawning of Conger myriaster in the western North Pacific: evidence for convergent migration strategies of anguilliform eels in the Atlantic and Pacific. Naturwissenschaften 98:537–543
McCleave JD, Miller MJ (1994) Spawning of Conger oceanicus and Conger triporiceps (Congridae) in the Sargasso Sea and subsequent distribution of leptocephali. Environ Biol Fish 39:339–355
Tsukamoto K (2006) Spawning of eels near a seamount. Nature 439:929
Chow S, Kurogi H, Mochioka N, Kaji S, Okazaki M, Tsukamoto K (2009) Discovery of mature freshwater eels in the open ocean. Fish Sci 75:257–259
Kurogi H, Okazaki M, Mochioka N, Jinbo T, Hashimoto H, Takahashi M, Tawa A, Aoyama J, Shinoda A, Tsukamoto K, Hideki T, Gen K, Kazeto Y, Chow S (2011) First capture of post-spawning female of the Japanese eels Anguilla japonica at the southern West Mariana Ridge. Fish Sci 77:199–205
Tsukamoto K, Chow S, Otake T, Kurogi H, Mochioka N, Miller MJ, Aoyama J, Kimura S, Watanabe S, Yoshinaga T, Shinoda A, Kuroki M, Oya M, Watanabe T, Hata K, Ijiri S, Kazeto Y, Nomura K, Tanaka H (2011) Oceanic spawning ecology of freshwater eels in the western North Pacific. Nat Commun 2:179. doi:10.1038/ncomms1174
Gorie S, Tanda M, Nagasawa K (2010) Movement and growth of white-spotted conger Conger myriaster in the Eastern Seto Inland Sea, Japan. Aquaculture Sci 58:233–242
Gorie S, Nagasawa K (2010) Habitat preference and feeding habits of juvenile white-spotted conger Conger myriaster in the Eastern Seto Inland Sea, Japan. Aquac Sci 58:167–179
Tabeta O, Mochioka N (1988) Leptocephali. In: Okiyama M (ed) An atlas of the early life stage fishes in Japan. Tokai Univ Press, Tokyo, pp 15–64 (in Japanese)
Smith DG (1989) Family Congridae: leptocephali. In: Böhlke EB (ed) Fishes of the Western North Atlantic, part 9, vol 2: Leptocephali. Sears Foundation for Marine Research, New Haven, pp 723–763
Inoue JG, Miya M, Tsukamoto K, Nishida M (2001) Complete Mitochondrial DNA sequence of Conger myriaster (Teleostei: Anguilliformes): novel gene order for vertebrate mitochondrial genomes and the phylogenetic implications for anguilliform families. J Mol Evol 52:311–320
Horie N, Utoh T, Yamada Y, Okamura A, Zhang H, Mikawa N, Akazawa A, Tanaka S, Oka HP (2002) Development of embryos and larvae in the common Japanese conger Conger myriaster. Fish Sci 68:972–983
Nishi T, Kawamura G (2005) Anguilla japonica is already magnetosensitive at the glass eel phase. J Fish Biol 67:1213–1224
Manabe R, Aoyama J, Watanabe K, Kawai M, Miller MJ, Tsukamoto K (2011) First observations of the oceanic migration of the Japanese eel using pop-up archival transmitting tags. Mar Ecol Prog Ser 437:229–240
Utoh T, Horie N (2011) Studies on the technique of seed production of the common Japanese conger Conger myriaster. Nippon Suisan Gakkaishi 77:775–778 (in Japanese)
Ohta H, Kagawa H, Tanaka H, Okuzawa K, Hirose K (1996) Changes in fertilization and hatching rates with time after ovulation induced by 17,20β-dihydroxy-4-pregnen-3-one in the Japanese eel, Anguilla japonica. Aquaculture 139:291–301
Horie N, Utoh T, Mikawa N, Yamada Y, Okamura A, Tanaka S, Tsukamoto K (2008) Influence of artificial fertilization methods of the hormone-treated Japanese eel Anguilla japonica upon the quality of eggs and larvae (comparison between stripping–insemination and spontaneous spawning methods). Nippon Suisan Gakkaishi 74:26–35 (in Japanese with English abstract)
Inoue JG, Miya M, Miller MJ, Sado T, Hanel R, López JA, Hatooka K, Aoyama J, Minegishi Y, Nishida M, Tsukamoto K (2010) Deep-ocean origin of the freshwater eels. Biol Lett 6:363–366
Aoki Y, Suga T, Hanawa K (2002) Subsurface subtropical fronts of the North Pacific as inherent boundaries in the ventilated thermocline. J Phys Oceanogr 32:2299–2311
Kimura S, Tsukamoto K (2006) The salinity front in the North Equatorial Current: a landmark for the spawning migration of the Japanese eel (Anguilla japonica) related to the stock recruitment. Deep Sea Res II 53:315–325
Hasunuma K, Yoshida K (1978) Splitting of the subtropical gyre in the Western North Pacific. J Oceanogr Soc Jap 34:160–172
Kaneko I, Takatsuki Y, Kamiya H, Kawae S (1998) Water property and current distributions along the WHP-P9 section (137–-142°E) in the western North Pacific. J Geophys Res 103:12959–12984
Qiu B, Chen S (2010) Interannual-to-decadal variability in the bifurcation of the North Equatorial current off the Philippines. J Phys Oceanogr 40:2525–2538
Qui B, Chen S (2010) Interannual variability of the North Pacific Subtropical Countercurrent and its associated mesoscale eddy field. J Phys Oceanogr 40:213–225
Tsukamoto K (1992) Discovery of the spawning area for Japanese eel. Nature 356:789–791
Kimura S, Tsukamoto K, Sugimoto T (1994) A model for the larval migration of the Japanese eel: roles of the trade winds and salinity front. Mar Biol 119:185–190
Shinoda A, Aoyama J, Miller MJ, Otake T, Mochioka N, Watanabe S, Minegishi Y, Kuroki M, Yoshinaga T, Yokouchi K, Fukuda N, Sudo R, Hagihara S, Zenimoto K, Suzuki Y, Oya M, Inagaki T, Kimura S, Fukui A, Lee TW, Tsukamoto K (2010) Evaluation of the larval distribution and migration of the Japanese eel in the western North Pacific. Rev Fish Biol Fish. doi:10.1007/s11160-010-9195-1
Kobashi F, Kawamura H (2001) Variation of sea surface height at periods of 65–220 days in the subtropical gyre of the North Pacific. J Geophys Res 106:26817–26831
Qiu B (1999) Seasonal eddy field modulation of the North Pacific subtropical countercurrent: TOPEX/Poseidon observations and theory. J Phys Oceanogr 29:2471–2486
Kim H, Kimura S, Shinoda A, Kitagawa T, Sasai Y, Sasaki H (2007) Effect of El Niño on migration and larval transport of the Japanese eel (Anguilla japonica). ICES J Mar Sci 64:1387–1395
Acknowledgments
The authors thank Captain Nobuyuki Nagai and the other crew members of the R/V Kaiyo Maru for supporting the sample collections; Kyoichi Kawaguchi, Tokio Wada, and Misao Arimoto for organizing this research cruise; Hiroto Imai, Kohichi Tahara, and Tsuyoshi Koga for their management of the R/V Kaiyo Maru cruises; Jun Aoyama, Akira Shinoda, and Atsushi Tawa for their enthusiastic sampling and sorting onboard; Yoichi Miyake for valuable comments on the oceanographic conditions; and Mayumi Sato for assisting with the DNA analysis. We also thank the participants in the Workshop on Conger-Eel Fisheries and Biology (ANA-KEN) for helpful discussions. We gratefully acknowledge the financial support of the Fishery Agency of Japan and the Fisheries Research Agency.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Kurogi, H., Mochioka, N., Okazaki, M. et al. Discovery of a spawning area of the common Japanese conger Conger myriaster along the Kyushu-Palau Ridge in the western North Pacific. Fish Sci 78, 525–532 (2012). https://doi.org/10.1007/s12562-012-0468-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12562-012-0468-6