
Abstract
The late early – early middle Eocene climate and vegetation evolution in northern Yakutia (Eastern Siberia) is quantitatively studied in detail for the first time, based on a palaeobotanical record. Palaeobotanical data for this time interval were obtained from 16 palynofloras from continental sediments on the southern shore of Tastakh Lake (the left bank of Indigirka River). In this first integrative study, we applied the Coexistence Approach for quantification of palaeoclimate, the Plant Functional Type (PFT) approach and Integrated Plant Record (IPR) vegetation analysis to quantify palaeovegetation. Our investigation documents the persistence of warm temperate and, at times, nearly tropical climate conditions in the earlier part of the record and the subsequent onset of a cooling trend. Overall humid climate conditions prevailed throughout the late early – early middle Eocene. Our reconstruction indicates low temperature seasonality and relatively strong seasonality in precipitation. Vegetation data as reconstructed are in good agreement with the climate evolution. PFT diversity spectra indicate the prevalence of mixed evergreen-deciduous forests of warm temperate character. The IPR method suggests that the mixed mesophytic forest biome persisted throughout the studied sedimentary succession. The presence of mangroves in our late early Eocene records (at the base of the section) is largely in line with our climate reconstruction and possibly can be related to the Early Eocene Climatic Optimum (EECO) hyperthermal event.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Today, more than 40% of the territory of Yakutia is located within the Arctic Circle and is mostly occupied by mountains, plateaus, and only 1/3 by lowlands. The main part of this region belongs to the middle taiga zone, which is replaced to the north by forest-tundra and tundra zones. Forests occupy about 4/5 of the territory, while meadows occur in river valleys and lowlands. Shrubby herbaceous vegetation and lichens dominate the coast and mountains tops. The Arctic tundra occupies the Lena River Delta and the Laptev Sea coast. The present-day climate of Yakutia is distinctly continental, characterised by long winters and short summers, with a temperature difference of 70–75 °C between the coldest and the warmest month and temperature minima as low as −70 °C, unparalleled in the Northern Hemisphere in terms of the annual number of days with negative temperatures (from 6.5 to 9 months per year) (Tsherbakov 1975).
According to the Intergovernmental Panel on Climate Change (IPCC 2014), the Arctic today is warming dramatically, faster than almost all other parts of our planet, attributing to “polar amplification” (Lee 2014), and impacting global climate feedback and Arctic biota. The consequences of modern climate warming, which are most pronounced in the Arctic, are usually viewed as negative and catastrophic. However, similar events have happened more than once in the past. Moreover, the state of the Earth, in which the poles are free of ice, is more characteristic of our planet. The early Eocene (Ypresian, ca. 56–47.8 Ma) was an exceptional warm interval with reduced pole-to-equator temperature gradients (Suan et al. 2017; Salpin et al. 2019; Suc et al. 2020) and high atmospheric CO2 concentrations ranging from ~1 000 to 3 000 ppm (Pearson and Palmer 2000; Beerling and Royer 2011). Globally, the early Cenozoic, especially Eocene, is increasingly recognised as a key period for understanding the climate and biogeography of warm intervals of Earth's history (Westerhold et al. 2020). Nevertheless, our knowledge of the warm climates of the Arctic and their role in forming Earth's climate is currently insufficient for confident forecasting of climatic events in the future. One of the main reasons for this is the currently prevailing qualitative characteristic of palaeoclimates, which is not suitable for constructing modern mathematical modeling. The solution to this problem is the application of new methods of analysis of fossil climate indicators, which provide quantitative data.
Extensive studies on climate proxies and models in the High Arctic have been conducted for the Cretaceous that revealed palaeoenvironmental conditions and vegetation responses in a “greenhouse” climate (e.g. Spicer and Parrish 1986; Spicer and Corfield 1992; Herman and Spicer 1996, 1997; Amiot et al. 2004; Spicer and Herman 2010; Herman et al. 2016; Spicer et al. 2014, 2019). By contrast, our knowledge of the climate and vegetation dynamics of the Eocene is still fragmentary. Stratigraphic analyses of Paleogene deposits in Northeast Russia in general and Yakutia in particular, as well as the reconstruction of the palaeoclimate and palaeovegetation, were previously tackled by Akhmetiev (1976, 1993, 2015), Akhmetiev et al. (1981, 2010, 2012), Aleksandrova (2016), Baranova and Biske (1979), Bakulina and Zharikova (1989), Belaya and Litvinenko (1989), Fradkina (1995), Fradkina et al. (1979), Grineniko and Kiseleva (1971), Grinenko et al. (1989, 1997), Kulkova (1973a, b, 1987), Kulkova and Laukhin (1975), Laukhin and Fradkina (1984), Laukhin et al. (1988, 1992), L'vova (1989), Narkhinova (1989), Nikitin (2006), Volkova et al. (1990), Volobueva et al. (1988), and others, which were mainly devoted to regional stratigraphy, description of plant fossil remains, new fossil taxa, taxonomic diversity of plants, or palaeofloristic aspects and generally based on qualitative interpretations of macro- and microfloral successions (Budantsev 1997, 1999; Akhmetiev 2015); quantitative studies were largely missing so far.
Recently, quantitative palaeoclimate and vegetation studies have been conducted for regions of the southern Russian Far East (Primorye and Amur Region), revealing major plant biome changes throughout the Paleogene and providing indications of the average annual temperature and the annual precipitation (Bondarenko et al. 2019, 2020a, b, 2021). Such studies are essential to compare results from other proxies or climate models. A most recent study on an early Eocene section located at high latitudes, in the area of the present Lena River Delta (northern Yakutia, Eastern Siberia), provided new insights into short-term climate and vegetation dynamics succeeding the Paleocene – Eocene Thermal Maximum (PETM) (Bondarenko et al. 2022).
Here we present a first higher-resolving time series for the climate and vegetation evolution of northern Yakutia (Eastern Siberia) before, during and after the Early Eocene Climatic Optimum (EECO). Our study is based on the investigation of 16 palynofloras sampled from continental sediments on the southern shore of Tastakh Lake (the left bank of the Indigirka River). We analyse changes in the taxonomic composition of the floras and the abundance of floral components and establish quantitative records to trace regional climate and vegetation dynamics at the time when early Cenozoic ecosystems became established. Moreover, we discuss climate conditions and mangrove’s growth in Arctic Siberia at the end of the early Eocene, and how global signals manifested themselves in northern Yakutia.
Study area
Geological settings and early Cenozoic strata
The Yano-Indigirskaya (Primorskaya) lowland is located in the north of Yakutia, along the shores of the Laptev Sea and the East Siberian Sea. In terms of structural units, the Yano-Indigirskaya lowland belongs to the Lower Indigirskaya Cenozoic plate. The basement includes the Mesozoic folded complex and the unevenly subsided blocks of the Khroma and Novosibirsk median massifs (Bogolepov 1962). The surface of the Yano-Indigirskaya lowland slightly dips towards the sea. Within the lowland relief, there are isolated outliers with a height of 20 to 100 m, composed of Mesozoic and, less often, Paleogene sedimentary and intrusive rocks. The sediment cover of the basement consists of Eocene, Oligocene, Miocene-Pliocene and Quaternary formations (Baranova et al. 1968), regionally, a weathering crust of white kaolin clays of Danian to early Oligocene age is preserved (Vollosovich 1930; Gusev 1954; Baranova et al. 1968). The presence of this weathering crust indicates preservation by long-term relative tectonic inactivity in a sufficiently warm and humid climate.
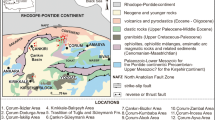
Early Cenozoic deposits of the Yano-Indigirskaya lowland come to the surface on the left bank of the Indirirka River, somewhat south of 70° N. The outcrops near Tastakh Lake (the section 1; Fig. 1a) and Jeloon-Sise Upland (the section 2; Fig. 1a) are erosional remnants rising above the plain level by 25-30 m, with absolute elevations up to 60 m. The studied materials are sampled from coastal cliffs of Tastakh Lake (lat 69°90′ N; long 145°60′ E; Fig. 1a). The base of the outcrop is situated at 21 m a.s.l. (Fig. 1b). The sampled deposits are composed of various facies of alluvium and lacustrine sediments, often changing along the strike. The lithology of the section on the southern shore of Tastakh Lake follows V.F. Goncharov (Goncharov and Kulkova 1970) (Fig. 2).

Map showing the location of the Tastakh Lake section (a) 1 Tastakh Lake section, 2 Jeloon-Sise section; geological profile of the southern shore of Lake Tastakh (b) according to Goncharov and Kulkova (1970)

Correlation chart showing the different stratigraphic interpretations of the Tastakhskaya Formation next to lithological profile of the studied section. The lithological description is taken from Goncharov and Kulkova (1970), depth levels of the samples refer to Kulkova (1973a, b). For the age model of the Tastakhskaya Formation, we follow Grinenko et al. (1997)
Stratigraphy and dating
Cenozoic deposits in Northern Yakutia have been known since the end of the nineteenth century. The first information about the finds of early Cenozoic sediments with layers of coal and amber near the shores of Lake Tastakh is from M. Gedenshtorm in 1830 (cited from Kulkova 1973a, b).
The problems concerning stratigraphy of these Paleogene deposits have been addressed in various studies (Vollosovich 1930; Krishtofovich 1958; Kajalainen 1967, 1970; Pogozhev and Semeykin 1959; Vakulenko 1968; Ivanov 1968, 1969; Dorofeev 1968; Grinenko 1968; Kulkova 1968; Goncharov and Kulkova 1970; Grinenko et al. 1989, 1997; others). The sediments near Tastakh Lake were named the Tastakhskaya Formation after the Interdepartmental Stratigraphic Meeting in 1957 in Magadan (Lobanov 1959; Decisions 1959). The formation of ideas about the age of the Tastakhskaya Formation (II and III horizons) is exemplified in Fig. 2, based on selected works.
A review of geological and palaeobotanical studies shows the early interpretation of the age of these deposits by various researchers (Fig. 2). According to Grinenko et al. (1997), based on materials of medium and large-scale geological survey and prospecting and exploration work, the results of pivot and parametric drilling, thematic and special (mainly palaeontological) long-term studies, the Tastakhskaya Formation is a stratotype of the Tastakhskii regional horizon (the second half of the early – the first half of the middle Eocene). The stratigraphic concept of Grinenko et al. (1997) for the Tastakhskii regional horizon is based on non-palaeontological (interregional profile correlation) and palaeontological (micro- and macrofloras, interregional pollen zones, dinoflagellates) data. Here we follow the stratigraphic concept of Grinenko et al. (1997).
Materials and methods
The floral record
In the present study, microfloras (PFs) from 16 depth levels of the continental Tastakhskaya Formation are analysed, exposed in a section on the southern shore of Tastakh Lake (left bank of Indigirka River) (Fig. 1a). All palynological data are taken from Kulkova (1973a, b). The microfloras yield evidence for a total of 97 different taxa (76 angiosperms, 13 gymnosperms, and 8 pteridophytes) as identified by I.A. Kulkova (Kulkova 1973a, b). Total taxonomical composition is listed in Table 1. The palynological data are represented in the diagram (Fig. 3). As reported in the original studies, up to 300 and more pollen grains were counted per sample. The material was collected by Kulkova in 1964-1970 for conducting complex studies in this territory and is stored in the Trofimuk Institute of Petroleum Geology and Geophysics, Siberian Branch of the Russian Academy of Sciences (IPGG SB RAS).

Palynological diagram (according to Kulkova 1973b)
Quantitative palaeoclimate reconstruction – Application of the coexistence approach (CA)
To reconstruct climate from the palynomorph record, we used the Coexistence Approach (CA) (Mosbrugger and Utescher 1997; Utescher et al. 2014). This approach is organ-independent, so that both macro- and microfossil plants are eligible as long as their modern botanical affinities are determinable (Mosbrugger and Utescher 1997; Utescher et al. 2007; Bruch et al. 2011). For a detailed description of the method, the reader is referred to the original papers describing the procedure (Mosbrugger and Utescher 1997; Utescher et al. 2014). For climatic requirements of extant plants, we used data sets from the Palaeoflora Database (Utescher and Mosbrugger 2018). Floral lists with corresponding nearest living relatives (NLRs) employed in this study and their climatic requirements are made available in ESM 1.
In this study, three temperature and four precipitation variables are reconstructed: mean annual temperature (MAT), cold and warm month mean temperature (CMMT, WMMT), mean annual precipitation (MAP), and mean monthly precipitation of the wettest, driest and warmest month (MPwet, MPdry, and MPwarm). Min, max and mean values of climatic parameters for each flora are given in Tables 2 and 3.
In the CA analysis, at least 10 NLR taxa contributing with climate data are required to obtain reliable results (Mosbrugger and Utescher 1997). The climatic resolution of the CA results also depends on the taxonomical level of NLR identification (Mosbrugger and Utescher 1997). Ginkgo biloba L. and Sciadopitys verticillata (Thunb.) Siebold et Zucc. were excluded from the analysis for being monotypic. For the monotypic genera (but not monotypic families) we used climate data for families (for example, climate data of Myricaceae Blume were used for Comptonia peregrina (L.) Coult., Taxodioideae Endl. ex K. Koch – for Glyptostrobus pensilis (Staunton ex D. Don) K. Koch, Sequoia sempervirens (D. Don) Endl. and Taxodium Richard).
Climate seasonality
In order to determine the temperature seasonality of the early Eocene climate of Yakutia, the mean annual range of temperature (MART) was calculated as the difference of WMMT and CMMT for each level (Table 3). To study precipitation seasonality, we calculated the mean annual range of precipitation (MARP – calculated as the difference of MPwet and MPdry) (Table 3). As a further estimate of precipitation seasonality, we calculated the ratio of MPwet and MPdry on MAP (RMPwet and RMPdry) (Table 3), also used as indication of past monsoon intensity (Jacques et al. 2011a). In general, increasing of RMPwet indicates higher proportion of precipitation in the wet season, while the decreasing of RMPdry suggests lower proportion of precipitation in the dry season.
Vegetation reconstruction methods
In order to reconstruct palaeovegetation based on fossil flora, several methods have been developed aiming at standardised, reproducible and thus comparable results, facilitating the analysis of spatio-temporal trends in the evolution of the vegetation cover. These methods differ mainly by the scale of reconstruction from local plant communities (PFT) to biome level (IPR).
Quantitative palaeovegetation reconstruction – Plant functional type approach (PFT)
The Plant Functional Type (PFT) concept goes back to the works of Prentice et al. (1992) and Prentice and Webb III (1998), and has been widely used to describe vegetation cover in vegetation modeling. A PFT is defined using traits and climatic thresholds of key taxa, and combines species related by morphological and phenological traits (François et al. 2011). The present study employs an extended PFT classification scheme comprising 26 herbaceous to arboreal PFTs, based on physiognomic characters and bioclimatic tolerances of plants, and complemented by an aquatic PFT (Popova et al. 2013) (Table 4). The allocation of fossil taxa to the single PFTs is based on the interpretation of their NLRs (Appendix B). Proportions of the different groups of PFTs are given in Table 5.
To exclude unlikely PFTs, we used the likelihood procedure according to François et al. (2011) and Henrot et al. (2017). In cases when several classes of arboreal PFTs are possible for a taxon, only those that can coexist with unambiguously verified classes identified at each site are retained. Here we exclude all PFTs of low and improbable levels of coexistence from the analysis (ESM 2). The PFT approach requires only information on the presence and absence of taxa and thus is comparatively robust towards taphonomic bias.
Plant biome reconstruction – Integrated plant record vegetation analysis (IPR)
The Integrated Plant Record vegetation analysis (IPR) is a semi-quantitative method first introduced by Kovar-Eder and Kvaček (2003) to assess zonal vegetation based on the fossil plant record (leaf, fruit, and pollen assemblages). In order to employ the IPR, thirteen basic taxonomic-physiognomic groups, termed components, defined to reflect key ecological characteristics of an assemblage (Kovar-Eder and Kvaček 2003, 2007; Kovar-Eder et al. 2008; Teodoridis et al. 2011) are used: conifer component (CONIF), broad-leaved deciduous component (BLD), broad-leaved evergreen component (BLE), sclerophyllous component (SCL), legume-like component (LEG), zonal palm component (ZONPALM), arborescent fern component (ARBFERN), dry herbaceous component (D-HERB), mesophytic herbaceous component (M-HERB). Azonal components, i.e. azonal woody component (AZW), azonal non-woody component (AZNW) and aquatic component (AQUA). The component PROBLEMATIC TAXA includes elements with uncertain taxonomic-physiognomic affinity. For further analysis, all taxa (but not their abundances) of every single assemblage have to be assigned to those components and their relative proportions have to be calculated. The complete flora lists, assigned NLRs and their allocation to the components are given in ESM 3. The number of taxa assigned to the components for each flora is given in Table 6.
To characterise zonal vegetation, we regarded the following proportions of components are relevant: (a) the proportion of the BLD, BLE, and SCL+LEG components of zonal woody angiosperms, where “zonal woody angiosperms” means sum of BLD+BLE+SCL+LEG+ZONPALM+ARBFERN components; (b) the proportion of the ZONAL HERB (D-HERB+M-HERB) component of all zonal taxa, where “zonal taxa” means sum of the CONIF+BLD+BLE+SCL+LEG+ZONPALM+ARBFERN+D-HERB+M-HERB components. The reliability of the results increases with an increasing number of zonal taxa preserved. Ten zonal taxa are regarded as a minimum to perform this method (Kovar-Eder et al. 2008). Recently, Kovar-Eder and Teodoridis (2018) raised the former threshold to 15 zonal taxa to apply the IPR-vegetation analysis. The proportions of the components were calculated for each flora and are given in Table 6.
Based on relative proportions of the components the following six zonal vegetation types are distinguished (Kovar-Eder and Kvaček 2007; Kovar-Eder et al. 2008): zonal temperate to warm temperate broadleaved deciduous forests (broadleaved deciduous forests, BLDF), zonal warm temperate to subtropical mixed mesophytic forests (mixed mesophytic forests, MMF), zonal subtropical broadleaved evergreen forests (broadleaved evergreen forests, BLEF), zonal subtropical, subhumid sclerophyllous or microphyllous forests (subhumid sclerophyllous forests, ShSF), zonal xeric open woodlands (open woodland), and zonal xeric grasslands or steppe (xeric grassland). Teodoridis et al. (2011) additionally defined ecotones between the BLDF and MMF and the BLEF and MMF and recently, Kovar-Eder and Teodoridis (2018) defined an ecotone between the MMF/ShSF.
Results
Climate
The analysis of 16 microfloras is based on 31 to 58 (mean 38.9) climate datasets of extant reference taxa (Table 2). In 17 out of 18 cases all NLRs can coexist, in one case – over 98 % of taxa, attributing high significance (Table 2). When reconstructing CA intervals of the MAT obtained from the microfloras, CA ranges are comparatively wide owing to the commonly high taxonomic level of NLR assignment. For MAT the width of CA intervals is ca. 3.6 °C at the mean (varies from 0.2 to 7.3 °C, std. 3.4 °C).
The mean values of MAT show a general cooling trend with some variability between single levels (Tables 2 and 3, Fig. 4). The highest MAT values are indicated for the PF 13 in the lower part – 21.8 °C (21.7–21.9 °C). The lowest MAT values are indicated for some floras in the middle and upper parts – 14.2 °C (13.8–14.5 °C). For the other floras, the MAT mean value is 17.5 °C.

Temperature and precipitation variables: MAT mean annual temperature, CMMT cold month mean temperature, WMMT warm month mean temperature, MAP mean annual precipitation, MPwet mean monthly precipitation of the wettest month, MPdry mean monthly precipitation of the driest month, MPwarm – mean monthly precipitation of the warmest month
The CMMT means generally demonstrate a cooling trend (Tables 2 and 3, Fig. 4). The highest values of CMMT are indicated for the PF 13 in the lower part – 15.2 °C. The lowest CMMT values are obtained for some floras in the middle and upper parts – 5.1 °C (3.1–7.1 °C). For the other floras, the mean values of CMMT vary from 6.6 to 9.4 °C.
WMMT means vary slightly but generally display a cooling trend (Tables 2 and 3, Fig. 4). The highest WMMT values are indicated for the PF 13 in the lower part – 27.7 °C (27.2–28.2 °C). The lowest values of WMMT are obtained for the PF 3 in the upper part – 23.3 °C (20.6–25.9 °C). For the other floras, the mean values of WMMT vary from 23.8 to 25.9 °C.
MAP shows a clear decreasing trend with means varying from 1 100 to 1 400 mm (Tables 2 and 3, Fig. 4). The highest MAP is indicated for the PF 13 – 1 414 mm (1 215–1 613 mm), the lowest for the PF 11 and 12 – 1 126 mm (897–1 355 mm). MPdry means in the early Eocene of northern Yakutia were at ca. 50–65 mm and show a smooth decreasing trend (Tables 2 and 3, Fig. 4). The mean values of MPwet attain very high values (173–276 mm) throughout the early Eocene (Tables 2 and 3, Fig. 4).
Climate seasonality, monsoon intensity
Temperature (MART) and precipitation (MARP) seasonality parameters and the mean values of related climatic parameters for each flora are given in Table 3 in comparison to the present-day values. The MART ranges within 12.4–19.7 °C (mean value of MART is 17.9 °C). The MARP varies from 109 to 205 mm. The RMPwet fluctuated within 15.1–20.7 %, while the RMPdry ranged from 4.1 to 5.1 % (Table 3, Fig. 5).

Lithological profile of the studied section next to curves for various climate and vegetation characteristics
Vegetation
Using the PFT approach, palaeovegetation data of Yakutia were obtained for 16 palynofloras. The number of fossil taxa in each sample is lower than the number of encountered PFTs (Table 5): therefore, our results are meaningful. Aquatic plants (PFT 27) are present in ten palynofloras in lower and middle parts, and the proportion of aquatic plants does not exceed 2.4 % (Table 5, Fig. 6). Herbaceous plants (PFTs 1–3) are present in all floras. The diversity of herbaceous PFTs varies from 4.1 to 9.1 % of the total diversity of the flora (Table 5, Fig. 6). Shrubs (PFTs 4–11) are represented in all floras studied – from 20.2 to 34.5 % (Table 5, Fig. 6). Arboreal plants (PFTs 12–26) are present in all floras studied, attaining 59.5–72.3 % of total diversity (Table 5, Fig. 6). Needleleaved PFTs (PFTs 12–18) are present in all floras, except the lowest flora – PF 16. The proportion of coniferous PFTs varies within the range of 0–16.0 % of total diversity (Table 5, Fig. 6). Broadleaved deciduous plants (PFTs 4–6, 22–25) are present in all floras with diversities varying from 32.4 to 45.7 % (Table 5). Broadleaved evergreens (PFTs 7–9, 19–21, 26) are present in all floras and attain diversities from 28.3 to 50.0 % of the total diversity of the flora (Table 5, Fig. 6).

Ecospectra of PFT diversity and vegetation types next to lithological profile of the studied section and curves of MAT and MAP: MAT mean annual temperature, MAP mean annual precipitation. PFTs are given Table 4
When regarding the diversity spectra of the early Eocene floras analysed (Fig. 6), it is shown that broadleaved summergreen shrubs and trees (PFTs 5, 6, and 24 – up to 22 %) are the most diverse functional types, followed by broadleaved evergreen shrubs and trees (PFTs 8/9 and 19–21) and needleleaved evergreens (PFTs 14–16 and 18). Needleleaved evergreen boreal or temperate cold trees (PFT 12), needleleaved summergreen boreal trees (PFT 17), broadleaved summergreen boreal or temperate cold trees (PFT 22) are completely absent from the spectra after having applied the likelihood procedure. Needleleaved summergreen subtropical swamp trees (PFT 18) are presented in all floras, with the exception of the two lowest floras where conifers are totally absent (Fig. 6). Broadleaved raingreen tropical trees (PFT 25) and broadleaved evergreen tropical trees (PFT 26) also are present in all floras (Fig. 6) but in low proportion. Herbaceous plants in the early Eocene floras of Yakutia are mainly represented by humid herbs (PFT 1).
Sixteen microfloras were analysed using IPR-vegetaion analysis (Table 6). Based on the relative proportions of the components, only one single zonal vegetation type was revealed for the early Eocene of northern Yakutia, namely warm temperate to subtropical mixed mesophytic forest (= mixed mesophytic forest, MMF). The following thresholds characterise floras assigned to this zonal vegetation type: BLD component from 59.0 to 73.8 %, BLE component – 16.9–27.9 %, SCL + LEG component – 6.1–16.2 %, and 9.5–22.0 % of herbs (Table 6, Fig. 7). The PF 6 is the only flora was revealed for ecotone between the BLDF and MMF with BLD component – 75.6 %, BLE component – 16.4 %, SCL + LEG component – 8.1 %, and 19.4 % of herbs (Table 6, Fig. 7).

Quantitative characteristics of the main plant groups according to palynology, PFT, and IPR methods next to lithological profile of the studied section and curves of MAT and MAP: MAT mean annual temperature, MAP mean annual precipitation, NL needleleaved plants, BLD broadleaved deciduous plants, BLE broadleaved evergreen plants, SCL+LEG sclerophyllous and legume-like plants
Discussion
General aspects of the evolution of early Eocene palynomorph complexes in northern Yakutia
According to Kulkova (1971, 1973a, b), the stratigraphic position of the palynological complex of the Tastakhskaya Formation is established by the presence of key taxa characteristic of the Eocene, and, particularly by the abundance of pollen Pistillipollenites mcgregorii Rouse. The characteristic features of the palynological complex of the Tastakhskaya Formation (Table 1, Fig. 3) are the absolute predominance of angiosperm pollen, a large amount of tricolporate and tricolpate pollen, including species determined by morphological and botanical classification (Quercus conferta Boitz. and Quercus graciliformis Boitz., Castanea crenataeformis Samig., Tricolpopollenites liblarensis Th. et Pfl. subsp. fallax R. Pot., T. microhenrici (R. Pot.) subsp. untragranulatus Pfl., Tricolporopollenites cingulum (R. Pot.) Th. et Pfl., etc.), the noticeable participation of pollen of Ulmoideipites And., Hamameidaceae R.Br. (Fothergilla Murr., Hamamelis L., Corylopsis Siebold et Zucc., Liquidambar L.), Myricaceae Blume (Myrica L., Comptonia L’Her.), Juglandaceae A. Richard ex Kunth (Engelhardia quietus (R. Pot.) Elsik, Carya simplex (R. Pot., Venitz.) Elsik, Pterocarya Kunth, Juglans L.), the constant presence of Loranthaceae Juss. (Loranthus elegans I. Kulk. and L. mirus I. Kulk.), Aralia L., Nyssa L., Rhus L., Alnus Mill., Fagus L., Tilia L., Cornus, Corylus L., and the abcense of pollen of Late Cretaceous relics. In a number of floras, pollen of Nypa sp., Sabal sp., Phoenix sp., Palmae gen. Indet., Proteacidites paradoxus I. Kulk., Cardiospermum notabile I. Kulk., Myrtaceidites jacuticus I. Kulk., Casuarinidites cainozoicus Cook. et Pike are found as single grains. Except for Taxodioideae pollen, gymnosperms have minor importance in the spectra. The diversity of spores is low, mainly Polypodiacae gen., Gleichenia sp., and Osmunda sp, and Azolla sp. occur in the upper part.
According to Fradkina (1995, 1996), the palynological assemblage of the Tastakhskaya Formation (second half of the early Eocene – first half of the middle Eocene) is generally similar to the palynological assemblage of the Kengdeiskaya Formation (first half of the early Eocene), but differs in an increasing role of angiosperms attaining up to 90–95 %, including evergreens up to 20–27%, small tricolporate and tricolpate pollen up to 55–59 %, and the appearance and/or more frequent occurrence of typical Eocene taxa.
According to Kulkova (1973a, b), the palynomorph complex of the early – middle Eocene Tastakhskaya Formation is undoubtedly similar to the early Eocene Kengdeiskaya Formation regarding the percentages of main forms (Pistillipollenites mcgregorii, Castanea sp., Rhus sp., Nyssa sp., Engelhardia quietus). However, these forms, combined with a somewhat different general systematic composition of the Kengdeiskaya Formation complex, in which angiosperm pollen and spores alternately dominate, cannot characterise them as time equivalent (Kulkova 1973a, b). The main background in the spectra of the Tastakhskaya Formation is represented by tricolporate and tricolpate pollen, namely Ulmoideipites, and Pistillipollenites Rouse, while in the palynomorph spectra of the Kengdeiskaya Formation – Betulaceae pollen, Quercites sparsus (Mart.) Sam., and Polypodiaceae spores are dominant (Kulkova 1973a, b). When comparing these complexes, attention is drawn to the presence of forms characteristic of each complex separately. So, among the forms not recorded in the complex of the Tastakhskaya Formation, for example, the pollen of Quercites sparsus can be distinguished. According to Boitsova (1966), Quercites sparsus had an extensive range in the Paleocene and was replaced in higher horizons of the Paleogene, in particular in the Eocene, by other characteristic species, such as Q. graciliformis and Q. conferta. In turn, Q. graciliformis and Q. conferta are typical taxa of the complexes of the Tastakhskaya Formation (Kulkova 1973a, b). In addition, the Paleocene and older forms, not found in the spectra of the Tastakhskaya Formation, but noted in the Kengdeiskaya Formation, include Psophosphaera lanceolata Bolkh., Psophosphaera magnoloides Bolkh., Dicksonia densa Bolkh., Podozamites bilateralis Bolkh., Parviprojectus echinaceus Mtch., and Beaupreaidites elegnasiformis Cooks. At the same time, there are morphotaxa, such as Loranthus elegans, L. mirus, Cardiespermum notablle, Tetraporites lubomirovae I. Kulk., Pleurospermaepollenites tastacliensis I. Kulk., and others, only observed in the palynomorph complex of the Tastakhskaya Formation and absent from the spectra of the Kengdeiskaya Formation (Kulkova 1973a, b). Moreover, it is shown that palynomorphs considered by Fradkina (1995) as typical Paleocene elements and Late Cretaceous relicts are absent from the palynomorph complex of the Tastakhskaya Formation. It indicates that the Tastakhskaya Formation complex is younger than the Kengdeiskaya Formation complex.
According to Grinenko et al. (1997) and Fradkina (1995, 1996), the Tastakhskaya Formation is the stratotype of the Tastakhskii regional horizon (the second half of the early Eocene – the first half of the middle Eocene). This horizon includes the Emgendyenskaya stratum and the Dirinskaya Formation of Yakutia, the Anzhuiskaya Formation of the New Siberian Islands, etc. In the stratotype, this horizon (Tastakhskaya Formation) has no geologically fixed upper and lower contacts (Grinenko et al. 1997). However, the temporal position of the section studied in the stratotype of Tastakh Lake is quite definite. Kulkova (1987) established the similarity of the Tastakh palynological complex with that of the marine sediments of Western Siberia, where she revealed a rich complex of dinoflagellates with zonal species. This made it possible to correlate the Tastakhskaya Formation and, accordingly, the regional horizon with the Ypresian – beginning of the Lutetian (Grinenko et al. 1997).
In the palynological complex of the Anzhuiskaya Formation, A.F. Fradkina (Grinenko et al. 1989) found Deflandrea oebisfeldensis Alberti that is part of the dinoflagellate complex of the Upper Lyulinvorskaya subformation of Western Siberia (Kulkova 1987), corresponding to the EECO. In addition, the Eocene Wetzeliella articulate (O. We.) Eisenack is also present here.
The Eocene age of the Tastakhskaya and Anzhuiskaya formations is also confirmed by the carpoflora. In particular, from the deposits of the Tastakhskaya Formation, Dorofeev (1968) identified Taxodium, Sciadopitys, Metasequoia, Glyptostrobus, Pinus, Larix, Picea, Tsuga, Fagus, Tilia, Acer, Trochodenron, Magnolia, Platanus, Betula sect. Costatae, Ficus, Morus, Ulmaceae, Rutospermum, Myrica, Staphylea, Stephanandra, Andromeda, Vaccinium, Aralia, Actinidia, Parthenocissus, Sambucus, Palaeosinomenium, Potamogeton, Butomus, Epipremnella, Nuphar, Menyanthes, Primulaceae, Cucurbitaceae, Azolla, etc. According to Dorofeev (1968), a number of forms in this list are very similar to fruits and seeds from the Eocene deposits of New Siberian Islands, the Irkutsk Basin, and England.
Initially studied by A.N. Krishtofovich (1958), and later L.Yu. Budantsev (1983) based on new collections made by S.A. Laukhin in 1979, the Tastakh leaf flora seemed extremely "exotic" for the north of Siberia due to the predominance of thermophilic large-leaved plants, and the almost complete absence of ferns and conifers. The critically evaluated list of macroremains includes Cephalotaxus sp., Menispermites sp., Trochodendroides arctica (Heer) Berry, Tr. richardsonii (Heer) Krysht., Liquidambar sp., Platanus latifolia (Knowlt.) Krysht., Ulmus pseudobraunii Hollick, Grewiopsis jacutica Krysht., Gr. vollosovitschii Krysht., Acer arcticum Heer, Cocculus eaxinii (Hollick) Budants., Alangium sp., Dombeyopsis splendida Krysht., D. aff. D. alaskana (Newb.) Krysht., Cissus jacutica Krysht., and Cissites vollosovitschii Krysht. Representatives of Liquidambar L., Quercus L., Fagus L. and Alangium Lam. are not typical for Paleocene boreal floras, but are widely present in Eocene and Oligocene floras. A characteristic feature of the Tastakh Lake flora is the large leaf size noted for Platanus, Trochodendroides Berry, Alnus, Acer L., and Alangium. However, many of the Arctic angiosperm leaves are large (Herman 1994), which is an advantageous adaptation to low and predominantly diffuse sunlight situations provided that water is abundant. This feature is also a characteristic of Cretaceous and Paleogene high latitude floras. For example, it is typical for the early early Eocene flora of Kengdei flora (Budantsev 1983; Grinenko et al. 1989). Despite the continuity in their development through common taxa, which include Trochodendroides arctica, Liquidambar, Acer arcticum, and Alangium, Grinenko et al. (1989) pointed the significant differences between the Tastakh flora and the Kengdei flora, and stated that the Tastakh flora is generally more thermophilic and younger than the Kengdei flora.
Evidence for Azolla event?
The end of the early Eocene is marked by the so-called “Azolla event”. A massive Azolla bloom is recorded in the central Arctic Ocean and all Nordic Seas in strata of the latest early Eocene age (49 Ma; van der Burgh et al. 2013; chron 22n-21r; Barke et al. 2012), interpreted as reflecting a massive freshwater discharge into the Arctic Ocean (Brinkhuis et al. 2006; Collinson et al. 2009; Greenwood et al. 2010; Barke et al. 2012) due to elevated precipitations in adjacent drainage areas. The duration of this Azolla bloom is estimated to have been ~800 kyr (Brinkhuis et al. 2006) or 1.2 myr (Barke et al. 2012).
Microspores of the freefloating freshwater fern Azolla are found as single grains in the upper part of the section (PFs 5-8; Table 1). In addition, Azolla was also identified in the carpoflora of the Tastakhskaya Formation (Dorofeev 1968). Of course, the content of Azolla microspores in the late early – early middle Eocene sediments of Tastakh Lake is too low to correlate it with the Azolla event (Fig. 8). However, Salpin et al. (2019) indicate the presence of only single grains of Azolla sp. before the peack (samples 1-22) in the late early Eocene age CaH2 section in the Mackenzie Delta (Arctic Canada). According to Salpin et al. (2019), the highest content of Azolla spores was observed in samples 23 and 34 (77 and 155 grains correspondently).

Proposed stratigraphic correlation of the studied section (green line) based on the pollen assemblages, and the climatic clues of the total set of analyses. PETM Paleocene–Eocene Thermal Maximum; ETM2 – Eocene Thermal Maximum 2; EECO Early Eocene Climatic Optimum; MECO Middle Eocene Climatic Optimum. The Azolla event and hyperthermal event the EECO are indicated in Zachos et al. (2008)
Taking into account the stratigraphic position of the deposits from the late early to early middle Eocene interval (Fig. 8), i.e. from the warm peak correlating with the EECO at the base to the stable cooling at the top of the section correlating with the early to middle Eocene transition ca. 48 Ma, when global temperatures declined, and the fact that the Azolla event is included in this interval, the presence of Azolla in palynofloras and carpofloras suggests the formation of these floras at that time (Fig. 9).

Proposed stratigraphic correlation of the early Eocene sites of northern Yakutia – Tastakh Lake (present study) and Bykovskaya Channel (Bondarenko et al. 2022), curve for various MAT and PFT spectra next to the composite deep-sea benthic foraminiferal oxygen isotope record after Zachos et al. (2008). PETM Paleocene–Eocene Thermal Maximum; ETM2 Eocene Thermal Maximum 2; EECO Early Eocene Climatic Optimum; PFTs are given Table 4
Climate evolution
For all known uncertainties and delimited climatic resolution primarily caused by the application of taxonomical methods in the early Paleogene in general, related uncertainties in NLR identification and their high taxonomical rank, basic conclusions on the early Eocene climate of the high-latitudinal northern part of Yakutia can be drawn. Given the scarcity of continental quantitative palaeoclimate data for the early – middle Eocene in general, even relatively crude estimates provide valuable information. According to our results, a warm temperate climate with MAT of 17.5 °C and CMMT at ca. 8–9 °C in the warmer floras and ca. 14.2 / 5.1 °C in the coolest floras of the record persisted throughout the lithological profile (Fig. 4). However, possibly also nearly tropical climate occurred with MAT of 21.8 °C and CMMT of 15.2 °C in the lower part (PF 13; Fig. 4). Overall, the climate was humid, with a high MAP of over 1 400 mm in the warmest phase and over ca. 1 100 mm during the cooler phases (Fig. 4). The warmest peak obtained for MAT, CMMT and WMMT in the lower part of the Tastakh Lake section (PF 13; Fig. 4), subsequent warming and cooling coinciding with the increasing and subsequent declining trends in d18O values in the global isotope stack throughout the late early – early middle Eocene (Zachos et al. 2008) (Figs. 8 and 9). According to the palynostratigraphical constraints outlined in the previous Section, this warm peak would comprise the EECO global hypothermal event.
These new findings complement results obtained for the first half of the early Eocene of northern Yakutia from a sequence of the Bykovskaya Channel in the area of the Lena River Delta (Bondarenko et al. 2022), illustrating the conditions following the PETM. According to Bondarenko et al. (2022), MAT and CMMT of the warmer floras of the early early Eocene record were at least ca. 16 °C and ca. 10 °C, respectively with the cooler floras at ca. 14.5 °C / 3.5 °C. Thus, a warm temperate, at times nearly tropical climate persisted throughout the deposition of the strata. Also, in the earliest part of the Eocene, the climate was overall humid, with high MAP rates of at least ca. 1 200 mm in the warmer phases and over ca. 1 000 mm during the cooler phases (Bondarenko et al. 2022). The warm peak in the upper part of the Bykovskaya Channel section (PF 3), with MAT at 17.5 °C had been referred to the ETM-2 (Bondarenko et al. 2022; Table 7, Fig. 9).
The comparison of both early Eocene, close located sites in northern Yakutia (Fig. 9), using the recently published and the Tastakh Lake record, allows us to assume that, in the Russian High Arctic, PETM, ETM-2, and the EECO with subsequent cooling can be traced. Both records suggest that, in the first half of the early Eocene, after the PETM, the climate of northern Yakutia was quite equable. In the second half of the early Eocene, after EECO, a period of fluctuations and subsequent cooling trends are observed (Fig. 9).
When considering the palaeolatitude of the studied section of about 73 °N (ODSN Plate Tectonic Reconstruction Service using the hotspot reference frame 1 for 50.0 Ma), CMMT in the study area was warmer than present by around 55 °C (modern climate data from WorldClim), a strikingly high value that is, however, in the order of CMMT anomalies with respect to present-day previously reconstructed from middle Eocene palaeobotanical sites of Eurasia N of 60 ° latitude, including Spitsbergen and Greenland (Utescher et al. 2011). Such high temperatures at very high latitudes are in line with a reconstruction based on early to middle Eocene palynofloras of coastal plain sediments on the Belkovsky and Faddeevsky Islands (New Siberian Islands, Russia) (Suan et al. 2017). In the Faddeevsky Island record, quantitative climate data were obtained with the Climatic Amplitude Method (Fauquette et al. 1998), with most likely values varying between ~16 and 21 °C for MAT, ~6 and 14 °C for CMMT, ~25 and 28 °C for WMMT, and 1 100 and 1 370 mm for MAP, all values in the order of the present reconstruction. Slightly cooler and drier climate conditions were obtained for the ~5-m.y.-younger Belkovsky Island record, where most likely MAT values oscillate between 17 and 19 °C, CMMT between 5.5 and 12 °C, WMMT between 25 and 26 °C, and MAP between 980 and 1340 mm (Suan et al. 2017). The Avicennia pollen records the northernmost mangrove growth ever documented and indicates early Eocene SSTs exceeding 13 °C in winter and 18 °C in summer (Suan et al. 2017).
Salpin et al. (2019), using the Climatic Amplitude Method, reconstructed even higher MAT values between 21.3 and 22 °C and MAP between 1 200 and 1 360 mm for the late early Eocene CaH2 section in the Mackenzie Delta (Arctic Canada). On the Siberian margin, floral assemblages indicate a paratropical humid climate (Akhmetiev and Beniamovskiy 2009; Suan et al. 2017). Also, from lower to middle Eocene strata of Arctic Canada and Greenland high temperatures with MAT (8 and 17 °C) and CMMT (3–9 °C) result from floral and faunal data (Eldrett et al. 2009; Huber and Caballero 2011; Wolfe et al. 2012; West et al. 2015). Floral and faunal assemblages indicative of exceptionally warmer and more humid conditions during the late Paleocene–early Eocene than today have also been reported in other localities around the Arctic Ocean (Ellesmere Island: Eberle et al. 2010; Schubert et al. 2012; Northeast Yukon: Vavrek et al. 2012; Axel Heiberg Island: McIver and Basinger 1999; Northwest Territories (NWT), Ellesmere and Axel Heiberg Islands: Greenwood et al. 2010; Canadian Arctic: Eberle and Greenwood 2012; Alaska: Sunderlin et al. 2011).
Evidence for exceptional warmth in the northern high latitudes during the early Eocene comes from other regions and also from non-botanical proxies. The results from the ACEX expedition for the Lomonosov Ridge indicate euxinic conditions and high primary productivity in the central Arctic Ocean during the Paleogene and early Eocene (Stein et al. 2006). Tetraether lipid-based proxies revealed an increase of air and sea surface temperatures (SST) of up to 8 °C during the PETM (Sluijs et al. 2006, 2009; Weijers et al. 2007) and a subsequent 3–5 °C SST warming during the ETM2 (Sluijs et al. 2009).
Very warm conditions in the early Eocene at high latitudes as evident from our data for the PF 13 are supported by results obtained from other continental and marine proxies. High MAT (~18–25 °C; Weijers et al. 2007 and ~16–21 °C; Suan et al. 2017) and SST (~18–25 °C; Sluijs et al. 2009) values based on tetraether membrane lipids are reported from Arctic lower Eocene strata of IODP Site 302, Lomonosov Ridge (Willard et al. 2019). More details on other paleontological/geological proxies are discussed in Bondarenko et al. (2022).
Temperature and precipitation seasonality
The modern regime of this region is characterised by pronounced temperature seasonality but almost no humidity seasonality. Today, MART is at a very high level of 45.0 °C, at a mean, while MARP is around 18 mm. The cold period (November – March) accounts for approximately 20–25 %, and the warm (April – October) 75–80 % of MAP. The minimum precipitation is observed in most of the territory in February – March. The modern RMPwet and RMPdry calculated based on the mean values using station data of Tiksi (Müller and Hennings 2000; New et al. 2002), are 14.1 % and 7.6 %, respectively (Table 3).
All our climatic data suggest a relatively weak seasonal control of the early – middle Eocene climate of northern Yakutia. The significantly higher-than-present early – middle Eocene WMMTs in combination with an even higher difference in CMMTs indicate a distinctly lower-than-present seasonality of temperature during the early – middle Eocene. Our MART values for the early – middle Eocene range from 18–20 °C, only occasionally dropping below 13 °C (Table 3, Fig. 5). Moreover, seasonal temperature reconstructions show that the warming was primarily a winter phenomenon, with little variability in summer temperatures throughout the profile (Fig. 5). According to Bondarenko et al. (2022), MART values for the first half of the early Eocene of northern Yakutia generally were slightly lower (ca. 15–16 °C).
The pronounced seasonality of precipitation (MPwet 173–276 mm; MPdry as 41–57 mm) in northern Yakutia weakly fluctuated during the early – middle Eocene and did not show any distinct increasing/decreasing trend. The early – middle Eocene MARP was significantly higher-than-present. However, the much higher past MPwet coupled with distinctly higher MPdry indicate that the climate of northern Yakutia was much more humid at that time. The calculated proportions of MPwet and MPdry to yearly precipitation (MPwet: 14.8–19.6 %; MPdry: 3.4–5.8 %) are slightly higher for MPwet and lower for MPdry, compared to present-day (Table 3, Fig. 5) and suggest more pronounced precipitation seasonality of the early – middle Eocene climate of northern Yakutia in comparison to the present-day one.
In general, the ratio of the wet month precipitation to the total amount of annual precipitation (RMPwet) is commonly considered a good proxy for East Asian monsoon intensity (Jacques et al. 2011a). RMPwet values calculated from our record could be interpreted in terms of a monsoonal type of climate. However, recent studies of modern monsoonal sites have shown that leaf physiognomy can be used in the identification of summer-precipitation-dominated monsoonal climates (Jacques et al. 2011b, 2014). This same signal would also be present in ecosystems that would become dormant during the long dark winters of high latitude regions (Royer et al. 2003; Jahren and Sternberg 2008). Leaf area (LA) analysis estimates of MAP may be biased towards precipitation during the Arctic summer, as a result of the winter dormancy. The Arctic summer would have spanned 6–8 months in duration (West et al. 2015). Regions that exhibit monsoons, or “monsoon-type” summer-wet precipitation (MAP >55%; Zhang andWang 2008) are not seasonally equable. Types of monsoonal precipitation regimes have been proposed using both proxies and modeling data to characterise Eocene hyperthermal conditions in several regions of the Earth, including the Arctic and Antarctic (Huber and Goldner 2012; Schubert et al. 2012; Jacques et al. 2014). However, in Polar Regions proposed summer-wet precipitation regimes are contradicted by other proxy evidence (Greenwood 1996; Eberle and Greenwood 2012). The results of West et al. (2015) for estimates of seasonal precipitation from early Eocene megafloras from Ellesmere Island (Arctic Canada) also contradict the monsoonal model, and rather are consistent with the modeling studies of Huber and Goldner (2012), that show Ellesmere Island and the surrounding region of the Arctic as “ever-wet”. Schubert et al. (2012) applied isotopic analysis to fossilised wood from the High Arctic, using δ13C to produce models that showed a high degree of seasonal precipitation: approximately 75% of the MAP falling during the summer polar light season. The dark polar winter as a result would have been comparatively dry. This degree of seasonality, where the ratio of summer precipitation to MAP is greater than 55% (Zhang and Wang 2008), implies a summer monsoonal precipitation regime for the early Eocene Arctic. Nevertheless, LA analysis of the Arctic megafloras has produced MAP estimates that far exceed the growing season precipitation estimates produced by the CLAMP analysis (West et al. 2015) while, for reasons cited above, LA analyses might not provide reliable results for High Arctic flora (see Herman 1994). Thus, at such high latitude, the revealed precipitation seasonality is not necessarily indicative of a monsoonal climate but could be explained by an enhanced hydrological cycle during the Arctic summers. Also, Eldrett et al. (2014) provide evidence for seasonally wetter summers briefly prior to and during the PETM interval in the North Sea and associate these shifts with enhanced hydrological cycling, a mechanism also operational in the Artic during the late Cretaceous that are supposed to cause high precipitation and humidity, under the presence of a permanent polar cloud cap (Spicer and Herman 2010).
Vegetation change
As is shown in Figs. 6 and 7, our late early – early middle Eocene data suggest an impact of climate change and variability on vegetational composition in northern Yakutia. This is mainly evident from changes in the ratio of broadleaved evergreen versus deciduous PFTs that largely co-varies with MAT exemplifying the affinity of evergreens to warmer climates.
Pollen evidence presented here indicates that mixed conifer-broadleaved forests occupied land masses near the studied locality during the late early Eocene. The relatively low percentages of bisaccate pollen (Pinus L., Picea A. Dietr., Abies Mill.) suggests that these components were transported from upland sites; the common occurrence of Juglandaceae and taxodioid pollen and fern spores points to the existence of forested wetlands near rivers and/or the coastline. The occurrence of these assemblages at very high latitudes coincides with other reconstructions for other continental parts of the NH. In the early Paleogene, higher latitudes of North America and Europe were covered by mixed conifer-broadleaved forests (Boulter and Manum 1989; Greenwood and Basinger 1994; Jolley and Whitham 2004; Jolley and Morton 2007; Smith et al. 2007; Jolley et al. 2009; Greenwood et al. 2010; Daly et al. 2011; Kender et al. 2012; Eldrett et al. 2014; West et al. 2015; Suan et al. 2017; Willard et al. 2019). Macrofloral data from Ellesmere Island show that the late Paleocene to early Eocene lowland terrestrial vegetation included coal-forming swamp forests dominated by conifers such as Glyptostrobus Endl. and Metasequoia, whereas upland forests were dominated by broadleaved taxa including Betulaceae Gray, Cercidiphyllaceae Engl./Trochodendraceae Eichler, Juglandaceae, Platanaceae T. Lestib. ex Dumort., Ulmus L., and Tilia L. (Eberle and Greenwood 2012).
Using PFT diversity spectra as proxies for vegetation types (Table 5, Figs. 6 and 7), the early to middle Eocene vegetation of northern Yakutia consisted of the mixed evergreen-deciduous forest. These results are in accordance with our data obtained from the IPR approach, indicating the MMF zonal vegetation type for all studied levels (Table 6, Fig. 7). According to both methods applied, the reconstructed zonal forest biomes have a warm temperate character.
It is known that vegetation changes are often induced by climate change. In many cases, the observed vegetation patterns and their changes through the early – middle Eocene can be correlated with warm temperate climate patterns and the overall continental palaeoclimate evolution as reconstructed from the palaeobotanical record. The change in the proportion of some groups of PFTs (needleleaved vs. broadleaved, broadleaved deciduous vs. broadleaved evergreen) throughout the early Eocene also reflects changes in some climatic parameters (Fig. 7). In general, a gradual decrease in the proportion of broadleaved evergreen PFTs clearly correlates with the cooling trend of MAT, while a gradual increase in the proportion of needleleaved PFTs is in accordance with increasing MART.
The constant presence of PFT 18 in all studied floras throughout the early – middle Eocene suggests a high humidity in general, and agrees with the precipitation reconstruction demonstrating considerably wetter conditions than at present. PFT 18 is also indicative of the existence of swamps at places and intrazonal swamp forest communities and thus points to the presence of inundated areas throughout the studied time-span. The constant presence of humid herbs (PFT 1) also coincides with the above-mentioned high humidity throughout the early Eocene. High MAP rates over ca. 1 000 mm were also obtained for the first half of the early Eocene of northern Yakutia from a sequence of the Bykovskaya Channel in the area of the Lena River Delta (Bondarenko et al. 2022). Nevertheless, high humidity in the Arctic under greenhouse climates is not confined to the Paleogene but is already known from the Late Cretaceous, e.g. reconstructed based on leaf floras from Alaska and NE Russia using CLAMP (Spicer et al. 2019). The humid polar conditions are probably referable to a then enhanced hydrological cycle and formation of clouds in the Arctic that also may explain the distinctly warm polar conditions, due to their radiative forcing effect (Niezgodzki et al. 2017).
The comparison of early Eocene sites in northern Yakutia, i.e. the Bykovskaya Channel section and Tastakh Lake record in the present study (Fig. 9), allows for tracing and discussing the vegetational succession in terms of PFTs together with the climate data. Early Eocene PFT diversity spectra of northern Yakutia show a good correlation with climate data and demonstrate very well that PFT spectra change in relation to palaeoclimate. In general, the colder periods coincide with the appearance of PFTs 13 and 23, while the warmer periods are characterised by an increase in evergreen, deciduous subtropical and tropical PFTs. More evidence for the co-variance of climate and vegetation is obtained from the diversity of tropical PFTs. In the late early to middle Eocene record of Tastakh Lake tropical PFTs (PFT 25 and 26) are present in almost all floras and may attain up to 7.1–10.1 % (Fig. 6). It should be noted that in the first half of the early Eocene of northern Yakutia, the proportion of PFT 26 was lower and did not exceed 4.5 % (Bondarenko et al. 2022). Moreover, IPR vegetation analyses provide a warmer vegetation type (MMF) with higher diversities of broadleaved evergreen PFTs for the late early Eocene compared to the earlier part, consistent with the reconstructed climate data and global climate evolution (Zachos et al. 2008). However, it needs to be stressed that, apart from a few taxa indicative for tropical conditions (e.g. Nypa Steck, see below), most taxa scoring for tropical PFTs may represent temperate types as well (for example, Hamamelidaceae R. Brown, Ilex L., Juglandaceae, Myrica L., Proteaceae Juss., etc.).
The succession of Tastakh Lake is generally characterised by a low abundance of conifers (5.0-23.1 %), except for the PF 16 where conifers are completely absent (Fig. 3). At the same time, there is a clear gradual increase in the proportion of conifers from the base to the top. This coincides with a gradual decrease in MAP and a gradual increase in MART (Fig. 5). Moreover, the Tastakh Lake record is characterised by a low abundance of Taxodioideae (0.8-7.6 %). In contrast, the early early Eocene Bykovskaya Channel record provided a high content of Taxodioideae – up to 53.3% (Bondarenko et al. 2022). At the same time, for the late early Eocene in general slightly higher mean MAP values are noted (see Section 5.3). The lower content of Taxodioideae pollen and sparsity in coals indirectly indicate a possible reduction in swamp vegetation by the end of the early Eocene.
Mangrove is very interesting and very specific vegetation. In our record (the palaeolatitude of ca. 73 °N), mangrove assemblage (including Nypa sp., Phoenis sp., Sabal sp. and Palmae gen. Indet.) occurs as single grains in the lower part only (PF 13; see Fig. 3). This flora is described from viscous, plastic, horizontally laminated clays with an admixture of kaolinite. The low but constant content of taxodioid pollen and fern spores suggests wetland habitats in the lowland. There is indirect evidence of favourable conditions for the growth of Nypa in the early Eocene in the study area. The fine-grained and laminated sediments suggest calm, stable sedimentation conditions, and the coastal-marine genesis of interlayers containing dinocysts indicates a transitional environment between continental and marine. Already described Nypa from the early early Eocene of Bykovskaya Channel (Bondarenko et al. 2022), found under comparable facies settings.
Other mangrove pollen in fossil records of high latitudes – Avicennia L. –appears confidently in the early Eocene in Europe (Chandler 1964; Sein 1961) and New Siberian Islands (Suan et al. 2017; Suc et al. 2020). Mangroves have particular climatic and geographic requirements as well as a distinctive habit of growth (Tomlinson 1986). These features play an essential role in the reconstruction of the palaeoclimate and vegetation patterns. The presence of Nypa fossils may indicate monsoonal or at least seasonal rainfall regimes and likely indicate tropical climates (Pole and Macphail 1996). Suan et al. (2017) for the early Eocene coastal plain sediments of the New Siberian Islands (Russia) estimate climate as subtropical (see Section 5.3). According to Suan et al. (2017), the Faddeevsky Island succession was deposited between the PETM and ETM-2 of the early Eocene, 56–54 Ma, when temperature rose globally before reaching a long-term maximum known as the EECO; sediments at Belkovsky Island were deposited near the early to middle Eocene transition ca. 48 Ma, when the global temperatures declined at the termination of the EECO.
In our records, mangroves are represented only at one floral level and in very low content (by single grains). This may indicate a depressed state or degradation of mangroves when considering that flora PF 13, in which Nypa is recorded, may reflect the vegetation immediately after the EECO, and, therefore a time of gradual degrading and disappearance of mangroves.
Conclusions
The Tastakh Lake record provides the first quantitative climate and vegetation data for the late early to early middle Eocene in the high latitudes of the Far East of Russia enlightening the environmental evolution around the EECO. Our results show a warm temperate and at times, nearly tropical climate in the lower part and a cooling trend in the upper part of the record. The reconstructed Eocene temperatures indicate a strikingly high anomaly with respect to present-day climate, with a CMMT difference of up to 55 °C. Seasonality of temperature was correspondingly low, but generally higher than recently reconstructed for the early early Eocene of nearby strata. Precipitation was at the high level of over 1 400 mm in wetter climate phases and over 1 100 mm in drier phases. Lower seasonality of precipitation in the earlier part of the succession points to more equable conditions near the EECO, thereafter increasing towards the onset of the middle Eocene.
Climate conditions supported a dense forest cover comprising various warm temperate vegetation types and at times even enabled the presence of tropical PFTs. According to IPR vegetation analysis, MMF predominantly existed over the studied time-span, including diverse broadleaved deciduous and evergreen components that thrived under long-lasting polar darkness at a palaeolatitude near 73 °N. Increasing conifers abundance in the later part of our record coincided with a cooling trend in MAT and a gradual increase of temperature seasonality. The low content of Taxodioideae pollen indicates a possible reduction in swamp vegetation during the late early Eocene, compared to neighboring strata dating to the early early Eocene. A floral level containing mangrove elements points to the intermittent existence of almost tropical climate conditions at such high latitude, and can possibly be related to the EECO. Our quantitative climatic and vegetational reconstructions support the existence of early Paleogene warmth in the High Arctic as postulated in reconstructions conducted in other regions of the NH and based on a variety of proxy data.
Data availability
All data generated or analysed during this study are included in this published article and its supplementary information files.
References
Akhmetiev, M. A. (1976). On climatic fluctuations in the Paleogene and Neogene in the middle and high latitudes of the Earth (according to paleobotanical data). International Geological Congress, 25th session (Sydney, 1976), reports of Soviet geologists: paleontology, marine geology (pp. 138–146). Academy of Sciences of the USSR, Ministry of Geology of the USSR. Moscow: Nauka. [in Russian]
Akhmetiev, M. A. (1993). Phytostratigraphy of Paleogene and Miocene continental deposits of boreal Asia. Transactions of the geological Institute of the Russian Academy of sciences 475. Moscow: Nauka. [in Russian]
Akhmetiev, M. A. (2015). High-latitude regions of Siberia and Northeast Russia in the Paleogene: Stratigraphy, flora, climate, coal accumulation. Stratigraphy and Geological Correlation, 23(4), 421–435.
Akhmetiev, M. A., Krasilova, I. N., Naidin, D. P., & Nemkov, G. I. (1981). Geochronological scale and principles of its construction. Phanerozoic. Itogi nauki i tekhniki: Seriya Stratigrafiya, Paleontologiya, 2, 1–121. [in Russian]
Akhmetiev, M. A., Zaporozhets, N. I., Iakonleva, A. I., Aleksandrova, G. A., Benyamovskiy, V. N., Oreshkina, T. V., Gnibidenko, Z. N., & Dolya, Z. A. (2010). Comparative analysis of marine Paleogene sections and biota from West Siberia and the Arctic region. Stratigraphy and Geological Correlation, 18(6), 635–659.
Akhmetiev, M. A., Zaporozhets, N. I., Benyamovskiy, V. N., Aleksandrova, G. A., Iakonleva, A. I., & Oreshkina, T. V. (2012). The Paleogene history of the Western Siberian seaway – A connection of the Peri-Tethys to the Arctic Ocean. Austrian Journal of Earth Sciences, 105(1), 50–67.
Akhmetiev, V. A., & Beniamovskiy, V. N. (2009). Paleogene floral assemblages around epicontinental seas and straits in northern Central Eurasia: Proxies for climatic and paleogeographic evolution. Geologica Acta, 7(1-2), 297–309.
Aleksandrova, G. N. (2016). Geological development of Chauna depression (northeastern Russia) in Paleogene and Neogene, 1: Paleogene. Bulletin of Moscow Society of Naturalists, Geological Series, 91(4-5), 148–164. [in Russian]
Amiot, R., Lecuyer, C., Beuffetaut, E., Frederic, F., Legendre, S., & Meartineau, F. (2004). Latitudinal temperature gradient during the Cretaceous upper-Campanian – Middle Maastrichtian: δ18O record of continental vertebrates. Earth and Planetary Science Letters, 226(1-2), 255–272.
Bakulina, N. T., & Zharikova, L. P. (1989). New data on the palynostratigraphy of the Cenozoic deposits of the Takhstakh trough of the Primorskaya lowland (Yakutia). In A. S. Zykina (Ed.), Cenozoic of Siberia and the northeast of the USSR. Trudy IGiG Sibirskogo otdeleniya AN SSSR 668 (pp. 115–120). Novosibirsk: Nauka, Siberian branch, [in Russian]
Baranova, Yu. P., & Biske, S. F. (1979). Paleogene and Neogene paleoclimates of Northeast Asia. In N. A. Schilo, Yu. P. Baranova (Eds.), Continental tertiary deposits of North-East Asia (stratigraphy, correlation, paleoclimates) (pp. 186–204). Novosibirsk: Publishing house “Nauka”, Siberian branch. [in Russian]
Baranova, Yu. P., Biske, S. F., Goncharov, V. F., Kulkova, I. A., & Titkov, A. S. (1968). Kainozoi Severo-Vostoka SSSR. Trudy Instituta geologii i geofiziki 38. Moscow: Nauka. [in Russian]
Barke, J., van der Burgh, J., van Konijnenburg-van Cittert, J. H., Collinson, M. E., Pearce, M. A., Bujak, J., Heilmann-Clausen, C., Speelman, E. N., van Kempen, M. M. L., Reichart, G. J., Lotter, A. F., & Brinkhuis, H. (2012). Coeval Eocene blooms of the freshwater fern Azolla in and around Arctic and Nordic seas. Palaeogeography, Palaeoclimatology, Palaeoecology, 337, 108–119.
Beerling, D. J., & Royer, D. L. (2011). Convergent Cenozoic CO2 history. Nature Geoscience, 4, 418–420.
Belaya, B. V., & Litvinenko, I. S. (1989). Structure and age of Cenozoic sediments in the Khatakhan - Yarovaya interfluve (Anui depression). In V. I. Volobueva (Ed.), Continental Paleogene and Neogene of the northeast of the USSR. Issue 3 (pp. 18–32). Chukotka. Magadan: Izdatel’stvo SVKNII DVO AN SSSR. [in Russian]
Bogolepov, K. V. (1962). Explanatory note to the map of Mesozoic and Cenozoic tectonics of Siberia and the FarEast, Scale 1 : 5 000 000. Moscow: Nauka. [in Russian]
Boitsova, E. P. (1966). Fossil pollen of the Quercus genus and its significance for the dissection of Paleogene deposits of the West Siberian lowland and Western Kazakhstan. In N. A. Bolchovitina, E. D. Zaklinskaya, A. A. Luber, L. A. Kuprianova, M. W. Oschurkova, I. M. Pokrovskaya, G. M. Romanovskaya, S. R. Samoilovich (Eds.), To the method of paleopalinological research. Leningrad: Izdatel’stvo VSEGEI, (pp. 236–249). Materials to the II International Palynological Conference, Holland, September 1966. [in Russian]
Bondarenko, O. V., Blokhina, N. I., & Utescher, T. (2019). Major plant biome changes in the Primorye region (Far East of Russia) during the Paleogene. Botanica Pacifica, 8(1), 3–18.
Bondarenko, O. V., Blokhina, N. I., Mosbrugger, V., & Utescher, T. (2020a). Paleogene climate dynamics in the Primorye region, Far East of Russia, based on a coexistence approach analysis of palaeobotanical data. Palaeobiodiversity and Palaeoenvironments, 100(1), 5–31.
Bondarenko, O. V., Utescher, T., Blokhina, N. I., Evstigneeva, T. A., & Kezina, T. V. (2020b). Temporal climate and vegetation gradient of the Paleocene in the Amur region (Far East of Russia). Botanica Pacifica, 9(2), 13–35.
Bondarenko, O. V., Blokhina, N. I., Henrot, A.-J., & Utescher, T. (2021). Paleogene vegetation changes in Primorye, Far East of Russia – A study based on diversity of plant functional types. Geological Journal, 56, 650–672.
Bondarenko, O. V., Blokhina, N. I., Evstigneeva, T. A., & Utescher, T. (2022). Short-term climate and vegetation dynamics in Delta Lena river (northern Yakutia, eastern Siberia) during the early Eocene. Palaeoworld. https://doi.org/10.1016/j.palwor.2021.09.006.
Boulter, M. C., & Manum, S. B. (1989). The Brito-Arctic igneous province flora around the Paleocene/Eocene boundary. In O. Eldholm, J. Thiede, E. Taylor, et al. (Eds.), Proceedings of the ocean Drill program, scientific results 104 (pp. 663–680). College Station, TX (ocean drilling program).
Brinkhuis, H., Schouten, S., Collinson, M. E., Sluijs, A., Damsté, J. S. S., Dickens, G. R., & Moran, K. (2006). Episodic fresh surface waters in the Eocene Arctic Ocean. Nature, 441(7093), 606–609.
Bruch, A. A., Utescher, T., & Mosbrugger, V. (2011). Precipitation patterns in the Miocene of Central Europe and the development of continentality. Palaeogeography, Palaeoclimatology, Palaeoecology, 304, 202–211.
Budantsev, L. Y. (1983). History of the Arctic flora of the early Kainophytic era. Moscow: Nauka. [in Russian]
Budantsev, L.Yu. (1997). Late Eocene flora of Western Kamchatka (Trudy BIN RAN, 19). Sankt-Peterburg: Izdatel'stvo “petro-RIF”. [in Russian]
Budantsev, L. Y. (1999). The reconstruction of the Cenozoic climates in eastern-North Asia based on palaeobotanical data. Botanicheskii Zhurnal, 84(10), 36–45 [in Russian]
Burgh, J. van der, Collinson, M. E., van Konijnenburg-van Cittert, J. H., Barke, J., & Brinkhuis, H. (2013). The freshwater fern Azolla (Azollaceae) from Eocene Arctic and Nordic Sea sediments: New species and their stratigraphic distribution. Review of Palaeobotany and Palynology, 194, 50–68.
Chandler, M. E. J. (1964). The lower tertiary floras of southern England IV. London: British Museum (Natural History).
Collinson, M. E., Barke, J., Burgh, J. van der, & van Konijnenburg-van Cittert, J. H. (2009). A new species of the freshwater fern Azolla (Azollaceae) from the Eocene Arctic Ocean. Review of Palaeobotany and Palynology, 155(1), 1–14.
Daly, R. J., Jolley, D. W., & Spicer, R. A. (2011). The role of angiosperms in Palaeocene arctic ecosystems: A palynological study from the Alaskan north slope. Palaeogeography, Palaeoclimatology, Palaeoecology, 309, 374–382.
Decisions of the Interdepartmental Meeting on the Development of Unified Schemes for the Northeast of the USSR (1959). Moscow: Gosgeoltekhizdat. [in Russian]
Dorofeev, P. I. (1968). Paleocarpological data on the history of Tertiary floras in the north of eastern Siberia. In Cenozoic history of the Polar Basin and its influence on the development of landscapes of the northern territories (materials of the symposium) (pp. 58-61): Leningrad: Izdatel'stvo geograficheskogo obshchestva SSSR. [in Russian]
Eberle, J. J., & Greenwood, D. R. (2012). Life at the top of the greenhouse Eocene world – A review of the Eocene flora and vertebrate fauna from Canada's high Arctic. Geological Society of America Bulletin, 124(1/2), 3–23.
Eberle, J. J., Fricke, H. C., Humphrey, J. D., Hackett, L., Newbrey, M. G., & Hutchison, J. H. (2010). Seasonal variability in Arctic temperatures during early Eocene time. Earth and Planetary Science Letters, 296, 481–486.
Eldrett, J. S., Greenwood, D. R., Harding, I. C., & Huber, M. (2009). Increased seasonality through the Eocene to Oligocene transition in northern high latitudes. Nature, 459, 969–973.
Eldrett, J. S., Greenwood, D. R., Polling, M., Brinkhuis, H., & Sluijs, A. (2014). A seasonality trigger for carbon injection at the Paleocene – Eocene thermal maximum. Climate of the Past, 10, 759–769.
Fauquette, S., Guiot, J., & Suc, J.-P. (1998). A method for climatic reconstruction of the Mediterranean Pliocene using pollen data. Palaeogeography, Palaeoclimatology, Palaeoecology, 144, 183–201.
Fradkina, A. F. (1995). Palynostratigrapy of Paleogene and Neogene sediments of northeastern Russia (transactions of the UIGGM SB RAS 806). Novosibirsk: Nauchno-izdatel’skii tsentr OIGGiM SO RAN. [in Russian]
Fradkina, A. F. (1996). Palynological assemblages and stratigraphy of the Paleogene and Neogene in Northeast Russia. Geologiya i Geofizika, 37(7), 39–46. [in Russian]
Fradkina, A. F., Trufanov, G. V., & Vakulenko, A. S. (1979). Eocene deposits of New Siberian Islands. In N.A. Schilo, Yu. P. Baranova (Eds.), Continental tertiary deposits of Northeast Asia (stratigraphy, correlation, paleoclimates) (pp. 22–30). Novosibirsk: Publishing house “Nauka”, Siberian branch. [in Russian]
François, L., Utescher, T., Favre, E., Henrot, A.-J., Warnant, P., Micheels, A., Erdei, B., Suc, J.-P., Cheddadi, R., & Mosbrugger, V. (2011). Modelling late Miocene vegetation in Europe: Results of the CARAIB model and comparison with palaeovegetation data. Palaeogeography, Palaeoclimatology, Palaeoecology, 304, 359–378.
Goncharov, V. F., & Kulkova, I. A. (1970). About the Eocene age of the Tastakh formation in the Yana-Indigirskaya lowland. Geologia i geofizika, 11. [in Russian]
Greenwood, D. R. (1996). Eocene monsoon forests in Central Australia? Australian Systematic Botany, 9, 95–112.
Greenwood, D. R., & Basinger, J. F. (1994). The paleoecology of high-latitude Eocene swamp forests from Axel Heiberg Island, Canadian high Arctic. Review of Palaeobotany and Palynology, 81, 83–97.
Greenwood, D. R., Basinger, J. F., & Smith, R. Y. (2010). How wet was the Arctic Eocene rain forest? Estimates of precipitation from Paleogene Arctic macrofloras. Geology, 38, 15–18.
Grinenko, O. V. (1968). Some paleogeographic features of the east Siberian coast of the Polar Basin in the Paleogene. In Cenozoic history of the Polar Basin and its influence on the development of landscapes of the northern territories (materials of the symposium). Leningrad: Izdatel'stvo geograficheskogo obshchestva SSSR. [in Russian]
Grineniko, O. V., & Kiseleva, A. V. (1971). On the age of coal-bearing deposits of the Bykovskaya Channel in the Lena River Delta. In A. I. Tomskaya & V. F. Vozin (Eds.), Palynological characteristics of the Paleozoic, Mesozoic and Cenozoic deposits of Yakutia (pp. 75–87). Yakutsk: Yakutskoye knizhnoye izdatel'stvo. [in Russian]
Grinenko, O. V., Zharikova, L. P., Fradkina, A. F., et al. (1989). Paleogene and Neogene of the northeastern USSR. Yakut Scientific Center SB AS USSR. [in Russian]
Grinenko, O. V., Sergeenko, A. I., & Belolubskiy, I. N. (1997). Stratigraphy of Paleogene and Neogene deposits of the northeast of Russia. Otechestvennaya geologiya, 8, 14–20. [in Russian]
Gusev, A. I. (1954). On the question of the genesis of fossil ice. Trudy NIIGA, 43(3). [in Russian]
Henrot, A.-J., Utescher, T., Erdei, B., Dury, V., Hamon, N., Ramstein, G., Krapp, M., Herold, N., Goldner, A., Favre, E., Munhoven, G., & Francois, L. (2017). Middle Miocene climate and vegetation models and their validation with proxy data. Palaeogeography, Palaeoclimatology, Palaeoecology, 467, 95–119.
Herman, A. B. (1994). Late Cretaceous Arctic platanoids and high latitude climate. In M. C. Boulter & H. C. Fisher (Eds.), Cenozoic plants and climates of the Arctic (pp. 151–159). Springer (NATO ASI subseries).
Herman, A. B., & Spicer, R. A. (1996). Palaeobotanical evidence for a warm Cretaceouc Arctic Ocean. Nature, 380, 330–333.
Herman, A. B., & Spicer, R. A. (1997). New quantitative palaeclimate data for the Late Cretaceous Arctic: Evidence for a warm polar ocean. Palaeogeography, Palaeoclimatology, Palaeoecology, 128, 227–251.
Herman, A. B., Golovneva, L. B., Shczepetov, S. V., & Grabovsky, A. A. (2016). The Late Cretaceous Arman flora of Magadan oblast, northeastern Russia. Stratigraphy and Geological Correlation, 24, 651–760.
Huber, M., & Caballero, R. (2011). The early Eocene equable climate problem revisited. Climate of the Past, 7, 603–633.
Huber, M., & Goldner, A. (2012). Eocene monsoons. Journal of Asian Earth Sciences, 44, 3–23.
Intergovernmental Panel on Climate Change (IPCC). (Ed.), (2014). Synthesis report. Contribution of working groups I, II and III to the fifth assessment report of the intergovernmental panel on climate change. Geneva: IPCC.
Ivanov, O. A. (1968). The main stages of the development of the northern plains of the northeast of the USSR in the Cenozoic. In Cenozoic history of the Polar Basin and its influence on the development of landscapes of the northern territories (materials of the symposium). Leningrad: Izdatel'stvo geograficheskogo obshchestva SSSR. [in Russian]
Ivanov, O. A. (1969). Cenozoic deposits of the Yano-Indigirskaya lowland and the New Siberian Islands (in connection with the prospective assessment of loose tin content). PhD. Thesis. Leningrad. [in Russian]
Jacques, F. M. B., Gou, S. X., Su, T., Xing, Y. W., Huang, Y. J., Li, Y. S., Ferguson, D. K., & Zhou, Z. (2011a). Quantitative reconstruction of the Late Miocene monsoon climates of Southwest China: A case study of the Lincang flora from Yunnan Province. Palaeogeography, Palaeoclimatology, Palaeoecology, 304, 318–327.
Jacques, F. M. B., Su, T., Spicer, R. A., Xing, Y. W., Huang, Y. J., Wang, W., & Zhou, Z. (2011b). Leaf physiognomy and climate: Are monsoon systems different? Global and Planetary Change, 76, 56–62.
Jacques, F. M. B., Shi, G., Li, H., & Wang, W. (2014). An early–middle Eocene Antarctic summer monsoon: Evidence of ‘fossil climates’. Gondwana Research, 25, 1422–1428.
Jahren, A. H., & Sternberg, L. S. (2008). Annual patterns within tree rings of the Arctic middle Eocene (ca. 45 ma): Isotopic signatures of precipitation, relative humidity, and deciduousness. Geology, 36, 99–102.
Jolley, D. W., & Morton, A. C. (2007). Understanding basin sedimentary provenance: Evidence from allied phytogeographic and heavy mineral analysis of the Palaeocene of the NE Atlantic. Journal of the Geological Society, 164, 553–563.
Jolley, D. W., & Whitham, A. G. (2004). A stratigraphical and palaeoenvironmental analysis of the sub-basaltic Palaeogene sediments of East Greenland. Petroleun Geosciences, 10, 53–60.
Jolley, D. W., Bell, B. R., Williamson, I. T., & Prince, I. (2009). Syn-eruption vegetation dynamics, paleosurfaces and structural controls on lava field vegetation: An example from the Palaeogene Staffa formation, Mull lava field, Scotland. Review of Palaeobotany and Palynology, 153, 19–33.
Kajalainen, V. I. (1967). Stratigraphy of the lower Cenozoic of the Yano-Indigirskaya (Primorskaya) lowland. Uchebnye zapiski NIIGA, regional'naya geologiya 10.
Kajalainen, V. I. (1970). The main questions of the stratigraphy and paleogeography of the Cenozoic of the Yano-Indigirskaya (Primorskaya) lowland. PhD. Thesis. Leningrad. [in Russian]
Kender, S., Stephenson, M. H., Riding, J. B., Leng, M. J., Knox, R. W. O., Peck, V. L., Kendrick, C. P., Ellis, M. A., Vane, C. H., & Jamieson, R. (2012). Marine and terrestrial environmental changes in NW Europe preceding carbon release at the Paleocene–Eocene transition. Earth and Planetary Science Letters, 353-354, 108–120.
Kovar-Eder, J., & Kvaček, Z. (2003). Towards vegetation mapping based on the fossil plant record. Acta Universitatis Carolinae, Geologica, 46(4), 7–13.
Kovar-Eder, J., & Kvaček, Z. (2007). The integrated plant record (IPR) to reconstruct Neogene vegetation: The IPR-vegetation analysis. Acta Palaeobotanica, 47(2), 391–418.
Kovar-Eder, J., & Teodoridis, V. (2018). The middle Miocene central European plant record revisited widespread subhumid sclerophyllous forests indicated. Fossil Imprint, 74(1-2), 115–134.
Kovar-Eder, J., Jechorek, H., Kvaček, Z., & Parashiv, V. (2008). The integrated plant record: An essential tool for reconstructing Neogene zonal vegetation in Europe. Palaios, 23, 97–111.
Krishtofovich, A. N. (1958). Fossil flora of the Penzhinskaya Bay, Lake Tastakh and Khr. Raritkin (Trudy BIN AN SSSR, Seria 8, vyp. 3). Moscow – Leningrad: Nauka. [in Russia]
Kulkova, I. A. (1968). Pollen Pistillipolite mcgregory Rousse in the Eocene deposits of Yakutia. Doklady Akademii Nauk SSSR, 182(6). [in Russian]
Kulkova, I. A. (1971). Eocene flora of the Yano-Indigirskaya lowland and its comparison with the floras of the same age in the northern hemisphere. In V. N. Saks & V. S. Volkova (Eds.), Cenozoic floras of Siberia according to palynological data (pp. 7–21). Moscow: Nauka. [in Russian]
Kulkova, I. A. (1973a). Palynological characterization of Eocene deposits of the Yana-Indigirka lowland. In E. D. Zaklinskaya (Ed.), The palynology of cenophytic (reports of the III international palynological conference) (pp. 105–109). Moscow: Nauka. [in Russian]
Kulkova, I. A. (1973b). Palynological research of Eocene sediments of Yano-Indigir lowland. Moscow: Nauka. [in Russian]
Kulkova, I. A. (1987). Stratigraphy of Paleogene deposits of Western Siberia according to palynological data. Geologiya i Geofizika, 6, 11–17. [in Russian]
Kulkova, I. A., & Laukhin, S. A. (1975). Flora of the continental Paleogene of the Yenisei ridge according to palynological data. Moscow: Nauka. [in Russian]
Laukhin, S. A., & Fradkina, A. F. (1984). Comparative characteristics of the pessimums of the Eocene-Oligocene and Oligocene-Miocene boundaries in Northeast Asia. Doklady Akademii Nauk SSSR, 278(6), 1439–1442. [in Russian]
Laukhin, S. A., Akhmetiev, M. A., Fradkina, A. F., & Zyryanov, E. V. (1988). Palynological characteristics of the Kunga Paleogene in the north of Yakutia. Doklady Akademii Nauk SSSR, 299(3), 686–689. [in Russian]
Laukhin, S. A., Grinenko, O. V., & Fradkina, A. F. (1992). The evolution of the Cenozoic climate of the Arctic coast of Northeast Asia. Izvestiya Akademii nauk SSSR, Seriya Geologicheskaya, 1, 131–139. [in Russian]
Lee, S. (2014). A theory for polar amplification from a general circulation perspective. Asia-Pacific Journal of the Atmospheric Sciences, 50, 31–43.
Lobanov, M. F. (1959). Tertiary continental coal bearing deposits of the northern Verkhoyansk and the New Siberian Islands. In Trudy Mezhvedomstvennogo sovetshaniya po razrabotke unifitsirovannykh stratigraficheskikh skhem Severo-Vostoka SSSR (pp. 406-411). Magadan: Magadanskoe knizhnoe izdatel’stvo. [in Russian]
L'vova, E. M. (1989). Results of spore-pollen analysis of sediments of the upper Cenozoic of the middle reaches of the river. Alazii (Yakutia). In A. S. Zykina (Ed.), Cenozoic of Siberia and the northeast of the USSR (pp. 120–124). Moscow: Nauka. [in Russian]
McIver, E. E., & Basinger, J. F. (1999). Early Tertiary floral evolution in the Canadian high Arctic. Annals of the Missouri Botanical Garden, 86, 523–545.
Mosbrugger, V., & Utescher, T. (1997). The coexistence approach – A method for quantitative reconstructions of tertiary terrestrial palaeoclimate data using plant fossils. Paleogeography, Palaeo-climatology, Palaeoecology, 134, 61–86.
Müller, M. J., & Hennings, D. (2000). The global climate data atlas on CD rom. University Flensburg, Institute für Geografie.
Narkhinova, V. E. (1989). Palynology of Cenozoic deposits of the Ust'-Pegtymel'skaya depression. In V. I. Volobueva, (Ed.), Continental Paleogene and Neogene of the northeast of the USSR (pp. 33–49). Issue 3. Chukotka. Magadan: Izdatel’stvo SVKNII DVO AN SSSR. [in Russian]
New, M., Lister, D., Hulme, M., & Makin, I. (2002). A high-resolution data set of surface climate over global land areas. Climate Research, 21, 1–25.
Niezgodzki, I., Knorr, G., Lohmann, G., Tyszka, J., & Markwick, P. J. (2017). Late Cretaceous climate simulations withdifferent CO2 levels and subarcticgateway configurations: A model-data comparison. Paleoceanography, 32, 980–998.
Nikitin, V. P. (2006). Paleocarpology and stratigraphy of the Paleogene and Neogene of Asian Russia. Novosibirsk: Izdatel’stvo “GEO”. [in Russian]
Pearson, P. N., & Palmer, M. R. (2000). Atmospheric carbon dioxide concentrations over the past 60 million years. Nature, 406, 695–699.
Pogozhev, A. G., & Semeykin, A. I. (1959). Tertiary deposits of the northeast of the USSR. Proceedings of the Interdepartmental Meeting on the Development of Unified Stratinographic Schemes for the Northeast of the USSR. Magadan. [in Russian]
Pole, M. S., & Macphail, M. K. (1996). Eocene Nypa from Regatta Point, Tasmania. Review of Palaeobotany and Palynology, 92, 55–67.
Popova, S., Utescher, T., Gromyko, D. V., Mosbrugger, V., Herzog, E., & François, L. (2013). Vegetation change in Siberia and the northeast of Russia during the Cenozoic cooling: A study based on diversity of plant functional types. PALAIOS, 28, 418–432.
Prentice, C., & Webb III, C. (1998). Biome 6000, reconstructing global mid-Holocene vegetation patterns from palaeoecological records. Journal of Biogeography, 25, 997–1005.
Prentice, C. I., Cramer, W., Harrison, S. P., Leemans, R., Monserud, R. A., & Solomon, A. M. (1992). A global biome model based on plant physiology and dominance, soil properties and climate. Journal of Biogeography, 19, 117–134.
Royer, D. L., Osborne, C. P., & Beerling, D. J. (2003). Carbon lose by deciduous trees in a CO2-rich ancient polar environment. Nature, 424, 60–62.
Salpin, M., Schnyder, J., Baudin, F., Suan, G., Suc, J.-P., Popescu, S.-P., Fauquette, S., Reinhardt, L., Schmitz, M.D., & Labrousse, L. (2019), Evidence for subtropical warmth in the Canadian Arctic (Beaufort-Mackenzie, Northwest Territories, Canada) during the early Eocene. In K. Piepjohn., J. V. Strauss, L. Reinhardt, & W. C. McClelland (Eds.), Circum-Arctic Structural Events: Tectonic Evolution of the Arctic Margins and Trans-Arctic Links with Adjacent Orogens. Geological Society of America Special Paper 541. https://doi.org/10.1130/2018.2541(27
Schubert, B. A., Jahren, A. H., Eberle, J. J., Sternberg, L. S. L., & Eberth, D. A. (2012). A summer time rainy season in the Arctic forests of the Eocene. Geology, 40(6), 523–526.
Sein, M. K. (1961). Fossil spores of the London Clay. Unpublished PhD Thesis. London: University of London.
Sluijs, A., Schouten, S., Pagani, M., Woltering, M., Brinkhuis, H., Damsté, J. S. S., & Moran, K. (2006). Subtropical Arctic Ocean temperatures during the Palaeocene/Eocene thermal maximum. Nature, 441, 610–613.
Sluijs, A., Schouten, S., Donders, T. H., Schoon, P. L., Röhl, U., Reichart, G.-J., Sangiorgi, F., Kim, J.-H., Sinninghe Damsté, J. S., & Brinkhuis, H. (2009). Warm and wet conditions in the Arctic region during Eocene thermal maximum 2. Nature Geoscience, 2, 777–780.
Smith, F., Wing, S., & Freeman, K. (2007). Magnitude of the carbon isotope excursion at the Paleocene–Eocene thermal maximum: The role of plant community change. Earth and Planetary Science Letters, 262, 50–65.
Spicer, R. A., & Corfield, R. M. (1992). A review of terrestrial and marine climates in the Cretaceous and implications for modeling the greenhouse earth. Geological Magazine, 129, 168–180.
Spicer, R. A., & Herman, A. B. (2010). The Late Cretaceous invironment of the Arctic: A quantitative reassessment using plant fossils. Palaeogeography, Palaeoclimatology, Palaeoecology, 295, 423–442.
Spicer, R. A., & Parrish, J. T. (1986). Paleobotanical evidence for cool north polar climates in Middle Cretaceous (Albian-Cenomanian) time. Geology, 14, 703–706.
Spicer, R. A., Herman, A. B., Yang, J., & Spicer, T. E. V. (2014). Why future climate change is likely to be underestimated: Evidence from palaeobotany. Journal of the Botanical Society of Bengal, 67, 75–88.
Spicer, R., Valdes, P., Hughes, A., Yang, J., Spicer, T., Herman, A., & Farnsworth, A. (2019). New insights into the thermal regime and hydrodynamics of the early Late Cretaceous Arctic. Geological Magazine, 157, 1729–1746.
Stein, R., Boucsein, B., & Meyer, H. (2006). Anoxia and high primary production in the Paleogene Central Arctic Ocean: First detailed records from Lomonosov Ridge. Geophysical Research Letters 33, https://doi.org/10.1029/2006GL026776.
Suan, G., Popescu, S.-M., Suc, J.-P., Schnyder, J., Fauquette, S., Baudin, F., Yoon, D., Piepjohn, K., Sobolev, N. N., & Labrousse, L. (2017). Subtropical climate conditions and mangrove growth in Arctic Siberia during the early Eocene. Geology, 45, 539–542.
Suc, J. P., Fauquette, S., Popescu, S. M., & Robin, C. (2020). Subtropical mangrove and evergreen forest reveal Paleogene terrestrial climate and physiography at the north Pole. Palaeogeography, Palaeoclimatology, Palaeoecology, 551, 109755. https://doi.org/10.1016/j.palaeo.2020.109755.
Sunderlin, D., Loope, G., Parker, N. E., & Williams, C. J. (2011). Paleoclimatic and paleoecological implications of the Paleocene–Eocene fossil leaf assemblage, Chickaloon formation, Alaska. Palaios, 26, 335–345.
Teodoridis, V., Kovar-Eder, J., Marek, P., Kvaček, Z., & Mazouch, P. (2011). The integrated plant record vegetation analysis: Internet platform and online application. Acta Musei Nationalis Pragae, Seria B, 67(3-4), 159–165.
Tomlinson, P. B. (1986). The botany of mangoves. Cambridge University Press.
Tsherbakov, I. P. (1975). Forest cover of the northeast of the USSR. Nauka, Siberian Branch. [in Russian]
Utescher, T., & Mosbrugger, V. (2018). The Palaeoflora Database. Available at http://www.palaeoflora.de. Last accessed 08 August 2020.
Utescher, T., Erdei, B., François, L., & Mosbrugger, V. (2007). Tree diversity in the Miocene forests of Western Eurasia. Palaeogeography, Palaeoclimatology, Palaeoecology, 253, 242–266.
Utescher, T., Bruch, A. A., Micheels, A., Mosbrugger, V., & Popova, S. (2011). Cenozoic climate gradients in Eurasia – A palaeo-perspective on future climate change? Palaeogeography, Palaeoclimatology, Palaeoecology, 304, 351–358.
Utescher, T., Bruch, A.A., Erdei, B., François, L., Ivanov, D., Jacques, F.M.B., Kern, A.K., Liu, C. (Y.S.), Mosbrugger, V., & Spicer, R.A. (2014). The coexistence approach – Theoretical background and practical considerations of using plant fossils for climate quantification. Palaeogeography, Palaeoclimatology, Palaeoecology, 410, 58–73.
Vakulenko, A. S. (1968). Palynological characteristics of the lower Cenozoic sediments of the lower reaches of the Indigirka River. Uchebnye zapiski NIIGA, paleontologiya i biostratigrafiya 21.
Vavrek, M. J., Evans, D. C., Braman, D. R., Campione, N. E., & Zazula, G. D. (2012). A Paleogene fl ora from the upper bonnet plume formation of Northeast Yukon territory, Canada. Canadian Journal of Earth Sciences, 49(3), 547–558.
Volkova, V. S., Kulkova, I. A., & Fradkina, A. F. (1990). Klimat Eotsena Sibiri (na primere Zapadnoi Sibiri) (Preprint NQ 4). Novosibirsk: Izdatel’stvo IGiG SO AN SSSR. [in Russian]
Volobueva, V. I., Gladenkov, Y. B., & Belaya, B. V. (1988). Paleogene of the northeastern USSR. In V. P. Pokhialainen & M. K. Gagiev (Eds.), Stratigrafiya i paleontologiya fanerozoya Severo-Vostoka SSSR (pp. 118–156). Magadan: SVKNII DVO AN SSSR. [in Russian]
Vollosovich, K. A. (1930). Geological observations in the tundra between the lower reaches of the Lena and Kolyma rivers. In Trudy komissii po izucheniyu YAASSR 13.
Weijers, J. W. H., Schouten, S., Sluijs, A., Brinkhuis, A., & Sinninghe Damsté, J. S. (2007). Warm arctic continents during the Palaeocene – Eocene thermal maximum. Earth and Planetary Science Letters, 261, 230–238.
West, C. K., Greenwood, D. R., & Basinger, J. F. (2015). Was the Arctic Eocene ‘rainforest’ monsoonal? Estimates of seasonal precipitation from early Eocene megafloras from Ellesmere Island, Nunavut. Earth and Planetary Science Letters, 427, 18–30.
Westerhold, T., Marwan, N., Drury, A. J., Liebrand, D., Agnini, C., Anagnostou, E., Barnet, J. S. K., Bohaty, S. M., De Vleeschouwer, D., Florindo, F., Frederichs, T., Hodell, D. A., Holbourn, A., Kroon, D., Lauretano, V., Littler, K., Lourens, L. J., Lyle, M. W., Pälike, H., et al. (2020). An astronomically dated record of Earth's climate and its predictability over the last 66 million years. Science, 369, 1383–1387.
Willard, D. A., Donders, T. H., Reichgelt, T., Greenwood, D. R., Sangiorgi, F., Peterse, F., Nierop, K. G. J., Frieling, J., Schouten, S., & Sluijs, A. (2019). Arctic vegetation, temperature, and hydrology during early Eocene transient global warming events. Global and Planetary Change, 178, 139–152.
Wolfe, A. P., Csank, A. Z., Reyes, A. V., McKellar, R. C., Tappert, R., & Muehlenbachs, K. (2012). Pristine early Eocene wood buried deeply in kimberlite from northern Canada. PLoS One, 7, e45537. https://doi.org/10.1371/journal.pone.0045537.
Zachos, J. C., Dickens, G. R., & Zeebe, R. E. (2008). An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature, 451, 279–283.
Zhang, S., & Wang, B. (2008). Global summer monsoon rainy seasons. International Journal of Climatology, 28, 1563–1578.
Acknowledgements
The authors are thankful to Prof. Dr. Funda Akgün and Prof. Dr. Christopher Yusheng Liu for carefully revising the manuscript and for their valuable suggestions. The research was carried out within the state assignment of Ministry of Science and Higher Education of the Russian Federation (theme No. 121031500274-4). This work contributes to NECLIME (Neogene Climate Evolution in Eurasia).
Funding
Open Access funding enabled and organised by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bondarenko, O.V., Utescher, T. Late early to early middle Eocene climate and vegetation change at Tastakh Lake (northern Yakutia, eastern Siberia). Palaeobio Palaeoenv 103, 277–301 (2023). https://doi.org/10.1007/s12549-022-00530-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-022-00530-6