
Abstract
The provenance and depositional setting of the human remains in the Dietersberg Cave, located in the Franconian Alb in Southern Germany, are evaluated based on 14C-dates and stable isotope analyses (C, N, O, Sr). Four basic scenarios are discussed: (1) human sacrifice, (2) ‘regular’ burial place for a small social unit, (3) special social group (e.g. slaves) and (4) special circumstances of death (e.g. fatal illness). Scenarios 1 and 2 are unlikely as the age distribution includes all ages and both sexes and the 14C-dates of the human remains span most of the Iron Age which would result in an implausible small burial community. Stable isotope analyses also render the deposition of slaves (scenario 3) implausible because a high proportion of the individuals were probably of local origin and their diet was not fundamentally different from that of contemporary populations. The archaeological evidence points to a social bias (i.e. low social standing) as reason for deposition. However, the high numbers of apotropaic objects and of perinatals also suggest that scenario 4 might be plausible for at least some of the individuals. The cave was probably a place of deposition not only for one category of individuals but also for those whose burial in the ‘regular’ cemetery was not considered appropriate.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
It has been argued that about 10% of the approximately 3000 caves of the Franconian Alb contain archaeological finds and that of these, 100 probably also contain human bones (Graf et al. 2008). For decades, these human skeletal remains were considered to reflect rites of sacrifice and even cannibalism (e.g. Torbrügge 1979, 62; Pauli and Glowatzki 1979, 147ff.; Ettel 2002).
However, the reassessment of the Iron Age finds from the Dietersberg Cave near Egloffstein/Franconian Jura by Norbert Baum in 1999 (Baum 1999), together with the publications of Peter-Röcher (1997, 1999, 2003), mark a paradigm-change in the interpretation of human remains from these caves. Baum found no evidence for either sacrificial or anthropophagic rituals on the human bones. He concluded based upon the fairly normal age distribution of the 46 individuals (see below) that the cave had been used as a ‘regular’ burial ground. He further speculated that perhaps a family of beekeepers had buried their dead in this cave for a few generations because of the bad condition of their teeth (Baum 1999, 101).
In order to assess the competing hypotheses outlined below and find evidence for possible alternative explanations, the human remains found in the Dietersberg Cave were 14C dated, and their carbon, nitrogen, oxygen, and strontium isotope compositions were analysed in a project “Settlement burials of the Early Iron Age” funded by the German Research Foundation (Deutsche Forschungsgemeinschaft).
A few scenarios have been put forward as plausible explanations for the deposition of human remains in caves like Dietersberg: (1) For a long time, the predominant paradigm was that humans were thrown into the cave as a kind of sacrifice. (2) The alternative interpretation put forward by Baum assumed that the cave served as a ‘regular’ burial ground for a small social unit. (3) Another possibility could be that the dead in the cave were part of a special social group, for example, individuals with particularly high or low social standing (e.g. slaves). (4) Finally, it seems conceivable that the humans fell victim to special circumstances of death or fatal illness and therefore were placed in the cave as special burial ground reserved for such individuals.
Assuming that Baum was correct in his interpretation of the dead in the Dietersberg Cave as members of a family of beekeepers and therefore considering the cave as a ‘regular’ burial place (scenario no. 2), we can expect – taking the relatively small sample size into accountFootnote 1 – that the depositions happened within a short time interval. Furthermore, if they were really beekeepers, differences in diet compared to contemporaneous groups are expected. On the other hand, if the humans were sacrificed in the cave (scenario no. 1), we might expect – assuming that they were slaves and/or prisoners of war – a provenance from different regions. This would be reflected in non-local strontium and possibly oxygen isotope signatures (if isotopically distinct water sources were used) of their bones and teeth. This would probably also hold for scenario no. 3. Only for scenario no. 4, isotope analyses a priori would seem of limited help. In this study, we combine 14C dating of bones with stable isotope analysis of teeth from the Dietersberg humans to assess their burial history, provenance and diet and to determine which of these different scenarios is most plausible.
The Dietersberg Cave: geology, stratigraphy and physical anthropology
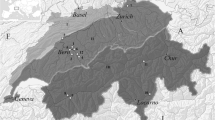
The Dietersberg Cave was discovered in the 1920s and excavated in 1928 by the local speleologist Josef Richard Erl.Footnote 2 The cave is situated on the western edge of the Franconian Alb at about 425 m above sea level near Egloffstein/Ldkr. Forchheim (cf. Fig. 1). This Dietersberg Cave developed due to karst processes of the Upper Jurassic marine limestones (‘Malm Delta’) and has a cave geometry typical of the Franconian Alb, consisting of a very narrow cleave with a depth of about 11 m. The cave entrance measures only 1.0 to 1.5 m so entering (and excavating) the cave is very difficult and would have been so in the past. It can therefore be assumed that items deliberately deposited in the cave were not meant to be recovered.

The Dietersberg Cave near Egloffstein and its geologic surrounding (simplified) (source: Geologische Karte der Bundesrepublik Deutschland 1:1.000.000, https://services.bgr.de/wms/geologie/gk1000/)
Archaeological finds were concentrated on a limestone debris cone, which marked the lower boundary of the deposits. Above the bone-bearing layer was another sediment layer, which yielded no archaeological material. This stratigraphic succession seems to indicate a continuing depositional practice within a relatively short time interval. The debris cone consisted of marly sediment in which many smaller and larger fragments of the limestone bedrock were embedded; in this substrate, bones and other archaeological objects occur without any apparent order. Only two bronze rings (see below) were found in situ still encircling arm bones.
Today, a minimum number of 46 individuals have been identified based on the mandible, which is the most common diagnostic bone present in the assemblage, because the human remains underwent heavy selection processes during and after excavation. Baum (1999, 88ff.), however, argues convincingly that complete bodies were originally deposited in the cave.
Apart from the human bones, only a few other items were found: 12 earrings, seven arm rings, 13 eye beads, four cowrie snails, one iron lance head, and one ceramic vessel. These finds are unambiguously dated to the Early Iron Age (periods HaD to LtB, c. 620–250 cal BC; cf. Baum, 1999, 88).
Of the 38 individuals sampled for the present study, only seven were sexed by N. Baum.Footnote 3 This is not surprising as he had only the mandibulae and in a few cases where a calvarium could be matched to a certain mandibula, the whole skull. Four individuals were classified as female and three as male. Age-at-death estimation was performed using X-ray images of the mandibulae (Baum 1999, 93). Based on this age distribution, Baum argued that the assemblage resembled that of a ‘regular’ cemetery (ibid. 105). However, many newborns are conspicuous, particularly when contrasted to the very low proportion observed in ‘regular’ cemeteries (see further below). Several non-metric, morphological traits of the skeletal remains were taken by Baum as indicators of family relationships within the individuals deposited in the cave in general.Footnote 4 As mentioned above, Baum put special emphasis on the bad condition of the teeth, many were lost intra vitam or when still present showed signs of caries (frequency of caries among the adults: 25.7%Footnote 5); this observation led him to suggest that these people had perhaps consumed large amounts of cariogenic substances such as honey.
In Table 1, the anthropological information of specimens analysed in this study is provided (based on Baum 1999).
Material and methods
A representative sample of 13 bones was radiocarbon dated in the Friedrich-Alexander-University of Erlangen-Nuremberg AMS 14C-laboratory. Ten mandibulae were sampled for 14C dating, while another three long bones (right femora) had already previously been 14C-dated (see Table 1 for details). Therefore, it cannot be ruled out that one or more individuals have been sampled twice.Footnote 6 The 14C-results were analysed with OxCal 4.2.4 (Bronk Ramsey 2009) to obtain calibrated 14C-ages and a probability density plot with the lower and upper boundaries of the age distribution (Fig. 2).

As far as possible, all individuals identified by Baum were sampled for stable isotope analyses (see Table 1). In addition, four animal bones from the cave (mandibulae belonging to a fox, pig and cow and the limb bone of a cat), registered in the Natural-Historical Museum Nuremberg, were sampled in order to obtain a local baseline value for 87Sr/86Sr.
Nitrogen and carbon isotopes are frequently used to reconstruct the diet of archaeological humans. Carbon isotopes (13C/12C expressed as δ13C value) permit the quantification of C4 plant consumption such as maize or millet (see review by Ben-David and Flaherty 2012) and are also used to detect the contribution of food from marine resources to diet (e.g. Sealy 2001). The consumption of both types of food sources (C4 and marine) results in higher δ13C values than consumption of resources from terrestrial C3 plant-based food webs. In contrast, freshwater fish consumption leads to variable, primarily low δ13C values compared to consumers of terrestrial food sources (Peterson and Fry 1987).
While δ13Ccollagen reflects the carbon isotope composition of the ingested protein, the δ13Ccarbonate reflects the carbon isotope composition of the bulk diet with a specific apatite–diet spacing (∆δ13Capatite–diet) (Ambrose and Norr 1993). This offset ∆δ13Ccarbonate-collagen between structural carbonate and collagen therefore indicates different amounts of ingested protein versus lipids/carbohydrates. This offset can vary due to differences in digestive physiology and diet but is used to distinguish consumption of animal and plant matter, i.e. herbivory from carnivory (Codron et al. 2018 and references therein), but also to assess the degree of protein intake from C3 and C4 plant-based resources in omnivorous humans (Ambrose et al. 2003; France and Owsley 2015; Goude et al. 2017). Since the region was dominated by C3 plants that time, ∆δ13Ccarbonate-collagen in the skeletons from the Dietersberg Cave should be largely a measure for the proportion of animal-derived food in the daily diet. Edible plants provide much less fat than animal products, and since fat is depleted in 13C, ∆δ13Ccarbonate-collagen is significantly lower in carnivores compared to herbivores. This way, a negative relationship between δ15N and ∆δ13Ccarbonate-collagen is expected.
Nitrogen isotopes (15N/14N expressed as δ15N value) increase systematically with each trophic level in a food chain (e.g. Caut et al. 2008). The δ15N values of bone collagen are proxies not only for the consumption of animal proteins, i.e. the amount of meat (e.g. Styring et al. 2010), but also for (mother’s) milk (Fogel et al. 1989; Kornexl et al. 1997; Fuller et al. 2003). Therefore, δ15N can also be used as a proxy to determine the age of weaning (e.g. Fuller et al. 2003, 2006a, 2006b).
In a Europe-wide study on stable isotopes of honey, Schellenberg et al. (2010, 772 Table 1) also included samples from Franconia. The samples (n = 40) showed a mean (± 1SD) δ13C value of − 26.2 ± 0.5‰ V-PDB and a mean δ15N value of 2.6 ± 1.6‰ AIR, respectively. The proportion of honey-derived products in the human diet required to result any significant isotopic effect can only be hypothesized. Based on isotopic values of modern honey, honey-consuming individuals would probably have δ13C values similar to individuals eating other C3 plant-derived food.
For C and N isotope analysis of collagen, a piece of each corpus mandibulae (minimum 250 mg) was taken. The collagen extraction followed a standard protocol (e.g. Becker and Grupe 2012). As a precaution, individuals with collagen, yielding abnormally low or high N (≤ 5.31 or ≥ 19.87 wt%) or C concentrations (≤ 13.64 or ≥ 53.0 wt%) or C/N ratios (≤ 2.85 or ≥ 3.65), were excluded. Only 28 individuals from the Dietersberg Cave yielded results in the acceptable parameter ranges. C and N isotope composition of the well-preserved collagen samples were measured using a Flash EA 2000 elemental analyser connected online to a Thermo Finnigan Delta V Plus mass spectrometer at the GeoZentrum Nordbayern (Prof. Dr. M. Joachimski). δ13C and δ15N values were determined with a precision of 0.1‰ and 0.2‰, respectively.
Oxygen isotopes (18O/16O expressed as δ18O value) of both the carbonate and the phosphate fraction of the bioapatite are related to meteoric water and are predominantly controlled by drinking water (Longinelli 1984; Daux et al. 2008). Thus, the use of isotopically distinct water resources from different water bodies, altitudes or latitudes can potentially be detected (e.g. White et al. 2004). δ18O values can vary by up to 1.5‰ within a human population as the result of metabolic processes (Lightfoot and O’Connell 2016). In addition, δ18O values from the structural carbonate fraction, especially in bone, are prone to diagenetic alteration. Considering these caveats, interpretation of inter-individual differences in δ18O should be regarded with caution.
The integrity of the bone apatite can only be assessed by the application of mineralogical methods. As a conservative measure, the δ18Oc values of those samples, which were excluded due to deviant collagen C and N concentrations and molar C/N ratios, were also not considered for interpretation, because of potential diagenetic changes. Thus, only for 28 individuals, reliable δ18Oc values are available. The δ18Oc values of the structurally bound carbonate in the bone bioapatite were measured after deproteinization of c. 100 mg bone powder (cf. Becker and Grupe 2012) using the classical phosphoric acid extraction using 100% H3PO4 in a GasBench II connected to a ThermoFisher Delta V Plus mass spectrometer at the GeoZentrum Nordbayern (Prof. Dr. M. Joachimski). The δ13C and δ18Oc values of the carbonate were measured with a precision of 0.1‰ and 0.15‰, respectively.
Strontium isotopes (87Sr/86Sr) are related to bioavailable Sr, which reflects the lithology and geological age of the bedrock substrate on which the food is ingested (e.g. Beard and Johnson 2000; Price et al. 2002; Bentley 2006; Maurer et al. 2012). Therefore, combined Sr and O isotope analyses can be used to trace the provenance and mobility of individuals (e.g. Chenery et al. 2010; Eckardt et al. 2009; Evans et al. 2006; Tütken et al. 2008a, b; Tütken 2010).
Only teeth, and whenever possible molars, were sampled for strontium isotope analyses (see Table 1 for details). All except one of the 34 individuals (Baum No. 6) (excluding the four perinatals) were measured successfully.
Sample preparation and strontium isotope analysis was performed using published protocols (Becker and Grupe 2012). In brief, the teeth were heated to 300 °C, and enamel was separated from dentin by hand-picking. The enamel fragments were then leached with suprapure 1 M acetic acid to remove any adhering diagenetic Sr. The cleaned enamel was then ashed at 550 °C and powdered in an agate mortar. The enamel powder was digested in distilled 12 M HNO3, the strontium was separated by ion chromatography using Sr-Spec (50–100 mesh) resin. The Sr fraction of each sample was diluted in 2% HNO3 to a 100 ppb solution to measure the 87Sr/86Sr with a ThermoFisher Neptune MC-ICP-MS at the University of Bonn. All data were normalized to a 87Sr/86Sr of 0.71025 for the NIST SRM 987 standard on a daily basis which was measured after each third sample. Some of the 87Sr/86Sr were measured with a Finnigan MAT 262 thermion mass spectrometer at the University of Munich. External reproducibility of 87Sr/86Sr for both laboratories was better than 0.000016.
Results
Radiocarbon ages
The 14C-dates (n = 13) of the human bones are spread relatively evenly across the Early Iron Age (see Table 1). A combined calibration of all data from the Dietersberg Cave generated using the “Combine” function in OxCal results in a chi-squared value of 63.1, which is much higher than the critical value of 21 for the 5%-probability level with 12 degrees of freedom (Suppl. 2). Therefore, it is extremely unlikely that the individuals (i.e. bones) were deposited in the cave in one single event. It is far more likely, as is implied by the boundaries of the modelled data (Fig. 2; Suppl. 3), that the deposition process continued for ~ 500 years. Deposition appears to have begun shortly before 800 cal BC and ended before 300 cal BC. This time span even exceeds the one reflected by the archaeological finds, which date to the archaeological periods HaD to LtB (see above), roughly equivalent to 620 to 250 cal BC (see, e.g. Hald 2009, 44ff. for a recent review of the absolute chronology of the Central European Iron Age).
Strontium isotopes
The three animal bones exhibit a wide range of values of 87Sr/86Sr of 0.71120 (fox), 0.71139 (pig) and 0.70947 (cow), respectively. The human individuals from the Dietersberg Cave also show a high variability (Fig. 3) with a mean value of 0.71037 ± 0.00119, ranging from 0.70823 to 0.71296 (n = 33). Most 87Sr/86Sr values cluster around 0.709, although also some values cluster at 0.710, 0.711 and 0.712. Only five values have radiogenic 87Sr/86Sr values > 0.712: infant no. 3 (87Sr/86Sr = 0.71201), adult male no. 8 (87Sr/86Sr = 0.71206), two adults nos. 18 and 19 (87Sr/86Sr = 0.71252; 0.71296) and juvenile no. 21 (87Sr/86Sr = 0.71202). The adult female individual no. 7 has the lowest 87Sr/86Sr of 0.70823. Nevertheless, the variability of 87Sr/86Sr measured in the teeth far exceeds the isotopic variability typical for individuals belonging to a local population (± 0.00015, Burton and Price 2013).

Enamel 87Sr/86Sr values of the teeth (n = 33) from human individuals and faunal bones (n = 3) from the Dietersberg Cave. The range of the individuals from the ‘regular’ cemetery of Dietfurt and the expected range for Jurassic Bedrock are plotted for comparison. For the individual values, see Table 1
Adult individuals cover a slightly larger range of values than non-adult ones. While the latter measure between 0.70869 and 0.71201 (n = 16), the former display a range from 0.70823 to 0.71296. Of the seven individuals sexed by Baum, only six were analysed for 87Sr/86Sr, including four female individuals and two of the three males. While it is obvious that no statistically meaningful results are to be expected from such a small sample size, it is still interesting that the two males show higher 87Sr/86Sr than any of the female individuals.
There is no correlation between the 14C-dates and 87Sr/86Sr (not shown), indicating that the spatial origin of the individuals deposited in the cave probably remained more or less the same over the time.
Stable oxygen isotopes
The oxygen isotope composition of the structural carbonate (δ18Oc) in all bones varies between −6.9 and −9.2‰ (n = 28) with a mean value of −8.0‰. Most of the δ18Oc values cluster between −8.0 and −7.3‰ (Fig. 4): however, in terms of age, interesting patterns emerge (Fig. 5).

87Sr/86Sr and δ18O values (n = 23) of the individuals of the Dietersberg Cave, differentiated according to age at death and sex (for 3 adult individuals). For the individual values, see Table 1. The ellipse shows the extent of the values of the individuals from the regular cemetery of Dietfurt (n = 7)

δ18O values (n = 28) of the individuals of the Dietersberg Cave, differentiated according to age at death. For the individual values, see Table 1
With two exceptions, only adults exhibit values < −8.3‰. The two exceptions are a perinatal individual (perinatal A) and a very young child aged 0.5–years (no. 38), both of which may still have been in isotopic equilibrium with their mothers (Britton et al. 2015). Apart from these, non-adult individuals (n = 15) show a very clear maximum between −7.7 and −8.0‰. In contrast, for the adults (n = 13), a bimodal distribution can be observed with higher values between −7.0 and −8.0‰ corresponding roughly to the youngest age class, while lower values between −8.7 and −9.5‰ are restricted to individuals older than 20 years. As there are too few sexed individuals who meet the quality criteria, it cannot be determined whether or not sex played a role in this differentiation.
Furthermore, we can neither falsify nor support Baum’s hypotheses concerning the family relationship of the individuals deposited in the cave. In terms of the ossiculae suturae lambdoideae for the two individuals sampled without these additional bones, the information is inconclusive: viewed from the 87Sr/86Sr perspective, they might have lived on the same bedrock substrate and thus potentially could have shared a common place of origin (no. 5: 0.71136; no. 10: 0.71125; the δ18O value for no. 10 had to be excluded). The individuals with these additional bones (n = 9) exhibit even higher variability of 87Sr/86Sr and δ18O. The three individuals with a persisting frontal suture (Sutura metopica) obviously came from different locations (no. 1: 87Sr/86Sr = 0.71001, δ18O = −9.0‰; no. 7: 87Sr/86Sr = 0.70823, δ18O excluded; no. 21: 87Sr/86Sr = 0.71202, δ18O = −7.3‰).
Stable nitrogen and carbon isotopes
The variability of the δ13C and δ15N values of the collagen (n = 29) is considerable. δ13C values vary between −21.8 and −16.8‰, with a mean value of −20.3 ± 1.0‰, while δ15N values range between 7.5 and 11.4‰, with a mean value of 8.9 ± 0.9‰. The δ13C values of bone carbonate (n = 28) vary between −13.8 and −5.0‰ with a mean value of −11.4‰. The spacing of the carbonate and collagen δ13C values (∆δ13Ccarbonate-collagen) is highly variable accordingly, and ∆δ13C values cover a large range between 4.3 and 14.9‰ (Table 1) with a mean value of 8.8 ± 1.9‰.
There does not seem to be a correlation between δ13C and δ15N values and 87Sr/86Sr values. Noteworthy, however, is the distribution according to age (Fig. 6). Children below an age of 3 years have δ15N values > 10.5‰, while children > 3 years have δ15N values similar to adults. On the other hand, the youngest individuals, the perinatals, largely fit into the group of adults. There is one outlier with a δ13C value of −17.5‰ (perinatal D), close to an adult individual at −16.8‰ δ13C (no. 23).

Collagen δ15N and δ13C values (n = 28) of the individuals of the Dietersberg Cave, differentiated according to age at death. For the individual values, see Table 1
Interestingly, the group of adults with δ18O values < −8.3‰ (n = 6) also have similar δ15N compositions which vary only slightly (between 8.5 and 9.3‰). In contrast, δ15N of the other adults with higher δ18O values (n = 7) range between 7.5 and 9.6‰ (Fig. 7).

δ15N and δ18O values (n = 28) of the individuals of the Dietersberg Cave, differentiated according to age at death and their sex (if determined). For the individual values, see Table 1
In terms of ∆δ13Ccarbonate-collagen, differences between the age classes show up too, and there is an almost perfect separation between adult and non-adult individuals. Only adults exhibit values below 8.5‰, with the exception of one adult (no. 30), while all non-adults have values of 8.5‰ or above (Fig. 8).

δ15N and ∆δ13Ccarbonate-collagen values (n = 28) of the individuals of the Dietersberg Cave, differentiated according to age at death and their sex (if determined). For the individual values, see Table 1
Discussion
The isotopic data from Dietersberg Cave were compared with a contemporary ‘regular’ burial place Dietfurt a. d. Altmühl ‘Tennisplatz’, located about 80 km to the south-southeast of the cave (Röhrig 1994). This is the nearest cemetery of reasonable size with the most complete data set. However, we also refer to other burial places further away when necessary and helpful.
87Sr/86Sr and δ18O baselines
For all four scenarios outlined in the introduction, the geographic origin of the individuals in the Dietersberg Cave bears immediate relevance. Therefore, it is important to establish baselines for locally bioavailable 87Sr/86Sr and δ18O ingested via food and water, respectively, which enable us to infer whether the humans were of local origin.
In provenance studies based on 87Sr/86Sr, the conservative approach of defining the ‘local’ baseline for bioavailable 87Sr/86Sr of a population is to compute the mean value for all individuals ± two standard deviations. This is, however, only appropriate if the 87Sr/86Sr values have a unimodal distribution, but the Dietersberg Cave specimens display a multimodal distribution. In this case, parametrics like arithmetic mean and standard deviation are not applicable.
Other approaches look at the geologic bedrock or try to analyse local faunal, plant or water samples, to characterize the 87Sr/86Sr of the biologically available strontium (e.g. Maurer et al. 2012). However, these approaches also have their own problems. For instance, it is well known that 87Sr/86Sr in a consumer’s tissue is dependent on the bioavailable 87Sr/86Sr entering the biosphere, which can differ considerably from the 87Sr/86Sr of the bedrock substrate (e.g. Maurer et al. 2012). Moreover, 87Sr/86Sr is strongly influenced by diet. As a consequence, variance in consumer 87Sr/86Sr is highly compressed compared to that in rocks: of 4885 human enamel samples from six continents, 95% of the 87Sr/86Sr fell into the narrow range between 0.7047 and 0.7190. The intra-population 87Sr/86Sr difference for humans from a single location is estimated at about 0.0003 (Burton and Price 2013).
The isotopic spacing of the three animal bones (0.00191) is already a magnitude higher than this estimate of a local range and is even considerably exceeded by the isotopic spacing of the human teeth (0.00473) from Dietersberg Cave. This implies a provenance of the individuals from different places or that the bioavailable 87Sr/86Sr in the surrounding area is more variable than usual. The Franconian Jura is dominated by marine limestones of Late Jurassic Age, which form the plateau of the Franconian Alb that extends north-south. The Alb is surrounded by predominantly siliciclastic sedimentary rocks of the Middle and Early Jurassic (Fig. 1). The Late Jurassic “Malm” limestones and marlstones have 87Sr/86Sr between 0.707 and 0.710. In the south, the bedrock substrate of the foothills of the Alps and the Molasse basin is composed of siliciclastic sediments of Tertiary and Cenozoic age. The area in the west is formed by Upper Triassic sedimentary rocks of the Keuper, mostly clay to sandstones, with 87Sr/86Sr between 0.708 and 0.711. Regions with higher (more radiogenic) 87Sr/86Sr > 0.711, like bunter, Palaeozoic granite and metamorphic bedrock of the Bohemian Massif, are found to the east of the Franconian Jura (Fig. 1) (see Stephan 2009 and references therein).
Archaeological studies applying 87Sr/86Sr as provenance tracer on faunal or human remains have so far mostly concentrated on Baden-Wurttemberg and Southern Bavaria. A large strontium isotope database of bedrock, water and human and animal remains has been compiled by E. Stephan (2009; s. a. Bentley et al. 2003; Bentley and Knipper 2005) along with a large faunal sample from the Iron Age. Stephan (2009) determined that modern and archaeological skeletal remains from Jurassic bedrock substrates in Baden-Wurttemberg have 87Sr/86Sr that varies between 0.70834 and 0.70978 (Stephan 2009).
In contrast, very few archaeological 87Sr/86Sr data from the Franconian Jura have been published so far. One Bell Beaker individual (Endneolithic) from Pommelsbrunn, about 30 km southeast from the Dietersberg Cave on similar (Middle and Late Jurassic) bedrock, has been analysed yielding 87Sr/86Sr values of 0.71054 (tooth) and 0.70998 (bone) (Grupe et al. 1997, 521; Price et al. 2004, 26).
Therefore, the three animal bones analysed in this study add considerably to the Sr isotope data available for this region. From the limited isotope data available, one would anticipate 87Sr/86Sr to range between 0.708 and 0.7105 for local animals. However, the values for the fox and pig from Dietersberg Cave exceed this range significantly. While it is conceivable that the animals (or their bones) were brought to the cave from further away, it seems much more likely that the animals were local; this is especially true for the fox (a synanthropic animal). For human remains (teeth) from the contemporaneous Iron Age cemetery Dietfurt, a similar 87Sr/86Sr range from 0.70967 to 0.71102 was determined (n = 13, unpubl.).
Taking into account the geological diversity of the area and the 87Sr/86Sr measured in the animal bones, the ‘local’ bioavailable 87Sr/86Sr for the Dietersberg Cave would range from roughly 0.709 to 0.7115. When we also take the neighbouring ‘tails’ of the human 87Sr/86Sr clusters in this range into account (Fig. 3), the ‘local’ 87Sr/86Sr values range from about 0.70867 to 0.71167. Values below and above this range can be considered as most likely ‘non-local’. This would apply to one and six individuals, respectively. This range of ‘local’ 87Sr/86Sr values is considerable and displays a multimodal distribution. Dietary preferences of the individuals and/or different food catchment areas may have caused this 87Sr/86Sr distribution pattern. Additionally, it could also be related to socially complex society where people had different individual access to certain food components.
Defining a baseline for δ18O is more straightforward. Judging from modern precipitation values, for the Franconian Alb, annual mean δ18OH2O values between −8.8 and −9.2‰ are to be expected (GNIP database n.d.: Hof-Hohensaas [1983–2016] and Regensburg [1978–2016], arithmetic means of monthly values, respectively). It is not certain that these modern meteoric water values prevailed also during prehistoric times, but it is the best assumption that can be made at this time. Whether bone δ18Oc values reflect these meteoric water values depends on the drinking behaviour of the humans. For instance, the type of drinking water source used (rivers, groundwater and lake water), seasonal variation in source water consumption and abnormal weather conditions in the years prior to death can all influence δ18Oc composition. Archaeological samples from pig enamel demonstrate that substantial intra- and inter-annual δ18OH2O variations can occur (Bentley and Knipper 2005, 631 Fig. 2). Nevertheless, δ18Oc values of Iron Age human skeletal remains allow us to isotopically characterize drinking water sources and to assess if different sources were used.
The cemetery of Dietfurt a. d. Altmühl around 80 km to the SSE yields a mean δ18Oc of the individuals of −8.12 ± 0.68‰ (1SD, n = 9; unpubl.). Another contemporaneous cemetery, Werneck-Zeuzleben, is located about 90 km WNW from Dietersberg Cave. Here, the mean δ18Oc of the individuals is −7.86 ± 0.93‰ (1SD, n = 12; unpubl.). The mean value of the individuals from the Dietersberg Cave of −8.0‰ falls right in between these two means and thus is in line with consumption of local drinking water.
Taking the mean ± 2 standard deviations (= 0.61‰, resulting in a range of −6.78 to −9.22‰) for all Dietersberg Cave individuals (n = 28) as the local baseline seems not appropriate given the bimodal distribution of the individuals older than 20 years. The individuals aged 19 or less (n = 15) exhibit a somewhat higher mean of −7.83‰ with a standard deviation of 0.46‰. Adding and subtracting two standard deviations lead to a range of −6.91 to −8.75‰. With this narrower range, the four adult individuals at the lower end of the distribution with values around − 9.0‰ would be classified as non-locals.
Overall, seven individuals out of 33 (= 21.2%) were considered non-local by means of 87Sr/86Sr if the baselines defined above are accepted. Among them are five adults and two non-adults. This implies that these individuals may have spent their early years elsewhere as molar crowns (mostly M1, m2 in the case of the child and P1 in the case of one of the adults [no. 8]) mineralize in early childhood. To this group, another four adults with very low δ18O values can be added. It is likely that they came from the south of the Dietersberg Cave where lower δ18O values occur. Taken altogether, eleven of 38 individuals (= 28.9%) or nine out of 18 adult individuals (= 50.0%) would be considered ‘non-local’, based on these assumptions.
However, one must keep in mind that isotope ratios within the ranges defined above are likely consistent with local origins but may also occur in non-local individuals from localities with similar geological and climatic/hydrologic conditions.
In regular Iron Age cemeteries, 50% non-local adults are common (Müller-Scheeßel et al. 2015). Therefore, the assumed percentage of non-local adults from the Dietersberg Cave seems to reflect the general high mobility of adults in the Iron Age.
Diet
Generally, the collagen δ15N and δ13C values of the humans from the Dietersberg Cave fit into the range of other populations dating to the Iron Age (Fig. 9). While their values certainly lie at the lower end of both ranges, there is no indication that their diet was fundamentally different from that of other contemporary groups. Similar values were recently found in three Iron Age individuals from the ‘Cherry Tree’ Cave situated in the same region (Seregély et al. 2015, 231 Tab. 14). The δ13C values fall mostly in the typical range expected for consumption of food from terrestrial C3-plant ecosystems, and the δ15N values indicate some degree of animal protein (i.e. meat) consumption, albeit perhaps at a low level. The stable isotope data is in line with a mixed, mainly cereal-based diet with additional consumption of some animal products, as typical for Iron Age populations. Thus, no specific diet could be detected which would confirm Baum’s hypothesis that the Dietersberg humans were beekeepers.

Arithmetic means and standard deviations of collagen δ15N and δ13C values (n = 13) of the adult individuals of the Dietersberg Cave in comparison to other Iron Age populations (after Knipper et al. 2017, 1318, Tab. 2)
Nevertheless, the differences in ∆δ13Ccarbonate-collagen between adults and non-adults indicate a variable diet between the different age groups, whereby adults exhibit on average lower carbonate-to-collagen spacings than the non-adults. With the exception of the very young children where δ15N indicates a nursing effect, non-adults with collagen δ15N signatures that fall into the same range as those of the adults have higher ∆δ13Ccarbonate-collagen values, indicative of a lower proportion of fat in the daily diet. Possibly, this energy-rich food component was largely reserved for the adults that had a higher energy use in the course of physical work. The finds from the Dietersberg Cave combine low δ15N values with high ∆δ13Ccarbonate-collagen spacings and are thus compatible with a diet largely based on cereals. A possible influence of C4 plants (see Ambrose et al. 2003), however, cannot be ruled out completely because millet was already a staple food at that time (Baitinger et al. 2010, 305, Fig. 10; Oelze et al. 2012; Knipper and Maus 2016; Knipper et al. 2017; Moghaddam et al. 2016; see also the survey by Lightfoot et al. 2013).

The high δ15N values > 10.5‰ of small children (< 3 years) most likely reflect breastfeeding, while children > 3 years have δ15N values similar to those of adults (Fig. 6) and were obviously already fully weaned. Thus, weaning presumably took place prior to or latest around the age of 3 years. On the other hand, the youngest individuals, the perinatals, are similar to the adults and thus were probably still in isotopic equilibrium with their mothers.
Anthropology
Baum claimed that the age distribution of the individuals is compatible with that of a ‘regular’ cemetery. At first glance, this could be true (Fig. 10) because all age categories (except the senile ones) are represented. However, upon closer inspection, the age distribution differs in some aspects from that of a ‘regular’ cemetery (here Dietfurt is used for comparison; data taken from Röhrig 1994): Such cemeteries frequently document a surprisingly low death rate in children; only for age class 1–5 years does the probability exceed 10%. From adulthood onwards, the rate increases steadily; the ‘dip’ in the mortality rates in individuals aged 40+ − also been observed in other contemporary cemeteries (e.g. Magdalenenberg: Zäuner and Wahl 2013, 144, Tab. 2a, column ‘Sterbewahrscheinlichkeit in %’) and could be due to inherent problems with anthropological methods for the determination of age at death. In contrast, the death rate for the children below an age of 10 years from the Dietersberg Cave is several times higher than for those from Dietfurt. For example, children of the age between 5 and 10 years had a probability to die of 22%, while for the children from Dietfurt, this value was only 8%. For the adult individuals, the probability curve for the Dietersberg Cave is very jagged, which undoubtedly is also due to the overall low numbers of individuals, which makes it hard to compare and interpret the data. Still, it is interesting to note that the adults aged 20 to 30 show a much lower probability of death than those from Dietfurt.
These differences do not mean, of course, that the living conditions were completely different and thus lead to very different mortality patterns. Instead, it can be taken as an indication that the age distribution of the people deposited in Dietersberg Cave differs from that of ‘regular’ cemeteries and might indicate burial of selected individuals.
The high frequency of caries among the adult individuals from the Dietersberg Cave of 25.7% is remarkable when compared with other contemporaneous populations. For Dietfurt, a lower frequency of caries (8.8%) is reported, which is in line with other contemporaneous ‘regular’ cemeteries (see Baum, 1999, 96 f., e.g.). A similar high percentage of one quarter of affected teeth is, however, also found among the individuals from the Durezza Cave in Austria (Fabrizii-Reuer and Reuer 1997, 174 Tab. 21).
Setting aside the possibility that the individuals from the Dietersberg Cave consumed a more cariogenic diet, as proposed by Baum, another explanation could be that the higher caries infection rate reflects an on average higher ontogenetic age of these individuals. This would fit with the slightly later peak in the age distribution described above.
Archaeological finds
The small number of archaeological finds is in accordance with observations in other caves of similar age (especially the Durezza Cave/Austria: Gleirscher 1997). In comparison with contemporary ‘regular’ burials, the absence of fibulae and belt buckles is particularly remarkable and contrasts with so-called offering places where such pieces are regularly found (Müller-Scheeßel 2013). It was suggested that the human bodies were brought to theses caves naked or only covered in shrouds but still adorned with jewellery (Gleirscher 1997, 233). However, the few pieces of jewellery found in the Dietersberg Cave could have adorned only some of the bodies. Some of the bronze objects were worn or reworked (Baum 1999, 100), and none of them could be regarded as ‘high-status’ jewellery as it appears in contemporaneous ‘regular’ burials. Still, the comparatively large number of eye beads and cowrie snails, which are frequently linked to apotropaic practices (Pauli 1975), is peculiar and requires further explanation.
Final consideration of the four burial scenarios for the dead in Dietersberg Cave
In the introduction, four basic scenarios were outlined for the deposition of human remains in the Dietersberg Cave: (1) victims of sacrifices, (2) ‘regular’ burial place (perhaps, of beekeepers), (3) special/deviant social group and (4) special/deviant circumstances of death. In the following, we discuss to what extent the available archaeological, anthropological and isotopic information can help us to evaluate these four competing hypotheses.
From the age distribution of the individuals, it seems obvious that the first scenario – human sacrifice – can be ruled out: It is hard to believe that humans of all ages and both sexes were chosen indiscriminately for ‘sacrifice’ (as already argued by Baum 1999).
On the other hand, the 14C dates indicate that the cave was used as place of deposition for human corpses for nearly 500 years, much longer than Baum originally hypothesized. Even assuming that the 46 individuals identified by Baum represent only the minimum number of individuals deposited, this would not allow for a social group of plausible size.Footnote 7 Thus, the dead in the cave can only represent a small portion of the actual burial community. Furthermore, there is no indication that the diet of the humans from the Dietersberg Cave was fundamentally different from that of other contemporary groups. However, it is not clear whether the diet of beekeepers (with a possibly higher portion of honey) is expected to be isotopically distinct from the typical diet of Iron Age populations. At best, the higher rate of caries in the Dietersberg assemblage could be taken as tentative indication for such a scenario (i.e. a more cariogenic diet). Overall, however, scenario 2 – the cave as a ‘regular’ burial place of a small group of specialized beekeepers – is also not convincing.
That leaves us with the other two options: deviant social group (scenario 3) or deviant mode of death (scenario 4).
Considering the moderate social complexity, which is usually assumed for Iron Age societies (e.g. Burmeister 2000), social deviancy would probably result in the deposition of individuals of especially low or high social positions in the cave. As described above, few archaeological objects were found, some of them even showed signs of wear or reworking, and none of which could be regarded as ‘high-status’ jewellery. This could be an indication that social inferiority might indeed have played a role. In this case, the dead in the cave could have been captives in some sense, being slaves or prisoners of war or both, or their descendants (the non-adults).Footnote 8 What is at odds with the ‘captive’ scenario, however, is the large number of seemingly ‘local’ individuals based on the 87Sr/86Sr and δ18O values: a priori, with slaves as well as prisoners of war, one would expect a much higher proportion of ‘non-locals’, raided in farer-away locations. However, only two non-adults were non-local, and the number of non-locals among the adults (50%) cannot be regarded as exceptional, as was been pointed out earlier. Though it must be conceded that we know virtually nothing of Iron Age slavery, scenario 3 – at least in this extreme form – is therefore not completely convincing either. However, the evidence pointing to low social standing for the individuals in question is still valid. Even if one regards the high rate of caries not as a sign of affiliation to a socially inferior group – high rates of caries are usually more linked to low-status individuals in pre-modern societies (Lanfranco and Eggers 2012, 18) – but instead as an indication of the higher proportions of older individuals in the assemblage, this hypothesis is still in accordance with the general understanding of Iron Age society in which only a small percentage of older adults gained in status (Burmeister and Müller-Scheeßel 2005).
The last alternative, deviant mode of death, is best supported by the unusual large amount of eye beads and cowrie shells discovered in the cave. It seems conceivable that they were given to the individuals in a state of crisis before or even after their death as a kind of amulet and were deposited with the individuals in the cave after death.
In this regard it is worth mentioning the very high number of perinatal individuals found in the cave. While it is entirely possible that they represent stillbirths, it is equally likely that they were placed there not on their own but unborn together with their mothers who had died in childbirth. Ethnographically it is well known that the corpses of women having died during childbirth are handled in a special way (Meyer-Orlac 1982, 75ff.; Shay 1985). Such a scenario seems especially likely for the perinatal and adult individual (of unknown sex) with outstandingly high collagen δ13C values > −18‰ (Fig. 6).
Conclusion
In summary, we state that our observations are partly explained by two of the four scenarios outlined in the introduction. We can exclude two popular hypotheses brought forward to explain the deposition of the dead humans in Dietersberg Cave – human sacrifice and ‘regular’ burial place for a small social group – as well as a non-local provenance of the majority of the deposited individuals. Overall, it seems that there are at least two main motivations for the deposition of the human remains in the cave: Some of the individuals obviously belonged to a lower social stratum of their Iron Age communities, whereas others perhaps suffered a ‘bad’ death such as death during childbirth.
While the distinction between deviant social group and deviant mode of death seems useful to us, it may very well be that in the Iron Age, this differentiation was not made at all. What these two groups have in common is that they were not considered appropriate to be buried in a ‘regular’ cemetery.
In this regard, it is worth emphasizing that the youngest age group, the infans I individuals, are relatively abundant in Dietersberg Cave but are typically under-represented in ‘regular’ burial places, possibly having not yet ‘earned’ their place in the ‘regular’ cemetery. Some of the adults with an isotopically inferred ‘non-local’ provenance might accordingly represent individuals that have died away from home and – because of missing family ties – were not buried ‘properly’ in a regular cemetery. Another reason for a special treatment of dead persons, also known ethnographically, might be dying without descendants. However, these last two cases are hard to prove archaeologically.
The Dietersberg Cave was thus probably not the depositional place for only one category of dead individuals, but rather for those individuals whose burial in the ‘regular’ cemetery was not considered appropriate – for various reasons. Social status, cause of death and (young) age were important aspects, but certainly not the only ones.
Notes
For example, the well-known contemporaneous grave mound ‘Magdalenenberg’ near Villingen-Schwenningen/Baden-Wurttemberg contained 141 individuals (Zäuner and Wahl 2013) which dated into a time-frame of about 50 years.
This and the following information are derived from Baum 1999.
15 calvaria were removed before excavation and later deliberately destroyed (Baum 1999, 84).
Baum 1999, 99; according to Baum three of 19 individuals with a preserved Os frontalis showed a persisting frontal suture (Sutura metopica) while 13 of 16 skull fragments of the Os occipitalis had additional bones in the Sutura lambdoidea (Ossicula suturae lambdoideae). However, in his catalogue only three of 16 individuals for Sutura metopica and eleven of 14 for the ossiculae are explicitly named. It is not clear how this divergence is to be explained.
According to the formula (teeth with caries + closed alveoles) * 100/observable tooth positions, this value differs from the value given by Baum 1999, 96, because we did not include alveoles with pathogenic signs as this might lead to double counting (there are many isolated teeth in the collection which Baum included); furthermore, Baum obviously did not add closed alveoles to the sample size.
Due to the nature of the assemblage, it is not possible to match individual bones to each other without extensive aDNA analyses.
Assuming 50 individuals and a period of deposition of 500 years, a single person would have died only every 10 years. With a theoretical life expectancy of 25 years, this would amount to a population size of only two to three individuals.
The existence of slaves or the habit of taking prisoners is hard to prove archaeologically. From later sources, we know that the inhabitants of Western and Central Europe traded slaves with their Mediterranean neighbours (e.g. Poseidonius: Diodorus 5, 26.3).
References
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate prehistoric human bone: archaeology at the molecular level. In: Lambert JB, Grupe G (eds) . Springer Berlin Heidelberg, Berlin, Heidelberg
Ambrose SH, Buikstra J, Krueger HW (2003) Status and gender differences in diet at Mound 72, Cahokia, revealed by isotopic analysis of bone. J Anthropol Archaeol 22:217–226
Baitinger H, Hansen L, Kalis AJ, Kreuz A, Pare CFE, Schäfer E, Schatz K, Stobbe A (2010) Der Glauberg: Ergebnisse der Forschungen in den Jahren 2004 bis 2009. In: Krausse D (ed) Fürstensitze‘ und Zentralorte der frühen Kelten: Abschlusskolloquium des DFG-Schwerpunktprogramms 1171 in Stuttgart, 12.–15. October 2009. Theiss, Stuttgart
Baum N (1999) Die Dietersberghöhle bei Egloffstein, Kr. Forchheim: von der Opferhöhle zum Bestattungsplatz. Praehistorische Zeitschrift 74:79–121
Beard BL, Johnson CM (2000) Strontium isotope composition of skeletal material can determine the birth place and geographic mobility of humans and animals. J Forensic Sci 45:1049–1061
Becker C, Grupe G (2012) Archaeometry meets archaeozoology: Viking Haithabu and medieval Schleswig reconsidered. Archaeol Anthropol Sci 4:241–262
Ben-David M, Flaherty EA (2012) Stable isotopes in mammalian research: a beginner’s guide. J Mammal 93:312–328
Bentley RA, Krause R, Price TD, Kaufmann B (2003) Human mobility at the early Neolithic settlement of Vaihingen: evidence from strontium isotope analysis. Archaeometry 45:471–486
Bentley RA, Knipper C (2005) Geographical patterns in biologically available strontium, carbon and oxygen isotope signatures in SW Germany. Archaeometry 47:629–644
Bentley RA (2006) Strontium isotopes from the earth to the archaeological skeleton: a review. J Archaeol Method Theory 13:135–187
Britton K, Fuller BT, Tütken T, Mays S, Richards MP (2015) Oxygen isotope analysis of human bone phosphate evidences weaning age in archaeological populations. Am J Phys Anthropol 157:226–241
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51:337–360
Burmeister S (2000) Geschlecht, Alter und Herrschaft in der Späthallstattzeit Württembergs. Waxmann, Münster, New York
Burmeister S, Müller-Scheeßel N (2005) Der Methusalemkomplex: Methodologische Überlegungen zu Geschlecht, Alter und Sozialstatus am Beispiel der Hallstattzeit Süddeutschlands. In: Müller J (ed) Alter und Geschlecht in ur- und frühgeschichtlichen Gesellschaften [Tagung Bamberg 2004]. R. Habelt, Bonn
Burton JH, Price TD (2013) Seeking the local 87Sr/86Sr ratio to determine geographic origins of humans. In: Armitage RA, Burton JH (eds) Archaeological chemistry VIII. American Chemical Society, Washington, DC
Caut S, Angulo E, Courchamp F (2008) Discrimination factors (Δ15N and Δ13C) in an omnivorous consumer: effect of diet isotopic ratio. Funct Ecol 22:255–263
Chenery C, Müldner G, Evans J, Eckardt H, Lewis M (2010) Strontium and stable isotope evidence for diet and mobility in Roman Gloucester, UK. J Archaeol Sci 37:150–163
Codron D, Clauss M, Codron J, Tütken T (2018) Within trophic level shifts in collagen–carbonate stable carbon isotope spacing are propagated by diet and digestive physiology in large mammal herbivores. Ecol Evol 8:3983–3995
Daux V, Lécuyer C, Héran M-A, Amiot R, Simon L, Fourel F, Martineau F, Lynnerup N, Reychler H, Escarguel G (2008) Oxygen isotope fractionation between human phosphate and water revisited. J Hum Evol 55:1138–1147
Eckardt H, Chenery C, Booth P, Evans JA, Lamb A, Müldner G (2009) Oxygen and strontium isotope evidence for mobility in Roman Winchester. J Archaeol Sci 36:2816–2825
Ettel P (2002) Höhlen und Schachthöhlen – die Nutzung von Höhlen als Kultplätze in Mitteleuropa vom Neolithikum bis in die Latènezeit. In: Zemmer-Planck L (ed) Kult der Vorzeit in den Alpen: Opfergaben – Opferplätze – Opferbrauchtum/Culti nella preistoria delle Alpi: Le offerte – i santuari – i riti. Athesia, Bozen
Evans JA, Chenery CA, Fitzpatrick AP (2006) Bronze age childhood migration of individuals near Stonehenge, revealed by strontium and oxygen isotope tooth enamel analysis. Archaeometry 48:309–321
Fabrizii-Reuer S, Reuer E (1997) Anthropologische Untersuchungen an 138 Skeletten der Durezza-Schachthöhle bei Villach, Kärnten. Neues aus Alt-Villach 34:125–212
Fogel ML, Tuross N, Owsley D (1989) Nitrogen isotope tracers of human lactation in modern and archaeological populations. In: Annual report of the director, geophysical laboratory. Carnegie Institution of Washington, Washington, DC, pp 1988–1989
France CAM, Owsley DW (2015) Stable carbon and oxygen isotope spacing between bone and tooth collagen and hydroxyapatite in human archaeological remains. Int J Osteoarchaeol 25:299–312
Fuller BT, Richards MP, Mays SA (2003) Stable carbon and nitrogen isotope variations in tooth dentine serial sections from Wharram Percy. J Archaeol Sci 30:1673–1684
Fuller BT, Fuller JL, Harris DA, Hedges RE (2006a) Detection of breastfeeding and weaning in modern human infants with carbon and nitrogen stable isotope ratios. Am J Phys Anthropol 129:279–293
Fuller BT, Molleson TI, Harris DA, Gilmour LT, Hedges REM (2006b) Isotopic evidence for breastfeeding and possible adult dietary differences from Late/Sub-Roman Britain. Am J Phys Anthropol 129:45–54
Gleirscher P (1997) Überlegungen zur Deutung der Durezza-Schachthöhle. Neues aus Alt-Villach 34:213–238
GNIP Database n.d.. IAEA/WMO. Global network of isotopes in precipitation. The GNIP Database https://nucleus.iaea.org/wiser.
Goude G, Rey L, Toulemonde F, Cervel M, Rottier S (2017) Dietary changes and millet consumption in Northern France at the end of prehistory: evidence from archaeobotanical and stable isotope data. Environ Archaeol 22:268–282
Graf N, Graf R, Pasda K (2008) Das Peterloch bei Woppental. Naturhistorische Gesellschaft, Nürnberg
Grupe G, Price TD, Schröter P, Söllner F, Johnson CM, Beard BL (1997) Mobility of Bell Beaker people revealed by strontium isotope ratios of tooth and bone: a study of Southern Bavarian skeletal remains. Appl Geochem 12:517–525
Hald J (2009) Die Eisenzeit im Oberen Gäu: Studien zur hallstatt- und latènezeitlichen Besiedlungsgeschichte. K. Theiss, Stuttgart
Knipper C, Maus M (2016) Isotopenanalysen zur Rekonstruktion von Mobilität und Ernährungsweise der Bestatteten der hallstattzeitlichen Nekropole von Mauenheim. In: Wamser L (ed) Mauenheim und Bargen: Zwei Grabhügelfelder der Hallstatt- und Frühlatènezeit aus dem nördlichen Hegau. Reichert, Wiesbaden
Knipper C, Pichler SL, Rissanen H, Stopp B, Kühn M, Spichtig N, Röder B, Schibler J, Lassau G, Alt KW (2017) What is on the menu in a Celtic town? Iron Age diet reconstructed at Basel-Gasfabrik, Switzerland. Archaeol Anthropol Sci 9:1307–1326
Kornexl BE, Werner T, Roßmann A, Schmidt H-L (1997) Measurement of stable isotope abundances in milk and milk ingredients – a possible tool for origin assignment and quality control. Zeitschrift für Lebensmitteluntersuchung und -Forschung A 205:19–24
Lanfranco LP, Eggers S (2012) Caries through time: an anthropological overview. In: Li M-Y (ed) Contemporary approach to dental caries. InTech, Rijeka
Lightfoot E, Liu X, Jones MK (2013) Why move starchy cereals? A review of the isotopic evidence for prehistoric millet consumption across Eurasia. World Archaeol 45:574–623
Lightfoot E, O’Connell TC (2016) On the use of biomineral oxygen isotope data to identify human migrants in the archaeological record: intra-sample variation, statistical methods and geographical considerations. PLoS One 11(4):e0153850
Longinelli A (1984) Oxygen isotopes in mammal bone phosphate: a new tool for paleohydrological and paleoclimatological research. Geochim Cosmochim Acta 48:385–390
Maurer A-F, Galer SJG, Knipper C, Beierlein L, Nunn EV, Peters D, Tütken T, Alt KW, Schöne BR (2012) Bioavailable 87Sr/86Sr in different environmental samples – effects of anthropogenic contamination and implications for isoscapes in past migration studies. Sci Total Environ 433:216–229
Meyer-Orlac R (1982) Mensch und Tod: Archäologischer Befund - Grenzen der Interpretation. K. Renner Verlag, Hohenschäftlarn
Moghaddam N, Müller F, Hafner A, Lösch S (2016) Social stratigraphy in Late Iron Age Switzerland: stable carbon, nitrogen and sulphur isotope analysis of human remains from Münsingen. Archaeol Anthropol Sci 8:149–160
Müller-Scheeßel N (2013) Im Tode gleich? Eisenzeitliche Bestattungen von Frauen und Männern in Siedlungskontexten und in ‚regulären Gräbern im Vergleich. In: Wefers S, Fries JE, Fries-Knoblach J, Later C, Rambuscheck U, Trebsche P, Wiethold J (eds) Bilder – Räume – Rollen: Beiträge zur gemeinsamen Sitzung der AG Eisenzeit und der AG Geschlechterforschung während des 7. Deutschen Archäologenkongresses in Bremen 2011. Beier & Beran, Langenweißbach
Müller-Scheeßel N, Grupe G, Tütken T (2015) In der Obhut von Verwandten? Die Zirkulation von Kindern und Jugendlichen in der Eisenzeit Mitteleuropas. In: Karl R, Leskovar J (eds) Interpretierte Eisenzeiten: Fallstudien, Methoden, Theorie. Tagungsbeiträge der 6. Linzer Gespräche zur interpretativen Eisenzeitarchäologie. Oberösterreichisches Landesmuseum, Linz
Oelze VM, Koch JK, Kupke K, Nehlich O, Zäuner S, Wahl J, Weise SM, Rieckhoff S, Richards MP (2012) Multi-isotopic analysis reveals individual mobility and diet at the Early Iron Age monumental tumulus of Magdalenenberg, Germany. Am J Phys Anthropol 148:406–421
Pauli L (1975) Keltischer Volksglaube: Amulette und Sonderbestattungen am Dürrnberg bei Hallein und im eisenzeitlichen Mitteleuropa. C. H. Beck’sche Verlagsbuchhandlung, München
Pauli L, Glowatzki G (1979) Frühgeschichtlicher Volksglaube und seine Opfer. Germania 57:144–152
Peter-Röcher H (1997) Menschliche Skelettreste in Siedlungen und Höhlen: kritische Anmerkungen zu herkömmlichen Deutungen. Ethnographisch-Archäologische Zeitschrift 38:315–324
Peter-Röcher H (1999) Die Höhle von Bycí Skála: Gaben an Götter und Ahnen. Mitt Berliner Ges Anthropol Ethnol Urgesch 18:47–56
Peter-Röcher H (2003) Ritual – Opfer – Totenkult: zur Kontroverse um die nacheiszeitliche Höhlennutzung. In: Metzner-Nebelsick C (ed) Rituale in der Vorgeschichte, Antike und Gegenwart: Studien zur Vorderasiatischen, Prähistorischen und Klassischen Archäologie, Ägyptologie, Alten Geschichte, Theologie und Religionswissenschaft. Interdisziplinäre Tagung vom 1.-2. Februar 2002 an der Freien Universität Berlin. M. Leidorf, Rahden/Westf
Peterson BJ, Fry B (1987) Stable isotopes in ecosystem studies. Ann Rev Ecol Systematics 18:293–320
Price TD, Burton JH, Bentley RA (2002) The characterization of biologically available strontium isotope ratios for the study of prehistoric migration. Archaeometry 44:117–135
Price TD, Knipper C, Grupe G, Smrcka V (2004) Strontium isotopes and prehistoric human migration: the Bell Beaker period in Central Europe. Eur J Archaeol 7:9–40
Röhrig K-H (1994) Das hallstattzeitliche Gräberfeld von Dietfurt a. d. Altmühl. Leidorf, Buch am Erlbach
Schellenberg A, Chmielus S, Schlicht C, Camin F, Perini M, Bontempo L, Heinrich K, Kelly SD, Rossmann A, Thomas F, Jamin E, Horacek M (2010) Multielement stable isotope ratios (H, C, N, S) of honey from different European regions. Food Chem 121:770–777
Sealy JC (2001) Body tissue chemistry and palaeodiet. In: Brothwell DR, Pollard AM (eds) Handbook of archaeological sciences. Wiley, Chichester
Seregély T, Burgdorf P, Gresik G, Müller MS, Wilk A (2015) ‘Tote Menschen und Tiere in finsteren Felsschächten …’: neue Dokumentationsmethodik und erste Untersuchungsergebnisse zur Kirschbaumhöhle in Oberfranken. Praehistorische Zeitschrift 90:214–244
Shay T (1985) Differentiated treatment of deviancy at death as revealed in anthropological and archeological material. J Anthropol Archaeol 4:221–241
Stephan E (2009) Rekonstruktion eisenzeitlicher Weidewirtschaft anhand archäozoologischer und isotopenchemischer Untersuchungen. In: Benecke N (ed) Beiträge zur Archäozoologie und Prähistorischen Anthropologie VII. Gesellschaft für Archäozoologie und Prähistorischen Anthropologie e.V, Langenweißbach
Styring AK, Sealy JC, Evershed RP (2010) Resolving the bulk δ15N values of ancient human and animal bone collagen via compound-specific nitrogen isotope analysis of constituent amino acids. Geochim Cosmochim Acta 74:241–251
Torbrügge W (1979) Die Hallstattzeit in der Oberpfalz I: Auswertung und Gesamtkatalog. Michael Lassleben, Kallmünz/Opf
Tütken T, Knipper C, Alt KW (2008a) Mobilität und Migration im archäologischen Kontext: Informationspotential von Multi-Element-Isotopenanalysen (Sr, Pb, O). In: Bemmann J, Schmauder M (eds) Kulturwandel in Mitteleuropa: Langobarden – Awaren – Slawen. R. Habelt, Bonn
Tütken T, Langenegger E, Wild W (2008b) Einheimisch oder fremd? Isotopenanalyse eines Frauenskelettes des 9. Jahrhunderts n. Chr. aus Elsau, Kanton Zürich, Schweiz / native or not? Isotope analysis of a female skeleton on the 9th century A.D. from Elsau, Canton Zurich, Switzerland. Anthropol Anz 66:19–50
Tütken T (2010) Die Isotopenanalyse fossiler Skelettreste - Bestimmung der Herkunft und Mobilität von Menschen und Tieren. In: Meller H, Alt KW (eds) Anthropologie, Isotopie und DNA - biografische Annäherung an namenlose vorgeschichtliche Skelette: 2. Mitteldeutscher Archäologentag vom 08. bis 10. Oktober 2009 in Halle (Saale). Landesamt für Denkmalpflege und Archäologie in Sachsen-Anhalt, Landesmuseum für Vorgeschichte, Halle/Saale
White C, Longstaffe FJ, Law KR (2004) Exploring the effects of environment, physiology and diet on oxygen isotope ratios in ancient Nubian bones and teeth. J Archaeol Sci 31:233–250
Zäuner S, Wahl J (2013) Zur demographischen Struktur der Bestattungen im späthallstattzeitlichen Grabhügel vom Magdalenenberg. Fundberichte aus Baden-Württemberg 33:133–145
Funding
Open Access funding provided by Projekt DEAL. Financial support by the German Research Foundation (grant no. MU 3053/1, PI: Müller-Scheeßel and grant no. TU 148/2-1, PI: Tütken) is thankfully acknowledged. Anja Staskiewicz provided great help in sampling the individuals. We further thank Jennifer Leichliter who brushed up the English.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Müller-Scheeßel, N., Grupe, G., Mühldorfer, B. et al. The dead of the Dietersberg Cave, Germany: new insights into burial practices of the Iron Age from 14C-dates and stable isotope (C, N, O, Sr) analyses of human bones and teeth. Archaeol Anthropol Sci 12, 68 (2020). https://doi.org/10.1007/s12520-020-01025-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-020-01025-1