
Abstract
Brazil is the world’s largest producer of sugarcane and one of the leading suppliers of sugar and ethanol worldwide. In the 2019–2020 crop season, the country produced 642.7 million tons of sugarcane in a harvest area of 8.44 million hectares. Historically, sugarcane breeding has contributed continuously to increasing yields by regularly releasing superior cultivars for use by the Brazilian industry. In the last 40 years, an average annual increase of 155.7 kg ha−1 of sugar yield has been reported, about half of which may be attributed to breeding programs. However, due to the size of the country, the intensive expansion of the crop to low-fertility soils in the last few years, especially in degraded pasture areas, and the widespread adoption of mechanization, new challenges have been imposed on national breeding programs. This review covers the current situation with sugarcane breeding in Brazil and the main advances that have allowed the country to maintain world leadership in developing the industry. Additionally, the history of sugarcane breeding, current national breeding institutions, germplasm development, key breeding objectives, selection stages and methodologies are summarized. An overview is also presented of biotechnological approaches which have become key tools for improving Brazilian traditional breeding programs. The adoption of strategies to increase Brazilian sugarcane yield, aiming to consolidate crop production in a food and energy matrix, is also discussed.
Similar content being viewed by others


Main Characteristics of the National Industry
Brazil is the world’s largest producer of sugarcane, followed by India, China, Thailand, Pakistan and Mexico (FAO-Food and Agriculture Organization of the United Nations 2020). The country has 404 sugar, flex-ethanol and distillery plants (sugarcane mills) and approximately 70,000 growers (UDOP-União Nacional da Bioenergia 2020; UNICA-União Indústria de Cana-de-Açúcar 2020). According to the Brazilian National Food Supply Company, during the 2019–2020 crop year, 642.7 million tons of sugarcane were produced in the country, in a harvest area of 8.44 million hectares (Fig. 1; Table 1). The Central-South region was responsible for 92% of domestic production, with more than 589 million tons, while the North and Northeast regions produced 52.8 million tons, which corresponded to 8% of production (CONAB-Companhia Nacional de Abastecimento 2020).

Harvested area and sugarcane production in Brazil—2005–2006 to 2019–2020 crop years
Starting in 2005, there was significant growth in sugarcane production in Brazil, as shown in Fig. 1. This growth may be explained by the increased ethanol demand with the adoption of vehicles with flex-fuel technology, which offer environmental and socioeconomic benefits (Antunes et al. 2019). In 2008, low sugar and ethanol prices, coupled with the global financial crisis, limited investment and affected the renewal of sugarcane plantations in the following years. In addition to the negative effects of the market, successive droughts further harmed Brazilian sugarcane production and agriculture as a whole. Significant decreases in sugarcane yield were observed especially after 2011 (Fig. 2). Moreover, the intense implementation of mechanized harvesting and expansion of the crop to low-fertility soils, especially in degraded pasture areas, also negatively impacted crop productivity (Antunes et al. 2019).

Sugarcane yield in Brazil—2005–2006 to 2019–2020 crop years
In the 2019–2020 crop year, the average cane yield in Brazil was 76.13 t ha−1, with a mean total recoverable sugar (TRS) of 139.3 kg t−1 and an average crop productivity, in tons of TRS per hectare, of 10.6 t ha−1, as shown in Fig. 2 (CONAB-Companhia Nacional de Abastecimento 2020). TRS is the total sugar contained in sugarcane and is used to quantify sugar and ethanol production (CONSECANA-Conselho dos Produtores de Cana-de-Açúcar Açúcar e Álcool do Estado de São Paulo-SP 2006). This information is given in kilograms of sugar per ton of sugarcane and is the most used metric in the country to represent sugarcane quality. The percentage of TRS allocated to ethanol or sugar varies mainly according to the market; in the 2019–2020 crop year, 65.1% of the TRS was allocated for ethanol production, while 34.9% was allocated for sugar production (CONAB-Companhia Nacional de Abastecimento 2020).
Brazilian ethanol production in the 2019–2020 crop year was 34 billion liters: 23.89 billion liters of hydrous ethanol and 10.12 billion liters of anhydrous ethanol. In Brazil, the mandatory percentage of anhydrous ethanol added in gasoline is 25%. The volume of ethanol exported was 1.9 billion liters, representing an increase of 6.2% compared to the previous crop year. The devaluation of the Brazilian currency (Real) against the US dollar and the increase in the supply of fuel in the country are the main reasons for this growth. Consequently, the balance between exports and imports of ethanol showed a surplus of 238.57 million liters, i.e., USD 286.06 million in value (CONAB-Companhia Nacional de Abastecimento 2020).
Regarding sugar, 29.8 million tons were produced in the 2019–2020 crop year, of which approximately 18.9 million tons were exported, generating approximately US$ 5.8 billion for the country. This value was 4.6% lower than in the previous cycle and can be explained by the increase in the global sugar supply, which forced the reduction in international prices. As a result, Brazilian production units showed reduced interest in supplying sugar to the international market. The drop could have been even greater, but exports recovered in the final half of the crop year after news of weather issues and declined production in India and Thailand (CONAB-Companhia Nacional de Abastecimento 2020).
In addition to its use for sugar and ethanol production, sugarcane in Brazil has also been increasingly used as a source of power, in a so-called cogeneration system. The country has the technical potential for the cogeneration of 146,000 GWh, if there was full use of the biomass present in the sugarcane fields. In 2018, 21,500 GWh were produced, i.e., only 15% of the potential (UNICA-União Indústria de Cana-de-Açúcar 2020). Other products that can be obtained from sugarcane are second-generation ethanol or cellulosic ethanol; biogas originating from vinasse; biomethane obtained from biogas, whose application is the same as natural gas; plastic; and other uses in the chemical industry.
Harvesting in Brazil is now predominately mechanized. The percentage of mechanized harvesting increased from 24.4% in 2007–2008 to 88.4% in 2019–2020. In the Central-South region, where the terrain is more favorable to mechanization, the adoption rate of mechanized harvesting is currently 92.9%. In the North and Northeast regions, the percentage of mechanized harvesting stands at 23%; in Alagoas and Pernambuco, which represent more than 60% of the harvested area in the region, the percentages of mechanized harvesting adoption are 21.3% and 1.9%, respectively (CONAB-Companhia Nacional de Abastecimento 2020). The low level of adoption of mechanized harvesting in the North and Northeast regions is explained by the rugged terrain in some of the production areas and the greater availability of labor for manually harvesting.
In terms of longevity, the index used in Brazil to evaluate the aging level of sugarcane plantations is the average cutting cycle. This index is measured in years and is calculated as the weighted average of the harvested area at each cutting cycle in a given mill or region. The higher the average cutting cycle is, the older the cane fields are, while the lower the average cutting cycle is, the younger the cane fields are. In the 2017–2018 crop year in the Central-South region, this index was 3.77, while the intensity of sugarcane field renewal, i.e., the rate of tilling the old stubble and re-establishment of new plantings, given by the planting/harvest ratio, was 13.7%. In the North and Northeast regions, the average cutting cycle was 4.39, indicating that their sugarcane fields are older than those in the Central-South region. On the other hand, the planting/harvest ratio was 14.7%; thus, the sugarcane field renewal intensity was higher than that of the Central-South region (Braga Júnior et al. 2019). Even so, the presence of older sugarcane plantations and the greater variations in water distribution explain the lower productivities in the North and Northeast Regions than in the Central-South region: In the 2019–2020 crop year, the average agricultural productivity of the former was 59.38 t ha−1 and that of the latter was 78.11 t ha−1 (CONAB-Companhia Nacional de Abastecimento 2020).
Other factors that may negatively impact the productivity of Brazilian sugarcane plantations are diseases and pests. In the country, the diseases that cause most damage to crops are ratoon stunting (bacterium, Leifsonia xyli subsp. xyli), leaf scald (bacterium, Xanthomonas albilineans), orange rust (fungus, Puccinia kuehnii), sugarcane smut (fungus, Sporisorium scitamineum), mosaic disease (sugarcane mosaic virus—ScMV) and brown rust (fungus, Puccinia melanocephala). Generally, the most recommended control method used in the country for these diseases is the planting of resistant varieties. In the case of ratoon stunting, leaf scald, sugarcane smut and mosaic disease, planting of healthy seedlings with resistant varieties is recommended. Although it is not possible to estimate the percentage of crop planted with certified disease-free cane, the demand for healthy propagating materials among Brazilian sugarcane growers has increased, particularly in the last two or three years.
The pests that cause greatest damage to sugarcane crops in Brazil include the sugarcane borer (Diatraea saccharalis), Sphenophorus (Sphenophorus levis), spittlebug (Mahanarva fimbriolata and Mahanarva posticata), Migdolus (Migdolus fryanus) and giant sugarcane borer (Telchin licus). The control of these pests is based on the adoption of various pest management practices throughout sugarcane cultivation, from soil preparation to sugarcane field renewal; the measures most used by Brazilian growers comprise biological and chemical control methods. In 2017, CTC20BT was commercially released, a genetically modified variety resistant to sugarcane borer. Use of this variety has been gradually expanded, and the results of pest control in fields planted in CTC20BT have been positive (Cheavegatti-Gianotto et al. 2019).
Brazilian sugarcane plantations are also subject to damage by abiotic stress. Among adverse environmental factors, drought is the most common, especially in the North and Northeastern regions and in some Central-South states. In the more southern states of the country, growers deal with low temperatures and frost, conditions that may occasionally also affect other states in the Central-South region. The flowering of sugarcane, which is influenced by environmental conditions, can also cause losses, especially in the North and Northeastern regions, where the conditions are extremely conducive to this phenomenon. In the Central-South region, flowering can also affect crop yield, but the frequency and intensity vary according to year and location.
History and Breeding Achievements
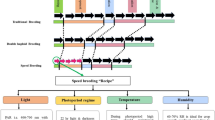
In Brazil, breeding has always provided a major contribution to increase the agroindustrial yield of the sugarcane production chain. For more than three centuries, cultivation of S. officinarum varieties, such as Creole and Caiana, was predominant. The first hybrids were introduced in the 1930s, including POJ 2878, Co 290, Co 419 and Co 331 (Andrade 1985; Landell and Bressiani 2008).
Pioneering milestones for sugarcane breeding in Brazil were achieved in the 1930s by Campos Brasil (CB), from the Experimental Sugarcane Station of Campos de Goytacazes/Rio de Janeiro, and the Ministry of Agriculture and the Agronomic Institute of Campinas (IAC, for its initials in Portuguese) in the state of São Paulo. In the CB variety program, CB41-76 and CB45-3 stood out in the subsequent decades (Fig. 3) (Andrade 1985; Cesnik and Miocque 2004). The IAC program is the oldest ongoing program in Brazil. Until the second half of the twentieth century, the most important varieties were IAC48-65, IAC50-134, IAC51-205 and IAC52-150. At the end of the last century, the program underwent a restructuring/reorganization of its activities, in partnership with the sugarcane industry, and a hybridization station was created in Uruçuca, Bahia (14°28′ S, 39°04′ W, 90 m) (Fig. 4). Since then, 19 commercial releases have occurred; currently, the most prominent varieties are IAC91-1099, IACSP95-5000 and IACSP95-5094 (Cesnik and Miocque 2004; Landell and Bressiani 2008; Instituto Agronômico de Campinas 2020).

Adapted from Dal-Bianco et al. (2012)
List of the ten main sugarcane varieties cultivated in Brazil between 1975 and 2019, with the respective cultivation ratios.

Sugarcane breeding programs and sugarcane germplasm collections in Brazil
In 1933, the federal government created the Sugar and Alcohol Institute (IAA, for its initials in Portuguese), which for more than half a century had the following missions: establishment of sugarcane production quotas for sugarcane mills and suppliers; administration of the price of sugarcane, sugar and alcohol; establishment of export rules; and other interventionist measures. Between 1960 and 1970, the average Brazilian yield was below four tons of sugar per hectare (IAA 1972; Barbosa et al. 2012). During this period, there was massive investment by the public–private sectors, both in the import of germplasm and in the implementation of new breeding programs. In 1966, the IAA introduced an early-maturing variety from Chacra Experimental Agrícola Santa Rosa—northern Argentina (NA56-79). This variety made important contributions, including in facilitating expansion of the harvesting period, and changed sugar production benchmarks in the 1980s. During that period, the cane payment method was also changed according to the sucrose content or the quality of the raw material in the Central-South region. Cultivation of NA56-79 ceased in the following decade due to its susceptibility to sugarcane smut (IAA 1972; Matsuoka et al. 2009) (Fig. 3). However, in 1966, the IAA and the production sector of the state of Alagoas created a hybridization station in Serra do Ouro, in Murici, Alagoas (09°13′ S, 35°50′ W, 500 m), where the RB (República do Brasil) varieties originated (Fig. 4).
In 1971, the IAA initiated the National Sugarcane Improvement Program (PLANALSUCAR), a research and development institution in agricultural and industrial areas, which was fundamental for the establishment in 1975 of the National Alcohol Program (PROÁLCOOL), which boosted the Brazilian sugar and alcohol industry, with the massive production of fuel ethanol to replace imported oil. In that year, PLANALSUCAR took over sugarcane hybridizations at Serra do Ouro, Alagoas, to obtain the RB cultivars (IAA 1972; Barbosa 2018). In two decades, this program released 19 varieties, with its greatest contribution being variety RB72454, which led in terms of planted sugarcane area in Brazil between 1995 and 2010 (Fig. 3). At that time, due to its extensive cultivation area, it was considered the most cultivated variety in the world. However, RB72454 is no longer planted due to its susceptibility to orange rust (Puccinia kuehnii) (Braga Júnior et al. 2011; Barbosa et al. 2012). In 1990, RB varieties occupied only 5% of the sugarcane plantations of Brazil, and in that year, the IAA/PLANALSUCAR was ended. The physical structures, human resources and technologies of the program were transferred to the federal universities in their areas of influence: Federal University of Alagoas (UFAL), Federal University of São Carlos-São Paulo (UFSCar), Federal Rural University of Pernambuco (UFRPE), Federal University of Viçosa-Minas Gerais (UFV), Federal Rural University of Rio de Janeiro (UFRRJ), Federal University of Paraná (UFPR), Federal University of Sergipe (UFS), Federal University of Goiás (UFG), Federal University of Mato Grosso (UFMT) and Federal University of Piauí (UFPI) (Fig. 4). As a result, the Inter-university Network for the Development of the Sugarcane Industry (RIDESA-Rede Interuniversitária Para o Desenvolvimento do Setor Sucroenergético) was established in Brazil. A public–private partnership was then established between RIDESA and companies from the national sugarcane sector, which started to provide research funding. In 30 years, RIDESA released 75 RB varieties. Several contributions were made by the network; in the Central-South region, for example, there were releases of early-maturing varieties that came to stand out—RB835054, RB835486, RB855156, RB855453 and RB855536. This maturation profile allowed the sugarcane crop season to start two months earlier in this region. Currently, RB855156, RB855453 and RB855536 are still very important in the national scenario, occupying the 5th, 6th and 10th positions, respectively, among the most cultivated varieties in the country (Table 2). However, the greatest contribution by that network in recent years was the release of variety RB867515, which has been the most cultivated variety in Brazil in the last 15 years, allowing planting in low-fertility soil conditions without a marked decline in yield (Table 2). RIDESA continues to justify the investments by the partner companies because in the last decade, two RB varieties have predominated in sugarcane plantations: RB92579 in the Northeast region and RB966928 in the Central-South region (Fig. 3; Table 2).
The contribution of private initiatives began in 1969 with Cooperativa Central dos Produtores de Açúcar e Álcool do Estado de São Paulo (COPERSUCAR), which produced the SP (São Paulo) varieties and established a hybridization station in Camamu, Bahia (13°91′S, 39°16′W, 100 m) (Fig. 4). Between 1985 and 2000, the SP varieties dominated sugarcane cultivation in Brazil (Fig. 3), especially SP70-1143 and SP71-1406. Subsequently, these two varieties stopped being used due to their susceptibility to brown rust (Puccinia melanocephala). SP71-6163 was subsequently shown to be very productive but was discarded due to susceptibility to sugarcane yellow leaf virus (ScYLV) (Cesnik 2004). Between 2000 and 2015, other SP varieties had significant cultivation areas—SP79-1011, SP80-1816, SP80-1842, SP81-3250 and SP83-2847. Among these, SP81-3250 was one of the most cultivated varieties but also stopped being cultivated due to its susceptibility to orange rust (Braga Júnior et al. 2011; Barbosa 2018) (Fig. 3). However, due to the large contribution of this variety to the area cultivated with sugarcane, it continues to be among the ten most cultivated varieties in the country (4%) despite its ongoing replacement with other varieties (Table 2). COPERSUCAR’s program ended its activities in 2003, and since 2004, it has been managed by the Sugarcane Technology Center (CTC, for its initials in Portuguese), which has been developing CTC varieties (Fig. 4). Since then, there has been a great increase in the cultivation area of these varieties, especially for CTC4 and CTC15, which are the 4th and 9th most cultivated varieties in Brazil, respectively (Table 2), and CTC9001 (Fig. 3).
According to the Brazilian Ministry of Agriculture, Livestock and Supply (Ministério da Agricultura and Pecuária e Abastecimento 2020), there are a total of 214 sugarcane cultivars registered in Brazil, among which 68 are RB, 38 are CTC, 37 are SP, 33 are IAC and 38 are other varieties. In addition to the breeding programs of the CTC, IAC and RIDESA, since 2012, BioVertis/GranBio, a Brazilian company founded in 2011, has been working on developing sugarcane varieties with high biomass (energycane) yield and low production cost, with the main objective of obtaining varieties with low-cost biomass and high energy efficiency and that can be grown in restrictive areas to avoid competition with food production. Currently, the company has 11 varieties registered in the country, under the name Vertix®. It should be noted that in 2003, with resources from the private group Votorantim and the technical staff of ex-professors/researchers linked to the Federal University of São Carlos, the program CanaVialis, whose varieties are named CV, was started. This program was acquired by Monsanto in 2008 and ended in October 2015; there are CV varieties that, although not yet released, are cultivated (free of charge) in representative areas in the country.
Among the four ongoing breeding programs in Brazil (Table 3), the CTC is the only one that has a publicly traded company with shares listed but not traded in the “Bovespa Mais” segment (CTCA3) and uses royalties as a source of revenue for financing the research and development of varieties. This model was also adopted by the BioVertis/GranBio breeding program. In contrast, both the IAC and the universities affiliated with RIDESA, all public, receive contributions through signed agreements with mills and sugarcane grower associations. In general, the values are based on the amount of sugarcane produced by the mills or delivered by the members in the different processing units. In addition, there are other financing models for different lines of research, such as biotechnology, physiology, microbiology and crop science, among others, which have been contributing continuously to public breeding programs. They include the FAPESP Bioenergy Research Program (BIOEN), the Funding Authority for Studies and Projects (FINEP) and the National Institute of Bioethanol Science and Technology (INCT-Bioethanol).
In Brazil, sugarcane breeding has enabled constant gains. In 1970, sugar productivity was 3712 kg ha−1, and in 2011, it reached 9148 kg ha−1, an average annual increase of 155.7 kg ha−1 year−1 from 1970 to 2011. This corresponds to 4% per year, with half attributed to breeding, representing a contribution to the country of 175 million dollars in the 2011–2012 crop year (Barbosa et al. 2012); 66% growth in agricultural productivity and 34% growth in cane sugar yield between 1975 and 2010 (Dal-Bianco et al. 2012); and a sugar yield gain of 1.5% per year in the state of São Paulo since 1945 (Hoffmann et al. 1999). In the same state, between 1986 and 1991, SP cultivars provided a gain of 130 million dollars (Fernandes and Tatizana 1991); in Alagoas, there was an annual increase of 80 kg ha−1 of sugar between 1975 and 1992 and of 140 kg ha−1 between 1993 and 2010, with the cultivar RB92579 contributing 1830 kg ha−1 more sugar than the standard cultivar in the 2010–2011 crop year, with a gain of USD 625.54 ha−1 or 50 million dollars (Barbosa 2018).
Barbosa (2018) reported that among the oldest varieties grown in northeastern Brazil, Co 331 and CB45-3 had a useful life of approximately 50 years and are the longest-lived cultivars in Brazil in the twentieth century (Matsuoka et al. 2009). Furthermore, the useful life of the most recent varieties is as follows: RB72454 and SP81-3250, 20 years; SP79-1011 and SP80-1842, close to 30 years; RB855156 and RB855453, 25 years; RB867515, 23 years; SP83-2847, 20 years; RB92579, 17 years; and RB966928, 10 years. In Brazil, the protection of plant varieties is regulated by the Ministry of Agriculture, Livestock and Food Supply, which is responsible for registering the varieties with the National Plant Variety Protection Service (SNPC, for its initials in Portuguese), and the protection time is 15 years.
Germplasm Collection and Development
Sugarcane germplasm collections in Brazil vary in size and are formed by species of the Saccharum complex, commercial cultivars and elite clones with advantageous traits for breeding (Table 4). Over the years, several efforts have been made to characterize accessions in terms of the main traits of economic value and to spur improvement. However, these activities are highly complex, and many accessions still need to be characterized for better use in breeding, which are performed at different improvement institutions. Currently, there are five germplasm collections, all located in northeastern Brazil, as it offers favorable climatic conditions for the profuse flowering of sugarcane and the production of fertile pollen. Two of these serve the RIDESA (RB) programs, one the CTC program, one the IAC program and one the BioVertis/GranBio program, with the latter dedicated exclusively to energycane breeding (Fig. 4). Despite the achievements in sugarcane breeding, almost all varieties planted worldwide are derived from a few parent plants, with an apparent narrowing of the genetic base of modern clones (Berding and Roach 1987). This factor, combined with the complex sugarcane genome, has limited breeding advances (Singh et al. 2008).
Over the course of decades, several expeditions were carried out, resulting in the incorporation of various species into two world collections, one in India and another in the USA. These collections serve as genetic reservoirs available to breeders for the production of new cultivars that are more productive and resistant to pests and diseases. With these accessions, since the 1960s, many programs began to perform introgression to expand the genetic base of their breeding populations and, consequently, increase the genetic variability of their progenies for various traits.
Moreover, with the growing need for renewable energy sources, interest in biomass emerged, but without much emphasis on sugar. In the 1980s in Brazil, some programs selected clones with high biomass but discarded them in the conventional selection process due to low sucrose and high fiber content. These programs were not successful in convincing the sugarcane industry of the importance of this strategy for selecting and developing more productive and fibrous sugarcane.
In the 1990s, a Brazilian program began an introgression project with Erianthus, specifically with E. arundinaceus, in the search for clones that maintained the current sucrose levels and presented high agricultural productivity, with upright architecture, higher fiber content, excellent sprouting, greater resistance to pests and tolerance to saline soils, drought and cold. Observations over the last 30 years showed that due to its genetic distance from the Saccharum genus, less than 30% of Erianthus accessions were able to produce viable seeds, especially when used as pollen suppliers. When Erianthus is employed as a female parent, seedlings are most often apomictic and are easily identified by the absence of dewlap.
In the last 20 years breeding programs in Brazil have imported hundreds of basic germplasm accessions and historical hybrids from the world collection in Miami for use in introgression and development of energycanes. Since then, several introgression activities with wild species, especially S. spontaneum, have started, directing selection toward resilient plants with high fiber and less sucrose but with high biomass productivity (Carvalho-Netto et al. 2014; Matsuoka et al. 2014). The main activities carried out in Brazil were developed by companies that are no longer in operation (Canavialis, Vignis and AGN Bioenergy) to obtain energycanes. Currently, these activities continue to be performed with less effort by IAC and RIDESA and with full dedication by BioVertis/GranBio.
Recent introgression efforts led to the registration and protection at the Ministry of Agriculture, Livestock, and Supply of 11 energycane cultivars developed by BioVertis/GranBio with the acronym Vertix; these cultivars are in the initial phases of multiplication by the companies in the sector. However, the success of this strategy will only be achieved in the coming years with greater commercial adoption and also with entrance of second generations technologies in the market.
Current Key Breeding Objectives
Sugarcane
In general, sugarcane breeding programs seek to obtain new varieties that are more favorable to different growing conditions, by maximizing the presence of desirable traits. However, this is a complex process due to several factors, such as the long cycle of generating a new variety, which can extend from 10 to 15 years (Gazaffi et al. 2010, 2016; de Morais et al. 2015). During this period, the economic value of traits may change, and new diseases may determine whether a genotype, previously considered appropriate, becomes obsolete. To that end, each breeding program establishes its objectives according to the needs of the sugarcane industry. The main objectives for RIDESA, in decreasing order of importance, are high cane and sugar yield, disease resistance, sprouting and ratoon ability (de Morais et al. 2015), among others, as described below.
The cane and sugar yield can be defined as the sugar production per area and can be obtained by two secondary components: tons of sugarcane per hectare (TCH) and sugar content (measured by Pol% cane and/or TRS). Both factors are extremely important because they are used to determine the price and payment method of sugarcane in Brazil (CONSECANA-Conselho dos Produtores de Cana-de-Açúcar Açúcar e Álcool do Estado de São Paulo-SP 2006). The TCH component has moderate heritability (Balsalobre et al. 2016) and can be inferred based on the traits height, diameter and number of stalks (tillering). The Pol% cane trait has high heritability (Balsalobre et al. 2016) and is a function of Brix, fiber and purity (Cursi et al. 2016). Therefore, the indirect components should also be considered for the composition of the main factor.
Disease resistance is the second most important trait because it replaces the need for the use of agrochemicals for the control of fungal diseases. The diseases with great importance for Brazilian agriculture and with genetic resistance mechanisms are orange rust (Puccinia kuehnii), brown rust (Puccinia melanocephala), sugarcane smut (Ustilago scitaminea) and mosaic disease (SCMV—sugar cane mosaic virus) (de Morais et al. 2015). In addition, the search for genetic resistance to other diseases is also highly desired, as is the case of ratoon stunting disease (RSD). However, due to the complexity in the selection of varieties resistant to RSD—arising from the difficulty in the rapid and efficient diagnosis of the disease—the use of clean seeds, hot water treatment and disinfection of equipment are sought as preventive methods for disease prevention. Nevertheless, according to Urashima et al. (2020), between 2013 and 2018, the mean RSD infestation in Brazilian sugarcane plantations was approximately 40%, when considering the five most planted varieties.
Sprouting and sugarcane ratoon ability have gained greater importance in Brazil since the introduction of mechanical harvesting and the gradual ban on controlled burnings for harvesting (Cortez and Baldassin Jr. 2016). The presence of straw has numerous benefits (Bordonal et al. 2018) but can also create challenges, such as greater difficulty in ratooning (Cortez and Baldassin Jr. 2016). Furthermore, increased use of heavy agricultural machinery can lead to increased soil compaction and, consequently, to the reduction in sugarcane ratoon yields. This issue is addressed by selecting genotypes with high sprouting ability.
Additional objectives include adaptability and yield stability; growth rate; upright growth habit; absence of flowering; self-trashing; high fiber content (depending on the objective); and resistance or tolerance to abiotic stresses, such as drought, low temperatures, frost, high aluminum concentration and low phosphorus levels.
Energycane
Different objectives can be considered in energycane improvement. At GranBio, there is an ongoing search for varieties with the following profiles:
-
1.
High biomass production potential, supporting the production of renewable energy that contributes to mitigating the greenhouse effect, with high conversion capacity of atmospheric C into organic C;
-
2.
High energy density, that is, having more efficient raw material than that of food plants;
-
3.
High resistance to biotic and abiotic stresses, allowing production with less input (less fertilizer, pesticide and energy) and in lands with lower agronomic value, i.e., less fertile, with less water availability, more extreme temperatures (both low and high) and more saline soils;
-
4.
Possessing a fasciculate and exuberant root system with high efficiency in soil C fixation and erosion control.
In general, the energycanes are classified into two categories: Type I and Type II. Type I is described as a cane closer to the conventional sugarcane but with lower sucrose content and thus lower purity but a higher fiber content than conventional sugarcane. Type II is a cane with only marginal sugar content but with higher fiber content than Type I, to be used exclusively for biomass production (Tew and Cobill 2008). For GranBio, it is expected that the cane fiber content for Type II varieties will be twice as high in the medium term, with a 20% to 50% lower sugar content in juice compared to sugarcane. It is also expected that energycane would be more tolerant to pests and diseases and that it has a much faster improvement cycle than that of sugarcane: four to six years instead of 8 to 12. Finally, it is believed that the multiplication rate of energycane is much higher than that of sugarcane and may reach 1:400 in one year, 16 times higher than that of sugarcane.
Based on the objectives listed above, GranBio established the product concept for the selection of Vertix type 1 and Vertix type 2 cultivars, as shown in Table 5.
Regardless of the type of sugarcane sought by breeding, it is possible that over time, new challenges will emerge for crops, for example, the introduction of new diseases and/or pests, that alter the priority list over time. In this case, knowledge of the crop and the prospecting ability are essential for the breeder in the development of a future variety (de Morais et al. 2015).
Breeding Methods
Sugarcane
Conceptually, a sugarcane breeding program can be organized into three main stages, namely generation of genetic variability, selection and cloning in the initial stages, characterized by experiments with few replications and small plots, and selection in the final stages, with a sufficient number of replicates and sites to perform experiments with high precision (Gazaffi et al. 2010). Usually, this structure is common both in Brazil and in other countries; therefore, the main difference is in the way each stage is organized and conducted.
The generation of genetic variability occurs through crossing heterozygous individuals and the selection of superior individuals in the segregating population. As a source of variability, sugarcane breeding programs in Brazil use different germplasm collections described previously. One common challenge for all breeding programs is the synchronization of flowering among the different genotypes present in the germplasm collection. Staggered planting can be performed to mitigate the asynchronization issue; however, it is not an efficient strategy for all cases. With the purpose of circumventing this type of adversity, in 2010, the IAC sugarcane breeding program implemented the first automated photoperiod chamber in Brazil (FAPESP-Fundação de Amparo à Pesquisa do Estado de São Paulo 2010a). This chamber is mainly used for energycane breeding (FAPESP-Fundação de Amparo à Pesquisa do Estado de São Paulo 2010b), and to our knowledge, it is still the only facility for this crop in Brazil. However, IAC crosses for sugarcane breeding mostly continue to be performed through natural induction of flowering.
Using a strategic approach, RIDESA constantly seeks to renew its germplasm collection through a recurrent selection process, with the annual inclusion of elite clones identified as performing well for key traits in advanced stages of selection as well as the introduction of domestic (other breeding institutions) and foreign clones. Historically, the network exchanges elite (protected) varieties with other Brazilian breeding programs. This strategy proved to be efficient because the kinship coefficient among the ten most planted varieties in Brazil has always remained relatively low since 1974 (Dal-Bianco et al. 2012). In recent years, the network has actively participated in variety exchange processes with several sugarcane-producing countries, such as Australia, Argentina, Colombia and Reunion Island, and more recently, new agreements were established with China, Ecuador, South Africa and the Philippines; approximately 60 varieties are exchanged annually. The main objective of this initiative is the introduction of new sources of variability for the different traits of agroindustrial importance and resistance to pests and diseases. In genetic terms, RIDESA constantly seeks to increase the germplasm collection to include new or increase the frequency of favorable alleles for continued improvement of varieties. The removal of unproductive parents is based on two main criteria: (1) the parental performance over family (progenies) trials, i.e., parents constantly contributing to low cane and sugar yield families and/or high incidence of diseases; (2) genotypes exhaustively used for crossing for RIDESA breeding programs. Currently, for some universities involved in RIDESA, the parental evaluation is fully based on family test results, where breeding values are estimated via a mixed models (REML/BLUP) procedure.
Various forms of crosses are performed for the production of progenies that can be selected for superior individuals. The most common are biparental crosses, polycrosses and free pollination (this strategy can be used due to the abundance of viable flowers). In RIDESA breeding programs, approximately 5000 crosses are performed annually. Of this, approximately half are performed in the Flowering and Crossing Station of Serra do Ouro, with the highest proportion of crosses corresponding to multiparental crosses (45%), followed by biparental crosses (31%), specific multiparental crosses (14%), free pollination (9%) and self-pollination (1%). The other half are performed by the Flowering and Crossing Station of Devaneio (08°19′ S, 35°24′ W, 514 m) (Fig. 4), with the following percentages: polycrosses (67%), biparental crosses (20%) and self-fertilization (13%). These numbers and proportions of crosses may vary according to the strategy of each breeding program. Among the 94 commercially released RB varieties, approximately half were generated by biparental crosses and the other half by polycrosses.
The correct choice of parents is one of the main challenges encountered by breeders because the total number of possible combinations is extremely high for a large number of parents, but due to technical and economic criteria, only a small part is used every year. The correct choice of parents ensures the genetic progress of future generations within breeding programs, and therefore, each institution has its own strategy. In RIDESA, the following concepts are considered when choosing parents:
-
1.
Behavior per se of the parents: Considering the presence of additive genetic effects for the most important economic traits, the use of elite clones assists in the process of obtaining superior varieties. All resulting varieties are then fed back into the germplasm collection in a recurrent process; therefore, the new accessions added to the germplasm collection are clones shown to be superior throughout the selection stages.
-
2.
Pedigree of the parent: This concept is widely used in animal breeding, emphasizing the importance of genetic diversity, which can be evaluated based on agronomic traits and/or the degree of kinship between genotypes. Crosses between distantly related genotypes are preferred.
-
3.
Combining ability: Estimating parameters such as general combining ability (GCA) and specific combining ability (SCA) are essential for the correct choice of parents. The disconnected factorial design has been widely used in some programs within the network, mostly with the formation of small groups of crossings, usually 3 × 3 or 4 × 4, aiming to maximize the number of parents to be evaluated in a single experiment (Burdon and Buijtenen 1990). In some programs, the use of reciprocal recurrent selection (RRS) schemes has also been considered to efficiently exploit both the nonadditive and additive effects involved in the expression of traits of economic importance. Preliminary studies have indicated promising results for establishing gene pools for the systematic exploration of heterosis (de Resende and Barbosa 2005; Mendes de Paula et al. 2020).
-
4.
History of the parent: An estimate of general combining ability can also be made based on the history of a parent in producing commercial cultivars. For example, RB72454 is considered to be a superior parent because it is the parent of 25 commercial varieties (Daros et al. 2015). Similarly, the cross between SP70-1143 and RB72454 generated five RB varieties of great importance and is considered a benchmark for high specific combining ability effects.
-
5.
Marker-assisted selection: The use of molecular markers for clone selection can be used to define crosses (parental selection). The best known, and to our knowledge the only, example for sugarcane is the gene Bru1, which is responsible for the control of brown rust resistance (Le Cunff et al. 2008; Costet et al. 2012). In this case, the crosses use parents, at least one of which has a resistance haplotype, to minimize the presence of genotypes susceptible to the disease in the population. We believe new results will be available in the future since several studies have been performed to characterize the genetic architecture of important sugarcane traits (see details in the molecular genetics and biotechnology section).
After generating variability, the following steps are established according to the strategy of each breeding program. At the Federal University of São Carlos (UFSCar), which operates in the main sugarcane-producing region of the country (Central-South) and is one of the ten federal universities that make up RIDESA, these stages are divided into progeny assessment trials (Phase 1), clonal assessment trials (Phases 2 and 3) and final assessment trials (Phase 4), as described below.
Phase 1: Currently, UFSCar and other universities in the network use a combined between- and within-family selection as a strategy in the initial breeding phase. According to previous studies, this methodology provides selection gains of approximately 11% over other methods (Cursi et al. 2019). This stage has the largest effective population size with approximately 200,000 seedlings produced by UFSCar alone and distributed to three different locations in the Central-South region of Brazil. Considering all the universities in the network, more than 3 million seedlings are planted annually throughout the country. These experiments are set up in an incomplete block design (unbalanced data), with the experimental unit consisting of two furrows that are 27 m in length and 1.5 m apart, with 108 seedlings per family (plot), spaced 0.5 m apart. In the plant cane stage (12-month-old cane), phenotypic data are collected at the family level for the sugarcane yield traits (tons of sugarcane per hectare—TCH), obtained through the full mechanized harvesting of each plot and weighed using a truck with a load cell. Due to the high number of clones at this phase and limited resources, the sugar content (Brix) is estimated by sampling ten stalks selected at random within each plot. In addition, incidence of diseased plants is assessed within each family. To minimize problems inherent to the initial phases, the mixed model approach is used to correct spatial variability, thus increasing the accuracy of the estimates (Gilmour et al. 1997). In the second year, selection occurs favoring the best families, i.e., the selection intensity is increased due to the smaller number of crosses involved. Families classified below average are not disregarded but rather subject to more intense selection.
Phase 2: In this stage, the number of individuals is reduced but is still a large sample that can range from 1000 to 3000 individuals, depending on the selection site of the initial phase. The selection intensity ranges from 1.5% (low-fertility soils) to 6% (high-fertility soils). The experimental design used is an augmented block design (Federer 1956), and the experimental unit consists of a single 5-m-long furrow, with only one replicate and the use of commercial varieties as controls. Morphological characteristics (conceptual grading scale), incidence of pests and diseases, and yield components are evaluated in both the plant cane and first-ratoon stages. Finally, clones with high potential are selected based on the sugarcane yield, second-ratoon sprouting and desirable morphological traits and disease resistance.
Phase 3: In this phase, around 200 clones are planted in a trial with a randomized block design, with individual plots consisting of two furrows that are 5 m in length and spaced 1.5 m apart, with two replicates. Approximately six different sites are considered within the states of São Paulo (SP) and Mato Grosso do Sul (MS), states that are affiliated with UFSCar, aiming at an initial identification of the interaction between genotype and environment. In addition to the desirable morphological and phytosanitary traits, the technological parameters are evaluated in plant cane and first-ratoon stages, and the clones with high potential in the second-ratoon stage are selected.
Phase 4: This phase is similar to Phase 3 but involves a smaller number of clones (approximately 25) such that it is possible to increase the number of observations (replicates) and evaluation sites. The experiment uses a randomized block design with a split plot design (different cuts). The experimental unit consists of four 12-m-long furrows (may vary with site), spaced 1.5 m apart, with four replicates. Approximately 10 different sites are considered among the states of SP and MS to capitalize on the identification of the interaction between genotype and environment and subsequent recommendations. Different phenotypic data are collected in the plant cane stage, first-ratoon stage, second-ratoon stage and, in some cases, until the fourth ratoon. For all phases, the harvests are performed mechanically, and the mixed model approach is considered for the best prediction of the genotypic values of each clone under experimentation. In addition, all promising clones, both in Phase 3 and Phase 4, are subjected to artificial and natural tests for resistance to the main diseases present in the country. Subsequently, validation steps are established with growers and partner mills for better dissemination and validation of the performance of potential clones for commercial release. Additionally, maturation curve experiments are conducted for a better understanding of the maturation process of the genotypes and, therefore, the correct recommendation of different harvesting times. It is important to note that all clones that show high potential in the final selection stages are distributed to the other RIDESA member universities, meaning that they are also tested in the different sugarcane regions of the country.
To complement the evaluation and selection stages, the supply of disease-free material for the establishment of nurseries, making the multiplication and commercial exploitation of new varieties more efficient, is a common objective among all breeding programs in Brazil. In 2019, UFSCar, in partnership with a company performing plant propagation from meristems (Explante Biotecnologia), started a project that delivered 2,794,000 healthy plantlets to 127 industries and growers associations in the Central-South region of Brazil. In 2020, it is estimated that 3,136,320 seedlings will be delivered to 132 units. In addition to the two initiatives, it is estimated that 5,930,320 meristem-derived seedlings will be supplied, without additional costs for the affiliated mills. Furthermore, the IAC Cane Program has disseminated methods to reduce the volume of seedlings necessary for the multiplication of new varietal technologies, with the objective of incorporating production gains through healthy seedlings. This method, called pre-sprouted seedling (PSS) (Landell et al. 2013), has been widely adopted by Brazilian growers and mills.
Notably, the breeding stages should be complemented with additional studies aiming to incorporate new knowledge and technologies to ensure genetic gains and maintain the continual commercial release of superior varieties. From this perspective, in recent years, RIDESA has invested in technologies to assist in the phenotyping process, i.e., inferring environmental variability to minimize residual effects (minimizing noise), e.g., using high-spectral-resolution field sensors (spectroradiometer), high-resolution proximal sensors (electromagnetic induction soil sensor) and multispectral subaerial sensors (UAVs, drones) (Natarajan et al. 2019). This information can be included in the statistical analyses, especially using the mixed model approach. Another newly developed technology is the use of a portable near-infrared spectrometer as a nondestructive, quick-read and environmentally friendly tool for the evaluation of the technological components of sugarcane, e.g., Pol, Brix and fiber, in the different breeding phases. Because these are new fields of study, the applicability of this type of technology is still ongoing, and it is currently in the development and validation phase.
Energycane
In the GranBio energycane improvement program, both the performance per se of the clone and the result of its progeny are considered for the selection of parents of a given population. A source of local selection can be found in the clonal selection phases. Early selection has been ongoing to reduce the recurrence time and, consequently, increase the generation gain. However, early selection involves an increase in the experimental error in assessments since the plots are smaller and are almost always in a single environment. Therefore, it is imperative to use progeny information as an auxiliary measure in the choice of parents and in clonal selection. If the progeny has a yield above the standard, the parent is identified as proven and used in a greater number of crosses. In addition, seeds remaining from the cross are used for germination and seedling utilization.
GranBio performs approximately 450 crosses annually, of which 300 are for obtaining Type II (F1 between commercial or pre-commercial hybrids crossed with Saccharum spontaneum genotypes from wild collection) energycane cultivars and 150 are for obtaining Type I (BC1 between F1 energycane types crossed with commercial or pre-commercial hybrids) energycane cultivars. Eighty percent of the crosses are exploratory (general combining ability), and 20% are considered proven (specific combining ability). The criteria for parental selection in energycane are the dry matter yield of the progenies both under GCA and SCA, associated with smut resistance, ratoon ability presence of rhizomes, no pithiness and absent of flowering. Yield of sugar per area which is the main issue in sugarcane is relevant only for the selection of energycane type 1, due to the commercial positioning of these type 1 varieties: poor environment of first-generation ethanol and sugar industry. For energycane type 1 selection, we add limits for fiber (lower than 35% up sugarcane standard) and sugars (higher than 75% of sugarcane standard).
Planting in Phase 1 is performed with the progenies grouped into families, and clonal selection is performed in a modified sequential manner (Bressiani et al. 2005). Families values from plant cane are used from parent selection and also for a ranking of the families to create the selection index, using families averages and genetic variations between and within families, that will suggest the number of individuals for the selection in first ratoon in each family. Individual selection at first ratoon is a visual selection taking into consideration number of stalks, stalk diameter, pithiness, flowering, disease resistance (smut, leaf scald, mosaic virus and rusts) and morphological aspects.
Phase 2 and in Phase 3, plots are planted in augmented blocs design (Federer) and selection is performed visually first for the same traits in stage 1 and then pre-selected individuals area submitted to biometrics and quality analysis in the laboratory at the plant cane and ratoon cane stages. Clones are confirmed to the next stage based on dry yield, fiber and sugar composition and grouped in type 1 and type 2 to continue the selection process.
In the experimental phase (Phase 4), a group of 20 clones plus 4 checks are planted in randomized block design experiments with three reps and five locations, and the plots are sampled for laboratory analysis, harvested manually or mechanically and weighed in their entirety. Tons of dry biomass, wet biomass, cane, fiber and sugars are measured. Also, disease and pests occurrence are quantified.
A summary of the selection steps of the GranBio improvement program is shown in Table 6.
Molecular Genetics and Biotechnology
Biotechnological approaches can be used to improve traditional breeding programs, mainly for understanding genetic structures, genomic locations and plant transformation. Due to the unusual complexity of the sugarcane genome, genomics studies and genetic manipulation are challenging tasks. However, efforts involving Brazilian germplasm have focused on the development of biotechnological approaches for this crop. The commercial sugarcane cultivar SP80-3280 has been used as a model for genomic, genetic and transgenic studies in Brazil, although other cultivars also have been studied.
Several molecular studies have elucidated aspects of sugarcane genome constitution and structure (Souza et al. 2011, 2019; de Setta et al. 2014). In this sense, the Brazilian Sugarcane Expressed Sequence Tags (ESTs) Sequencing Project (SUCEST) database is an important resource for managing sugarcane genome data and since 2003 has accumulated over 238,000 ESTs from different sugarcane tissues and cultivars grouped into approximately 43,000 sugarcane assembled sequences (SASs) (Vettore et al. 2003; Dal-Bianco et al. 2012). This database integrates the Sugarcane EST Project (SUCEST) (Vettore et al. 2003), the Sugarcane Gene Index (SGI), gene expression data (Papini-Terzi et al. 2009; Waclawovsky et al. 2010; Ferreira et al. 2016) and records of phenotypic data of the main Brazilian sugarcane cultivars (http://sucest-fun.org). Recently, Souza et al. (2019) and collaborators presented a gene space assembly for SP80-3280, including 373,869 putative genes and their potential regulatory regions, and showed that SP80-3280 has particular regulatory elements involved in sucrose synthesis not found in its ancestor Saccharum spontaneum.
Transcriptome and metabolomics projects have contributed to advances in the understanding of gene regulation systems of sugarcane, such as nitrogen supply metabolic pathways using RB975375 and RB937570 (Bassi et al. 2018), bud sprouting potential among 16 sugarcane Brazilian varieties (Ferreira et al. 2018) and drought responses using CTC15 and SP90-3414 (Budzinski et al. 2019). Several transcriptome analyses have been conducted considering Brazilian cultivars and with different purposes, such an RNA-Seq transcriptome profile for lignin content using IACSP04-065 and IACSP04-627 (Vicentini et al. 2015); leaf transcriptomes for six sugarcane genotypes (SP81-3250, RB925345, SP80-3280, RB835486, IACSP96-3046 and IACSP95-3018) involved in biparental crosses (Cardoso-Silva et al. 2014), transcription of circadian rhythm genes from different organs in field-grown SP80-3280 (Dantas et al. 2020); and ethylene-induced transcription responses at ripening (Cunha et al. 2017). Additionally, microtranscriptomes (miRNA expression patterns) (Ferreira et al. 2012; Gentile et al. 2013) have provided a working model of the defense strategies that might be regulated by miRNAs in sugarcane exposed to drought (Ferreira et al. 2017). These genes can be promising targets for genetic modification of the studied traits, although confirmation of their function is still needed. In these transcripts, SNP alleles can be found, which may be within the regulatory sequences that cause trait variation by modifying the expression levels of transcript. Such SNPs can be used as promising markers to direct crossings in the initial phases of breeding programs.
Understanding the molecular mechanisms of infection in sugarcane is important and can allow identifying molecular markers for disease resistance (dos Santos et al. 2017; Yang et al. 2018). An RNA-Seq approach was used to investigate the changes in temporal expression of transcripts by the sugarcane immune system when challenged by P. kuehnii. The authors revealed that P. kuehnii initially suppressed sugarcane genes involved in plant defense systems and that late overexpression of specific regulatory pathways suggested the possibility of an inefficient recognition system by a susceptible sugarcane genotype (Correr et al. 2020). Other pathosystems, such as sugarcane smut (Schaker et al. 2016; Peters et al. 2017) and sugarcane ratoon stunting (Cia et al. 2018), have also been studied. Moreover, molecular markers have been useful for molecular diagnosis in sugarcane, especially for Puccinia. The Bru1 frequency among Brazilian cultivars indicates that this gene is the prevalent source of brown rust resistance in Brazilian sugarcane breeding programs and could be used to predict resistant phenotypes (Barreto et al. 2017; Neuber et al. 2017). Additionally, molecular markers associated with resistance to orange rust (e.g., G1 marker, Yang et al. (2018)) have been evaluated as molecular tools that can be used by Brazilian breeding programs in the search for resistant sugarcane cultivars (Fier et al. 2020).
In recent years, numerous initiatives to produce genetically modified (GM) sugarcane have been put into place in Brazil. Genetic transformation mediated by Agrobacterium and biobalistic systems has been the most commonly adopted method for generating transgenic events. In proof-of-concept experiments, variety SP80-3280 has usually been used because the protocols of in vitro regeneration are well described. For GM sugarcane, several traits are being studied, e.g., herbicide tolerance; pest resistance (cry genes); resistance to abiotic stresses, such as drought (higher accumulation of proline and trehalose); accumulation of sugar and biomass (Falco et al. 2000; Molinari et al. 2007, 2019; Ramiro et al. 2016; Ferreira et al. 2017; Cristofoletti et al. 2018; de Souza et al. 2019). In late 2017, CTNBio, the Brazilian government regulatory authority responsible for the approval of biotechnology-derived products, approved the first GM sugarcane, Cry1Ab-expressing sugarcane, for cultivation in Brazil (Velini 2017). Developed by CTC, this transgenic commercial event was obtained by bombarding embryogenic cells (callus) of the immature inflorescences of sugarcane cultivar CTC20 with microprojectiles containing purified CTC2.nptII DNA fragments from the pGH-CTC2.nptII vector, which contains the cry1Ab and nptII genes, generating insect-resistant sugarcane. Currently, there are at least three commercially approved GM sugarcane varieties in Brazil, all of which are insect resistant (Bt technology: Cry1Ab and Cry1Ac gene). Cry1Ab and Cry1Ac proteins target receptors found only in Lepidoptera, causing selective toxicity. These proteins have proved very effective, as their toxicity is specific, and can thus be used to target specific lepidopteran pests (Kennedy et al. 2018). Few studies have been carried out with genome editing technology, but some have focused on sugarcane genome editing using CRISPR/Cas9 for targeted mutagenesis of ethylene biosynthesis.
Molecular markers have been used in genetic diversity studies, cultivar identification and genetic mapping (de Morais et al. 2015). The markers commonly used in sugarcane, such AFLP, TRAP, SSRs and SNPs, are able to differentiate cultivars and enable cost-effect mapping studies. The genotyping costs depend on several factors, including the nature of the study, number of genotypes, the type and density of markers, the required coverage of the genome, the time of analysis, and the level of knowledge and experience of workers (Puckett 2017; Rasheed et al. 2017). For sugarcane, the genotyping cost per sample can range between US$50.00 and US$80.00 for sequencing-based markers and between US$5.00 and US$20.00 for gel-based markers. These genotyping values should be considered carefully as they are determined by sample and not by data point. In the first case, hundreds to thousands of markers can be obtained per sample, while in the second case, the number of markers is much smaller, in the tens if multiplexing is used (Slater et al. 2013; Thomson 2014; Puckett 2017; Rasheed et al. 2017; Ahmad et al. 2018). In Brazil, the use of molecular markers has advanced over time, and some approaches are employed by sugarcane breeding programs. As discussed below, genetic diversity studies to support the choice of parents and introgression of different genetic backgrounds and biparental and associative mapping studies with the objective of identifying genomic regions have been performed to further assist in the selection and development of diagnostic markers.
Diversity studies have been carried out with Brazilian and foreign germplasms using several types of molecular markers (Creste et al. 2010; Garcia et al. 2013; Manechini et al. 2018). Recently, Medeiros et al. (2020) evaluated the expanded Brazilian Panel of Sugarcane Genotypes (BPSG), composed of 254 accessions of the Saccharum complex, with eight TRAP markers anchored in sucrose and lignin metabolism genes. The authors suggest that even for genes under selection processes, such as those involved in sucrose and lignin, there is still a possibility of the introgression of new and favorable alleles. This further development is possible because these genes showed, through TRAP markers, sufficient variability to separate groups of accessions and to identify exclusive fragments. Therefore, the TRAP markers could be used to evaluate foreign accessions in a targeted manner, opening a path to germplasm exchange and assisted selection with functional molecular markers. In addition, Kanthack Junior et al. (2020) evaluated the genetic variability of 96 sugarcane genotypes through TRAP markers derived from candidate genes involved in sugar and lignin metabolism. The authors suggest that not only S. officinarum but also S. spontaneum made significant contributions to the genetic variability of genes involved in sugar metabolism of Brazilian commercial sugarcane cultivars and that considerable variability in lignin metabolism genes remains underexplored by sugarcane breeding programs. Diversity studies have also enabled a practical application of molecular markers to confirm the genetic fidelity of the sugarcane seedlings produced or resolve doubts about a variety cultivated in the field (Manechini et al. 2018).
Sugarcane genetic maps and mapping studies using Brazilian clones or cultivars were initiated by Garcia et al. (2006), who developed the first integrated genetic map from a cross of two precommercial cultivars, SP80-180 x SP80-4966. Subsequently, Oliveira et al. (2007) added to the pre-existing genetic map molecular markers developed from expressed sequences, namely EST-SSRs and EST-RFLPs. Additionally, quantitative trait locus (QTL) mapping studies were performed with progeny of a cross between SP80-180 and SP80-4966. Pinto et al. (2010) evaluated fiber percent, cane yield (TCH), sucrose content (Pol) and tonnes of sugar per hectare (TSH) and detected 120 marker-trait associations (MTAs). Pastina et al. (2012) identified 46 QTLs (13 for TCH, 14 for TSH, 11 for fiber percent and eight for Pol) through a strategy for QTL detection in multiharvest-location trial data, based on interval mapping and mixed models. Furthermore, Margarido et al. (2015) evaluated fiber content, Pol and TCH and detected QTLs using multitrait multienvironment mixed models with a multiple QTL mapping approach. A total of 13 QTLs exhibiting QTLs by location, QTLs by harvest or a three-way interaction were found. Similarly, Palhares et al. (2012) constructed a genetic map for a population derived from the cross between cultivars IAC66-6 and TUC71-7. This map was constructed with AFLP markers, EST-SSRs and scIvana_1-based markers. scIvana_1 is an LTR retrotransposon family member of the Copia superfamily. Two other studies used a population that originated from the cross between IACSP95-3018 and IACSP93-3046. The first, by Santos et al. (2015), evaluated MTAs and epistasis for brown rust resistance using SSR and AFLP markers, and the second, by Costa et al. (2016), was the first genetic map in which SNP markers segregated in a 1:2:1 fashion, and therefore, codominant, were included together with information about ploidy level. Associative mapping has also been performed; Barreto et al. (2019) used a mini core collection of sugarcane composed of 134 accessions from the BPSG to perform a genome-wide association study (GWAS) for Brix, SH, stalk number (SN), SW, and TCH using 100 SSR markers. Four subpopulations were detected, and linkage disequilibrium was stronger in the first 15 cM and present in a large extension. The GWAS detected MTAs for all traits. The four MTAs with the highest percentages of explained phenotypic variation indicate that the presence of at least one copy of the allele could also be important for driving strategies in breeding programs. Therefore, these MTAs should be validated as an initial approach to support breeding programs with introgression or selection processes. In addition, new methods of analysis are still in development to increase the understanding of complex genomes and enable mapping and association studies with further levels of allelic information. The BPSG has been expanded and is evaluating SNP markers with single and multiple dosage information. These results showed the advances in molecular studies of sugarcane in Brazil, including different molecular markers, populations with different genetic backgrounds and analysis approaches. Furthermore, this timeline demonstrates the intention of breeding programs to include molecular information in the process of selection and/or choice of parents for crosses. A successful case, already reported in breeding programs around the world, is the use of the marker associated with brown rust resistance gene, Bru1, as an auxiliary tool in the selection of resistant genotypes (Barreto et al. 2017).
Despite this progress, the genetic complexity of sugarcane continues to challenge the integration between quantitative traits and molecular data. Thus, the incorporation of new technologies is important to decrease this gap (Rasheed et al. 2017). New technologies for the high-throughput genotyping of sugarcane populations were used by Balsalobre et al. (2017), who reported, for the first time, the development and application of genotyping-by-sequencing (GBS) for mapping studies in sugarcane. Single-dose SNP results from GBS data analysis were combined with SSR and TRAP markers to construct an integrated genetic map of the population that originated from the cross between SP80-3280 and RB835486. The map length was 3682.04 cM, with an average marker density of 3.70 cM, the highest density for sugarcane genetic maps to date. The QTL map for Pol, Brix, SD and FIB identified seven QTLs. In addition, QTLs for Brix and FIB traits had markers linked to candidate genes that could be validated in other populations. Another initiative to use high-throughput genotyping was carried out by Brum (2018), who used SNPs markers and evaluated the efficiency of genomic selection in the CTC breeding program. Clones of two breeding cycles were genotyped, and plot weight, Brix, fiber and sucrose content were phenotyped. The genomic prediction accuracy of the model ranged from 0.07 to 0.39 in cross-validation within a breeding cycle and from 0.01 to 0.32 in predictions across cycles, showing that overall the fiber content exhibited the highest values. A strong genotype by year interaction was observed. Furthermore, the association analysis and genomic prediction showed advantages when high dosage of alleles were considered. Therefore, MAS and genomic selection have advanced in Brazil, primarily genotyping and multiple dose analysis approaches. Improving the phenotyping of quantitative traits for a large number of genotypes will be essential to allow the efficient use of genomic selection in breeding programs.
Conclusions
In the last 40 years, Brazil has increased sugar production by 155.7 kg ha−1 per year. In addition, the country has become a world leader in the production of byproducts, such as biomass used for the cogeneration of electricity and bioethanol used as fuel for vehicles. In addition, in this period, the frontiers of cultivation of sugarcane were expanded, achieving an approximate eightfold increase in area. Despite this expansion, the main sugarcane-growing regions in the country remain hundreds of kilometers away from the Amazon rainforest edges (Fig. 4). However, most expansions occurred in “nontraditional” regions, mostly those with low-fertility soils and low and/or irregular rainfall, thus challenging the resilience of this crop; in association with new diseases, accelerated adoption of mechanical harvesting and less investment in sugarcane field a nonsignificant change in yield can be observed in the last decade (Fig. 2). With the expansion of industrial parks, the demand for raw material increased, as did that for an expanded crop season, which was achieved by Brazilian breeding efforts. In addition, more than 200 high-yield varieties adapted to different production environments and exhibiting longevity, disease resistance and mechanization adaptation were developed and made available to the industry. Despite great advances, the national average production is still far below the theoretical potential of the crop and lower than that of some other countries. Conventional sugarcane breeding in Brazil has consistently been a major contributor to the agroindustrial yield improvement in the country and provided security against pest and diseases outbreaks. However, due to the high genetic complexity of the crop, the low narrow-sense heritability of most traits of economic importance and the crop’s long breeding cycle, detailed knowledge of quantitative genetics and possibly new innovative breeding strategies are required to continue advance breeding. The emergence of new technologies has enabled a deeper understanding of genetic and environmental factors and consequently a more accurate understanding of the architecture of additive and non-additive genetic components. In addition, the use of biotechnology in breeding platforms, driven by recent advances in genomic resources in sugarcane, is already a reality in the search for genetic progress in traditional breeding programs in Brazil. For example, GM cultivars with insect tolerance are already marketed; however, difficulties remain in obtaining sugarcane bred for complex characteristics, e.g., higher sucrose and dry matter content. Furthermore, pre-breeding activities remain a key factor in the expansion of genetic diversity, especially for alleles conferring resistance to pests and diseases, and in the search for allelic complementation for some traits, such as sugar content. Measures for strengthening and ensuring the continuity of germplasm exchange among the breeding programs of the world will be essential to expand the genetic base of breeders’ working collections without total dependence on the variability present in ancestral species. Introgression processes should be used predominantly to compose the gene pool of the energycane ideotype. Furthermore, multidisciplinary approaches and the development of trained teams will be key factors to ensure genetic progress in sugarcane crop production.
References
Ahmad, Ali, Jin-Da Wang, Yong-Bao Pan, Rahat Sharif, and San-Ji Gao. 2018. Development and use of simple sequence repeats (SSRs) markers for sugarcane breeding and genetic studies. Agronomy 8: 260.
Andrade, Jose Clovis. 1985. Escorço histórico de antigas variedades de cana-de-açúcar. Maceió: ASPLANA.
Antunes, Felipe A.F., Anuj K. Chandel, Ruly Terán-Hilares, Thais S.S. Milessi, Beatriz M. Travalia, Felipe A. Ferrari, Andrés F. Hernandez-Pérez, Lucas Ramos, Paulo F. Marcelino, Larissa P. Brumano, Gilda M. Silva, Marcus B.S. Forte, Júlio C. Santos, Maria G.A. Felipe, and Silvio S. da Silva. 2019. Biofuel production from sugarcane in Brazil. In Sugarcane biofuels: Status, potential, and prospects of the sweet crop to fuel the world, eds. Muhammad Tahir Khan, and Imtiaz Ahmed Khan, 99–121. Cham: Springer.
Balsalobre, Thiago W.A., Melina C. Mancini, Guilherme da S. Pereira, Carina O. Anoni, Fernanda Z. Barreto, Hermann Paulo Hoffmann, Anete P. de Souza, Antonio A.F. Garcia, and Monalisa S. Carneiro. 2016. Mixed modeling of yield components and brown rust resistance in sugarcane families. Agronomy Journal 108: 1824–1837.
Balsalobre, Thiago Willian Almeida, Guilherme da Silva Pereira, Gabriel Rodrigues Alves Margarido, Rodrigo Gazaffi, Fernanda Zatti Barreto, Carina Oliveira Anoni, Cláudio Benício Cardoso-Silva, Estela Araújo Costa, Melina Cristina Mancini, Hermann Paulo Hoffmann, Anete Pereira de Souza, Antonio Augusto Franco Garcia, and Monalisa Sampaio Carneiro. 2017. GBS-based single dosage markers for linkage and QTL mapping allow gene mining for yield-related traits in sugarcane. BMC Genomics 18: 72.
Barbosa, Geraldo Veríssimo de Souza. 2018. Inovações tecnológicas desenvolvidas em Alagoas para o setor canavieiro. Maceió: FAPEAL/Imprensa Oficial Graciliano Ramos.
Barbosa, Márcio Henrique Pereira, Marcos Deon Vilela Resende, Luiz Antônio dos Santos Dias, Geraldo Veríssimo de Souza Barbosa, Ricardo Augusto de Oliveira, Luiz Alexandre Peternelli, and Edelclaiton Daros. 2012. Genetic improvement of sugar cane for bioenergy: The Brazilian experience in network research with RIDESA. Crop Breeding and Applied Biotechnology 12: 87–98.
Barreto, Fernanda Zatti, João Ricardo Bachega Feijó Rosa, Thiago Willian Almeida Balsalobre, Maria Marta Pastina, Renato Rodrigues Silva, Hermann Paulo Hoffmann, Anete Pereira de Souza, Antonio Augusto Franco Garcia, and Monalisa Sampaio Carneiro. 2019. A genome-wide association study identified loci for yield component traits in sugarcane (Saccharum spp.). PLoS ONE 14: e0219843.
Barreto, Fernanda Zatti, Thiago Willian Almeida. Balsalobre, Roberto Giacomini Chapola, Hermann Paulo Hoffmann, and Monalisa Sampaio Carneiro. 2017. Validação de marcadores moleculares associados à resistência à ferrugem marrom em cana-de-açúcar. Summa Phytopathologica 43: 36–40.
Bassi, Denis, Marcelo Menossi, and Lucia Mattiello. 2018. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Scientific Reports 8: 2327.
Berding, Nils, and Brain T. Roach. 1987. Germplasm collection, maintenance, and use. In Sugarcane improvement through breeding, ed. Don J. Heinz, 143–210. Amsterdam: Elsevier.
Bordonal, Ricardo de Oliveira, João Luís Nunes Carvalho, Rattan Lal, Eduardo Barretto de Figueiredo, Bruna Gonçalves de Oliveira, and Newton La Scala. 2018. Sustainability of sugarcane production in Brazil. A review. Agronomy for Sustainable Development 38: 13.
Braga Júnior, Rubens Leite do Canto, Marcos Guimarães de Andrade Landell, Daniel Nunes da Silva, Márcio Aurélio Pitta Bidóia, Thiago Nogueira da Silva, José Roberto Thomazinho Júnior, Victor Hugo Palverqueires da Silva, and Ivan Antônio dos Anjos. 2019. Censo varietal IAC de cana-de-açúcar no Brasil - safra 2017/18 e na região Centro-Sul - safra 2018/19. Campinas: Instituto Agronômico.
Braga Júnior, Rubens Leite do Canto, I.A. Oliveira, and J.A. Raizer. 2011. Evolução das áreas cultivadas com variedades de cana-de-açúcar no Brasil nos últimos vinte anos. STAB Açúcar, Álcool e Subprodutos, Piracicaba 30: 46–50.
Bressiani, J.A., R. Vencovsky, and W.L. Burnquist. 2005. Modified sequential selection in sugarcane. Proceedings of International Society of Sugarcane Techologists 25: 459–467.
Brum, I.J.B. 2018. Performance of genomic prediction for a sugarcane commercial breeding program. Doctoral dissertation, Cornell University, USA.
Budzinski, Ilara Gabriela, Fabricio Edgar Frasson, Thais de Moraes, Regiani Cataldi, Lívia Maria Franceschini, and Carlos Alberto Labate. 2019. Network analyses and data integration of proteomics and metabolomics from leaves of two contrasting varieties of sugarcane in response to drought. Frontiers in Plant Science 10: 1524.
Burdon, Rowland David, and J.P. Van Buijtenen. 1990. Expected efficiencies of mating designs for reselection of parents. Canadian Journal of Forest Research 20: 1664–1671.
Cardoso-Silva, Claudio Benicio, Estela Araujo Costa, Melina Cristina Mancini, Thiago Willian Almeida Balsalobre, Lucas Eduardo Costa Canesin, Luciana Rossini Pinto, Monalisa Sampaio Carneiro, Antonio Augusto Franco Garcia, Anete Pereira de Souza, and Renato Vicentini. 2014. De novo assembly and transcriptome analysis of contrasting sugarcane varieties. PLoS ONE 9: e88462.
Carvalho-Netto, Osmar V., Jose A. Bressiani, Hugo L. Soriano, Celso S. Fiori, Joao M. Santos, Geraldo V.S. Barbosa, Mauro A. Xavier, Marcos G.A. Landell, and Goncalo A.G. Pereira. 2014. The potential of the energy cane as the main biomass crop for the cellulosic industry. Chemical and Biological Technologies in Agriculture 1: 1–8.
Cesnik, Roberto, and Jacques Miocque. 2004. Melhoramento da cana-de-açúcar, 307. Brasília: Embrapa Informação Tecnológica.
Cheavegatti-Gianotto, A., et al. 2019. Development of CTC20BT, the first genetically modified sugarcane commercially available in the world. Proceedings International Society of Sugar Cane Technologists Congress 30: 1272–1279.
Cia, Mariana Cicarelli, Giselle de Carvalho, Ricardo Antunes Azevedo, Claudia Barros Monteiro-Vitorello, Glaucia Mendes Souza, Milton Yutaka Nishiyama-Junior, Carolina Gimiliani Lembke, Raphael Severo da Cunha Antunes de Faria, João Paulo Rodrigues Marques, and Maeli Melotto. 2018. Novel insights into the early stages of ratoon stunting disease of sugarcane inferred from transcript and protein analysis. Phytopathology 108: 1455–1466.
CONAB-Companhia Nacional de Abastecimento. 2020. Acompanhamento da safra brasileira de cana-de-açúcar, safra 2019/20, quarto levantamento. https://www.conab.gov.br/info-agro/safras. Accessed 14 May 2020.
CONSECANA-Conselho dos Produtores de Cana-de-Açúcar Açúcar e Álcool do Estado de São Paulo-SP. 2006. Manual de instruções. Piracicaba, Brazil: CONSECANA.
Correr, Fernando Henrique, Guilherme Kenichi Hosaka, Sergio Gregorio Pérez Gómez, Mariana Cicarelli Cia, Claudia Barros Monteiro Vitorello, Luis Eduardo Aranha Camargo, Nelson Sidnei Massola, Monalisa Sampaio Carneiro, and Gabriel Rodrigues Alves Margarido. 2020. Time-series expression profiling of sugarcane leaves infected with Puccinia kuehnii reveals an ineffective defense system leading to susceptibility. Plant Cell Reports 39: 873–889.
Cortez, Luis A.B., and Ricardo Baldassin, Jr. 2016. Polices towards bioethanol and their implications: Case Brazil. In Bioetanol de cana-de-açúcar – P&D para produtividade e sustentabilidade, ed. Luis A.B. Cortez, 142–162. São Paulo, Brazil: Blucher.
Costa, E.A., C.O. Anoni, M.C. Mancini, F.R.C. Santos, T.G. Marconi, R. Gazaffi, M.M. Pastina, D. Perecin, M. Mollinari, and M.A. Xavier. 2016. QTL mapping including codominant SNP markers with ploidy level information in a sugarcane progeny. Euphytica 211: 1–16.
Costet, L., L. Le Cunff, S. Royaert, L.M. Raboin, C. Hervouet, L. Toubi, H. Telismart, O. Garsmeur, Y. Rousselle, J. Pauquet, S. Nibouche, J.C. Glaszmann, J.Y. Hoarau, and A. D’Hont. 2012. Haplotype structure around Bru1 reveals a narrow genetic basis for brown rust resistance in modern sugarcane cultivars. Theoretical and Applied Genetics 125: 825–836.
Creste, S., D.M. Sansoli, A.C.S. Tardiani, D.N. Silva, F.K. Gonçalves, T.M. Fávero, C.N.F. Medeiros, C.S. Festucci, L.A. Carlini-Garcia, and M.G.A. Landell. 2010. Comparison of AFLP, TRAP and SSRs in the estimation of genetic relationships in sugarcane. Sugar Tech 12: 150–154.
Cristofoletti, Plinio T., Edson L. Kemper, Adriana N. Capella, Sandra R. Carmago, Juliana L. Cazoto, Fernanda Ferrari, Tederson L. Galvan, Luciane Gauer, Gustavo A. Monge, Marcelo A. Nishikawa, Nicolas Z. Santos, Altair A. Semeao, Leandro Silva, Alan R. Willse, Almir Zanca, and Michael D. Edgerton. 2018. Development of transgenic sugarcane resistant to sugarcane borer. Tropical Plant Biology 11: 17–30.
Cunha, Camila P., Guilherme G. Roberto, Renato Vicentini, Carolina G. Lembke, Glaucia M. Souza, Rafael V. Ribeiro, Eduardo C. Machado, Ana M.M.A. Lagôa, and Marcelo Menossi. 2017. Ethylene-induced transcriptional and hormonal responses at the onset of sugarcane ripening. Scientific Reports 7: 43364.
Cursi, Danilo Eduardo, Rodrigo Gazaffi, Hermann Paulo Hoffmann, Roberto Giacomini Chapola, A.R. Fernandes Júnior, M.S. Carneiro, and Antonio Augusto F. Garcia. 2019. Genetic gain prediction by different family-selection strategies. In Proceedings of the international society of sugar cane technologists, 693–697. Tucumán: ISSCT.
Cursi, Danilo Eduardo, Hermann Paulo Hofmann, Roberto Giacomini Chapola, A.R. Fernandes Júnior, Rodrigo Gazaffi, and Antonio Augusto F. Garcia. 2016. Análise de trilha visando seleção de genótipos com elevado rendimento de sacarose em fases iniciais do melhoramento da cana-de-açúcar. In 10° Congresso nacional da sociedade dos técnicos açucareiros e alcooleiros do Brasil - STAB, 21–26. Ribeirão Preto: PMGCA.
Dal-Bianco, Maximiller, Monalisa Sampaio Carneiro, Carlos Takeshi Hotta, Roberto Giacomini Chapola, Hermann Paulo Hoffmann, Antonio Augusto Franco Garcia, and Glaucia Mendes Souza. 2012. Sugarcane improvement: How far can we go? Current Opinion in Biotechnology 23: 265–270.
Dantas, Luíza Lane de Barros, Felipe Marcelo Almeida-Jesus, Natalia Oliveira de Lima, Cícero Alves-Lima, Milton Yutaka Nishiyama-Jr, Monalisa Sampaio Carneiro, Glaucia Mendes Souza, and Carlos Takeshi Hotta. 2020. Rhythms of transcription in field-grown sugarcane are highly organ specific. Scientific Reports 10: 6565.
Daros, Edelclaiton, Bruna Gonçalves de Oliveira, and Geraldo Veríssimo de Souza Barbosa. 2015. 45 anos de variedades RB de cana-de-açúcar: 25 anos de Ridesa. Curitiba: Graciosa.
de Morais, Lizz Kezzy, Marcelo Sfeir de Aguiar, Paulo de Albuquerque e Silva, Tassiano Maxuell Marinho Câmara, Danilo Eduardo Cursi, Antônio Ribeiro Fernandes Júnior, Roberto Giacomini Chapola, Monalisa Sampaio Carneiro, and João Carlos Bespalhok Filho. 2015. Breeding of sugarcane. In Industrial crops: Breeding for bioenergy and bioproducts, eds. Von Mark V. Cruz, and David A. Dierig, 29–42. New York: Springer.
de Resende, Marcos Deon Vilela, and Márcio Henrique Pereira Barbosa. 2005. Melhoramento genético de plantas de propagação assexuada. Colombo: Embrapa Informação Tecnológica.
de Setta, Nathalia, Cláudia Barros Monteiro-Vitorello, Cushla Jane Metcalfe, Guilherme Marcelo Queiroga Cruz, Luiz Eduardo Del Bem, Renato Vicentini, Fábio Tebaldi Silveira Nogueira, Roberta Alvares Campos, Sideny Lima Nunes, Paula Cristina Gasperazzo Turrini, Andreia Prata Vieira, Edgar Andrés Ochoa Cruz, Tatiana Caroline Silveira Corrêa, Carlos Takeshi Hotta, Alessandro de Mello Varani, Sonia Vautrin, Adilson Silva da Trindade, Mariane de Mendonça Vilela, Carolina Gimiliani Lembke, Paloma Mieko Sato, Rodrigo Fandino de Andrade, Milton Yutaka Nishiyama, Jr., Claudio Benicio Cardoso-Silva, Katia Castanho Scortecci, Antônio Augusto Franco Garcia, Monalisa Sampaio Carneiro, Changsoo Kim, Andrew H. Paterson, Hélène Bergès, Angélique D'Hont, Anete Pereira de Souza, Glaucia Mendes Souza, Michel Vincentz, João Paulo Kitajima, and Marie-Anne Van Sluys. 2014. Building the sugarcane genome for biotechnology and identifying evolutionary trends. BMC Genomics 15: 540.
de Souza, Wagner Rodrigo, Nelson Geraldo de Oliveira, Felipe Vinecky, Ana Paula Ribeiro, Marcos Fernando Basso, Raphael Augusto das Chagas Noqueli Casari, Bárbara Andrade Dias Brito da Cunha, Karoline Estefani Duarte, Thaís Ribeiro Santiago, Polyana Kelly Martins, Carlos Eduardo Aucique-Perez, Sílvio Carlos Cristofoletti Júnior, Alexandre Lima Nepomuceno, Carlos Antônio Ferreira de Sousa, Adilson Kenji Kobayashi, Kazuo Nakashima, Kazuko Yamaguchi-Shinozaki, and Hugo Bruno Correa Molinari. 2019. Field evaluation of AtDREB2A CA overexpressing sugarcane for drought tolerance. Journal of Agronomy and Crop Science 205: 545–553.
dos Santos, Fernanda RC., Maria I. Zucchi, Jong-Won. Park, Thiago R. Benatti, Jorge A. da Silva, Glaucia M. Souza, Marcos G.A. Landell, and Luciana R. Pinto. 2017. New sugarcane microsatellites and target region amplification polymorphism primers designed from candidate genes related to disease resistance. Sugar Tech 19: 219–224.
Falco, M.C., A. Tulmann Neto, and E.C. Ulian. 2000. Transformation and expression of a gene for herbicide resistance in a Brazilian sugarcane. Plant Cell Reports 19: 1188–1194.
FAO-Food and Agriculture Organization of the United Nations. 2020. Base de dados Faostat—Agriculture. http://www.fao.org/faostat/en/#rankings/countries_by_commodity. Accessed 15 May 2020.
FAPESP-Fundação de Amparo à Pesquisa do Estado de São Paulo. 2010a. Câmara de fotoperíodo possibilitará cruzamento de variedades. https://namidia.fapesp.br/camara-de-fotoperiodo-possibilitara-cruzamento-de-variedades/39639. Accessed 17 May 2020.
FAPESP-Fundação de Amparo à Pesquisa do Estado de São Paulo. 2010b. Câmara simula clima para cana. https://namidia.fapesp.br/camara-simula-clima-para-cana/35488. Accessed 14 September 2020.
Federer, Walter T. 1956. Augmented (hoonuiaku) designs. Hawaiiwan Planter’s Record 55: 191–208.
Fernandes, A.C., and S.A Tatizana. 1991. Retorno econômico do programa de melhoramento de cana-de-açúcar da COPERSUCAR nas safras 86/87 a 90/91. In V seminário de tecnologia agronômica, 79–95. Piracicaba: COPERSUCAR.
Ferreira, Thaís, Agustina Gentile, Romel Vilela, Gustavo Costa, Lara Dias, Laurício Endres, and Marcelo Menossi. 2012. microRNAs associated with drought response in the bioenergy crop sugarcane (Saccharum spp.). PLoS ONE 7: e46703.
Ferreira, Savio Siqueira, Carlos Takeshi Hotta, Viviane Guzzo de Carli Poelking, Debora Chaves Coelho Leite, Marcos Silveira Buckeridge, Marcelo Ehlers Loureiro, Marcio Henrique Pereira Barbosa, Monalisa Sampaio Carneiro, and Glaucia Mendes Souza. 2016. Co-expression network analysis reveals transcription factors associated to cell wall biosynthesis in sugarcane. Plant Molecular Biology 91: 15–35.
Ferreira, Danilo A., Marina C.M. Martins, Adriana Cheavegatti-Gianotto, Monalisa S. Carneiro, Rodrigo R. Amadeu, Juliana A. Aricetti, Lucia D. Wolf, Hermann Paulo Hoffmann, Luis G.F. de Abreu, and Camila Caldana. 2018. Metabolite profiles of sugarcane culm reveal the relationship among metabolism and axillary bud outgrowth in genetically related sugarcane commercial cultivars. Frontiers in Plant Science 9: 857.
Ferreira, Thais H.S., Max S. Tsunada, Denis Bassi, Pedro Araújo, Lucia Mattiello, Giovanna V. Guidelli, Germanna L. Righetto, Vanessa R. Gonçalves, Prakash Lakshmanan, and Marcelo Menossi. 2017. Sugarcane water stress tolerance mechanisms and its implications on developing biotechnology solutions. Frontiers in Plant Science 8: 1077.
Fier, Ícaro, Thiago Willian Almeida Balsalobre, Roberto Giacomini Chapola, Hermann Paulo Hoffmann, and Monalisa Sampaio Carneiro. 2020. Field resistance and molecular detection of the orange rust resistance gene linked to G1 marker in Brazilian cultivars of sugarcane. Summa Phytopathologica 46: 92–97.
Garcia, Antonio Augusto F., Ederson Akio Kido, A.N. Meza, Henrique Marques-Souza, Luciana Rossini Pinto, Maria Marta Pastina, C.S. Leite, Jorge Alberto da Silva, Eugenio C. Ulian, Antonio Figueira, Anete Pereira de Souza. 2006. Development of an integrated genetic map of a sugarcane (Saccharum spp.) commercial cross, based on a maximum-likelihood approach for estimation of linkage and linkage phases. Theoretical and Applied Genetics 112: 298–314.
Garcia, Antonio A.F., Marcelo Mollinari, Thiago G. Marconi, Oliver R. Serang, Renato R. Silva, Maria L.C. Vieira, Renato Vicentini, Estela A. Costa, Melina C. Mancini, and Melissa O.S. Garcia. 2013. SNP genotyping allows an in-depth characterisation of the genome of sugarcane and other complex autopolyploids. Scientific Reports 3: 3399.
Gazaffi, Rodrigo, Danilo Eduardo Cursi, Roberto Giacomini Chapola, João Messias dos Santos, Antonio Ribeiro Fernandes Júnior, Monalisa Sampaio Carneiro, Geraldo Veríssimo de Souza Barbosa, and Hermann Paulo Hoffmann. 2016. RB varieties: A major contribution to the sugarcane industry in Brazil. In Proceedings of the international society of sugar cane technologists, 1677–1682. Chiang Mai: ISSCT.
Gazaffi, Rodrigo, Karine Miranda Oliveira, Anete Pereira de Souza, and Antonio Augusto Franco Garcia. 2010. Sugarcane: Breeding methods and genetic mapping. In Bioetanol de cana-de-açúcar – P&D para produtividade e sustentabilidade, ed. Luis A.B. Cortez, 333–352. São Paulo: Blucher.
Gentile, Agustina, Thaís H. Ferreira, Raphael S. Mattos, Lara I. Dias, Andrea A. Hoshino, Monalisa S. Carneiro, Glaucia M. Souza, Tercílio Calsa, Rejane M. Nogueira, and Laurício Endres. 2013. Effects of drought on the microtranscriptome of field-grown sugarcane plants. Planta 237: 783–798.
Gilmour, Arthur R., Brian R. Cullis, Ar.ūnas P. Verbyla, and Arunas P. Verbyla. 1997. Accounting for natural and extraneous variation in the analysis of field experiments. Journal of Agricultural, Biological, and Environmental Statistics 2: 269–293.
Hoffmann, Hermann Paulo, A.L. Francelli, Sizuo Matsuoka, and Y. Masuda. 1999. Contribuição de variedades de cana-de-açúcar no Estado de São Paulo, nos últimos cinquenta anos. In Congresso nacional da STAB, 49–52. Londrina: STAB.
IAA. 1972. BRASIL/AÇÚCAR. Rio de Janeiro: MIC/IAA.
Instituto Agronômico de Campinas. 2020. Centro de Cana. Campinas. http://www.iac.sp.gov.br/areasdepesquisa/cana/. Accessed 5 May 2020.
Junior, Kanthack, Carlos Augusto Dias, João Ricardo Vieira Manechini, Ronan Xavier Corrêa, Ana Christina Rossini Pinto, Juliana Borges, Thais da Costa, Monteiro Favero, and Luciana Rossini Pinto. 2020. Genetic structure analysis in sugarcane (Saccharum spp.) using target region amplification polymorphism (TRAP) markers based on sugar-and lignin-related genes and potential application in core collection development. Sugar Tech 22: 641–654.
Kennedy, Reese D., Adriana Cheavegatti-Gianotto, Wladecir S. de Oliveira, Ronald P. Lirette, and Jerry J. Hjelle. 2018. A general safety assessment for purified food ingredients derived from biotechnology crops: Case study of Brazilian sugar and beverages produced from insect-protected sugarcane. Frontiers in Bioengineering and Biotechnology 6: 45.
Landell, M.G.A., and J.A. Bressiani. 2008. Melhoramento genético, caracterização e manejo varietal. In Cana-de-açúcar, ed. L.L. Dinardo-Miranda, A.C.M. Vasconcelos, and M.G.A. Landell, 101–179. Campinas: IAC.
Landell, Marcos Guimarães de Andrade, Mário Pércio Campana, and Pery Figueiredo. 2013. Sistema de multiplicação de cana-de-açúcar com uso de mudas pré-brotadas (MPB), oriundas de gemas individualizadas. Documentos IAC, 109. Revista Campinas: Instituto Agronômico.
Le Cunff, Loïc, Olivier Garsmeur, Louis Marie Raboin, Jérome Pauquet, Hugues Telismart, Athiappan Selvi, Laurent Grivet, Romain Philippe, Dilara Begum, Monique Deu, Laurent Costet, Rod Wing, Jean Christophe Glaszmann, and Angélique D’Hont. 2008. Diploid/polyploid syntenic shuttle mapping and haplotype-specific chromosome walking toward a rust resistance gene (Bru1) in highly polyploid sugarcane (2n approximately 12x approximately 115). Genetics 180: 649–660.
Manechini, João Ricardo Vieira, Juliana Borges da Costa, Bruna Turcatto Pereira, Luciana Aparecida Carlini-Garcia, Mauro Alexandre Xavier, Marcos Guimarães de Andrade Landell, and Luciana Rossini Pinto. 2018. Unraveling the genetic structure of Brazilian commercial sugarcane cultivars through microsatellite markers. PLoS ONE 13: e0195623.
Margarido, G.R.A., M.M. Pastina, A.P. Souza, and A.A.F. Garcia. 2015. Multi-trait multi-environment quantitative trait loci mapping for a sugarcane commercial cross provides insights on the inheritance of important traits. Molecular Breeding 35: 175.
Matsuoka, Sizuo, Jesus Ferro, and Paulo Arruda. 2009. The Brazilian experience of sugarcane ethanol industry. Vitro Cellular & Developmental Biology—Plant 45: 372–381.
Matsuoka, Sizuo, Anthony J. Kennedy, Gustavo D. Eder, André L. dos Santos Tomazela, and Luis Claudio S. Rubio. 2014. Energy cane: Its concept, development, characteristics, and prospects. Advances in Botany 2014: 1–13.
Medeiros, Carolina, Thiago Willian Almeida Balsalobre, and Monalisa Sampaio Carneiro. 2020. Molecular diversity and genetic structure of Saccharum complex accessions. PLoS ONE 15: e0233211.
Mendes de Paula, Thiago Otavio, Bruno Brasileiro, Danilo Cursi, Edjane Freitas, João Santos, Marcos Resende, Collins Kimbeng, and Marcio Barbosa. 2020. Establishment of gene pools for systematic heterosis exploitation in sugarcane breeding. Agronomy Journal. https://doi.org/10.1002/agj1002.20299.
Ministério da Agricultura, and Pecuária e Abastecimento. 2020. Registro Nacional de Cultivares – RNC. Brasília. http://sistemas.agricultura.gov.br/snpc/cultivarweb/cultivares_registradas. Accessed 2 May 2020.
Molinari, Hugo Bruno, Celso Jamil Correa, Edelclaiton Daros Marur, Marília Kaphan Freitas De Campos, Jane Fiuza Rodrigues Portela De Carvalho, João Carlos Bespalhok Filho, Luiz Filipe Protasio Pereira, and Luiz Gonzaga Esteves Vieira. 2007. Evaluation of the stress-inducible production of proline in transgenic sugarcane (Saccharum spp.): Osmotic adjustment, chlorophyll fluorescence and oxidative stress. Physiologia Plantarum 130: 218–229.
Natarajan, Sijesh, Jayampathi Basnayake, Xianming Wei, and Prakash Lakshmanan. 2019. High-throughput phenotyping of indirect traits for early-stage selection in sugarcane breeding. Remote Sensing 11: 2952.
Neuber, Ana Caroline, Fernanda Raquel Camilo dos Santos, Juliana Borges da Costa, Maicon Volpin, Mauro Alexandre Xavier, Dilermando Perecin, Roberto Carlos Villavicencio Burbano, Marcos Guimarães de Andrade Landell, and Luciana Rossini Pinto. 2017. Survey of the Bru1 gene for brown rust resistance in Brazilian local and basic sugarcane germplasm. Plant Breeding 136: 182–187.
Oliveira, Karine M., Luciana R. Pinto, Thiago G. Marconi, Gabriel R.A. Margarido, Maria Marta Pastina, Helena M. Laura, Antônio V. Teixeira, Eugênio César Figueira, Antônio Augusto Ulian, F. Garcia, and Anete Pereira Souza. 2007. Functional integrated genetic linkage map based on EST-markers for a sugarcane (Saccharum spp.) commercial cross. Molecular Breeding 20: 189–208.
Palhares, Alessandra C., Taislene B. Rodrigues-Morais, Marie-Anne Van Sluys, Douglas S. Domingues, Walter Maccheroni, Hamilton Jordão, Anete P. Souza, Thiago G. Marconi, Marcelo Mollinari, and Rodrigo Gazaffi. 2012. A novel linkage map of sugarcane with evidence for clustering of retrotransposon-based markers. BMC Genetics 13: 51.
Papini-Terzi, Flávia S., Flávia R. Rocha, Ricardo Z.N. Vêncio, Juliana M. Felix, Diana S. Branco, Alessandro J. Waclawovsky, Luiz E.V. Del Bem, Carolina G. Lembke, Maximiller D.L. Costa, Milton Y. Nishiyama, Jr., Renato Vicentini, Michel G.A. Vincentz, Eugênio C. Ulian, Marcelo Menossi, and Glaucia M. Souza. 2009. Sugarcane genes associated with sucrose content. BMC Genomics 10: 120.
Pastina, Maria Marta, Marcos Malosetti, R. Gazaffi, Marcelo Mollinari, Gabriel Rodrigues Alves Margarido, K.M. Oliveira, Luciana Pinto, Anete Pereira de Souza, Fred A. van Eeuwijk, and Antonio Augusto F. Garcia. 2012. A mixed model QTL analysis for sugarcane multiple-harvest-location trial data. Theoretical and Applied Genetics 124: 835–849.
Peters, Leila P., Giselle Carvalho, Milca B. Vilhena, Silvana Creste, Ricardo A. Azevedo, and Claudia B. Monteiro-Vitorello. 2017. Functional analysis of oxidative burst in sugarcane smut-resistant and -susceptible genotypes. Planta 245: 749–764.
Pinto, Luciana, Antonio Augusto F. Garcia, Maria Marta Pastina, Laura Helena Marcon Teixeira, Jose Antonio Bressiani, Eugenio C. Ulian, M.A.P. Bidoia, and Anete Pereira de Souza. 2010. Analysis of genomic and functional RFLP derived markers associated with sucrose content, fiber and yield QTLs in a sugarcane (Saccharum spp.) commercial cross. Euphytica 172: 313–327.
Puckett, Emily E. 2017. Variability in total project and per sample genotyping costs under varying study designs including with microsatellites or SNPs to answer conservation genetic questions. Conservation Genetics Resources 9: 289–304.
Ramiro, Daniel Alves, Danila Montewka Melotto-Passarin, Mariana de Almeida, Flavio Barbosa, Sergio dos Santos, Gregorio Perez Gomez, Nelson Sidnei Massola. Júnior, Eric Lam, and Helaine Carrer. 2016. Expression of arabidopsis bax inhibitor-1 in transgenic sugarcane confers drought tolerance. Plant Biotechnology Journal 14: 1826–1837.
Rasheed, Awais, Yuanfeng Hao, Xianchun Xia, Awais Khan, Xu Yunbi, Rajeev K. Varshney, and Zhonghu He. 2017. Crop breeding chips and genotyping platforms: Progress, challenges, and perspectives. Molecular Plant 10: 1047–1064.
RIDESA-Rede Interuniversitária Para o Desenvolvimento do Setor Sucroenergético. 2020. Censo Varietal Brasil 2017/18. https://www.ridesa.com.br/censo-varietal. Accessed 16 May 2020.
Santos, Fernanda R.C., Luciana R. Pinto, Luciana A. Carlini-Garcia, Rodrigo Gazaffi, Melina C. Mancini, Bruna S. Gonçalves, Cibele N.F. Medeiros, Dilermando Perecin, Antônio A.F. Garcia, and Anete P. Souza. 2015. Marker-trait association and epistasis for brown rust resistance in sugarcane. Euphytica 203: 533–547.
Schaker, Patricia D.C., Alessandra C. Palhares, Lucas M. Taniguti, Leila P. Peters, Silvana Creste, Karen S. Aitken, Marie-Anne Van Sluys, João. P. Kitajima, Maria L.C. Vieira, and Claudia B. Monteiro-Vitorello. 2016. RNAseq transcriptional profiling following whip development in sugarcane smut disease. PLoS ONE 11: e0162237.
Singh, Ram Kushal, Shraddha Srivastava, Sujeet Pratap Singh, M.L. Sharma, Tribhuban M. Mohopatra, Nagendra K. Singh, and S.B. Singh. 2008. Identification of new microsatellite DNA markers for sugar and related traits in sugarcane. Sugar Tech 10: 327–333.
Slater, Anthony T., Noel O.I. Cogan, and John W. Forster. 2013. Cost analysis of the application of marker-assisted selection in potato breeding. Molecular Breeding 32: 299–310.
Souza, Glaucia Mendes, Helene Berges, Stephanie Bocs, Rosanne Casu, Angelique D’Hont, João Eduardo Ferreira, Robert Henry, Ray Ming, Bernard Potier, and Marie-Anne Van Sluys. 2011. The sugarcane genome challenge: Strategies for sequencing a highly complex genome. Tropical Plant Biology 4: 145–156.
Souza, Glaucia Mendes, Marie-Anne Van Sluys, Carolina Gimiliani Lembke, Hayan Lee, Gabriel Rodrigues Alves Margarido, Carlos Takeshi Hotta, Jonas Weissmann Gaiarsa, Augusto Lima Diniz, Mauro de Medeiros Oliveira, Sávio de Siqueira Ferreira, Milton Yutaka Nishiyama, Felipe Ten-Caten, Geovani Tolfo Ragagnin, Pablo de Morais Andrade, Robson Francisco de Souza, Gianlucca Gonçalves Nicastro, Ravi Pandya, Changsoo Kim, Hui Guo, Alan Mitchell Durham, Monalisa Sampaio Carneiro, Jisen Zhang, Xingtan Zhang, Qing Zhang, Ray Ming, Michael C. Schatz, Bob Davidson, Andrew H. Paterson, and David Heckerman. 2019. Assembly of the 373k gene space of the polyploid sugarcane genome reveals reservoirs of functional diversity in the world's leading biomass crop. GigaScience 8: giz129.
Tew, Thomas L., and Robert M. Cobill. 2008. Genetic improvement of sugarcane (Saccharum spp.) as an energy crop. In Genetic improvement of bioenergy crops, ed. Wilfred Vermerris, 273–294. New York, NY: Springer.
Thomson, Michael J. 2014. High-throughput SNP genotyping to accelerate crop improvement. Plant Breeding and Biotechnology 2: 195–212.
UDOP-União Nacional da Bioenergia. 2020. Usinas/Destilarias no mundo. https://www.udop.com.br/index.php?item=unidades&cn=am&id_pais=1#topo. Accessed 15 May 2020.
UNICA-União Indústria de Cana-de-Açúcar. 2020. UNICADATA. http://unicadata.com.br/. Accessed 15 May 2020.
Urashima, Alfredo Seiiti, M.F. Silva, N.F. Coraini, and Rodrigo Gazaffi. 2020. Temporal incidence of Leifsonia xyli subsp. xyli in sugarcane propagating materials of Brazilian cultivars. Crop Protection 128: 104976.
Velini, Edivaldo Domingues. 2017. Extrato de parecer técnico N°5.483/2017. CTNBio-Comissão Técnica Nacional de Biossegurança. http://www.in.gov.br/materia/-/asset_publisher/Kujrw0TZC2Mb/content/id/19141678/do1-2017-06-27-extrato-de-parecer-tecnico-n-5-483-2017-19141574. Accessed 19 May 2020.
Vettore, André L., Felipe R. da Silva, Edson L. Kemper, Glaucia M. Souza, Aline M. da Silva, Inês T. Maria, Flavio Henrique-Silva Ferro, Eder A. Giglioti, Manoel V.F. Lemos, Luiz L. Coutinho, Marina P. Nobrega, Helaine Carrer, Suzelei C. França, Mauricio Bacci Júnior, Helena S. Maria, Suely L. Goldman, Luiz R. Gomes, Luis E.A. Nunes, Walter J. Camargo, Marie-Anne Van. Siqueira, Otavio H. Sluys, Eiko E. Thiemann, Roberto V. Kuramae, Celso L. Santelli, Maria L.P.N. Marino, Jesus A. Targon, Henrique C.S. Ferro, Danyelle C. Silveira, Eliana G.M. Marini, Claudia B. Lemos, José H.M. Monteiro-Vitorello, Dirce M. Tambor, Patrícia G. Carraro, Vanderlei G. Roberto, Gustavo H. Martins, Regina C. Goldman, Daniela de Oliveira, Carlos A. Truffi, Magdalena Rossi Colombo, Paula G. de Araujo, Susana A. Sculaccio, Aline Angella, Marleide M.A. Lima, Vicente E. de Rosa, Fábio Siviero Júnior, Virginia E. Coscrato, Marcos A. Machado, Laurent Grivet, Sonia M.Z. Di Mauro, Francisco G. Nobrega, Carlos F.M. Menck, Marilia D.V. Braga, Guilherme P. Telles, Frank A.A. Cara, Guilherme Pedrosa, João Meidanis, and Paulo Arruda. 2003. Analysis and functional annotation of an expressed sequence tag collection for tropical crop sugarcane. Genome Research 13: 2725–2735.
Vicentini, Renato, Alexandra Bottcher, Michael Dos Santos Brito, Adriana Brombini Dos Santos, Silvana Creste, Marcos Guimarães de Andrade Landell, Igor Cesarino, and Paulo Mazzafera. 2015. Large-scale transcriptome analysis of two sugarcane genotypes contrasting for lignin content. PLoS ONE 10: e0134909.
Waclawovsky, Alessandro J., Paloma M. Sato, Carolina G. Lembke, Paul H. Moore, and Glaucia M. Souza. 2010. Sugarcane for bioenergy production: An assessment of yield and regulation of sucrose content. Plant Biotechnology Journal 8: 263–276.
Yang, Xiping, Md. S. Islam, Sushma Sood, Stephanie Maya, Erik A. Hanson, Jack Comstock, and Jianping Wang. 2018. Identifying quantitative trait loci (QTLs) and developing diagnostic markers linked to orange rust resistance in sugarcane (Saccharum spp.). Frontiers in Plant Science 9: 350.
Acknowledgements
The authors thank the sugarcane mills and grower associations for their support in the development of new varieties for the Brazilian sugarcane sector.
Funding
The Technical-Scientific Cooperation Program in the Area of Sugarcane Breeding (Proc. 0890/1999-56 - Proex/UFSCar) supported this research.
Author information
Authors and Affiliations
Contributions
All authors contributed equally to this work.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cursi, D.E., Hoffmann, H.P., Barbosa, G.V.S. et al. History and Current Status of Sugarcane Breeding, Germplasm Development and Molecular Genetics in Brazil. Sugar Tech 24, 112–133 (2022). https://doi.org/10.1007/s12355-021-00951-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12355-021-00951-1