
Abstract
Timing adjustment is an important ability for living organisms. Wild animals need to act at the right moment to catch prey or escape a predator. Land plants, although limited in their movement, need to decide the right time to grow and bloom. Humans also need to decide the right moment for social actions. Although scientists can pinpoint the timing of such behaviors by observation, we know extremely little about how living organisms as actors or players decide when to act – such as the exact moment to dash or pounce. The time measurements of an outsider-observer and the insider-participants are utterly different. We explain how such essential operations of timing adjustment and temporal spanning, both of which constitute a single regulated set, can be carried out among organisms. For this purpose, we have to reexamine the ordinary conception of time. Our specific explanatory tools include the natural movement known as the upbeat (anacrusis) in music, a rhythmic push for the downbeat that follows, which predicts future moves as an anticipatory lead-in. The scheme is situated in and is the extension of our formulation of E-series time, i.e., timing co-adjusted through interaction, which is derived from the semiotic/communicative perspectives. We thereby demonstrate that a prediction-based timing system is not mechanical but communicative and entails meanings for future anticipation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Although we humans resort to mechanical clocks to measure time based on the global standard time, other living organisms neither use such clocks nor count time apart from their actions or living circumstances. For these other living organisms, time and actions are unitary, not separated from one another, where time may not move linearly forward as we generally conceive. For us humans, time independently exists regardless of one’s being or acting, as it passes even while we sleep; however, we are not sure if this occurs to reptiles, insects, fish, plants or bacteria. Is it then safer for us to assume the independent existence of time for the world of organisms, since no human being has ever seen time? What can we do? Although the controversy on whether the presence of time apart from our existence is scientifically sound or not is beyond the scope of the present paper, it seems to be important to place ourselves on a logically better ground concerning the temporality issues in biology. The observation seems warranted that what is visible to us is not time per se but organisms’ act of timing adjustment. This is perhaps a safer empirical ground on which we can rebuild a reliable theory of time. The goal of the present paper is to construct a practical theory of timing from the viewpoint of organisms, not from the perspective of outsider observers. Such a theory of timing naturally entails anticipatory characters due to organisms taking action right at hand, because timing adjustment cannot be judged by a clock sitting outside the organism but must rely on the organism’s own ability to regulate the timing for operation, that is, the ability of time-ing.
Our research team has been exploring avenues to the comprehensive time theory that covers biological behaviors that are experienced from within, i.e., from the organisms’ perspective, and our concern has been how to obtain a dependable framework that permits us to see time as a semiotic/communicative process (Nomura and Matsuno 2016; Nomura et al. 2018, 2019). The underlying thesis of our theory is the notion called punctuation, i.e., the act of making a distinction by drawing a boundary somewhere in the course of an event. Without sensing some type of distinction or difference, it is difficult to attain meaning or information. For example, the sonic frequencies that can be heard by humans range between 20 Hz and 20,000 Hz; thus, vibrations beyond and below the range are not detected, that is, there is no difference and no distinction for humans. Dogs can detect up to 50,000 Hz, but humans cannot. If the environment is not punctuated, then the unheard or the unobserved area is experientially non-existent. This general principle in communication has been applied to the scheme of viewing time as punctuation or as some type of demarcation. To measure time or recognize its passage, living organisms must rely on a distinction whatever type it is. We have then reformulated an alternative picture of time by converting the philosophical time series outlined by McTaggart (1908, 1927) into our semiotic/communicative series based on perception. Accordingly, the ontological question of whether or not time exists in philosophy is modified to the practical question of how organisms address and distinguish time in communication.
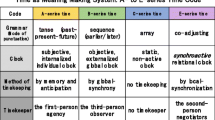
Another reformulation of ours concerning McTaggart’s time scheme is an inclusion of a new time series, the E-series, which is absent in the philosopher’s model. In addition to the objective time of the global standard time, which is called the B-series, or the subjective time of one’s sense of the past, present, and future, which is called the A-series, we have additionally proposed the inter-subjective time of the E-series that consists of the co-adjustment of timing. This is a different time scheme, but no cooperative actions are possible without it, such as humans singing in a chorus, birds flying in formations, or a guide dog leading a blind person. The inter-subjective time of the E-series made up of communicative interaction comprises local synchronization in contrast to the global synchrony of the standard time (Nomura and Matsuno 2016), and we explain this more in the subsequent section. Additionally, the C-series time and its subsidiary concept of the D-series indicate the static non-active clock such as timetables and calendars (McTaggart 1927). We thus employed the “E”-series for the proper order.
A sharp focus on timing adjustments by organisms makes the present paper unique. We have already discussed synchronicity in relation to time and its semiotic nature (Nomura and Matsuno 2016; Nomura et al. 2018) and how time in the E-series flows through interactions (Nomura et al. 2019). Thus, we currently turn to “time-ing” in terms of action (as a verb), which could be more relevant to organisms as participant-players than “timing” in terms of a time spot (as a noun). Unlike our dispassionate clocks, these acts cannot be mechanical because they are relational to meaning, i.e., anticipation. The meaning-making aspects of timing adjustment are explained with the upbeat/downbeat from the rhythmic cycle, which is connected to the command/report functions in communication. We explain these one by one in the sections that follow. Our aim here is to explain that timing adjustment is a rhythmic operation accompanied by meaning making, which is a striking contrast to the impersonal observation of timing by scientists. Migratory birds can decide when to fly off for a long journey in response to seasonal changes, periodical cicadas can decide how long to wait to appear on the ground, and hibernating bears can decide when to wake up and leave the den. All of these examples are a matter of time-ing for the participant-players. The current investigation should open a new dimension of organisms with a time-plus-environment, since measuring temporal spans and timing adjustments are the two essential aspects of a single unitary operation of how organisms live and experience.
Time as Punctuation: Our Current Position
Time, a momentous concept for most of us, is sufficiently forceful to permit a broad understanding and a wide variety of approaches. It defies against simplistic and sweeping generalizations and remains mysteriously impenetrable. Although various ways exist of interpreting the enigmatic nature of time (Aristotle 1930, p. 217b-224a, Augustine 1961, p. 253–280, Heidegger 1996, p. 279–398, Ricoeur 1984, p. 3–87), we view time as a relational or meaning-making system based on punctuation (Nomura and Matsuno 2016; Nomura et al. 2018, 2019). Humans feel the passage of time when noticing the hands of a clock progressing, seeing the sun tilting, or hearing church bells, and all of these recognitions are the result of sensing continuous states or events that have been marked off or punctuated. To measure time or recognize its passage, organisms must rely on some distinction. As already addressed, migratory birds, periodical cicadas, and hibernating bears all choose their timings for flying off, appearing on the ground, or leaving the den, respectively. These choices or action-takings correspond to punctuation. Even bacteria adjust their timing and synchronize the behavior of all members of the group once their population density reaches a threshold concentration; scientists have named this quorum sensing (Bassler and Losick 2006; Rutherford and Bassler 2012). Bacteria’s timing adjustment, that is, their taking action at the threshold concentration, represents punctuation.
Punctuation is boundary making in the course of an event or a state that causes an organism to sense a sign or news of difference through some type of distinction. Thus, the progression of time is noticed only if organisms detect such news of a difference that matters or “a difference which makes a difference” (Bateson 1972, p. 457–461). The important function of punctuation in communication is that it restrains subsequent coming actions by pushing them into the expected scope of possible alternatives (Bateson 1972, p. 130–131) and thereby creates more adaptable meanings for the actions to take. When a comma punctuates a sentence, for instance, the next sentence starts in light of the previous one. When a certain rhythm gets started, we can predict the next rhythms to come. Based on these observations, we can deduce that punctuation leads the participant to anticipation under the restraint, which may develop further meanings. That is, restraint and anticipation are almost synonymous but are only looked at by different observers. For an outsider-observer, punctuation appears to limit the number of choices for subsequent actions out of the possible alternatives. For the insider-participant, however, punctuation looks predictive and anticipatory and provides clues for his/her future actions.
We summarize our premises as follows based on our previous works (Nomura and Matsuno 2016; Nomura et al. 2018, 2019).
-
1.
Time is not about substance but is communication between participating agents, namely, a sign activity that uses a series of distinctions or punctuations. This claim, which is based on the relational activity of communication between participating agents, turns our traditional notion of time upside down. That is, instead of considering time as something that exists as a physical entity, we maintain that living organisms generate time by making distinctions and punctuations in the course of their communicative interaction with their immediate environment or with other individuals. How is this claim justified? From the point of view of communication, if a distinction is not perceived or if a difference does not make any difference, then time has not occurred. In the absence of any indexical time marker – whatever the type – no one, no organism, and no cell experiences the passage of time. The phenomenon called time occurs in the domain of communication.
-
2.
Thus, a differently punctuated series would lead to different time experiencing. Just as we observe a different beat in music with rhythms (punctuation) and tempo (speed), a different series of punctuations should represent a different series of indexical temporal signs and, therefore, a different time series. Specifically, time cannot carry only one implication. Thus, a different time series possesses a different mode of punctuation, which corresponds to the grammar of this particular series.
-
3.
The time grammar (mode of punctuation) of the A-series rests on the past, present and future tenses, which together support a sense of duration or an unbroken succession of events. Tense is a linguistic property; thus, this time series primarily applies to humans, although it does not exclude living organisms completely. The A-series corresponds to subjective time and assumes a single agent that has access to its own memory and anticipation. The personally felt slow progression of time or the recollection of past events from the vantage point of the present may be examples. When such a flow of time is sensed by the first-person agency, time appears to him/her in the A-series. Within the short span from the immediate past to the immediate future, living organisms such as a giant anteater (Myrmecophaga tridactyla) may also experience time in the A-series through its continued activity of capturing ants one after another in its subjective tempo.
-
4.
Time in the B-series is characterized as tense-less, and the mode of punctuation depends on the earlier/later (or before/after) relations, not tenses such as the past, present, and future. The B-series corresponds to the objective time that is read by a third-person outsider observer. Our ordinary clock may be the representative example par excellence. That is, when punctuated sequences of events are seen from the perspective of earlier/later, time appears to the observer to be in the B-series. The mode of punctuation in the B-series is unidirectional so that it does not permit our participation. Our global standard time shows the B-series characteristic of moving in one direction with global synchrony. It is a lineal Newtonian time that permits no creatural participation in its progress, and only the third-person onlooker situated outside reads its time. B-series time is of paramount importance for human social life, mass production, and modern science, without which our techno-centered civilization would be difficult to live in. However, it might be an unnecessary formality for paramecium.
-
5.
Time in the C-series is a static array. For example, when you map your time frame on a piece of paper, the product would be a static picture of time with no progression, which represents non-active, tense-free C-series time. This map removes both tense and earlier-later relations from the sequence. For example, the face of a clock, timetables, music scores, itineraries, or calendars are all related to time progress but remain inactive and stationary and indicate no specification of the direction of movement. This map of time, similar to a map of terrain, does not specify the direction of movement – you can use it for either climbing up or going down. The map itself does not specify the direction. Thus, C-series time is a time pre-scenario before being used. In relation to circadian rhythms and biological clocks, we have also discussed that internal models and feedback loops of gene circuits as a scheme or design represent C-series time to the external observer (Nomura et al. 2018, p. 73–77).
-
6.
Time in the E-series is interactive, which results in the co-adjusted temporal spans in rhythms and movements. When you conjointly determine the timing of punctuation with other people through interaction and if the effects are reciprocal, the process would represent time in the E-series. The E-series is neither objective nor subjective but inter-subjective. It is in a continuous process where two or more organisms or people communicate with one another – like a dialogue – and mutually punctuate toward a collective alignment of their rhythms. Such mutual temporal spanning is cooperatively achievable in conversation; it is a pair dancing, such as two kittens playing with one another, schooling fish, mosquito swarms, etc. The examples in the E-series exhibit a local synchronization of movement that contrasts with the standard time of the controlled global synchrony in the B-series. Time in the E-series is a joint effort of the participants to share the now from one moment to the next through continuous negotiation.
What Is Timing?
Consider the word timing, which consists of a combination of a verb, time, and the progressive mode of –ing. Although “timing” is ordinarily used as a noun, which means “the precise moment for acting effectively,” it can be considered to be a gerund that means “timing.” According to the Merriam-Webster dictionary, time as a verb (transitive) includes the following definition: “to set the tempo, speed or duration of” as in the case of “She timed her leap perfectly.”
In this paper, timing (n.) indicates the noun usage of timing, whereas timing (v.) indicates verb usage. Timing (n.) as a noun belongs to the B-series, because the time point of “the precise moment for action” is specified on the tense-less earlier/later scale, which accords with the definition of the B-series. In contrast, timing (v.) as a verb belongs to the E-series, because ongoing timing adjustments of “tempo and duration” are in constant negotiation with one’s communicative partners or with the environment.
Imagine, for instance, a case of walking hand in hand with a small child. You must match the tempo with the toddler by considering both the length of your stride and the timing (v.) of your steps. However, it is obvious that only a single operation occurs. As another example, think of a bird feeding her chicks. Baby birds open their mouths all at once when the mother arrives at the nest with food. Chicks can decide when to open their mouths and for how long. Their time management, specifically, the timing (v.) of when to open their mouth and the measurement of duration, are all determined interactively.
Turn taking is one of the most basic rules for human conversations, where it is customary for one participant to speak at a time in an alternating manner (Sacks et al. 1974). When to open a mouth for us to speak is also a matter of timing (v.). Interestingly, turn taking is also observed among songbirds. Two territorial male birds in California, namely, the wrentit (Chamaea fuscata) and Bewick wren (Thryomanes bewickii), alternate their turn to sing in the same chaparral environment (Cody and Brown 1969). The cycles of the two species are asynchronous so that each species sings most frequently when the other bird’s singing is at a low ebb. To avoid acoustic interference from similar signals of other species, Amazonian songbirds display turn taking between species for using different time slots of the day. White-browed antbird (Myrmoborus leucophrys) and warbling antbird (Hypocnemis cantator striata) are found to sing their “dawn chorus” primarily within 30 min of sunrise, while chestnut-backed antshrike (Thamnophilus palliatus palliatus) and black-faced antthrush (Formicarius analis) primarily sing from one to two hours after sunrise (Luther 2008). Although the ornithologists do not use the term turn taking, we find it to be consistent to include such temporal acoustic partitioning in the notion of turn taking. Since timing (v.) is one of the essential operations for living organisms to survive, the principle of timing (v.) applies widely to a large domain of biological behavior.
These are the practical aspects of timing (v.) that do not assume time as a given or a priori. That is, when we consider the nature of timing (v.), we do not need to presuppose the concept of time, just as organisms in general never do. Consequently, it becomes unnecessary to presume the existence of time; this interpretation corresponds to time as a sign activity (cf., II – 1). Because living organisms do not presuppose the existence of time, by timing (v.), we emphasize “setting the tempo” as a verb rather than “the right moment” as a noun. “Play” comes into existence only during play. It does not exist apart from the action of play. Time is similar in character.
Adjusters of Timing (V.): Upbeat and Anacrusis
The key idea that we rely on to explain timing (v.) is the natural movement known as anacrusis or upbeat, which is a concept used in musicology. Anacrusis is a rhythmic push-up for the downbeat that participating agents or performers follow. For example, we often prepare for the start of our physical action by saying “Ready, Go!” Here, “Ready” is not the action itself but a part of the same process – momentum building – that leads to an occurrence of the main action. “Ready” thus corresponds to anacrusis. Alternatively, when we take a snapshot photo, we say “One, Two, Three” before pressing the shutter. “One” and “Two” are not exactly a part of snapping the photo, but they help the participants synchronize their joint actions. “One” and “Two” correspond to the anacrusis or upbeat.
Anacrusis in music indicates a set of notes or beats that precede the legitimate part of a musical phase. This upbeat consists of the unaccented notes before the first downbeat in a bar, which is often called a “pick up”. Since the anacrusis is a lead-in or a push-up for the subsequent downbeat, an anacrusis is anticipatory and directed toward a future event (Hasty 1997: 120–129).
Here, John Denver sings “Take Me Home Country Roads,” and notes (1), (2) and (3) correspond to the anacrusis.
To further understand this notion, it may be helpful for the reader to imagine, for example, a case of listening to and following a musical piece with a fast tempo. To follow the piece of music, our attention must be relatively continuous; otherwise, we will no longer be with the ongoing event. Notice that to follow the music is not to trail along behind the notes or even to keep up with them “at every instant”. Instead, to follow requires above all that we keep moving ahead and that we anticipate what is about to occur to follow what is happening. Anticipation in this sense is not the projection of a definite outcome but a readiness to interpret emerging novelty in light of what has gone before (Hasty 1997: 69). It is a retrocausal action where the immediate future event becomes the cause of the present behavior in terms of “acting for the present in the immediate future” (Nomura et al. 2019).
Anacrusis is not a symbol but rather an indexical sign toward the participating practitioners. It is a sort of lead-in that is anticipatory. Living organisms actively utilize rhythmic reverberations between the perceptions of upbeats and actions that occur with a downbeat and move back and forth. We suspect that these rhythmic cycles serve to better secure organisms’ chances for survival with regard to phenomena such as heartbeats, bird songs, and internal circadian rhythms.
We have so far presented a theoretical framework of anacrusis or upbeat. We now provide examples to illustrate our theory. The reader is once again reminded that our purpose is not to explain the outsider observer’s action that tracks along a prepared time scale. Instead, our purpose is to explain the insider’s way of measuring a temporal span through his/her interactive movements that adjusts timing for action (Fig. 1).

Bill Danoff, Taffy Nivert & John Denver
Example 1
Conducting Triple Time – Let us suppose that you conduct an orchestra in triple time (see Fig. 2). If Movement 1–2 is downbeat, then Movement 2–3 becomes upbeat in preparation for the next action, which builds momentum for the coming locomotion.

Conducting triple time
The use of a baton for conducting does not occur in a vacuum. The orchestra members play in concert so that the conductor’s act is a part of the entire movements of the ensemble, all of which head for local synchronization, including the audience. Leading an orchestra is therefore based on mutual adjustments of timing.
Example 2
Caterpillar Eating a Leaf – When the larva of a butterfly eats a leaf in an arc shape, the repetitious and rhythmic movement shows its anticipation for the next round of leaf grazing. Watching at the very end of a biting round, we notice that the caterpillar engages in the following two different activities: (a) its punctuating action to complete the preceding grazing round and (b) an anticipatory push-up to move itself to the subsequent starting point. The anticipatory push-up (b) constitutes an upbeat (anacrusis) that gathers momentum, and the subsequent grazing round may correspond to a downbeat. An upbeat in this case also contributes to the economy of energy expenditure in that the smooth transition to the initial position would make rhythmical repetition easier and faster.
Example 3
Frog Swimming in Water – Most frogs swim well. They kick their legs backward to propel themselves forward in the water. Upbeat and downbeat each apply to their movements of bending and stretching their hind legs. When they draw their hind legs upward toward their body, the movement may represent an upbeat to prepare for a kick. Then, when they shoot their legs out backward, the movement may represent a downbeat for the execution.
This rhythmical cycle of the frog is affected by the water conditions that happen to surround it; thus, kicking and swimming occur in an environmental context. When the frog acts against the water, the frog and water together constitute a feedback loop in which information is transmitted as if via cybernetic circuitry. Thus, the frog’s locomotion occurs in the E-series time frame because of its interaction with the surrounding water.
Example 4
Courtship Sequence of the Stickleback – Tinbergen’s well-known report on the three-spined sticklebacks’ courtship (Tinbergen 1953) is a good example that illustrates the upbeat-downbeat sequences in animal interaction. Compared to the previous three cases (i.e., the triple time, the caterpillar, and the frog), interactional characteristics are clearly seen in the stickleback’s courtship, with mutual engagements of a male and female fish. Fig. 5 shows each step of their courtship sequence from the male-female encounter to the final stages of spawning and fertilizing.
The zigzag dance is the male’s first gambit when a female appears in the territory. This may or may not appeal to a particular female; however, a receptive female that is attracted by the zigzag dance turns toward the male instead of fleeing. Then, the male turns around and swims toward the nest, and the female follows it. After arriving at the nest, the male shows the nest entrance to the female, and the female slips in. After the male prods the female’s tale with his snout, the female spawns after some time. Then, the male enters the nest, in turn, and fertilizes the eggs. The male then chases the female away as a part of the sticklebacks’ courtship behavior (Tinbergen 1953: 8–14).
Such interactive sequences have some basic principles in common. The animal’s actions toward the other – whether they are sending acoustic cues or displaying nonverbal behaviors – always exhibit communicative functions at two different levels simultaneously. One level reports the content of the message, and the other level commands some action of the partner (Ruesch and Bateson 1951: 179–181). For instance, the male stickleback that leads the female to the nest in Step 4 reports what he has done and at the same time commands the female by virtually saying “Follow me!” Alternatively, when you receive a greeting card from your friend, the card gives you a sort of command to send it back to him/her apart from the message content. Any single message communicated between any pair of participating agents reports and commands simultaneously.
The above notions become clearer if we consider all the arrows in Fig. 5. The male’s zigzag dance performed when a female appears displays – among other things – his fit bodily condition as the message content, which corresponds to the level of a report. At the same time, the dance makes an appeal to the female to “like” him in human terms or to court, which belongs to the level of a command (as indicated by the arrow from Step 2). That is, the results from the preceding acts set the conditions for initiating the succeeding acts (Matsuno 2020). We admit that these explanations are somewhat anthropocentric if both report and command are taken to be symbols that have already been accepted in our languages. However, we have applied these terms as indices. The agents that maneuver these indexical signs are both male and female sticklebacks, not ourselves.
However, the two indexical functions within a single message, namely, command and report, are operationally connected to the ideas of upbeat and downbeat. Since the zigzag dance is a reaction to the appearance of a female, the appearance of a female is the immediate cause or a lead-in to the male’s dance, which works as an upbeat. That is, the appearance of the female not only reports “her appearance” but also commands or upbeats the male to display the zigzag dance (the arrow from Step 1). His zigzag dance (Step 2) is then a report about the appearance of the female (as a downbeat) and simultaneously a command for the female to be attracted to him (as an upbeat). The report corresponds to a downbeat, while the command corresponds to an upbeat (anacrusis). Specifically, a report is about the completed act so far, and a command is about a de novo act yet to come. Similarly, his zigzag dance now plays the role of an upbeat toward the female and solicits the female to court (Step 3). The female courting as a report about the zigzag dance then functions as an upbeat and prompts the male to lead her to the nest as a downbeat (the arrow toward Step 4), and so on.
Thus, in an interactive context, we can assign both an upbeat and a downbeat to a single communicative act, which potentially possesses the two message levels of a report by one agent and a command to another agent. If we consider the entire sequence of interaction between the male and female sticklebacks from the point of view of the participants, then the observation sheds light on the nature of upbeats and downbeats in a more lucid way.
Example 5
Synchronized Hatchings of Stink Bugs – The timing of hatching varies in response to multiple environmental risks and opportunities, which are mediated by a variety of cues and mechanisms, and this is called environmentally cued hatching (ECH) (Warkentin 2011). Although we tend to think that hatching is a developmental event that occurs at some fixed stage, we should not assume that hatch timing is fixed. That is, when the cost/benefit ratio is more favorable outside the capsule than inside, embryos should hatch; thus, hatching is essentially a “decision” based on information, i.e., it depends on a specific behavioral or physiological process that is environmentally cued (Warkentin 2011: 17).
One of the environmental cues on hatch timing includes the effects of sibling contacts. Although there is some evidence in several freshwater turtles for synchronous hatching via responses to cues from siblings (e.g., Spencer and Janzen 2011), the scope of embryo-embryo communication is not fully known. The following is a case illustration from insects. Embryos of the brown marmorated stink bug (Halyomorpha halys) synchronize hatching by responding to vibrations made by siblings cracking their eggshells (Endo et al. 2019). The vibration spreads across a clutch and induces explosive hatchings. This highly synchronized pattern is promoted by immediate hatching responses to vibrational stimuli generated by the cracking of previously hatched eggs (see Fig. 6).
Consistent with our argument, the command aspect of previous egg hatching may be equated to an upbeat, while the report aspects of subsequent hatchings may be equated to a downbeat. The near synchronous hatching may be made possible through the quick progression of upbeats and downbeats. Thus, eggshell cracking, which is a rigorous process itself, becomes a report to siblings via the medium of vibration and simultaneously becomes a stimulus or a command to the other embryos within the clutch to follow suit. In this way, the previous egg cracker commands (or upbeats) the other siblings to crack their own eggs, where cracking becomes a report (or downbeat). In this species, synchronized hatching is crucial for survival because individuals that hatch earlier than the rest of the clutch may cannibalize sibling eggs that have not cracked for nutrient supplements.
Environmentally cued hatching (ECH) should be the embryo’s act of timing adjustment, i.e., timing (v.), regardless of whether hatching with siblings is synchronous or asynchronous or whether hatching occurs in a laboratory or in the field. As described by Warkentin (2011) earlier, when the cost/benefit ratio is found by the embryo to be more favorable outside the capsule than inside, hatching, which is a “decision” based on an information process, should follow. A case was reported on an asynchronously hatching bird, the glaucous-winged gull (Larus glaucescens), to examine whether the chicks modify their hatch timing in response to sibling proximity in a laboratory and in the natural field (Schwagmeyer et al. 1991). The study found that the sibling eggs incubated in the field received cues from parents, colony milieu and other ambient sources to deliver normal hatching, whereas the artificially incubated eggs in close contact to siblings in a laboratory received their cues for “normal” asynchronous hatching, but the eggs placed physically apart experienced a retarded hatch. The eggs in a laboratory without close contact with their sibling eggs apparently failed to receive a news of difference (i.e., report) that makes a difference (i.e., command). The timing (v.) for hatching seems to be environmentally determined or interactively negotiated within each environment, which indicates that time management is not the sequentially fixed developmental event in the B-series but the negotiated co-adjusting relational event in the E-series.
Discussion
We now clarify the following three points based on what we have argued thus far, including our new proposition that timing (v.) has organizational effects on living systems.
(i) Upbeat/Downbeat (or Command/Report) as Tense Markers
We have observed that the phenomena of upbeat and downbeat in a rhythm sequence are accompanied by communication functions such as command and report. The case of the sticklebacks (Fig. 5) has shown that the sequentially listed action (Steps 1 through 10) plays such complex roles. In Fig. 5, it may be more accurate to indicate that all the steps (Steps 1–10) correspond to downbeat (and report) and that all the arrows correspond to upbeat (and command). Thus, the male stickleback’s act of showing the nest entrance (Step 6) is a downbeat and a report to the female about the nest’s location, and it is also an upbeat and a command to the female to slip into the nest (as indicated by the arrow toward Step 7). Upbeat as a message has a command function, and downbeat as a message has a report function, and these two sets continuously alternate their turns in interaction.
At this point, two linguistic tenses are relevant – i.e., the present progressive tense and the present perfect tense – for the interacting players (Matsuno 2013a, 2013b), since organisms that participate in interaction in such cases as the sticklebacks jointly “progress” toward measuring temporal spans (all the arrows) and collaboratively set timing for action as a tentative “perfection” (Steps 1–10). Tense is viewed as a grammatical category of an indexical nature that expresses a time reference – such as the past tense, the present tense, the future tense, etc. For the current case of interactive sequences, however, the present progressive tense and the present perfect tense in English may be the two promising candidates for a linguistic time reference. These interactive processes are neither subjectively based on a single agent’s past, present and future (in the A-series) nor objectively based on the outsider’s view of earlier/later relations (in the B-series) but inter-subjectively based on the participants’ mutual time referring (in the E-series). In the E-series, these two tenses are connected and alternate with one another through the players’ efforts for the mutual time referring that is destined to leave slight misalignments between them so that the collaborative making-up for the misalignments would make the interaction continue (Nomura et al. 2019: 271).
Thus, the stickleback’s act of showing the nest entrance as a report (Step 6) registers the linguistic time that represents the present perfect tense, whereas the arrow commanding or being directed toward the female to enter the nest (the arrow toward Step 7) represents the linguistic time of the present progressive tense. Alternatively, when conducting in triple time, as in Fig. 2, Movement 1–2 serves as a downbeat that can be seen as the present perfect tense, while Movement 2–3 can be seen as the present progressive tense. The reader may notice that upbeats and commands should roughly be parallel to the present progressive tense, and downbeats and reports should roughly be parallel to the present perfect tense. This is the reason that upbeat/downbeat and report/command are considered to be indexical tense markers for the insider players to indicate the flow of linguistic time.
If command/report can also be considered tense markers, then the entire meaning-making sequence as a single system – the sticklebacks’ interactions in Fig. 5, for example – can be regarded as an “E-series time clock.” Although this “clock” is, of course, different from the tense-less B-series time clock that we generally experience that is based on the global standard time, it is a relational or meaning-based clock that may remain legitimate for insiders as the organisms that participate in an action. Thus, the rhythm-making upbeat/downbeat and the meaning-making command/report can both become tense markers that register linguistic time. Since Examples 1 through 5 suggest updating the two tenses — the present progressive tense and the present perfect tense — repeatedly, we presume that the meaning-making acts in the five examples (Figs. 2, 3, 4, 5 and 6) all function as tense markers for the insider players that participate in the operation. Thus, the indexical sign activities of upbeats/downbeats or commands/reports could be temporal expressions for the internal observers. From the point of view of tense, each series time is summarized in Table 1.

A caterpillar (i.e., larva) of a tiger swallowtail butterfly eating a leaf. Permission obtained from ViralHog https://www.youtube.com/watch?v=7k4w7nwIQAs

Frog swimming in water

Courtship behavior of the three-spined sticklebacks (Tinbergen 1953, p. 8–14)

Synchronized hatchings by stink bugs (Endo et al. 2019)
Accordingly, an interaction can become a clock for the players in a system. A clock placed outside a system specifies the predetermined B-series for a third-person observer, whereas a clock placed inside a system expresses the continuous process of the determination of the E-series for second-person (inter-subjective) participants. Time ticks differently for someone who observes a system from the outside than for someone (organisms) who acts on a system from within; the two time series are different in character and yet are made commensurable with one another, although the participating sticklebacks do not care about such a relationship to be delineated between the series.
(ii) Prediction and Anticipation are Synonymous with Command/Report
The illustrations in Figs. 2, 3, 4, 5 and 6 assume predictive behaviors, namely, a person who builds momentum by expecting to swing a baton down (Fig. 2), the caterpillar’s anticipatory moves toward the starting point for grazing leaves (Fig. 3), a frog bending its hind legs for swimming and anticipating subsequent kicking (Fig. 4), the male and female stickleback’s successive incitements for anticipated actions to follow (Fig. 5), and the coordinated hatching of stink bug embryos to avoid cannibalization (Fig. 6). These can all be interpreted as examples of anticipatory behaviors.
When anticipation is involved, the responses are not mechanical but presumed to be communicative, and they entail meanings unique to the participating players. Prediction and anticipation, which are both tinted psychological concepts, are transferred to communication terms, and the introduced notions of command/report may be able to substitute for them. Since any message in the communication environment has the two functions of command and report, the command function conveys information for the expected future, which helps the organisms make a prediction in an anticipatory manner. For example, when a sports player senses a command message from his/her teammates, this perception leads to the prediction and anticipation for his/her next moves. Command permits room for the capacity of making choices, that is, to help focus on the relevant context out of which to make choices. Command is a message about the message in focus, since it instructs what to do with the content of report. Your friend shouts, for example, “I’m in a hurry!” This instructs you not to speak to him/her at the moment. “Not to speak to” is the meaning derived from the reported message so that it is the message about the message, “I’m in a hurry!” The deduced meaning of “not to speak to” is an anticipatory meaning. For this reason, we say command is a rough synonym for anticipation, although a command is not separated from a report.
We can utilize the notions of report and command to circumvent the likely charge of anthropocentrism. We employ report and command to refer exclusively to joint indexical signs, not symbolic signs. That is, report in this paper refers not to a statement but to a state that is communicated with the partner. For example, there is a remarkable finding that bumble bees damage the leaves of flowerless plants in a way that accelerates flowering time (as much as 30 days) when pollen is scarce (Pashalidou et al. 2020: 881–884). With climate change or phenological irregularity, floral resources are sometimes scarce when bumble bee workers need to collect pollen to establish their spring colony. Instead of passively waiting for timing (n.) to bloom, bees actively engage in making the plant’s internal clocks move faster by timing (v.). The pollinator’s behavior of making a cut into foliage is not only damage to the leaf but also stimuli, specifically, a report message, which also contains a command message that induces the plant to accelerate flower production. Without having an anticipation of early flowering, the bees would not “ask” the plants for help in providing them food. Plants, in turn, get successfully pollinated by meeting the bees’ requests. Accelerating flower production is therefore an adaptive strategy for both organisms connected via communication functions of report and command – without resorting to so-called languages or symbols.
As we noted before, anticipation is not the projection of a definite outcome but a readiness to interpret emerging novelty in light of what has already gone before (Hasty 1997: 69). Thus, the subsequent act is not automatically predetermined (such as deciding exactly when to bloom) but is sufficiently constrained to assume the role of an indexical signs (to bloom earlier due to the instigation of the cuts made into foliage). Such constraints may be the nature of prediction among organisms (or among bumble bees).
(iii) Timing (v.) is Organizational
Biologically speaking, it should be more realistic to equate a unit of life to “an organisms-in-its-environment” (Bateson 1972) or to an organism’s subjective environment, Umwelt (Uexküll and Kriszat 1957), than to an individual organism that is separated from the outside by its skin or cell membrane. In our view, what survives is organisms-plus-environment as a whole, and this framework is the basis of the subsequent discussion. The examples of the smallest unit of life, organisms-plus-environment, may include the caterpillar and the leaf (in Fig. 3), the frog and the water (in Fig. 4), a single egg among many in the clutch (in Fig. 5) or bumble bees and the plants. Without water, there is no kicking by a frog. Thus, organisms attempt to organize all the organisms plus the local environment, which is the smallest unit of survival.
This subjective environment (Umwelt), that is, how organisms perceive and interpret their own immediate surroundings, may be identical to the organism’s view of its environment or the organism’s internal model of the environment (Franklin and Wolpert 2011, Kawato 1999, Wolpert et al. 1995). An internal model is a scheme and a map of the internalized environment or semiotic scaffolding, that is, a web of checks and balances established through the communication between organisms (Hoffmeyer 2015: 154) and constructed and manipulated through processes of signification. The organisms utilize their maps of the environment (i.e., the C-series) to process biological and ecological functions. The C-series, which is a static picture of time, thus corresponds to non-measuring, tense-free pre-scenarios. Similarly, chemical cycles and gene circuits are also static pictures of some dynamic movements expressed without time in the C-series (Nomura et al. 2018: 73–77).
How then can organisms obtain an active moving process of trial-and-error out of the static scheme of the subjective environment? Time factors may be needed to explain the process, and this is where the notion of timing (v.) – not timing (n.) – comes in. What mobilizes the biological system forward cannot be based on externally measured pace and tempo but internally measured concerted action – such as the working of pacemaker cells that regulate our heartbeat from within. Furthermore, organisms-plus-environment does not stand still – it is a dynamic process of changing, evolving and balancing that requires managing and coordinating the system to keep it going. Accordingly, organisms-plus-environment must be organizational in the sense that the present state is constantly updated, and simultaneously, the balance of the surviving relationships is kept resilient. That is, the time to bloom is updated through communication between the bees and the plants, while their relationship (i.e., pollinator and floral resource provider) is kept resiliently unchanged.
Thus, we operationally define organizational to refer to the indexical sign activities for updating the state and maintaining the relationship within the organism-plus-environment to increase the chances for survival. Here, sign activities are the patterns of interaction that have been historically handed over for moving the system forward, as noted by Hoffmeyer (2015).
Sustaining one’s survival is a matter of duration. The duration requires time operation in a coordinated fashion within its subjective environment. As we have introduced in the previous section, the two sets of upbeat/downbeat and command/report enable both timing adjustment and meaning making for living organisms. We clarify the organizational effects of these two sets and suggest that timing (v.) composed with these sets can also be organizational.
(iii-1) Upbeat/Downbeat is Organizational: Renewing and maintaining rhythms are made possible by upbeats gathering momentum and downbeats marking punctuations. Upbeats drive the continuation forward to help prepare a partner’s subsequent beginning (downbeat), and vice versa. This is a mutual adjustment – between the participating organisms or between the organism and its immediate environment – of temporal spanning that allows the joint coming together of punctuations. Such an agential operation entails alternating turns of the two tenses, the present progressive tense and the present perfect tense, where upbeat and downbeat function as semiotic scaffolding, or tense markers, for the joint making of a rhythmic sequence.
Upbeats/downbeats are therefore organizational in achieving the timing (v.) of rhythmic coordination as seen in the examples of conducting an orchestra, a caterpillar eating leaves, or a frog swimming in water. They are organized in relation to their immediate local environment (i.e., the subjective environment). In an interactive setting, the turns of upbeats and downbeats in the sticklebacks’ sequential moves become apparent with successive courtship behavior. Since these sequences are not mechanical but communicative, that is, each step and each move is constantly affected by the other parts of the organism-plus-environment, they consequently leave room for a certain flexibility and freedom. Thus, to be organizational, a living system maintains itself by constantly moving forward and constantly leaving some margin for leeway to the extent that communication is unpredictable. Such maintenance is necessary not to pass over fatal deviations in the system.
(iii-2) Command/Report is Organizational: Although the idea of upbeat/downbeat is necessary to emphasize the aspect of temporal management, the notion of command/report is useful to illustrate the meaning-making aspect of interactional exchange. The report message level displays some action, while the command level stimulates the players to take action. Since the participants mutually determine the timing of punctuation for the engaging event, these functions have been considered to be tense markers in the E-series. Such sequences virtually punctuate temporal spans and incite the partner to anticipate the next action in the near future; therefore, they are organizational, which corresponds to a player-dependent act of meaning. In the case of the sticklebacks, the male showing the nest entrance (report) makes the female anticipate (command) the next action, which is entering the nest. The male’s intent may be shared with the female only when the timing is right. Anticipation and prediction have time-dependent meanings for each participant; however, we might extend them by saying that communication in terms of meaning making is always time dependent. That is, timing (v.) may be a context out of which scientists can image the organism’s view of itself and its environment. The meaning-based report/command is an aspect of timing (v.) and is organizational in playing a significant role in prediction and anticipation. Since prediction and anticipation are types of readiness to interpret emerging novelty instead of projections of a definite outcome (Hasty 1997), they are communicative, not mechanical, where even the next expected move is bound by context, which leaves the possibility for a different presentation or different interpretation along the course of the exchange.
Upbeat/downbeat and command/report are inseparable and are two sides of the same coin that constitute the organizing principle for living systems. In possessing these two functions, timing (v.) is organizational, because upbeat together with downbeat pick up momentum to organize the subsequent actions to follow and because command together with report guide the near future to carry a meaning of anticipation.
We supplement our argument with a few further remarks. While organisms’ actions and interactions are seamless and unbroken, we humans cut them into segments using such concepts as the upbeat or command to understand the insider players’ living worlds. Although the use of concepts is unavoidable when seeking to grasp scientific reality, it gives the reader an impression that the upbeat or command could exist as an independent phenomenon. However, for organisms to participate in their environment with right timing or synchronize with other individuals, it is imperative for them to view the downbeat-upbeat or report-command as a single, inseparable unit and as occurring simultaneously. The zigzag dance of a male Stickleback, for example, is downbeat and upbeat “at the same time” or report and command “at the same time”. Such would be the insider participants’ frame of reference or the way in which the E-series time clock works, permitting unbroken, continuous action by organisms.
Timing adjustment becomes acutely challenging in inter-specific contexts. We have presented bumble bee-plant interaction as an example of such coordinated action of time-ing, with each organism changing its responses to the other while keeping the pollinator-resource provider relationship unchanged. The bees’ goal of collecting pollen matches the plant’s goal of being pollinated. The trial and error process of both species results in bumble bees damaging plant leaves and the flowering plants setting their internal clocks ahead to hasten flower production. Such adaptations for mutual benefit may be a small piece of real-time coevolution on site. Of course, the phenomena of co-evolution are diverse and tremendously complex, involving predator-prey or herbivore-plant relations, but scientists indicate that pollinators are dominant agents of plant evolution (Gervasi and Schiestl 2017, Agrawal et al. 2012).
Relying on an E-series time clock is not a guarantee that organisms are always successful in their timing adjustments. They often miss a good moment to catch prey, inadvertently get injured, or fail to get a chance to mate. Despite their unsuccessful attempts, their behaviors and efforts do not stop; they continually move ahead and are sustained without cessation. This is partly due to the organisms’ capacity to learn. While a simple machine’s behavior is specific and fixed and such a machine is incapable of learning by trial and error -- the right or wrong is not subject to correction -- organisms change their responses by correcting prior errors, which improves their future skills. The former Zero learning is in contrast to the latter Learning I (Bateson 1972: 283–292). In this vein, a behavioral blunder is not exactly a mistake or a failure, since organisms are constantly learning through trial-and-error and error-and-trial rhythms, where the error is an upbeat inseparable from the next trial as a downbeat.
Learning a skill is an anticipatory act. If an organism has a future goal in mind, such as in the case of catching prey, then the expected future state becomes the cause of the present behavior. The direction of causality is reversed – since the cause resides in the future, it is called retrocausality or reverse causation. Here, the errors committed in the cut-and-try ongoing process are cancelled out and nullified. In other words, an error is no longer an error when aiming at the target, since the errors are the way in which organisms approach the target. We elaborated such semiotic mechanisms of predictive movement with the term retrocausal scaffolding, in which the corrective actions are constantly updated so that the present choice of action erases past errors in the recursive feedback loop run by the E-series time clock (Nomura et al. 2019: 279–284). It is in the B-series time code that past events, such as failures or successes, remain frozen in the record.
Conclusions
We have attempted to clarify the way in which organisms adjust timing for action, i.e., time-ing or timing (v.), in relation to other individuals or their environments. We have also indicated that such timing (v.) processes are sensitive to context, which implies the participation of a player that can identify the context. Young chili plants (Capsicum annuum), for example, are found to be able to sense their neighbors from as early as the seed stage, and as seeds grow into seedlings, they can discriminate among neighboring species and modify their growth patterns accordingly (Gagliano et al. 2012). As another example, as we noted, quorum sensing is a bacteria’s way of identifying its context, that is, the group-wide detection of information about cell density and species composition of the vicinal community, which enables bacteria to collectively execute behaviors (Rutherford and Bassler 2012). These are not mechanically based because the subsequent actions are not predetermined but left undetermined for the inter-player negotiations. We applied the musical concept of upbeat/downbeat to explain the rhythmic mechanism of timing (v.), and this led us to the communication functions of command/report as it is operationally tied to. One support for our contextual-meaning scheme comes from the fact that sensing the command level of a message predicts or generates anticipation, which is supposedly meaning procreation. Thus, rhythm making (upbeat/downbeat) for living organisms comes together with meaning making (command/report) to theoretically separate living cycles from mechanical oscillations.
Furthermore, the parallels to the linguistic time references – the present progressive tense and the present perfect tense – have suggested that interactive processes constantly convey temporal information along with tense markers as the semiotic scaffolding for insider players, whose reading helps succeed in the adjustment of timing. Timing (v.) builds a momentum and guides the near future, for which the building of momentum indicates movement, and guiding the future indicates meaning. Unlike physical movements, the movement for time-ing accompanies the crossing between the different tenses.
Biological systems are actuated with meaning, and time in the E-series as a meaning procreator is synonymous with organizational, which is because of the system moving forward and sustaining its basic scheme, or supposedly the identity (Matsuno 2004) of the organisms-plus-environment. Thus, timing (v.) observed by insider players or participants in the living world – not by the external outsider observers – belongs to a different time series than our human clock (i.e., B-series time). Biological time is communicative and belongs to the E-series; thus, the assumption of B-series time – that is, one can measure the entity of time – cannot hold. Human clock time is just another local time.
Finally, the contrast between predetermined B-series time and the ongoing process of determining E-series time should be mentioned. Our concept of timing (v.) is based on the E-series in a practical discourse with the participants’ perspective so that the time operation remains local and context-bound. Conversely, timing (n.) is measured globally by an outsider observer in the B-series expressed in the present tense without the consideration of boundary conditions. Thus, timing (n.) belongs to the B-series, and timing (v.) belongs to the E-series. A comprehensive understanding of biological phenomena may require both the B-series and the E-series time. External measurement represents the single-handed mono-metric B-series that is all predetermined by itself, whereas internal measurement (Matsuno 2004, 2015) represents the dialogue-like dia-metric E-series that is constantly in the process of negotiation.
A novelty that we can claim for this paper may be that we have shown the meaning-based biological mechanism of how an adjustment of timing (v.) leads to another subsequent timing (v.) by increasing momentum and anticipating the near future by using the conceptual linkages between upbeat/downbeat and command/report.
References
Agrawal, A. A., Hasting, A. P., Johnson, M. T. J., Maron, J. L., & Salminen, J.-P. (2012). Insect herbivores drive real-time ecological and evolutionary change in plant populations. Science, 338, 113–116.
Aristotle. (1930). The works of Aristotle. Translated and Edited by D. Ross. Oxford University Press.
Augustine. (1961). Confessions. Translated by R. S. Pine-Coffin. Harmondsworth: Penguin Books.
Bassler, B. L., & Losick, R. (2006). Bacterially speaking. Cell, 125(2), 237–246. https://doi.org/10.1016/j.cell.2006.04.001.
Bateson, G. (1972). Steps to an ecology of mind. New York: Ballantine Books.
Cody, M. L., & Brown, J. H. (1969). Song asynchrony in neighboring bird species. Nature, 222, 778–780. https://doi.org/10.1038/222778b0.
Endo, J., Takanashi, T., Mukai, H., & Numata, H. (2019). Egg-cracking vibration as a cue for stink bug siblings to synchronize hatching. Current Biology, 29(1), 143–148. https://doi.org/10.1016/j.cub.2018.11.024.
Franklin, D. W., & Wolpert, D. M. (2011). Computational mechanisms of sensorimotor control. Neuron, 72, 425–442. https://doi.org/10.1016/j.neuron.2011.10.006.
Gagliano, M., Renton, M., Duvdevani, N., Timmins, M., & Mancuso, S. (2012). Out of sight but not out of mind: Alternative means of communication in plants. PLoS One, 7(5), e37382. https://doi.org/10.1371/journal.pone.0037382.
Gervasi, D. & Schiestl, F. P. (2017). Real-time divergent evolution in plants driven by pollinators. Nature Communications, 8, 14691. https://doi.org/10.1038/ncomms14691.
Hasty, C. (1997). Meter as rhythm. Oxford: Oxford University Press.
Heidegger, M. (1996). Being and time. Translated by J. Stambaugh. New York: State University of New York Press.
Hoffmeyer, J. (2015). Introduction: Semiotic scaffolding. Biosemiotics, 8, 153–158.
Kawato, M. (1999). Internal models for motor control and trajectory planning. Current Opinion in Neurobiology, 9, 718–727. https://doi.org/10.1016/s0959-4388(99)00028-8.
Luther, D. A. (2008). Signaller: Receiver coordination and the timing of communication in Amazonian birds. Biology Letters, 4, 651–654. https://doi.org/10.1098/rsbl.2008.0406.
Matsuno, K. (2004). Internal measurement in the present progressive tense and cohesion. Revue International de Philosophie, 228, 173–188.
Matsuno, K. (2013a). Toward accommodating biosemiotics with experimental sciences. Biosemiotics, 6, 125–141. https://doi.org/10.1007/s12304-012-9156-2.
Matsuno, K. (2013b). Making biological theory more down to earth. Progress in Biophysics and Molecular Biology, 113, 46–56. https://doi.org/10.1016/j.pbiomolbio.2013.03.004.
Matsuno, K. (2015). On the physics of the emergence of sensorimotor control in the absence of the brain. Progress in Biophysics and Molecular Biology, 119(3), 313–323. https://doi.org/10.1016/j.pbiomolbio.2015.08.004.
Matsuno, K. (2020). Making the onset of semiosis comprehensible with use of quantum physics. Biosemiotics, 13, 271–283. https://doi.org/10.1007/s12304-020-09377-w.
McTaggart, J. E. (1908). The unreality of time. Mind; A Quarterly Review of Psychology and Philosophy, 17, 456–473.
McTaggart, J. E. (1927). The nature of existence (Vol. II). Cambridge: Cambridge University Press.
Nomura, N., & Matsuno, K. (2016). Synchronicity as time: E-series time for living formations. Cybernetics and Human Knowing, 23(3), 69–77.
Nomura, N., Muranaka, T., Tomita, J., & Matsuno, K. (2018). Time from semiosis: E-series time for living systems. Biosemiotics, 11(1), 65–83. https://doi.org/10.1007/s12304-018-9316-0.
Nomura, N., Matsuno, K., Muranaka, T., & Tomita, J. (2019). How does time flow in living systems? Retrocausal scaffolding and E-series time. Biosemiotics, 12(2), 267–287. https://doi.org/10.1007/s12304-019-09363-x.
Pashalidou, F. G., Lambert, H., Peybernes, T., Mescher, M. C., & De Moraes, C. M. (2020). Bumble bees damage plant leaves and accelerate flower production when pollen is scarce. Science., 368(6493), 881–884. https://doi.org/10.1126/science.aay0496.
Ricoeur, P. (1984). Time and narrative (vol. 1). Translated by K. Mclaughlin and D. Pellauer. Chicago: University of Chicago.
Ruesch, J., & Bateson, G. (1951). Communication: The social matrix of psychiatry. New York: Norton.
Rutherford, S. T., & Bassler, B. L. (2012). Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harbor Perspectives in Medicine, 2(11), a012427. https://doi.org/10.1101/cshperspect.a012427.
Sacks, H., Schegloff, E. A., & Jefferson, G. (1974). A simplest systematics for the organization of turn-taking for conversation. Language, 50(4), 696–735.
Schwagmeyer, P. L., Mock, D. W., Lamey, T. C., Lamey, C. S., & Beecher, M. D. (1991). Effects of sibling contact on hatch timing in an asynchronously hatching bird. Animal Behaviour, 41(5), 887–894. https://doi.org/10.1016/S0003-3472(05)80355-0.
Spencer, R.-J., & Janzen, F. J. (2011). Hatching behavior in turtles. Integrative and Comparative Biology, 51, 100–110. https://doi.org/10.1093/icb/icr045.
Tinbergen, N. (1953). Social behaviour in animals. London: Chapman and Hall.
Uexküll, J., & Kriszat, G. (1957). A stroll through the worlds of animals and men. In C. H. Schiller (Ed.), Instinctive behavior: The development of a modern concept (pp. 5–80). New York: International University Press.
Warkentin, K. M. (2011). Environmentally cued hatching across taxa: Embryos respond to risk and opportunity. Integrative and Comparative Biology, 51(1), 14–25. https://doi.org/10.1093/icb/icr017.
Wolpert, D. M., Ghahramani, Z., & Jordan, M. I. (1995). An internal model for sensorimotor integration. Science, 269, 1880–1882. https://doi.org/10.1126/science.7569931.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nomura, N., Matsuno, K., Muranaka, T. et al. Toward a Practical Theory of Timing: Upbeat and E-Series Time for Organisms. Biosemiotics 13, 347–367 (2020). https://doi.org/10.1007/s12304-020-09398-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12304-020-09398-5