
Abstract
A network of 15 Surface Elevation Tables (SETs) at North Inlet estuary, South Carolina, has been monitored on annual or monthly time scales beginning from 1990 to 1996 and continuing through 2022. Of 73 time series in control plots, 12 had elevation gains equal to or exceeding the local rate of sea-level rise (SLR, 0.34 cm/year). Rising marsh elevation in North Inlet is dominated by organic production and, we hypothesize, is proportional to net ecosystem production. The rate of elevation gain was 0.47 cm/year in plots experimentally fertilized for 10 years with N&P compared to nearby control plots that have gained 0.1 cm/year in 26 years. The excess gains and losses of elevation in fertilized plots were accounted for by changes in belowground biomass and turnover. This is supported by bioassay experiments in marsh organs where at age 2 the belowground biomass of fertilized S. alterniflora plants was increasing by 1,994 g m−2 year−1, which added a growth premium of 2.4 cm/year to elevation gain. This was contrasted with the net belowground growth of 746 g m−2 year−1 in controls, which can add 0.89 cm/year to elevation. Root biomass density was greater in the fertilized bioassay treatments than in controls, plateauing at about 1,374 g m−2 and 472 g m−2, respectively. Growth of belowground biomass was dominated by rhizomes, which grew to 3,648 g m−2 in the fertilized treatments after 3 years and 1,439 g m−2 in the control treatments after 5 years. Depositional wetlands are limited by an exogenous supply of mineral sediment, whereas marshes like North Inlet could be classified as autonomous because they depend on in situ organic production to maintain elevation. Autonomous wetlands are more vulnerable to SLR because their elevation gains are constrained ultimately by photosynthetic efficiency.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Tidal marshes and mangroves are valued for their ecosystem services (Barbier et al. 2010; Boesch and Turner 1984; Costanza et al. 2008; Li et al. 2018; Zu Ermgassen et al. 2021). They improve water quality, attenuate waves (Anderson and Smith 2014; Fagherazzi 2014; Ma et al. 2013; Möller et al. 2014), reduce flooding, sequester carbon, and provide nursery habitat for fin and shellfish, aquatic invertebrates, and birds (Adams et al. 2021; Irving et al. 2011; Mazzocco et al. 2022; Nagelkerken et al. 2015). These ecosystems face existential threats from population growth and coastal development on one hand and climate change on the other. Coastal wetlands and human populations are on a collision course. It is estimated that in 2015 between 0.625 and 1.1 billion persons globally lived in the ≤ 10 m low-elevation coastal zone (MacManus et al. 2021). Thirty percent of the 2000 global population lives in the 100-year flood plain and the number is growing (Neumann et al. 2015). The greater threat of development or climate on a global scale has not been determined, but there is an interaction because of the need for coastal wetlands to migrate inland.
Coastal wetlands have kept pace with rising sea level for millennia (Gehrels 1999; Kelley et al. 1995; Kirwan et al. 2011; Redfield 1965), but a prevailing view is that their resilience is threatened by rising sea level, coastal development, reduced sediment delivery from major rivers, sinking of riverine deltas, and increased coastal erosion (Leuven et al. 2019; Murray et al. 2019; Osland et al. 2022). Elevation capital can extend the time a marsh maintains its areal extent but does not remove the long-term risk of drowning when marsh elevation cannot keep pace with sea-level rise (Langston et al. 2021; Törnqvist et al. 2021). Alternatively, rather than losses, a net gain in the extent of coastal wetlands is possible, if there is sufficient accommodation space and sediment supply remains at present levels (Kirwan et al. 2016; Schuerch et al. 2018). The question of resilience depends on (1) the relative importance of gains in marsh platform elevation versus transgression or inland migration and (2) the relative importance of gains of exogenous mineral sediments versus in situ primary production. The present paper examines the importance of minerals and organic production for the North Inlet estuary in South Carolina. We arrive at generalities that may help to identify coastal ecosystems at risk before they are lost. When identified, mitigating the risk of loss may be more successful than restoration.
The original work on elevation change in North Inlet marshes was a 2002 publication (Morris et al. 2002) that described how marshes keep pace with sea level. The model became known as the Marsh Equilibrium Model (MEM). The main points are that the productivity of marsh vegetation, Spartina alterniflora in this case, responds to changes in mean sea level (MSL). Note that taxonomists placed Spartina in the genus Sporobolus in 2014 (Bortolus et al. 2019; Peterson et al. 2014). Spartina alterniflora grows within a limited vertical range approximately between MSL and mean higher high water (MHHW) (McKee and Patrick 1988; Morris et al. 2013b), limited at the upper end by osmotic stress or interspecific competition (Bertness 1991) and at the lower end by hypoxia. There is a preferred or optimum elevation approximately in the middle of the range (Morris et al. 2013b). If marsh elevation is higher than the optimum, then the productivity and rate of elevation gain will increase with a rise in MSL, and an equilibrium ensues (Morris et al. 2002). If the elevation is suboptimal, productivity and gains will decrease with a rise in MSL, which leads to marsh loss.
We revisit the model in this paper and examine its predictions in light of 30 years of elevation change measurements in North Inlet. Also, we report the results of a bioassay experiment using marsh organs to directly test the hypothesis that there is an optimal relative elevation for primary production of S. alterniflora, the dominant salt marsh species on the southeast coast of North America. And we report results of the effects of fertilization of marsh plots designed originally with the expectation that fertilization would decrease marsh elevation because of enhanced decay of soil organic matter. This was quickly disproven, and the experiment was refocused on the effect of enhanced productivity on elevation gain. The observation that fertilization increased the rate of gain of marsh elevation (Morris et al. 2013a) inspired additional bioassay experiment designed to test the influence of nutrients and plant age on belowground production and biomass.
Methods
Surface Elevation Change
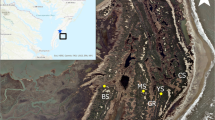
Elevation change was measured with a 9-pin Surface Elevation Table (SET) (Cahoon et al. 2002a, b) consisting of a portable arm that fits into a permanently installed benchmark pipe. SET sites were established in three groups (Fig. 1). For the first group, thirteen original-style SET benchmark pipes were installed early in 1990 or 1991 in six Spartina alterniflora-dominated salt marsh sites distributed across North Inlet by Dan Childers and Fred Sklar. SET measurements were made for two years as part of the NIN LTER program. The work was resumed by Morris and technicians in 1997 and has continued approximately annually through 2022. One benchmark post at each site was surveyed for elevation by the South Carolina Geodetic Survey in June 2001 using a Trimble RTK GPS. The remaining SET pipes were surveyed for elevation with a Trimble RTK GPS in April 2022. Boardwalks constructed at each site to access the pipes have deteriorated, but we have been careful not to trample the footprints of the SET pins when we visited the sites. Three of the original pipes eroded into creeks.

Location of SET pipes in North Inlet estuary, SC shown as green and turquoise bubbles. The bubble size is proportional to the rate of elevation change (turquoise is loss, green is gain). The locations of submerged SET platforms are denoted by skull and crossbones
For the second and third groups of sites, six original-style SET pipes were installed at Goat Island in North Inlet in the high- and low-marsh (spring 1996 and 2001, respectively). The 3 high-marsh pipes were driven 1 m below the sand/sediment interface to isolate the upper 1.0–1.5 m of the substrate where the biological processes of root growth and decomposition occur. The 3 low-marsh pipes were driven to the point of refusal. Each high-marsh pipe was established in the center of paired plots, consisting of a control plot on one side and a fertilized plot on the opposite side. One low-marsh SET pipe was surveyed by the South Carolina Geodetic Survey in June 2001. The remaining 5 pipes at Goat Island were surveyed for elevation with a Trimble RTK GPS in 2009. The high- and low-marsh SETs at Goat Island were first sampled in June 1996 and April 2001 (respectively) and continued to be measured approximately monthly through 2022. Boardwalks were installed to protect the SET platforms and are maintained. While we refer to the upper elevation sites at Goat Island as high marsh, they are in the zone occupied by a monoculture of S. alterniflora. The actual high marsh at North Inlet is a community of Salicornia, Borrichia, and Juncus. The choice of nomenclature is regrettable but has a historical precedent.
Each SET pipe has notches that receive the portable arm, so measurements can be made in 4 (first installation) or 6 (second and third installations) compass directions. To make measurements, the arm is placed on the pipe, leveled, and 9 fiberglass pins (the original pins were brass) were lowered to the sediment surface through a plate at the end of the arm until they touch, but do not penetrate the sediment surface. The pin lengths above the plate were measured to the nearest mm. This process was repeated at each compass heading, resulting in 4 subplot means in the original group of SET platforms and 6 subplot means at each of the Goat Island group of SET pipes. Elevation of the marsh surface, Z (cm NAVD) at each pin was calculated using the surveyed SET pipe elevation, the geometry of the SET apparatus, and pin length at each time point as Z = (E + S) − (PT − P), where E is the surveyed elevation of the top of the pipe (cm NAVD), S is the vertical distance between the SET plate and the top of the pipe (cm), PT is the total pin length (cm), and P is the pin length (cm) above the SET plate.
The rate of change of marsh elevation at each SET subplot was computed from a linear regression (SAS Institute Inc. 2009) of the mean subplot elevation versus time (Table S1). Subplot regression slopes were used when fitting the rate of elevation change (Eq. 1) to the entire collection using a static, nonlinear parameter estimation procedure. An assessment of the error of measurement of elevations using the SETs at Goat Island, made independently by two operators gave a mean of 2.4 mm (n = 36) for the absolute value of differences in estimates of subplot elevations, while at the plot and treatment level, the error was 1.3 mm (n = 12) (Morris et al. 2002). The amount of disturbance in each subplot (e.g., by crab burrows, mussels, or other means) can be gauged by the average variation (relative standard error) in 9 pin measurements at a single time point. Measured over all time the standard error of measurements was 0.5% of the mean in controls at Goat Island, 1% in fertilized plots, and 2.3% elsewhere.
Field SET Fertilization Experiment
One side of each of the three Goat Island high-marsh SETs (three SET arm positions each) was fertilized starting in June 1996 and ending in August 2004. Fertilizer was in the form of NH4NO3 plus P2O5 applied at a dosage of 30 mol N and 15 mol P m−2 year−1. The fertilizer was applied in granular form to four slices per m2 plot made in the marsh surface with a trowel and then closed. Fertilization in these plots with (NH4)2SO4 and P2O5 was resumed at the same dosage starting in May 2017 and continued through 2023. The frequency has varied from bimonthly through 2004, to monthly since 2017. The control and fertilized sides of the SET plot were separated by about 0.5 m and bisected by a boardwalk. The growth of the S. alterniflora plants on the control and fertilized sides was very different and similar to the growth in nearby control and fertilized plots used for biomass and productivity measurements, which indicates that there was no appreciable nutrient dispersion from fertilized to control plots.
Bioassay Experiments
Three experimental planters (Marsh Organs) described in (Morris 2007) were placed along a creekbank in North Inlet, Georgetown, SC. All were facing south to eliminate self-shading. The first marsh organ was established in 2002 and the first planting and first data collection were in 2005. Two additional marsh organs were established in 2006. Marsh organ pipes were constructed from varying lengths (in 15-cm increments) of 6″ diameter PVC. All of the PVC pipes are open at the bottom and grounded on a creekbank. Each marsh organ had between 6 and 8 levels; each level had 6 replicate pipes. The pipes were filled with sediment from the nearby marsh or creek at least one day before planting to allow for settling. Planting was accomplished by transplanting sediment cores containing S. alterniflora plants from the marsh nearby. The transplant cores were 10 cm in diameter by 23 cm in depth and contained plants of uniform size and density. Each pipe was planted with a core with its top flush with the top of the PVC pipe and packed with additional sediment. Elevations of every level of all marsh organs were measured in October 2009 using a Trimble R8 GNSS receiver, Trimble Ranger Handheld Computer, and Trimble HPB450 Radio. In 2021, the elevations ranged from ~ 15 cm above MSL to ~ 35 cm above MHHW. For marsh organ experiments with multiple rows of planters at 6 or more elevations and multiple years, depth was expressed as a dimensionless metric (DDim) computed as:
and corrected annually for changes in mean annual MHW and MLW.
From 2010 to 2016 an experiment examining the effects of plant age on growth and biomass of S. alterniflora from the marsh organs was initiated. All pipes in all three organs were planted in 2010. From 2010 through 2015 subsets of the pipes were harvested (and replanted the following spring), which resulted in plants of various ages occupying the marsh organ pipes through 2016. In 2016, all pipes were harvested. The ages of plants harvested during the 7 growing seasons of the experiment ranged from 1 to 5 years.
In 2007 and 2008, marsh organ experiments examining the effects of fertilization on growth and biomass of S. alterniflora were conducted. In 2007, all pipes in one marsh organ were fertilized with NH4NO3 plus P2O5, and all pots were harvested at the end of the growing season. In 2008, half of the pipes in each of two marsh organs were fertilized again with NH4NO3 and P2O5, and all pots of both organs were harvested at the end of the growing season. The fertilization rate was the same as that in the field plots (30 and 15 mol m−2 year−1 of N and P). Fertilizer was applied to individual organ pipes by pouring dry fertilizer pellets into a 20-cm-deep borehole through a funnel followed by a DI water rinse. The hole was then backfilled with sediment from the adjacent marsh.
In 2017 an experiment that examined permutations of fertilization and plant age was initiated. All pipes in two of the three marsh organs were fertilized with (NH4)2HPO4 as described above. Fertilizer was applied seasonally for the duration of the experiment. All pipes in the third marsh organ served as unfertilized controls. In 2017 and 2018 subsets of the pipes were harvested and replanted the following spring, which resulted in fertilized plants occupying the marsh organ pipes for 1–3 years. In 2019, all pipes were harvested.
Lignin Analyses
In February 2019, several whole plants were harvested from the north side of the North Inlet, Oyster Landing causeway. The plants were sorted into live roots, live rhizomes, and live leaves. The tissues were oven dried and about 30 g of each composited fraction was analyzed for lignin content by Dairy One Forage Testing lab in Ithaca NY. Klason lignin concentrations in roots, rhizomes, and leaves were 10.7%, 4.8%, and 5.9%, respectively. We use 10% in the model for lignin concentration and refractory fraction in the model (Morris et al. 2021).
Sediment Cores
Cores were collected, one from each of 3 census plots (Morris and Haskin 1990) in the low marsh and 3 in the high marsh at Goat Island. Cores were 7.6 cm diameter aluminum tubes by 30 cm in length. The sediment was extruded in 5 cm sections, dried at 60 °C, and combusted in a muffle furnace at 550 °C to determine organic matter concentration, expressed as loss-on-ignition (LOI).
The Coastal Wetland Equilibrium Model (CWEM)
The CWEM computes the change in surface elevation (dz/dt) as the sum of mineral sedimentation and net growth of belowground biomass:
where the organic and mineral contributions are \(\frac{d{z}_{org}}{dt}\mathrm{ and}\frac{d{z}_{min}}{dt}\). Both terms are functions of the marsh’s surface depth below the mean high water (MHW) level of the tide.
The net addition of organic matter (cm/year) is calculated as
Constants in the equation are the refractory fraction kr (g/g) the root-shoot ratio \(\xi\) (g/g), the turnover rate of belowground biomass \(\delta\) (y−1), and the self-packing density (Morris et al. 2016) k1 (g/cm3). Variable Bs is the maximum annual standing biomass (g/cm2). Refractory fraction is the portion of organic matter that is stable in an anoxic environment.
The inorganic sedimentation (g cm−2 year−1) is given by
Parameters q, m, f, D,\(\tau\), and k2 are the dimensionless capture coefficient, the concentration of suspended inorganic matter (g/cm3), flood frequency (y−1), depth below MHW (cm), inundation time (\(0\le \tau \le 1)\) (y/y), and self-packing density k2 (g/cm3), respectively.
The model was fitted to the time series of measured marsh elevations and rates of change using two procedures, depending on the availability of ancillary data. The annual mean elevation at SET benchmarks at Goat Island was fitted by integrating the model using a nonlinear, dynamic least-squares integration (SAS Institute Inc. 2010). The elevations were paired with observed water levels, mean suspended sediment concentrations, and biomass data first described in Morris and Haskin (1990). The sediment concentrations were measured by the NIWB NERR program, and water levels (1990–2022) at the nearby NOAA Springmaid Pier gauge (station 8661070).
A static nonlinear regression was also made to fit the model dz/dt to the measured slopes of the entire collection of SET time-series data. The slope (dz/dt) of each elevation time-series, one per subplot, was computed by linear regression (SAS Institute Inc. 2009), and the rates of elevation change (the slopes) were paired with the time-zero subplot elevations (Zo), MHW (70 cm), and mean sediment concentration (20 mg/L).
The maximum annual standing biomass \({B}_{S}\) was calculated as
where the coefficients a, b, and c are empirical constants taken from Miller et al. (2019) and Morris et al. (2013b). Depth D or (MHW-Z) was taken from the mean elevation of each SET subplot, and for each subplot there was a single rate of elevation change, single biomass, and single depth.
Results
Elevation Change
There were 15 SET platforms (pipes) in this study deployed among 7 sites between 1990 and 1996 (Fig. 1). Absolute elevation has increased at a majority of platforms since they were deployed but 5 have lost elevation including 3 that are now in creeks. All 3 lost SET pipes were close to creekbanks; otherwise, there has been no discernable spatial pattern of vertical elevation gains or losses. The lost SET sites appeared to succumb to bank erosion.
Of 73 elevation time series in control plots, 61 have elevation gains less than the rate of sea-level rise (0.34 cm/year, Fig. 2). Twelve have gains that exceed the rate of SLR (Reg Slope in Table S1). The average rate of elevation change for sites initially at elevations less than 10 cm NAVD was − 0.1 cm/year. The static fit of CWEM to elevation change across all these sites was statistically significant, and the negative rates observed at lower elevations were reflected in the negative capture coefficient q of − 0.75.

Distribution of rates of elevation change versus initial marsh elevation at the time of deployment of SET platforms. The least-squares fit of CWEM to SET controls and the recent rate of sea-level rise are also shown. A least-square fit of the model returned parameter values q of − 0.75 ± 0.42 (P < 0.08) and \(\xi\delta{k}_{r}\) of 0.13 ± 0.08 (P < 0.0001)
Our a priori expectation was that the refractory contribution of organic production kr should be about 0.1, based on the lignin concentration in live tissue. The total refractory contribution was calculated as the maximum standing aboveground biomass, given by Eq. 5, multiplied by the root and rhizome:shoot ratio \(\xi\)= 2, turnover rate \(\delta\) = 0.5, and the refractory fraction kr = 0.1, the product of which is 0.1. However, the fitted value of \(\xi \delta {k}_{r}\) = 0.66. The negative fitted value of q and the larger-than-expected value of \(\xi \delta {k}_{r}\) suggest that the marsh surface was eroding mineral sediment, at least at lower elevations, and that the major contribution to elevation gain was organic production. We build on this theme below with evidence from fertilization studies in the field and in bioassays.
The SET platforms in low and high marsh control sites at Goat Island (GI-5 and GI-18, Fig. 1) had vertical gains of 2.8 mm/y and 1.1 mm/y (Fig. 3), based on the fit of CWEM. The fertilized SET platforms in the high marsh had gains of 4.7 mm/y until the treatment was suspended in 2005. Fertilization was suspended for about 11 years and during that time the standing biomass and elevation declined over several years. The time series of elevations in the low marsh was more dynamic than control elevations in the high marsh. From 2005 to 2010, the low-marsh plots gained 2.5 cm (0.5 cm/year), and from 2010 to 2013, they lost 1.4 cm. Then from 2013 to 2021, they gained 5.7 cm (0.7 cm/year). Although the model accounted for 84% of the variation in elevation of the low-marsh SET plots, it was not able to simulate the short-term changes in elevation.

Annual mean marsh elevations (3 SETs and 2 subplots per treatment, 12 monthly measurements) made at SET stations at Goat Island, North Inlet, SC and results of a dynamic fit of CWEM (-—-). The procedure returned coefficient values of \(\xi\delta{k}_{r}\) (y−1) and q (g/g) equal to 0.11 ± 0.004 (P < 0.0001) and 3.1 ± 0.2 (± Std. Err., P < 0.0001), respectively. Constants entered as knowns were the self-packing densities k1 and k2 (0.085 and 1.99 g/cm3) (Morris et al. 2016)
Bioassay Experiments
Bioassay experiments in marsh organs (Fig. 4) carried out over 3 years showed clear evidence of (1) the existence of an optimum elevation for growth of both control and fertilized plants, expressed here as dimensionless depth (Eq. 1), (2) an increase with age in the aboveground and rhizome biomass in fertilized treatments, and (3) a higher optimum depth (lower relative elevation) in fertilized treatments compared to controls. Aboveground biomass of ages 1 and 3 plants differed significantly across planter depth \(\left({D}_{Dim}\right)\) as did age 1 live rhizomes (Table S2). The fertilization treatment had a significant effect on above- and belowground biomass for both age groups. We saw no evidence that maximum and minimum elevations differed between treatments and no evidence of a difference between the optimum elevation of rhizomes and aboveground biomass (Fig. 4) (Table S2).

Aboveground and live rhizome biomass of Spartina alterniflora at different relative elevations (dimensionless depth) and age in control and fertilized marsh organs. A Aboveground biomass at the end of one growing season (age 1). B Aboveground biomass after 3 growing seasons (age 3). C Live rhizome biomass after 3 growing seasons. The polynomial curves are from least-squares fitting of the model\(B={aD}_{Dim}+b{{D}_{Dim}}^{2}+c\). The optimum dimensionless depth is computed by solving for \({D}_{Dim}\) when the derivative \(dB/({dD}_{Dim} )\) is set to zero:\({D}_{opt}=-a/2b\). See Table S3 for coefficient values
The optimum dimensionless depth \({D}_{opt}\) for age 1 aboveground biomass in control treatments was 0.07 (Fig. 4), meaning it was close to the elevation of MHW (Eq. 1). After three years it was virtually unchanged at 0.04 (Fig. 4, Table S2). In the fertilized treatment \({D}_{opt}\) was 0.14 in year 1 and 0.27 in year 3, about midway between MHW and MSL. \({D}_{opt}\) for age 3 live rhizomes was 0.16 in fertilized plants, and the poor fit of the model precluded any conclusions being made about the other treatments (Table S2).
The maximum aboveground biomass from the polynomial fits of plant weights in the control and fertilized treatments in year 1 was 896 and 1,169 g/m2, respectively (Table S2). And by year 3, the maximum aboveground biomass had increased to 926 in controls and 5,281 g/m2 in fertilized plants (Fig. 4, Table S2). The trend in live rhizome biomass was similar. In year 3, the maximum rhizome biomass was 721 in controls and 3,109 g/m2 in the fertilized treatment. Maximum live rhizome biomass in year 1 was 1,075 g/m2 in the fertilized treatment and could not be determined in the controls (Table S2).
Averaged over all elevations, the growth rates of control and fertilized plants in marsh organs were significantly different (Fig. 5). In fertilized treatments, total plant biomass grew for 2 years at a rate of 3,949 g m−2 year−1. After 3 years the fertilization treatment was terminated because the rhizomes became pot-bound (Fig. 6). The total biomass of controls grew rapidly in year 1 to about 1,897 g m−2 and then quickly leveled off to an asymptotic level of about 2,994 g m−2. The asymptotic level is from a fit of the equation \(B=2994\left(1-{0.42}^{Age}\right)\) in Fig. 5A.

Total plant above and belowground biomass by age and treatment, composited across all elevations. A Time-series of the growth of total plant biomass by treatment. B Time-series of the growth of total belowground biomass (live and dead). C Stacked bar graph of control above and belowground biomass by age. D Stacked bar graph of fertilized above and belowground biomass by age. Geometric means and standard errors are in Table S2

Photo of third-year harvests of roots and rhizomes from A a washed and sorted control and B a washed, fertilized marsh organ pipe. The width was 15 cm. Length was about 35 cm. CT scans of cores taken from the field are shown in B a control and D a fertilized field site at Goat Island. CT scans were previously published (Wigand et al. 2015)
Most of the total growth was belowground, proportionally more so in rhizomes (Fig. 7). The net growth rate of total belowground biomass was 2,293 g m−2 y−1 in the fertilized treatment in the first 2 years or 63% of the total plant growth rate. In the controls, belowground biomass expanded quickly in the first year and then leveled off to an asymptotic level of 2,708 g m−2 (Fig. 5B). The net growth rate of total belowground biomass in controls was 758 g m−2 year−1 or 64% of the growth of total plant biomass. The mean ratio of total below:aboveground biomass was 2.8 ± 0.12 across age classes and treatments (Table S3).

Total root and rhizome biomass by age and treatment. A Time series of the growth of total rhizome biomass (live and dead) by treatment. Time series regressions were done on treatment means. B Time series of the growth of total root biomass (live and dead). C Stacked bar graph of control live and dead rhizome biomass by age. D Stacked bar graph of fertilized live and dead rhizome biomass by age. Geometric means and standard errors are in Table S2
Total rhizome biomass accounted for 0.4 ± 0.1 of total plant biomass and live rhizomes accounted for 0.42 ± 0.19 of total belowground biomass across all treatments. In the fertilized treatment total rhizome biomass increased until at least age 3 but was slowing in the third year when the experiment was terminated. Total rhizome biomass increased from 966 g m−2 at age 1 to 3,648 g m−2 at age 3. In the control treatment, total rhizome biomass increased from 731 g m−2 at age 1 to 1,437 g m−2 at age 5 (Fig. 7A).
The fractional turnover of roots was greater than the turnover of rhizomes and suggests that rhizomes are long-lived while roots probably turnover annually. The fraction live rhizome/total rhizome biomass in age 3 plants was 0.76 ± 0.03 in controls and 0.64 ± 0.14 in fertilized plants. This contrasts with the distribution of live and dead root biomass. In the controls, age 3 plants had 119 ± 19 g m−2 of live roots and 151 ± 16 g m−2 of dead roots. Fertilized plants had 182 ± 13 and 797 ± 64 g m−2 of live and dead roots (Table S3).
Sediment Organic Matter Content
The Goat Island site had measured LOI concentrations of 12.3% ± 0.44 (± 1 SD) to 13.4% ± 0.95 in the top 15 cm of low-marsh sediment, declining to 8.6% ± 1.4 in the 20–25-cm section. LOI in the high marsh was 20% ± 5.4 in the top 5 cm of the high marsh sediment, decreasing to 5.25% ± 2.9 at 20–25 cm. The depth-averaged LOI of the sediments in the modeled cohorts was 5.25% in high marsh control sites, 9% in fertilized high marsh, and 8.6% in the low marsh. The predicted values are net of live belowground biomass. The version of the CWEM used here did not simulate sediment cohorts. For an analysis with the cohort version of the model, see (Morris et al. 2022).
Discussion
Variation in Elevation Change
North Inlet is losing marsh, judging by the 3 lost SET sites (Fig. 1), and elevation gains less than the rate of SLR at a majority of sites (Fig. 2). We think the losses of SET sites were due to an increase in the tidal prism and consequent edge erosion. The rising tidal prism is chipping away at the margins, but when the platform drowns the loss will be relatively fast and cover a large area. Of 73 SET subplots monitored on control plots, 84% had elevation gains less than the local rate of SLR (0.34 cm/year); the mean rate of vertical change on these subplots was 0.08 ± 0.16 cm/year. The initial mean elevation of those sites was 17.9 ± 17.2 cm NAVD, which represents an elevation capital sufficient to maintain the marsh for 69 years at the current rates of mean elevation change and SLR. The 12 subplots with gains greater than the current rate of SLR had a mean gain of 0.43 ± 0.10 cm/year. At the current rate of SLR, those sites should be stable for more than a century. If the sea level accelerates in a century to a level 100 cm higher than today, CWEM predicts these sites should drown in about 75 years.
At North Inlet as in other estuaries like Poplar Island (Morris and Staver 2023), there is a great deal of variation or noise in elevation change, independent of the relative elevation (Fig. 2). There are hot spots for erosion and accretion. We cannot extract statistically the small mineral signal from the biological noise. Sediment is distributed as a function of inundation time and concentration. Inundation time varies seasonally and with astronomical cycles. Superimposed on this orderly behavior are the chaotic patterns imposed by fluid mechanics and shear stress. Regardless of the hydrodynamics, there is little opportunity for mineral sedimentation when relative elevation is high. If the marsh surface is at or above MHW, the delivery of sediment to the surface approaches zero (Freidrichs and Perry 2001; French and Spencer 1993; Krone 1985; Pethick 1981). Biovolume accounts for an increasingly greater share of elevation gain as relative marsh elevation increases and as the duration and depth of tidal flooding decrease. But there is a limit to organic accretion, and if the combination of subsidence and rising sea level exceeds organic and mineral gains, the wetland will drown sooner or later depending on the elevation capital.
The Dominance of Organic Production
After 5-y total plant biomass in control bioassays had reached a steady level of 2,994 g m−2 based on the fitted equation (Fig. 5A). Total belowground biomass had reached 2,761 g m−2 after 5 year and was still slowly rising (Fig. 5B). The most striking feature of these data is the large investment in belowground biomass relative to aboveground biomass in both controls and fertilized treatments. Based on those same regressions, total plant biomass in age 5 controls was 612 g m−2 greater than total belowground biomass, which gives a ratio of below:aboveground biomass was 3.8:1. This probably is inflated due to the decline in leaf biomass at the end of the growing season when marsh organs were harvested. The corresponding ratio in age 3 fertilized plants was 1.4:1, but the belowground biomass of age 3 fertilized plants was 2,911 g m−2 greater than age 5 controls. The model assumes a turnover of belowground biomass of 0.5 year−1 and preservation of 10%. Applying these to the total year-five belowground biomass of controls gives a production of refractory organic matter of 117 ·10−4 g cm−2 year−1 and dividing this by the particle density of organic matter, 0.084 g/cm3 (Morris et al. 2016), gives 0.14 cm/year, which is equal to the measured elevation gains on control SET plots of 0.14 ± 0.2 cm/year (Fig. 3).
From the sediment LOI, self-packing densities (Morris et al. 2016), and rate of elevation gain, it is possible to approximate the relative importance of organic and inorganic inputs. For example, the Goat Island high marsh had measured LOI concentrations near the surface of control plots of about 20% including live biomass, declining to about 5% below the root zone. For an elevation gain of 0.14 cm/year (Fig. 2) and assuming an average LOI of 12.5%, the required inputs of inorganic (y) and organic (x) material can be calculated from two equations. The elevation gain is x/k1 + y/k2 = 0.14 cm/year, where k1 = 0.084 g/cm3 and k1 = 1.99 g/cm3, and the LOI is x/(x + y) = 0.125 g/g. The calculated values are x = 0.0091 g cm−2 year−1 and y = 0.0635 g cm−2 year−1. The volumes in 0.14 cm of sediment are x/k1 = 0.11 cm3 organic matter per cm2 of marsh surface and y/k2 = 0.032 cm3 mineral matter per cm2 of marsh surface. Therefore, the required organic input is only 13% of the weight, but 77% of the volume.
The fit of CWEM to the SET data (Fig. 3) and marsh organ data (Figs. 5 and 7) indicate that marsh elevation change at North Inlet is dominated by biovolume production, driven by the growth and turnover of roots and rhizomes. The statistical insignificance (P = 0.08) and estimated error (± 0.42) of the capture coefficient q indicate that rates of elevation change across North Inlet estuary are not highly correlated with mineral inputs, or in this case, losses. Consequently, there is not a high degree of confidence in q. The static fit of the model to estuary-wide rates of elevation change (Fig. 2) gave a value of \(\xi \delta {k}_{r}\) = 0.65 ± 0.07 year−1 (defined in Eq. 3). The product \(\xi \delta {k}_{r}\) was highly significant (P < 0.0001), but there is not a high degree of confidence in the individual parameter values. Probably the greatest uncertainty is in the turnover rate \(\delta\). Our a priori expectation was for the product \(\xi \delta {k}_{r}\) to be close to 0.1 based on a root:shoot ratio of 2, a turnover rate of 0.5, and a refractory fraction of 10%. The refractory fraction is the proportion of the belowground organic production that is preserved. This is likely related to the lignin concentration in plant tissue, which for Spartina spp. is about 10% of dry weight (Buth and Voesenek 1987; Hodson et al. 1984; Wilson 1985). The fit of CWEM to the Goat Island elevations (Fig. 3) returned a value of \(\xi \delta {k}_{r}\) (0.13 ± 0.08 year−1) very close to the expected quantity. Assuming average standing biomass of 1,280 g/m2 (Miller et al. 2019), the expected vertical change \({k}_{r}\xi \delta {B}_{S}/{k}_{1}\) should be 0.198 cm/year [0.13 year−1 ·1280 g/m2 ·10−4 m2/cm2/(0.084 g/cm3)], which is close to the average observed rate of elevation change across all SET control subplots of 0.14 ± 0.20 cm/year (Fig. 2, Table S1) and indicates that organic production can account for the total average elevation gain.
Using a delta-growth geometric model, Lorenzo-Trueba et al. (2012) showed that the organic volume fraction in marsh sediment is maximized when the organic matter accumulation and accommodation rates are balanced. In equilibrium, the vertical gains and creation of accommodation space by rising sea level are equal. Inorganic sedimentation fills the fraction of the accommodation space not occupied by organics. Inorganic sediment cannot account for elevation gains in marshes that generate organic volume sufficient to fill the accommodation space. If SLR exceeds the organic contribution, relative marsh elevation will fall. At elevations super-optimal for growth, falling relative marsh elevation will result in rising rates of elevation gain until a new equilibrium is achieved. Rates of elevation gains in US East Coast marshes have risen during the past century as SLR has accelerated (Weston et al. 2023), which suggests that these marshes have been on the positive side of the growth curve or the side of suboptimal depth. If the depth below MHW of the marsh surface is greater than optimal, an increase in the rate of SLR will depress primary production, reduce elevation gain, and raise the inorganic sediment input, decreasing sediment LOI. Ultimately, the sediment composition and elevation gains are controlled by the rate of SLR and the creation of accommodation space, which control primary productivity and relative elevation.
The Fertilization Effect
Most growth was belowground and this is especially the case with fertilized plants in marsh organs that were gaining belowground biomass through age 2 at a rate of 1,994 g m−2 year−1, which could add 2.4 cm/year of elevation. This rate is 5 times greater than the elevation change we observed in the fertilized Goat Island SET plots (Fig. 3). This may be due to differences in the efficiency of fertilization between bioassay and field, and/or it may reflect the growth premium of plants in the marsh organ.
The fertilization experiments (Figs. 3, 4, 5, and 7) suggest that S. alterniflora biomass expands to the limit of its limiting nutrient supply or until some other resource limitation is reached, such as space, water, or light. Whether it is old field succession (Odum 1960), crop yield (Sinclair 1994), grassland productivity (Burke et al. 1997; Reich et al. 2006), forests fertilized with CO2 (Norby et al. 2010), or chronic, long-term marsh fertilization (Valiela et al. 2023), the productivity and biomass will increase to a higher level when the supply of limiting resources increases. The concept that marsh vegetation expands to fill its resource space is supported by field studies of marsh construction (Craft et al. 2003; Morris and Staver 2023) and fertilization experiments (Figs. 3, 4, 5, 6, and 7). The volume of soil on the fertilized Goat Island sites increased for 10 years at a rate four times that of the controls, and the evidence, including bioassay data, points to an expansion of belowground biomass as the primary mechanism, and rhizomes constituted the majority of belowground growth (Fig. 7). In the fertilized bioassay treatment, the growth of rhizomes appeared to be slowing by age 3 and had reached a biomass of 3,648 g m−2 (Fig. 7A). Based on the regression models, age 2 fertilized rhizomes were growing at a rate of 1,347 g m−2 year−1, or 68% of the growth of total belowground biomass, 1,994 g m−2 year−1 (Fig. 5B). In contrast, rhizome biomass in controls plateaued at about 1,254, almost within the first year.
The Growth Premium
The growth premium is the volume of sediment accrued by the expansion of belowground biomass during the early growth and development of a marsh (Morris et al. 2023; Morris and Staver 2023). The rapid change in elevation of the fertilized plots relative to controls (Fig. 3) is a good example. The rapid rise in elevation was due to the growth of belowground biomass, and the decline in elevation when nutrient additions were suspended was from the decay of labile necromass and reduction in live root and rhizome biomass. Volumes of coarse root and rhizome biomass in fertilized and control plots were 301 and 176 ml in sediment cores 20 cm in length and 15 cm in diameter (Wigand et al. 2015). For these cores, 178 cm2 in area, the difference in biovolume was (301–175.6) cm/178 cm2 = 0.7 cm. That difference represents the gain in the volume of live roots and rhizomes during the first phase of fertilization from 1996–2005. Then, starting in 2005, the biomass on fertilized plots gradually declined to the same level as controls until 2016 when fertilization was resumed. During that time (2005–2016) the elevation in the fertilized plots decreased by 1 cm while the controls were gaining 0.1 cm/year (Fig. 3). It follows that any increase in production will increase vertical elevation gain, whether by growth and development of a newly planted or colonized site (Morris and Staver 2023) or by an increase in resource supply.
Autonomous Marshes
Although relatively young marshes like North Inlet and Poplar Island (Morris and Staver 2023) have mineral-rich sediments by weight, they are nevertheless organogenic, self-regulating to a large extent, and essentially autonomous. In autonomous marshes, vertical gains will be proportional to net ecosystem production. Others depend on large, exogenous sediment loads from rivers or in some cases sediment scouring by macro tidal forces (van Proosdij et al. 2006). The distinction between the two modes of accretion is important. The autonomous type is probably more vulnerable to accelerating SLR except where flood control measures isolate depositional wetlands from their sediment supply, and vulnerability is especially high in marshes with little elevation capital. Autonomous and depositional wetland types represent the ends of a continuum, although the end members can flip from one mode to the other as can be seen in the alternating deposits of peat and silt in deep deltaic sediments (Törnqvist et al. 2008). The depositional type is more likely found in tropical latitudes where river basins can produce and transport more sediment than basins of similar scale located in the temperate zone (Syvitski et al. 2017). Currently, the greatest losses of marsh area, measured remotely, are due to edge erosion (Campbell et al. 2022), which is a consequence of rising sea level and the subsequent increase in the tidal prism volume (Gardner and Bohn 1980). This can be quantified with remote imagery whereas loss of elevation is not easily quantified until the marsh platform drowns.
The Role of Mineral Sediment
What is the role of mineral sediment in an autonomous type of marsh? Redfield (1972) summarized it when he wrote about marsh development on Cape Cod, saying that “…the size of the inlets and the currents through them are smaller and the supply of sediment to the basins is thus diminished. The result has been that the rate of deposit of sediment has not been sufficient in the face of rising sea level to fill the basins to a level where marsh can develop.” Mineral sedimentation establishes an elevation sufficient for colonization by marsh vegetation, and once established, organic production is largely responsible for raising the elevation to a higher plane. A good example was discussed by Cahoon et al. (2011). Once colonized, the vegetation becomes the dominant actor in generating new sediment volume with exceptions such as the mouths of large deltas (Falcini et al. 2012; Sidik et al. 2016). But the marshes must have access to those sediment loads (Boesch et al. 1994).
Nutrient Uptake
The asymptotic density of root biomass in both bioassay treatments must have been sufficient to supply the total plant growth with nutrients or to exhaust the nutrient supply. The fertilizer treatment was seasonal and continued throughout the experiment. The total growth rate of fertilized plants, based on the derivative of the regression evaluated at year 2, was estimated to be 2,625 g m−2 year−1 (Fig. 5A). With a tissue N of 1.8% (Ornes and Kaplan 1989), the N uptake would need to be about 47 g N m−2 year−1 compared to 210 g N m−2 year−1 added in fertilizer, which suggests that the fertilized plants were N-saturated. This level of nitrogen supply was able to support the growth of total plant biomass (Fig. 7B) for at least 3 years. Based on the average live root weight, 327 g m−2 (Table S3), the unit N uptake was about 0.14 g N year−1 g root−1, equivalent to 16 μg N hr−1 g root−1. This is less than the hourly uptake measured in the laboratory, 46.5 μg NH4-N hr−1 g root−1 (Morris and Dacey 1984), and half of the estimated Vmax of 32 μg NH4-N hr−1 g root−1 in the presence of sulfide (Bradley and Morris 1990). Such comparisons are difficult because growth and nutrient uptake are seasonal and diffusion gradients in sediment are steeper than in stirred vessels, but this level of agreement is still remarkable. In the controls, where the growth of total biomass over the first 2 year was 1,233 g m−2 year−1 and average live root biomass was 182 g/m2 (Fig. 5A, Table S3), the nitrogen uptake should be about 8 μg N hr−1 g root−1or 45% of the unit N-uptake in the fertilized treatment. The functional balance model of Davidson (1969) posits that the activities of roots and leaves are balanced and, therefore predicts that an increase in nutrient uptake must accompany an increase in total growth. Our results show that the increase in total N-uptake is accomplished by increases in both root biomass (Fig. 7B and Table 2S) and unit N-uptake.
The Original 2002 Model
The original MEM model (Morris et al. 2002) was highly abstracted: \(dZ/dt=(q+k{B}_{s})D\), and the contribution of organic production and sediment trapping on leaf surfaces were assumed to be proportional to biomass expressed by a single term, kBsD, where D is depth below MHW, Bs is maximum standing biomass, and k is an empirical constant. The model is elegant because it can be solved for the depth, D, that is in equilibrium with the rate of sea-level rise: \(dZ/dt=SLR\) or \(k{bD}^{3}+ka{D}^{2}+ \left(q+kc\right)D=SLR\) after substituting for Bs from Eq. 5. The 2002 study was done on Goat Island at an elevation high in the tidal frame where access by tides and sediments was limited. The conclusion about the importance of inorganic sedimentation was based on the erroneous assumption that kBD mostly represented inorganic sediment inputs, supported we thought by 210Pb analyses of 3 sediment cores (Vogel et al. 1996) and on mineral accumulation over marker horizons that had been in place for only 3 months. The marker horizon deposits were not substantial, not consolidated, and the relationship between the bulk density and volume (Morris et al. 2016) had not been worked out. The 1996 210Pb study reported that North Inlet sediments were 80% mineral by weight, which is consistent with the LOI concentration measured in the present study. Based on the mixing model (Morris et al. 2016) we estimate that minerals of this weight percentage should occupy only about 14% of the volume (0.8 g mineral/1.99 g cm−3 vs 0.2 g organic/0.084 g cm−3).
There are several important generalities from the 2002 study: (1) feedback between primary production and relative elevation moves marsh elevation toward an equilibrium with MSL, (2) relative elevation and productivity are determined by the rate of SLR, and (3) there is a relative elevation at which primary production and carbon sequestration are maximized, but this is also a tipping point. These generalities are still valid and supported by the present study. We conclude that kBD in the original model mostly represented organic inputs. CWEM does not resolve the difference between mineral deposition by settlement onto the marsh surface and by trapping on leaf surfaces. Direct measurements are needed to resolve this question because of the autocorrelation among biomass, sedimentation, and relative elevation.
Summary
Marsh elevations have been measured with SETs at North Inlet marshes beginning as early as 1990. The time series of elevations at these sites show that the majority are losing elevation relative to sea level. Rates of elevation change measured by linear regressions showed that 84% of the total have gained less than the local rate of SLR; 5 have lost elevation including 3 that have eroded into creeks. A field experiment showed that fertilization with N&P significantly increased elevation gain to about 4 × above controls due to the volume generated by the growth of belowground biomass. North Inlet marshes are in an autonomous class because organic production accounts for most vertical change and equilibrium with MSL. We hypothesize that elevation gain in an autonomous marsh is directly proportional to net ecosystem production. A marsh organ experiment showed that above- and belowground biomass have a growth range that spans the upper half of the tidal frame and an optimum relative elevation in the middle. A fertilization experiment showed (1) nutrients increased belowground biomass, including root biomass, and 2) rhizomes accounted for a majority of total belowground production, but on a fractional basis the turnover of roots was greater. Estimated NH4 uptake rates were consistent with laboratory measurements and suggest that plants in the fertilized marsh organs were N-saturated.
Data Availability
Data for this publication is available upon request from the corresponding author.
References
Adams, J.B., J.L. Raw, T. Riddin, J. Wasserman, and L. Van Niekerk. 2021. Salt marsh restoration for the provision of multiple ecosystem services. Diversity 13: 680.
Anderson, M.E., and J.M. Smith. 2014. Wave attenuation by flexible, idealized salt marsh vegetation. Coastal Engineering 83: 82–92.
Barbier, E., S. Hacker, C. Kennedy, E. Koch, A. Stier, and B. Silliman. 2010. The value of estuarine and coastal ecosystem services. Ecological Monographs.
Bertness, M.D. 1991. Zonation of Spartina patens and Spartina alterniflora in New England Salt Marsh. Ecology 72: 138–148.
Boesch, D.F., M.N. Josselyn, A.J. Mehta, J.T. Morris, W.K. Nuttle, C.A. Simenstad, and J.P. Swift. 1994. Scientific assessment of coastal wetland loss, restoration and management in Louisiana. Journal of Coastal Research Special Issue 20.
Boesch, D.F., and R.E. Turner. 1984. Dependence of fishery species on salt marshes: The role of food and refuge. Estuaries 7: 460–468.
Bortolus, A., P. Adam, J.B. Adams, M.L. Ainouche, D. Ayes, M.D. Bertness, T.J. Bouma, J.F. Bruno, I. Caçador, J.T. Carlton, J.M. Castillo, C.S.B. Costa, A.J. Davy, L. Deegan, B. Duarte, E. Figueroa, J. Gerwein, A.J. Gray, E.D. Grosholz, S.D. Hacker, A.R. Hughes, E. Mateos-Naranjo, I.A. Mendelssohn, J.T. Morris, A.F. Muñoz-Rodríguez, F.J.J. Nieva, L.A. Levin, B. Li, W. Liu, S.C. Pennings, A. Pickart, S. Redondo-Gómez, D.M. Richardson, A. Salmon, E. Schwindt, B.R. Silliman, E.E. Sotka, C. Stace, and M. Sytsma. 2019. Supporting spartina: Interdisciplinary perspective shows Spartina as a distinct solid genus. Ecology 100: e02863.
Bradley, P.M., and J.T. Morris. 1990. Influence of oxygen and sulfide concentration on nitrogen uptake kinetics in Spartina alterniflora. Ecology 71: 282–287.
Burke, I.C., W.K. Lauenroth, and W.J. Parton. 1997. Regional and temporal variation in net primary production and nitrogen mineralization in grasslands. Ecology 78: 1330–1340.
Buth, G.J.C., and L.A.C.J. Voesenek. 1987. Decomposition of standing and fallen litter of halophytes in a Dutch salt marsh. In Vegetation between land and sea: Structure and processes, ed. A.H.L. Huiskes, C.W.P.M. Blom and J. Rozema, 146–162. Dordrecht: Springer Netherlands.
Cahoon, D., J. Lynch, B. Perez, B. Segura, R. Holland, C. Stelly, G. Stephenson, and P. Hensel. 2002a. High-precision measurements of wetland sediment elevation: II. The rod Surface Elevation Table. Journal of Sedimentary Research - J SEDIMENT RES 72: 734–739.
Cahoon, D.R., J.C. Lynch, P. Hensel, R. Boumans, B.C. Perez, B. Segura, and J.W. Day Jr. 2002b. High-precision measurements of wetland sediment elevation: I. Recent improvements to the sedimentation-erosion table. Journal of Sedimentary Research 72: 730–733.
Cahoon, D.R., D.A. White, and J.C. Lynch. 2011. Sediment infilling and wetland formation dynamics in an active crevasse splay of the Mississippi River delta. Geomorphology 131: 57–68.
Campbell, A.D., L. Fatoyinbo, L. Goldberg, and D. Lagomasino. 2022. Global hotspots of salt marsh change and carbon emissions. Nature 612: 701–706.
Costanza, R., O. Pérez-Maqueo, M.L. Martinez, P. Sutton, S.J. Anderson, and K. Mulder. 2008. The value of coastal wetlands for hurricane protection. Ambio 37: 241–248.
Craft, C., P. Megonigal, S. Broome, R. Stevenson, R. Freese, J. Cornell, L. Zheng, and J. Sacco. 2003. The pace of ecosystem development of constructed Spartina alterniflora Marshes. Ecological Applications - ECOL APPL 13: 1417–1432.
Davidson, R.L. 1969. Effects of soil nutrients and moisture on root/shoot ratios in Lolium perenne L. and Trifolium repens L. Annals of Botany 33: 570–577.
Fagherazzi, S. 2014. Storm-proofing with marshes. Nature Geoscience 7: 701–702.
Falcini, F., N.S. Khan, L. Macelloni, B.P. Horton, C.B. Lutken, K.L. McKee, R. Santoleri, S. Colella, C. Li, G. Volpe, M. D’Emidio, A. Salusti, and D.J. Jerolmack. 2012. Linking the historic 2011 Mississippi River flood to coastal wetland sedimentation. Nature Geoscience 5: 803–807.
Freidrichs, C., and J. Perry. 2001. Tidal salt marsh morphodynamics. Journal of Coastal Research 27: 7–37.
French, J.R., and T. Spencer. 1993. Dynamics of sedimentation in a tide-dominated backbarrier salt marsh, Norfolk, UK. Marine Geology 110: 315–331.
Gardner, L.R., and M. Bohn. 1980. Geomorphic and hydraulic evolution of tidal creeks on a subsiding beach ridge plain, North Inlet, S.C. Marine Geology 34: M91–M97.
Gehrels, W.R. 1999. Middle and late holocene sea-level changes in eastern maine reconstructed from foraminiferal saltmarsh stratigraphy and AMS 14C dates on basal peat. Quaternary Research 52: 350–359.
Hodson, R., R. Christian, and A. Maccubbin. 1984. Lignocellulose and lignin in the salt marsh grass Spartina alterniflora: Initial concentrations and short-term, post-depositional changes in detrital matter. Marine Biology 81: 1–7.
Irving, A.D., S.D. Connell, and B.D. Russell. 2011. Restoring coastal plants to improve global carbon storage: Reaping what we sow. PLoS ONE 6: e18311.
Kelley, J.T., W.R. Gehrels, and D.F. Belknap. 1995. Late Holocene relative sea-level rise and the geological development of tidal marshes at Wells, Maine, U.S.A. Journal of Coastal Research 11: 136–153.
Kirwan, M.L., A.B. Murray, J.P. Donnelly, and D.R. Corbett. 2011. Rapid wetland expansion during European settlement and its implication for marsh survival under modern sediment delivery rates. Geology 39: 507–510.
Kirwan, M.L., S. Temmerman, E.E. Skeehan, G.R. Guntenspergen, and S. Fagherazzi. 2016. Overestimation of marsh vulnerability to sea level rise. Nature Climate Change 6: 253–260.
Krone, R.B. 1985. Simulation of marsh growth under rising sea levels. In Hydraulics and hydrology in the small computer age., ed. W.R. Waldrop, 106–115. Lake Buena Vista, FL: Hydraulics Div. ASCE.
Langston, A.K., C.R. Alexander, M. Alber, and M.L. Kirwan. 2021. Beyond 2100: Elevation capital disguises salt marsh vulnerability to sea-level rise in Georgia, USA. Estuarine, Coastal and Shelf Science 249: 107093.
Leuven, J.R., H.J. Pierik, and M.v.d. Vegt, T.J. Bouma, and M.G. Kleinhans. 2019. Sea-level-rise-induced threats depend on the size of tide-influenced estuaries worldwide. Nature Climate Change 9: 986–992.
Li, X., R. Bellerby, C. Craft, and S.E. Widney. 2018. Coastal wetland loss, consequences, and challenges for restoration. Anthropocene Coasts 1: 1–15.
Lorenzo-Trueba, J., V. Voller, C. Paola, R. Twilley, and A. Bevington. 2012. Exploring the role of organic matter accumulation on delta evolution. Journal of Geophysical Research (Earth Surface) 117.
Ma, G., J.T. Kirby, S.-F. Su, J. Figlus, and F. Shi. 2013. Numerical study of turbulence and wave damping induced by vegetation canopies. Coastal Engineering 80: 68–78.
MacManus, K., D. Balk, H. Engin, G. McGranahan, and R. Inman. 2021. Estimating population and urban areas at risk of coastal hazards, 1990–2015: How data choices matter. Earth Syst. Sci. Data 13: 5747–5801.
Mazzocco, V., T. Hasan, S. Trandafir, and E. Uchida. 2022. Economic value of salt marshes under uncertainty of sea level rise: A case study of the Narragansett Bay. Coastal Management 50: 306–324.
McKee, K.L., and W.L. Patrick. 1988. The relationship of smooth. cordgrass ( Spartina alterniflora ) to tidal datums: A review. Estuaries 11: 143–151.
Miller, G.J., J.T. Morris, and C. Wang. 2019. Estimating aboveground biomass and its spatial distribution in coastal wetlands utilizing Planet Multispectral Imagery. Remote Sensing 11: 2020.
Möller, I., M. Kudella, F. Rupprecht, T. Spencer, M. Paul, B.K. van Wesenbeeck, G. Wolters, K. Jensen, T.J. Bouma, M. Miranda-Lange, and S. Schimmels. 2014. Wave attenuation over coastal salt marshes under storm surge conditions. Nature Geoscience 7: 727–731.
Morris, J.T. 2007. Estimating net primary production of salt-marsh macrophytes. In Principles and Standards for Measuring Primary Production, ed. T.J. Fahey and A.K. Knapp, 106–119. Oxford: Oxford University.
Morris, J.T., and J.W.H. Dacey. 1984. Effects of O2 on ammonium uptake and root respiration by Spartina alterniflora. American Journal of Botany 71: 979–985.
Morris, J.T., and B. Haskin. 1990. A 5-y record of aerial primary production and stand characteristics of Spartina alterniflora. Ecology 71: 2209–2217.
Morris, J.T., D.C. Barber, J.C. Callaway, R. Chambers, S.C. Hagen, C.S. Hopkinson, B.J. Johnson, P. Megonigal, S.C. Neubauer, T. Troxler, and C. Wigand. 2016. Contributions of organic and inorganic matter to sediment volume and accretion in tidal wetlands at steady state. Earth’s Future 4: 110–121.
Morris, J.T., D.R. Cahoon, J.C. Callaway, C. Craft, S.C. Neubauer, and N.B. Weston. 2021. Marsh equilibrium theory: Implications for responses to rising sea level. In Salt Marshes: Function, Dynamics, and Stresses, ed. D.M. FitzGerald and Z.J. Hughes, 157–177. Cambridge: Cambridge University Press.
Morris, J.T., J.Z. Drexler, L.J.S. Vaughn, and A.H. Robinson. 2022. An assessment of future tidal marsh resilience in the San Francisco Estuary through modeling and quantifiable metrics of sustainability. Frontiers in Environmental Science 10.
Morris, J.T., J.A. Langley, W.C. Vervaeke, N. Dix, I.C. Feller, P. Marcum, and S.K. Chapman. 2023. Mangrove trees outperform saltmarsh grasses in building elevation but collapse rapidly under high rates of sea-level rise. Earth's Future 11: e2022EF003202.
Morris, J.T., and L.W. Staver. 2023. Modeling elevation change on constructed Poplar Island, Chesapeake, MD marshes: II. The importance of marsh development time Estuarine and Coasts in review.
Morris, J.T., P.V. Sundareshwar, C.T. Nietch, B. Kjerfve, and D.R. Cahoon. 2002. Responses of coastal wetlands to rising sea level. Ecology 83: 2869–2877.
Morris, J.T., G.P. Shaffer, and J.A. Nyman. 2013a. Brinson Review: perspectives on the influence of nutrients on the sustainability of coastal wetlands. Wetlands 33: 975–988. https://doi.org/10.1007/s13157-013-0480-3.
Morris, J.T., K. Sundberg, and C.S. Hopkinson. 2013b. Salt marsh primary production and its responses to relative sea level and nutrients in estuaries at Plum Island, Massachusetts, and North Inlet, South Carolina, USA. Oceanography 26: 78–84.
Murray, N.J., S.R. Phinn, M. DeWitt, R. Ferrari, R. Johnston, M.B. Lyons, N. Clinton, D. Thau, and R.A. Fuller. 2019. The global distribution and trajectory of tidal flats. Nature 565: 222–225.
Nagelkerken, I., M. Sheaves, R. Baker, and R.M. Connolly. 2015. The seascape nursery: A novel spatial approach to identify and manage nurseries for coastal marine fauna. Fish and Fisheries 16: 362–371.
Neumann, B., A.T. Vafeidis, J. Zimmermann, and R.J. Nicholls. 2015. Future coastal population growth and exposure to sea-level rise and coastal flooding–a global assessment. PLoS ONE 10: e0118571.
Norby, R.J., J.M. Warren, C.M. Iversen, B.E. Medlyn, and R.E. McMurtrie. 2010. CO<sub>2</sub> enhancement of forest productivity constrained by limited nitrogen availability. Proceedings of the National Academy of Sciences 107: 19368–19373.
Odum, E.P. 1960. Organic Production and Turnover in Old Field Succession. Ecology 41: 34–49.
Ornes, W.H., and D.I. Kaplan. 1989. Macronutrient status of tall and short forms of Spartina alterniflora in a South Carolina salt marsh. Marine Ecology Progress Series 55: 63–72.
Osland, M.J., B. Chivoiu, N.M. Enwright, K.M. Thorne, G.R. Guntenspergen, J.B. Grace, L.L. Dale, W. Brooks, N. Herold, J.W. Day, F.H. Sklar, and C.M. Swarzenzki. 2022. Migration and transformation of coastal wetlands in response to rising seas. Science Advances 8: eabo5174.
Peterson, P.M., K. Romaschenko, Y.H. Arrieta, and J.M. Saarela. 2014. A molecular phylogeny and new subgeneric classification of Sporobolus (Poaceae: Chloridoideae: Sporobolinae). Taxon 63: 1212–1243.
Pethick, J.S. 1981. Long-term accretion rates on tidal salt marshes. Journal of Sedimentary Research 51: 571–577.
Redfield, A.C. 1965. Ontogeny of a Salt Marsh Estuary. Science 147: 50–55.
Redfield, A.C. 1972. Development of a New England salt marsh. Ecological Monographs 42: 201–237.
Reich, P.B., S.E. Hobbie, T. Lee, D.S. Ellsworth, J.B. West, D. Tilman, J.M. Knops, S. Naeem, and J. Trost. 2006. Nitrogen limitation constrains sustainability of ecosystem response to CO2. Nature 440: 922–925.
SAS Institute Inc. 2009. Introduction to regression procedures. In SAS/STAT ® 9.2 User’s Guide, 72–111. Cary, NC: SAS Institute Inc.
SAS Institute Inc. 2010. The MODEL Procedure. In SAS/ETS® 9.22 User’s Guide. Cary, NC: SAS Institute.
Schuerch, M., T. Spencer, S. Temmerman, M.L. Kirwan, C. Wolff, D. Lincke, C.J. McOwen, M.D. Pickering, R. Reef, A.T. Vafeidis, J. Hinkel, R.J. Nicholls, and S. Brown. 2018. Future response of global coastal wetlands to sea-level rise. Nature 561: 231–234.
Sidik, F., D. Neil, and C.E. Lovelock. 2016. Effect of high sedimentation rates on surface sediment dynamics and mangrove growth in the Porong River, Indonesia. Marine Pollution Bulletin 107: 355–363.
Sinclair, T.R. 1994. Limits to Crop Yield? In Physiology and Determination of Crop Yield, 509–532.
Syvitski, J., A. Kettner, I. Overeem, G. Brakenridge, and S. Cohen. 2017. Latitudinal controls on siliciclastic sediment production and transport, 1–15. Tulsa, OK, USA: Latitudinal Controls on Stratigraphic Models and Sedimentary Concepts; SEPM Special Publication.
Törnqvist, T.E., D.R. Cahoon, J.T. Morris, and J.W. Day. 2021. Coastal wetland resilience, accelerated sea-level rise, and the importance of timescale. AGU Advances 2: e2020AV000334.
Törnqvist, T.E., D.J. Wallace, J.E.A. Storms, J. Wallinga, R.L.V. Dam, M. Blaauw, M.S. Derksen, C.J.W. Klerks, C. Meijneken, and E.M.A. Snijders. 2008. Mississippi Delta subsidence primarily caused by compaction of Holocene strata. Science & Engineering Faculty.
Valiela, I., K. Chenoweth, J. Lloret, J. Teal, B. Howes, and D. Goehringer Toner. 2023. Salt marsh vegetation change during a half-century of experimental nutrient addition and climate-driven controls in Great Sippewissett Marsh. Science of the Total Environment 867: 161546.
van Proosdij, D., R.G.D. Davidson-Arnott, and J. Ollerhead. 2006. Controls on spatial patterns of sediment deposition across a macro-tidal salt marsh surface over single tidal cycles. Estuarine, Coastal and Shelf Science 69: 64–86.
Vogel, R.L., B. Kjerfve, and L.R. Gardner. 1996. Inorganic sediment budget for the North Inlet salt marsh, South Carolina, U.S.A. Mangroves and Salt Marshes 1: 23–35.
Weston, N.B., E. Rodriguez, B. Donnelly, E. Solohin, K. Jezycki, S. Demberger, L.A. Sutter, J.T. Morris, S.C. Neubauer, and C.B. Craft. 2023. Recent Acceleration of wetland accretion and carbon accumulation along the U.S. East Coast. Earth's Future 11: e2022EF003037.
Wigand, C., E. Davey, R. Johnson, K. Sundberg, J. Morris, P. Kenny, E. Smith, and M. Holt. 2015. Nutrient effects on belowground organic matter in a minerogenic salt marsh, North Inlet, SC. Estuaries and Coasts 38: 1838–1853.
Wilson, J.O. 1985. Decomposition of [14C] lignocelluloses of Spartina alterniflora and a comparison with field experiments. Applied and Environmental Microbiology 49: 478–484.
Zu Ermgassen, P.S., R. Baker, M.W. Beck, K. Dodds, and S.O. zu Ermgassen, D. Mallick, M.D. Taylor, and R.E. Turner. 2021. Ecosystem services: Delivering decision-making for salt marshes. Estuaries and Coasts 44: 1691–1698.
Acknowledgements
Support from the US National Science Foundation is gratefully acknowledged. The work described here has been possible only with the assistance of numerous individuals including Dan Childers and Fred Sklar who installed the first SET platforms at North Inlet with support from the NIN LTER, then Don Cahoon who helped install the Goat Island SET platforms with support from the USGS, and students, including James Edwards, Gwen Miller, Brant Priest, PV Sundareshwar, Siobhan Kitchen, Weihong Zanazzi, Whitney Kiehn, and Emily Van Seeters, technicians Betsy Haskin, Robin Crest, Melissa Day, Jeff Jefferson, and Warren Hankinson, and visiting scientists Karim Alizad, Yihui Zhang, Katherine Renken, and Chuan Tong.
Funding
This study was supported by funds from the US National Natural Science Foundation LTREB (grant DEB 1654853).
Author information
Authors and Affiliations
Contributions
JM: writing—original draft, review and editing, modeling, data analysis, funding, methodology, and project administration. KS: review and editing, methodology, data acquisition, and curation.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Communicated by Nathan Waltham
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Morris, J.T., Sundberg, K. Responses of Coastal Wetlands to Rising Sea-Level Revisited: The Importance of Organic Production. Estuaries and Coasts (2024). https://doi.org/10.1007/s12237-023-01313-8
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12237-023-01313-8