
Summary
A taxonomic revision is presented of the Nengella group of the palm genus Hydriastele (Arecaceae: Arecoideae) in New Guinea, which comprises slender understorey or midstorey palms with small, protandrous inflorescences. Seven species are accepted: Hydriastele aprica, H. divaricata sp. nov., H. flabellata, H. montana, H. pinangoides, H. simbiakii sp. nov., and H. splendida sp. nov. All species are illustrated and their distributions are mapped. In addition, a key to species and preliminary IUCN Red List Assessments are provided.
Similar content being viewed by others


Introduction
Hydriastele H. Wendl. & Drude (Arecoideae: Areceae) is a highly variable palm genus that comprises 49 accepted species (Govaerts et al. 2017), distributed from Sulawesi in Indonesia, through Papuasia to northern Australia, Fiji and Palau (Dransfield et al. 2008). The genus is most diverse in New Guinea where some 30 species are reported to occur (Baker & Couvreur 2012), including minute, understorey species, through midstorey palms to canopy emergents. In this paper, as part of ongoing research on New Guinea palms, we present a taxonomic revision of one lineage of Hydriastele in New Guinea, a monophyletic group (Loo et al. 2006), termed here the Nengella group.
The Nengella group consists of slender palms (stem diameter 0.5 – 7.5 cm), typically of the understorey and midstorey (up to 10 m height) of lowland to lower montane rainforests. They have protandrous inflorescences that are spicate or branched to one order with up to 6 rachillae (usually fewer), bearing triads that are usually spirally arranged. The male flowers tend to be showy, displaying shades of scarlet, pink, purple and lavender, and in some species even the inflorescence branches are brightly coloured. The female flowers have free sepals and petals, the petals bearing a triangular limb at the tip. The leaves often have conspicuously cuneate leaflets with prominent praemorse distal margins, although this is not universally the case, and young leaves are often bronze-coloured on emergence. Prior to this treatment, the Nengella group consisted of 11 accepted species, including one of the commonest understorey species in New Guinea, Hydriastele pinangoides (Becc.) W. J. Baker & Loo (Young 1985; Dowe & Ferrero 2000a; Baker & Loo 2004). Our monograph reduces these 11 species to four (H. aprica (B. E. Young) W. J. Baker & Loo, H. flabellata (Becc.) W. J. Baker & Loo, H. montana (Becc.) W. J. Baker & Loo and H. pinangoides); and includes three further species described as new here (H. divaricata, H. simbiakii and H. splendida).
Taxonomic history
Beccari erected the genus Nengella Becc. in 1877 when he described the species Nengella montana Becc. and Nengella flabellata Becc., along with Nenga pinangoides Becc. and Nenga affinis Becc. The genus Nenga H. Wendl. & Drude had already been established in 1875 (Wendland & Drude 1875), and to reflect the distinctive morphology in flowers and foliage displayed by Nenga pinangoides and Nenga affinis, Beccari further classified these taxa within ‘Subgenus II Gronophyllum Scheff.’ along with a species from Sulawesi, Nenga selebica Becc. Gronophyllum Scheff., which had been described as a genus in 1876 by Scheffer, was thus given infra-generic status by Beccari (1877). In 1885, Leptophoenix Becc. was erected and combined with Nenga pinangoides and Nenga affinis, whilst Nenga selebica was recombined in Gronophyllum, which was reassigned generic status (Beccari 1885). In the following decades, new species were described in either Leptophoenix or Nengella, until Burret (1936) eventually sank the former into the latter. Burret regarded the main difference between the genera, the condition of the seed endosperm (ruminate in Leptophoenix, homogeneous in Nengella), too trivial to warrant a generic distinction. Essig & Young (1985), encouraged by John Dransfield (pers. comm.), later sunk Nengella into Gronophyllum noting that a continuum of variation connected the two genera and that their circumscriptions had a geographical bias, meaning that the name Nengella seemed to be applied only to New Guinea species, despite Nengella-like palms being known from outside New Guinea in Sulawesi and the Moluccas. We here define the ‘Nengella group’ nomenclaturally to comprise all species that were referred to the genus Nengella immediately before Essig & Young (1985) recombined it, plus two further species that were described directly in Gronophyllum after this point, G. apricum B. E. Young (1985) and G. cariosum Dowe & M. D. Ferrero (2000a).
Following detailed molecular phylogenetic studies, which found Gronophyllum not to be monophyletic, Loo et al. (2006) placed Gronophyllum with Gulubia Becc. and Siphokentia Burret in synonymy within a greatly broadened genus Hydriastele H. Wendl. & Drude (Baker & Loo 2004). In addition, this study indicated that the Nengella group is monophyletic and that its closest relatives may be Hydriastele brassii (Burret) W. J. Baker & Loo, which resembles a very large species of the Nengella group, and H. beguinii (Burret) W. J. Baker & Loo and H. dransfieldii (Hambali, Maturb., Wanggai & W. J.Baker) W. J. Baker & Loo, which were formerly placed in the genus Siphokentia (Baker et al. 2000). It should be noted that species from Sulawesi (H. selebica (Becc.) W. J. Baker & Loo, H. nannostachys W. J. Baker & Loo, H. kjellbergii (Burret) W. J. Baker & Loo, H. sarasinorum (Burret) W. J. Baker & Loo,) and the Moluccas (H. microcarpa (Scheff.) W. J. Baker & Loo, H. oxypetala (Burret) W. J. Baker & Loo) that may belong to the Nengella group were not sampled by Loo et al. (2006). These species remain under-collected and very poorly known, and in the absence of further evidence, we exclude them from this revision, treating the Nengella group here as restricted to New Guinea and immediately adjacent islands.
Key to the species of the Nengella group
-
1. Triads spirally arranged with distinct alternating verticils of 3; rachillae often displaying shades of pink; endosperm ruminate .................................................................................................... 2
-
Triads spirally arranged with more or less well-defined verticils, or opposite and decussately arranged; rachillae normally yellow to green; endosperm homogeneous .................................................................................................... 5
-
2. Bearing c. 4 – 5 leaves per crown; leaf lamina entire bifid; inflorescence with 3 – 4 rachillae .................................................................................................... 7. H. splendida
-
Bearing 5 – 10 leaves per crown; leaf lamina pinnately compound; inflorescence with 2 – 5 (– 6) rachillae .................................................................................................... 3
-
3. Clustering palm; stem pliable and often leaning; leaflets 14 – 16 per side, regularly arranged, linear; restricted to river banks .................................................................................................... 6. H. simbiakii
-
Clustering or solitary palm; stem rigid and erect; leaflets 4 – 13 per side, regularly or irregularly arranged, linear or cuneate; occurrence not limited to river banks .................................................................................................... 4
-
4. Clustering palm to 4.5 m tall; stem diam. 0.9 – 1.7 cm; leaflets 9 – 12 per side, rather uniform in shape and size, linear and sub-regularly arranged, never grouped, basal and middle leaflets divaricate, i.e. appearing to be extending nearly perpendicular to the leaf rachis; inflorescence spicate or with 2 rachillae .................................................................................................... 2. H. divaricata
-
Clustering or solitary palm to 7 (– 10) m tall; stem diam. 1.5 – 7.5 (rarely thinner in some high-altitude forms); leaflets 5 – 13 per side, very variable in shape and size, broadly cuneate and irregularly arranged with several distinct groups usually present or more rarely narrowly cuneate and ± regularly arranged; inflorescence with 2 – 5 (– 6) rachillae .................................................................................................... 5. H. pinangoides
-
5. Solitary palm; stem diam. c. 3 cm; lamina with 15 – 23 cuneate leaflets per side, leaflets abaxially with ramenta on midrib; inflorescence 15 – 26 cm long, with 4 – 6 rachillae; triads opposite and decussate; fruits subglobose .................................................................................................... 1. H. aprica
-
Clustering or gregarious palm; stem diam. 0.5 – 2 cm; lamina entire bifid or pinnate with 2 – 6 cuneate leaflets or c. 9 – 11 leaflets per side, ramenta apparently lacking; inflorescence 7 – 15 cm long, spicate (more rarely with 2 rachillae); triads spirally arranged; fruits ellipsoid to fusiform .................................................................................................... 6
-
6. Gregarious palm; stem 5 – 8 mm in diam.; petiole c. 3 – 8 cm long; lamina with c. 9 – 11 regularly arranged, narrowly linear leaflets; rachilla bracts ± conspicuous .................................................................................................... 4. H. montana
-
Clustering palm; stem 1 – 2 cm in diam.; petiole 10 – 30 cm long; lamina entire bifid or with up to 6 regularly or irregularly arranged, cuneate leaflets; rachilla bracts often conspicuously cucullate .................................................................................................... 3. H. flabellata
Taxonomic treatment
1. Hydriastele aprica (B. E. Young) W. J. Baker & Loo (2004: 62). Gronophyllum apricum B. E. Young (1985: 139). Type: Papua New Guinea, Sandaun, Telefomin, Essig & Young LAE 74082 (holotype LAE!, isotype BH).
Solitary, slender palm to 5 m tall, bearing c. 7 leaves in crown. Stem c. 3 cm in diam. Leaves 48 – 80 cm long including petiole; sheath 21 – 23 cm long, sparsely covered with small brown to purple lacerate-peltate scales; petiole 12 – 18 cm long, indumentum as sheath; rachis 36 – 62 cm long, indumentum as sheath; leaflets 15 – 23 each side of rachis, irregularly arranged and variously grouped, borne 0 – 5.5 cm apart, drooping, cuneate, praemorse apically, discolorous, with ramenta present on the abaxial side of the midrib; basal leaflets 20 – 24 × 3 – 3.5 cm; middle leaflets 12 – 13 × 2 – 2.5 cm; terminal leaflets 8 – 10 × 1 – 2 cm. Inflorescences 15 – 26 cm long including 3.2 – 5.5 cm peduncle, branched to 1 order; prophyll 21 – 29 cm long, purplish, indumentum as sheath; rachillae 14 – 25 × 0.1 – 0.3 cm; triads opposite and decussate. Staminate flowers 3 – 7 × 0.5 – 3 mm in bud (when dry), cream-coloured with purplish tips; calyx 0.5 – 1 × 0.5 – 1 mm, consisting of three basally connate sepals; petals 3 – 7 × 0.5 – 3.0 mm, lanceolate and valvate; stamens 6; anthers 0.5 – 2.5 × 0.1 – 0.5 mm, filaments up to 1 × up to 0.2 mm; pistillodes 1 – 3 lobes. Pistillate flowers 1.5 – 7 × 1 – 2 mm in bud (when dry), dark purple; sepals 0.5 – 2 × 0.5 – 2 mm, imbricate; petals 1 – 7 × 0.5 – 2, broadly imbricate, with long and thin, valvate tips; ovary 1 – 2.5 × 0.5 – 1.5, ellipsoidal; staminodes c. 3, tooth-like. Fruits c. 8 × 7 mm, subglobose, red. Seeds c. 5 × 4 mm; endosperm homogeneous (Fig. 1).

Hydriastele aprica. A leaf diagram; B leaf apex; C mid-leaf portion; D inflorescence; E staminate flower whole and in longitudinal section; F pistillate flower; G fruit whole and in longitudinal section. Scale bar: A = 18 cm; B – C = 4 cm; D = 3 cm; E – G = 4 mm. A, F – G from Essig & Young LAE 74049; B – E from Essig & Young LAE 74072. drawn by lucy t. smith.
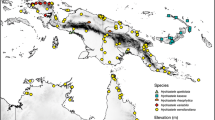
distribution. Known from a few localities in a remote part of northern central New Guinea (Map 1).

Distribution map of Hydriastele aprica.
specimens examined. papua new guinea. Sandaun Province: Telefomin Subprovince, on ridge above junction of “clear-water” stream with Frieda R., c. 2 km upstream from Carpentaria Exploration Company airstrip camp, 300 m, 26 April 1978, Essig & Young LAE 74049 (BH, LAE!, USF); Telefomin Subprovince, Rainforest below Carpentaria Exploration Co. helipad K-27 on exposed ridge, [4°45'S 141°50'E], 900 m, 1 May 1978, Essig & Young LAE 74082 (BH; LAE!); On exposed ridge near Carpentaria Exploration Co. “Antap Mt” helipad, [4°45'S 141°50'E], 1200 m, 29 April 1978, Essig & Young LAE 74072 (BH, LAE!, USF).
habitat. Sun-exposed limestone ridges in montane forest; 300 – 1200 m elevation.
vernacular names and uses. None known.
conservation status. Data deficient (DD). More data are needed about the distribution and abundance of this species.
notes. Hydriastele aprica is unique within the Nengella group in having mostly opposite and decussate triads, and the numerous irregularly arranged, cuneate and drooping leaflets add to the distinctiveness of this species. H. aprica is unlikely to be confused with other species in the group. The habitat preference is highly unusual, as noted by Young (1985).
2. Hydriastele divaricata Heatubun, Petoe & W. J. Baker sp. nov. Type: Indonesia, Papua Province, Mimika Regency (Previously Fakfak Regency), Timika, PT. Freeport Indonesia Area of Work, Mile 39, 23 Feb. 1998, Baker et al. 876 (holotype K!, isotypes BH!, BO!, L!, MAN!).
http://www.ipni.org/urn:lsid:ipni.org:names:77175849-1
Clustering, very slender palm with c. 5 stems per clump, to 4.5 m tall. Stem c. 9 – 17 mm in diam.; internodes 6 – 15 cm long, smooth, green, with small purple to black lacerate-peltate scales. Leaves to c. 80 cm long including petiole; sheath c. 25 cm long, indumentum as upper internodes and additionally coated with larger white to black scurfy scales, crownshaft c. 50 cm long; petiole c. 27 – 30 cm long, indumentum as sheath; rachis c. 46 cm long, indumentum as sheath; leaflets 9 – 12 each side of rachis, sub-regularly arranged with some divaricate pairs, borne 2 – 7.5 cm apart, narrowly linear, praemorse apically, slightly discolorous, papery; basal leaflets c. 30 – 33 × 1 cm, single-fold; middle leaflets 37 – 38 × 1 – 1.5 cm, single-fold; terminal leaflets 19 – 20 × 0.5 – 1.5 cm, comprising c. 2 folds. Inflorescences 10 – 12 cm long including 2 – 2.5 cm peduncle, pendent, spicate or branched to 1 order; prophyll not seen; rachillae 9 – 10 × 0.2 – 0.3 cm, 1 – 2 per inflorescence; triads spirally arranged with alternating verticils of 3. Staminate flowers not seen. Pistillate flowers 4 – 4.5 × 3 – 3.3 mm at fruiting stage; sepals c. 1.5 – 2 × 3 mm; petals 3 – 3.5 × 3 – 3.3 mm with conspicuous triangular tips; ovary c. 4.5 × 3 mm, ovoid; staminodes not seen. Fruits c. 12 × 3 mm, cylindrical, pale green to pinkish. Seeds c. 8 × 2 mm, elongate-cylindrical; endosperm shallowly ruminate (Fig. 2).

Hydriastele divaricata. A leaf diagram; B leaf sheath with portion of petiole; C leaf apex; D mid-leaf portion; E attached inflorescence with pistillate flowers; F attached infructescence; G portion of rachilla with developing fruit; H fruit whole, in longitudinal section, and in transverse section. Scale bar: A = 30 cm; B – D = 6 cm; E – F = 3 cm; G – H = 7 cm. All from Baker et al. 876. drawn by lucy t. smith.
recognition. Distinguished by its sub-regularly arranged, narrowly linear leaflets which are divaricate in the basal and middle section of the leaf.
distribution. Known only from the type locality near to Timika in Papua Province (Indonesia) on the lower slopes of the Mt Jaya region (Dransfield et al. 2000; Map 2).

Distribution map of Hydriastele divaricata.
specimens cited. indonesia. Papua Province: Mimika Regency (Previously Fakfak Regency), Timika, PT. Freeport Indonesia Area of Work, Mile 39, [4°20'S 136°58'E], 100 m, 23 Feb. 1998, Baker et al. 876 (BH!, BO!, K!, L!, MAN!).
habitat. The transition zone between lowland alluvial rain forest and heath forest; 100 m elevation.
vernacular names and uses. None recorded.
conservation status. Data deficient (DD). More data are needed about the distribution and abundance of this species.
notes. Hydriastele divaricata is readily distinguished by its leaves with widely spreading, narrowly linear leaflets (hence the choice of species epithet). The limited available material displays inflorescences consisting of just one (spicate) or two rachillae, prompting comparison with H. flabellata and H. montana, but these two species have homogeneous endosperm and spirally arranged triads with more or less well-defined verticils, whereas H. divaricata has ruminate endosperm and triads spirally arranged with distinct alternating verticils of three, like H. pinangoides. Vegetatively, H. divaricata is highly distinct from H. pinangoides, which typically bears irregularly arranged, cuneate leaflets. Rare forms of H. pinangoides display regularly arranged, narrowly cuneate leaflets, but these are never narrowly linear and divaricate.
3. Hydriastele flabellata (Becc.) W. J. Baker & Loo (2004: 64). Nengella flabellata Becc. (Beccari 1877: 34). Gronophyllum flabellatum (Becc.) Essig & B. E. Young (1985: 134). Type: Indonesia, Papua Barat, Sorong, Ramoi, Beccari 427 (holotype FI!).
Nengella calophylla var. montana Becc. (Beccari 1914: 27). Nengella pleurocarpa Burret (1936: 314). Gronophyllum pleurocarpum (Burret) Essig & B. E. Young (1985: 136). Hydriastele pleurocarpa (Burret) W. J. Baker & Loo (2004: 67). Type: Papua New Guinea, Madang, Schlechter 16251 (holotype B†, isotype FI!). synon. nov.
Nengella calophylla var. rhopalocarpa Becc. (Beccari 1914: 28). Nengella rhopalocarpa (Becc.) Burret (1936: 314). Gronophyllum rhopalocarpum (Becc.) Essig & B. E. Young (1985: 136). Hydriastele rhopalocarpa (Becc.) W. J. Baker & Loo (2004: 67). Type: Papua New Guinea, Morobe, Waria R., Schlechter 17466 (holotype B†, isotypes FI!, K!). synon. nov.
Nengella gracilis Burret (1939: 206). Gronophyllum gracile (Burret) Essig & B. E. Young (1985: 134). Hydriastele gracilis (Burret) W. J. Baker & Loo (2004: 65). Type: Papua New Guinea, Western, Palmer R., Brass 7083 (holotype A!). synon. nov.
Gronophyllum cariosum Dowe & M. D. Ferrero (2000a: 161). Hydriastele cariosa (Dowe & M. D. Ferrero) W. J. Baker & Loo (2004: 63). Type: Papua New Guinea, Sandaun, Bewani Mts, Dowe et al. 514 (holotype BRI). synon. nov.
Clustering, very slender palm forming clumps of up to 6 stems, to 4 m tall, bearing 4 – 8 leaves per crown. Stem 1 – 2 cm in diam., shiny green and becoming dark and dull brown; internodes 3 – 15 cm long. Leaves 35 – 90 cm long including petiole; sheath 8 – 23 cm long, green with small dark scales densest towards the apex, crownshaft 14.5 – 40 × 1.5 – 1.8 cm; petiole 10 – 30 cm, indumentum as sheath, sometimes with additional scurfy scales; rachis 5 – 30 cm long, indumentum as petiole; lamina 20 – 65 cm long, entire bifid, bijugate or more finely dissected with up to 6 leaflets each side of rachis, pale to mid green sometimes with a metallic quality, slightly discolorous, ± papery, ramenta not seen; leaflets (when present) regularly or irregularly arranged, single- or multi-fold, borne 2 – 13 cm apart, c. 9 – 24 × 0.8 – 15 cm, cuneate and praemorse apically. Inflorescences 8 – 15 cm long including 1 – 3 cm peduncle, spicate or branched to 1 order, pendent or erect; prophyll 8.5 – 15 × 1.5 – 3 cm, cream to light green; rachillae 7 – 12 cm long, 1 – 2 per inflorescence (up to 3 in one cultivated specimen), somewhat stiff, green to yellow; rachilla bracts often rather conspicuous (at least when dry) and cucullate; triads spirally arranged with ± well-defined verticils. Staminate flowers 8 – 10 × 2 – 3 mm in bud, cream-coloured to violet; calyx 1 – 2 × 2 – 3 mm consisting of three basally connate sepals; petals, 7 – 9 × 2 – 3 mm, lanceolate, valvate; stamens 6; anthers c. 3 – 6 × 1 mm: filaments c. 1 × 0.1 mm, variously epipetalous; pistillodes 1 – 3 lobes. Pistillate flowers 3 – 5 × 2 – 3.5 mm in bud, cream to light green or lavender; sepals 0.5 – 2 × up to 3 mm; petals 3 – 4 × 2 – 3 mm, with conspicuous triangular tips; ovary 1 – 1.5 × 1 mm; staminodes not seen. Fruits 12 – 15 × 5 – 8 mm, ellipsoid to fusiform, red, smooth or striate, glossy. Seeds 10 – 12 × 4 – 7 mm, ellipsoid; endosperm homogeneous (Fig. 3).

Hydriastele flabellata. A habit; B attached leaf; C – F leaf diagrams; G prophyll; H inflorescence; J portion of rachilla with triads; K staminate flower in longitudinal section; L pistillate flower in longitudinal section; M fruit whole, in longitudinal section, and in transverse section. Scale bar: A = 20 cm; B = 6 cm; C – F = 18 cm; G – H = 2 cm; J = 5 mm; K = 4 mm; L = 3 mm; M = 7 mm. A – C, G – L from Baker et al. 879; D from Schlechter 17466; E – F from Baker 643; M from Heatubun 406. drawn by lucy t. smith.
distribution. Scattered across New Guinea with collections known from parts of Papua Barat Province (Indonesia), the area around Timika in Papua Province (Indonesia), and from several major areas in Papua New Guinea south and north of the central New Guinea highlands (Map 3).

Distribution map of Hydriastele flabellata.
specimens examined. indonesia. Papua Barat Province: Sorong, Ramoi, [0°57'S 131°20'E], 1872, Beccari 427 (FI!); Sorong Selatan Regency, Sayal, Maampow Forest, [1°28'S 131°53'E], 10 m, 21 Feb. 2013, Heatubun 406 (K (spirit)!, MAN!); same locality as preceding, [1°28'S 131°53'E], 10 m, 21 Feb. 2013, Heatubun 426 (K (spirit)!, MAN!); Kaimana Regency, Teluk Etna, Waribun, Km-27 road to Kaltim Hutama, 100 m, 31 Jan. 2001, Heatubun 329 (K!, MAN!); Surroundings of Ayawasi, [1°5'S 132°17'E], 450 m, 16 Jan. 1996, Polak 1000 (BO, L!); Papua Province: Maribu village, Cyclops Mts Nature Reserve, [2°29'S 140°23'E], 850 m, 21 Sept. 1998, Maturbongs 576 (BO, K!, MAN,); Mimika Regency, Timika, Kali Kopi, [4°26'S 136°57'E], 95 m, 24 Feb. 1998, Baker et al. 879 (BO!, K!, MAN!); Kuala Kencana, [4°25'S 136°51'E], 50 m, 19 Feb. 1998, Witono 23 (BH, BO!, K!, MAN!); Jayapura, Cyclops Mts, path to Ormu, [2°26'S 140°37'E], 700 m, 18 Aug. 1998, Heatubun 285 (BO, FTG, K!, L, MAN). papua new guinea. Morobe Province: Morobe Sub-province, Mayama village, [7°35'S 147°10'E], 100 m, 17 June 1981, Katik LAE 74935 (BRI, L!, LAE, NSW, USF); Kamiali Wildlife Management Area, ridge to Blue Mt, near starting point of the Nembebah PABITRA plot, [7°17'S 147°5'E], 650 m, 28 Feb. 2005, Takeuchi 18982 (K!, LAE); Waria R., [7°58'S 147°28'E], 13 March 1908, Schlechter 17466 (B†, FI!, K!); Madang Province: 12 July 1907, Schlechter 16251 (B†, FI!); Sandaun Province: Roundhouse village, [3°2S 141°7'E], 200 – 250 m, 27 Nov. 1996, Barfod 413 (AAU!, LAE); Bewani, [3°1'S 141°8'E], 0 m, 19 March 2000, Barfod 499 (AAU!, K!, LAE); Lumi Sub-distr., Karataiem, off road between Lumi Govt. Station and Karataiem Catholic Mission, [3°27'S 142°1'E], Jan. 1974, Frodin s.n. (K!, UPNG); Telefomin Sub-distr., Prospect Creek near Frieda R., [4°42'S 141°48'E], 500 m, 23 June 1969, Henty NGF 42555 (BH, LAE!); Amanab Sub-distr., Imonda, along road to Bapi R., [3°20'S 141°10'E], 300 m, 25 Nov. 1971, Essig LAE 55094 (BH, CANB!, LAE!); Southern Highlands Province: Kutubu patrol area, Waro, 500 – 600 m, 3 Aug. 1991, Takeuchi 7284 (A!, LAE); same locality as preceding, 500 – 600 m, 3 Aug. 1991, Takeuchi 7312 (A!, LAE); Kantobo, [6°44'S 143°35'E] 470 m, 9 Feb. 1996, Baker et al. 643 (K!, LAE!); Mt Bosavi, Wasaso, Bona village, [6°26'S 142°47'E], 700 m, 2 Feb. 1996, Baker et al. 611 (BH, FTG, K!, LAE); Mt Bosavi, northern side, [6°26'S 142°50'E], 900 m, 26 Oct. 1973, Jacobs 9470 (L!, LAE); Western Province: Palmer R., 2 min. below junction Black R., [5°47'S 141°42'E], 100 m, June 1936, Brass 7083 (A!); Same locality as preceding, [5°47'S 141°42'E], 100 m, June 1936, Brass 7368 (A!). cultivated. united states. hawaiian islands: Hawai’i, Cultivated Floribunda Palms and Exotics, 14 Nov. 2017, Baker 1448 (K!).
habitat. The understorey of primary rainforest on slopes, ridges and riverbanks, on a diversity of soils (e.g. waterlogged alluvium, limestone karst and volcanic soil). This species should mainly be considered a lowland rainforest species as its habitat barely stretches into the premontane vegetation zone; 0 – 700 m elevation.
vernacular names and uses. Two local names are recorded for this species; mplemponik (Sayal), filiawoi yamu (Bewani).
The leaves are used as food wrapping in the Sayal area of Sorong Selatan in Indonesia (Heatubun 2005) and in the Bewani Mts of Sandaun Province in Papua New Guinea (Dowe & Ferrero 2000a). The stems are used for making arrow shafts and fish spear handles in Southern Highlands Province (Papua New Guinea).
conservation status. Least Concern (LC). The EOO (c. 404,000 km2) of this species indicates non-threatened status and the low AOO (44 km2) is thought to be an underestimate resulting from under-collecting. Even the relatively large EOO is likely to be a conservative figure.
notes. This widespread and variable species is distinguished by its variation in leaf lamina morphology ranging from entire bifid to pinnately compound with up to 6 cuneate leaflets per side. The inflorescence is spicate or occasionally with 2 rachillae (up to three rachillae has been confirmed from one cultivated specimen; Baker 1448), and it has spirally arranged triads with more or less well-defined verticils of 3, that are often subtended by conspicuously cucullate rachilla bracts. Hydriastele flabellata can be confounded with H. pinangoides because the foliage can appear similar, however the leaves of H. pinangoides are never entire bifid and usually include more leaflets, and its somewhat longer inflorescences bear 2 – 5 rachillae with spirally arranged triads in distinct alternating verticils of 3, subtended by inconspicuous rachilla bracts. In addition, the endosperm of H. flabellata is homogeneous, whereas H. pinangoides is ruminate. The only species within the Nengella group with a similar inflorescence structure to H. flabellata is H. montana but this species differs in having 9 – 11 narrowly linear leaflets per side and by forming gregarious associations.
There is a considerable span of variation in leaf morphology across specimens of Hydriastele flabellata with some individuals displaying an entire bifid leaf lamina (e.g. Barfod 413) and others a distinctly pinnate blade with up to 6 leaflets per side (e.g. Schlechter 17466; Fig. 3D). Between these outer extremes we observed intermediate forms and substantial within-specimen variation in the form of both near-entire and bijugate leaves (Baker et al. 643; Fig. 3E, F) and leaves with 3 – 5 leaflets per side (Baker et al. 879; Fig. 3C) present within single individuals. When we combined the continuum in leaf lamina morphology with the general uniformity and presence of continua observed across other characters studied, e.g. habit, leaf texture, inflorescence morphology, and seed endosperm condition, it became evident that H. rhopalocarpa, H. cariosa, H. pleurocarpa and H. gracilis can no longer be maintained and we therefore consequently place these names in synonymy under H. flabellata.
We make note here of a deviating specimen, Heatubun 285 from the Cyclops Mountains (Heatubun 2000). It resembles Hydriastele flabellata except that it has metallic leaves, inflorescences with up to 3 rachillae (known from one cultivated specimen; Baker 1448), and somewhat curved fruits when dry. However, there is insufficient material available currently to determine whether it should be recognised as a new species or not.
4. Hydriastele montana (Becc.) W. J. Baker & Loo (2004: 66). Nengella montana Becc. (Beccari 1877: 33). Gronophyllum montanum (Becc.) Essig & B. E. Young (1985: 134). Type: Indonesia, Papua, Arfak Mts, Gunon Morait, Beccari s.n. (holotype FI!, isotype K!).
Kentia beccarii F. Muell. (Mueller 1880: 163). Type: Indonesia, Papua, Arfak Mts, Beccari s.n. nom. superfl. & nom. illeg.
Gregarious palm to 1.5 m high, very slender, bearing 4 – 5 leaves per crown, forming new stems through vigorous rhizomatous growth. Stem 5 – 8 mm in diam.; internodes 3 – 8 cm long, smooth, covered with small dark lacerate-peltate scales. Leaves c. 25 – 50 cm long including petiole; sheath 10 – 15 cm long, indumentum as upper internodes if more dense; petiole 3 – 8 cm long, indumentum as sheath; rachis 19 – 38 cm long, indumentum as sheath; leaflets 9 – 11 each side of rachis, regularly arranged, alternate to sub-opposite, borne 1 – 4.5 cm apart, narrow linear, ± concolorous, ramenta not seen, papery; basal leaflets 18 – 25 × 0.8 – 1.3 cm, single- or bi-fold, pointed apically; middle leaflets c. 21 – 27 × 0.8 cm, single-fold, pointed and truncately praemorse apically; terminal leaflets 14 – 21 × 0.6 – 1.7 cm, single- or bi-fold, praemorse apically. Inflorescences 7 – 9 cm long including 1 – 1.5 cm peduncle, spicate, pendent; prophyll c. 10 × 1 cm; spike 6 – 7.5 × 0.3 – 0.4 cm, yellowish to green; rachilla bracts ± conspicuous; triads spirally arranged with ± well-defined verticils. Staminate flowers 9 – 10 × 1.5 – 2.5 mm in bud; calyx 1.5 – 2.5 × 1.5 – 2.5 mm consisting of three basally connate sepals; petals 9 – 9.5 × 1.5 – 2 mm, lanceolate, valvate; stamens 6; filaments c. 0.5 × 0.3 mm; anthers c. 3.5 – 4 × 0.5 mm; pistillodes 2 – 3, c. 0.5 × 0.3 mm, column shaped. Pistillate flowers 4 – 4.5 × 3 – 3.5 mm in bud; sepals 1.5 – 2 × 2.5 – 3 mm, rounded; petals 3.5 – 4 × 2.5 – 3 mm, rounded and imbricate at the base, with long valvate tips; ovary c. 2.5 × 2 mm, subglobose; staminodes c. 3, tooth-like, rudimentary. Fruits c. 12 – 19 × 4 mm (when dry), fusiform, red. Seeds c. 12 – 14 × 3 mm (when dry), elongate-turbinate; endosperm homogenous (Fig. 4).

Hydriastele montana. A stem basal portion with running rhizomes and roots; B leaf diagram; C apical and mid-leaf portion; D portion of stem with attached inflorescences; E portion of rachilla with pistillate flowers; F pistillate flower whole and in longitudinal section. Scale bar: A, C = 4 cm; B = 18 cm; D = 2 cm; E = 7 mm; F = 4 mm. All from Baker et al. 1380. drawn by lucy t. smith.
distribution. Known from two localities in the Tamrau Mountains on the Bird’s Head Peninsula (Map 4).

Distribution map of Hydriastele montana.
specimens cited. indonesia. Papua Barat Province: Kabupaten Tambrauw, Gunon [Gunung] Morait, [0°45'S 132°30'E], 1300 – 1500 m, July 1875, Beccari s.n. (FI!, K!); Kabupaten Tambrauw, Bamusbama distr., road to Fef, [0°46'S 132°16'E], 950 m, 28 Jan. 2013, Baker et al. 1380 (AAU!, BO!, K!, L!, MAN!); Locality info lacking, Iwanggin 132 (MAN, K!).
habitat. The understorey of premontane forest, on ridges and slopes; 950 – 1300 (– 1500) m elevation.
vernacular names and uses. None known.
conservation status. Data deficient (DD). More data are needed about the distribution and abundance of this species.
notes. Hydriastele montana is distinguished by its gregarious habit meaning that it occurs in groups and is locally dominant. This is distinct from the caespitose habit which is not uncommonly observed amongst understorey members of Hydriastele. It is arguably the production of multiple running rhizomes (Fig. 4A) that results in the colony formation. Another distinctive feature of this species is the presence of a spicate inflorescence, a character state that is only shared with H. flabellata within the Nengella group. However, H. flabellata has a bifid leaf lamina or the blade is pinnately compound and consisting of up to 6 regularly or irregularly arranged, cuneate leaflets on each side of the rachis, whereas H. montana has 9 – 11 narrowly linear, regularly arranged leaflets per side. Only H. divaricata has similar leaflets, but they are divaricate and it also has triads that are spirally arranged with distinct verticils of 3 and seeds with ruminate endosperm. Hydriastele montana has spirally arranged triads with more or less well-defined verticils, and homogeneous endosperm.
The name Kentia beccarii F. Muell has been highlighted as a superfluous replacement name for Hydriastele montana and is therefore illegitimate (Dowe 2017).
5. Hydriastele pinangoides (Becc.) W. J. Baker & Loo (2004: 66). Nenga pinangoides Becc. (Beccari 1877: 28). Leptophoenix pinangoides (Becc.) Becc. (Beccari 1885: 82). Nengella pinangoides (Becc.) Burret (1936: 315). Gronophyllum pinangoides (Becc.) Essig & B. E. Young (1985: 135). Type: Indonesia, Papua Barat, Ramoi, Beccari 430 (holotype FI!, isotype K!).
Nenga affinis Becc. (Beccari 1877: 29). Leptophoenix affinis (Becc.) Becc. (Beccari 1885: 82). Nengella affinis (Becc.) Burret (1936: 316). Gronophyllum affine (Becc.) Essig & B. E. Young (1985: 136). Hydriastele affinis (Becc.) W. J. Baker & Loo (2004: 62). Type: Indonesia, Papua, Kapaor, Beccari s.n. (holotype FI!, isotype K!). synon. nov.
Nenga calophylla K. Schum. & Lauterb. (Schumann & Lauterbach 1900: 208). Nengella calophylla (K. Schum. & Lauterb.) Becc. (Beccari 1914: 27). Type: Papua New Guinea, Morobe, Sattelberg, Lauterbach 564 (holotype B†, type photo FI!).
Leptophoenix minor Becc. (Beccari 1905: 298). Nengella minor (Becc.) Burret (1936: 315). Type: Papua New Guinea, San Giuseppe R., Loria s.n. (holotype FI!).
Gronophyllum densiflorum Ridl. (Ridley 1916: 232). Leptophoenix densiflora (Ridl.) Burret (1936: 205). Nengella densiflora (Ridl.) Burret (1936: 316). Type: Indonesia, Papua, Mt Carstenz, Kloss s.n. (holotype BM, isotype K!).
Leptophoenix incompta Becc. (Beccari 1923: 452). Nengella incompta (Becc.) Burret (1936: 316). Type: Papua New Guinea, East Sepik, Ettapenberg, Ledermann 9017 (holotype B†).
Leptophoenix mayrii Burret (1933: 709). Nengella mayrii (Burret) Burret (1936: 314). Gronophyllum cyclopense Essig & B. E. Young (1985: 136). Hydriastele cyclopensis (Essig & B. E. Young) W. J. Baker & Loo (2004: 64). Type: Indonesia, Papua, Arfak Mts, Mayr 24 (holotype B†, isotype BO — see notes section). synon. nov.
Leptophoenix micrantha Burret (1933: 710). Nengella micrantha (Burret) Burret (1936: 314). Gronophyllum micranthum (Burret) Essig & B. E. Young (1985: 136). Hydriastele micrantha (Burret) W. J. Baker & Loo (2004: 65). Type: Indonesia, Papua, Wandammen Mts, Mayr 253 (holotype B†, isotype BO!). synon. nov.
Leptophoenix pterophylla Becc. (Beccari 1934: 131). Nengella pterophylla (Becc.) Burret (1936: 316). Type: Cultivated in Bogor Botanic Garden ex New Guinea, X D 114 (holotype FI!).
Leptophoenix yulensis Becc. (Beccari 1934: 130). Nengella yulensis (Becc.) Burret (1936: 316). Type: Papua New Guinea, Central, von Mueller s.n. (holotype MEL!, isotype FI!).
Leptophoenix brassii Burret (1935: 339). Nengella brassii (Burret) Burret (1936: 316). Gronophyllum leonardii Essig & B. E. Young (1985: 134). Type: Papua New Guinea, Western, Kubuna, Brass 5631 (holotype A, isotypes BRI!, BO!, NY!).
Leptophoenix macrocarpa Burret (1935: 340). Nengella macrocarpa (Burret) Burret (1936: 316). Type: Papua New Guinea, Central, Mafulu, Brass 5299 (holotype B†, isotypes BRI!, NY!).
Leptophoenix microcarpa Burret (1935: 342). Nengella microcarpa (Burret) Burret (1936: 316). Type: Papua New Guinea, Central, Dieni, Brass 3998 (holotype B†, isotypes A!, NY, BRI, BO!).
Nengella rhomboidea Burret (1939: 207). Type: Papua New Guinea, Western, Palmer R., Brass 7201 (holotype A!, isotype BRI).
Solitary or clustering, very slender to slender palm to 7 (– 10) m tall, often shorter, bearing 5 – 10 leaves per crown. Stem (0.8 –) 1.5 – 7.5 cm in diam.; internodes 4 – 16 cm long, smooth, grey to greenish, ± speckled and lepidote. Leaves 50 – 157 cm long including petiole, reddish to bronze when newly emerged; sheath 15 – 52 cm long, green with small brown to purple lacerate-peltate scales covered by larger scurfy scales, crownshaft 23 – 60 (– 112) × 2 – 6 cm; petiole 15 – 54 cm long, indumentum as sheath; rachis 30 – 90 cm long, indumentum as sheath; leaflets 5 – 10 (– 13) each side of rachis, very variable in size and shape, irregularly arranged usually in 2 – 4 widely spaced groups sometimes with portions of the lamina regularly pinnate, very rarely the entire blade regularly pinnate, broadly (rarely narrowly) cuneate, praemorse apically, mid to dark green adaxially and somewhat paler abaxially, ± discolorous, leathery; basal leaflets 16 – 36 × 4 – 11 cm, comprising (1 –) 3 – 6 folds; middle leaflets 12 – 50 × 2 – 20 cm, single- or multi-fold; terminal leaflets 7 – 36 × 3 – 24 cm, multi-fold. Inflorescences (13 –) 18 – 30 cm long including 1.5 – 3 cm peduncle, branched to 1 order, arching; prophyll 12 – 31 × 2 – 5 cm, cream-green; rachillae 8 – 25 × 0.2 – 0.5 cm, 2 – 5 (– 6) per inflorescence, with various shades of pink; triads spirally arranged with alternating verticils of 3. Staminate flowers 6 – 8 × 1 – 3 mm in bud (when dry), vinaceous; sepals 1 – 2 × 1.1 – 2 mm, basally connate, rounded or sometimes acuminate apically; petals 5 – 7 × 1.5 – 2 mm, lanceolate and valvate; stamens 6; filaments c. 1 – 1.3 × 0.2 – 0.3 mm; anthers 3.5 – 4 × 0.5 mm; pistillodes not seen. Pistillate flowers c. 4 × 3.5 mm at fruiting stage, crimson; sepals c. 2 × 3 mm, rounded, imbricate; petals c. 3.5 × 3.5 mm, basally imbricate, with conspicuous valvate tips; staminodes several, small, tooth-like. Fruits 10 – 16 × 4 – 8 mm, ellipsoid or cylindrical to fusiform, smooth and shiny, pink, red, purple or blackish. Seeds 6 – 10 × 2 – 4 mm broadly to narrowly ellipsoid; endosperm ruminate (Fig. 5).

Hydriastele pinangoides. A leaf diagram; B leaf apex; C mid-leaf portion; D leaf base; E inflorescence with attached peduncular bract; F infructescence; G portion of rachilla with triads; H staminate flower whole and in longitudinal section; J fruit whole, in longitudinal section, and in transverse section. Scale bar: A = 24 cm; B – D = 8 cm; E – F = 4 cm; G – H = 5 mm; J = 7 mm. A – H from Pullen 5767; J from Baker et al. 1057. drawn by lucy t. smith.
distribution. Occurs throughout New Guinea including the islands of Misool, Waigeo, and Kobroor in the Aru Archipelago (Map 5).

Distribution map of Hydriastele pinangoides.
specimens cited. indonesia. Maluku Province: Aru Islands, Pulau Kobroor, [6°15'S 134°17'E], 0 m, 6 Nov. 1994, van Balgooy 6855 (K!, L!); Papua Barat Province: Ramoi, [0°57'S 131°20'E], 1872, Beccari PP 430 (FI!, K!); Kabupaten Pegunungan Arfak, Arfak Mts, Minyambou, Mt Nerimbau, [1°8'S 133°51'E], 1600 m, 19 May 1962, Koster BW 13879 (L!, LAE!, MAN); Kabupaten Teluk Bintuni, Concession of PT. Manokwari Mandiri Lestari, Blok RKT 2010, Jalur 22 Petak 18Y, 250 m, 18 July 2010, Heatubun CH 1040 (K!, MAN); Kabupaten Kaimana, Teluk Arguni, Distrik Arguni Bawah, Kampung Jawera, Hutan Manggai, [3°4'S 133°40'E], 0 m, 3 Oct. 2010, Heatubun CH 1074 (K!, MAN); Kabupaten Teluk Wondama, Distrik Wombu (Wasior Barat), Wosimi, Kambi Forest, [3°0'S 134°32'E], 50 m, 18 Aug. 2006, Heatubun CH 777 (BO, K!, MAN); Kabupaten Manokwari, Warmare; Valley of R. Prafi, new road to Manyambo, [0°47'S 133°58'E], 350 m, 25 Aug. 1995, Dransfield JD 7605 (BO, K!); Manokwari, Kebar, Kebar Valley, forest immediately due N of Andjai in valley, [0°55'S 133°3'E], 550 m, 30 April 1995, Davis 685 (K!); Manokwari Distr., Warmare, Prafi R. valley, new road to Manyanbo, [0°47'S 133°58'E], 350 m, 25 Aug. 1995, Zona 689 (FTG, K!); Manokwari Distr., Nuni, Sungei Asai, between Mt Manggombo and Mt Marwadibau, [0°45'S 133°56'E], 0 m, 15 Aug. 1995, Zona 671 (FTG, K!); Vogelkop Peninsula, Segior, NW side of Lake Ajamaru, [1°14'S 132°12'E], 250 m, 10 March 1962, Vink BW 15285 (A, K!, L!); Sorong, Tourism Park, Klasaman, KM14, [0°55'S 131°22'E], 50 m, 25 July 1996, Wally 490 (K!, MAN); Sorong, Waifoi village, Waigeo Island, Raja Ampat Islands, [0°14'S 130°50'E], 200 m, 25 June 1997, Wally 702 (K!, MAN); Sorong, Waifoi Village, Waigeo Island, Raja Ampat Islands, [0°14'S 130°50'E], 400 m, 25 June 1997, Maturbongs RAM 506 (K!, MAN); Sorong, Mamiai, Waigeo Island, [0°20'S 131°9'E], 50 m, 25 June 1997, Heatubun 87 (K!, MAN); Manokwari, Cultivated, in the front of a mosque within the university area, [0°51'S 134°4'E], 100 m, 13 Aug. 1995, Keim AK 2 (K!); Wandammen Peninsula, Wasior Distr., Manokwari Regency, Kowi, near Wondiwoi village (formerly Kobiari Village), c. 9 km S of Wasior, [2°48'S 134°32'E], 900 m, 24 Feb. 2000, Baker et al. 1057 (BO, K!, MAN); Siwi village, Ransiki Sub-distr., Manokwari, [1°30'S 134°2'E], 28 Jan. 1999, Heatubun CH 290 (BO, FTG, K!, MAN); Raja Ampat Regency, Misool Island, Motlol, [1°53'S 129°44'E], 0 m, 22 Jan. 2002, Heatubun CH 358 (K!, MAN); Sorong Distr., Cape Seget, Seget Camp, [1°20'S 131°0'E], 0 m, 26 Jan. 2002, Heatubun CH 367 (K!, MAN); Sorong Distr., around Wavari Logging Camp, 10 km SW of Limalas village, 0 m, 21 Jan. 2002, Wanggai 04 (BO, K!, LAE, MAN); Sorong Distr., N Misool Island, about 20 km W of Waigama village, near Motlol Camp, [1°53'S 129°44'E], 0 m, 22 Jan. 2002, Maturbongs 699 (BO, K!, LAE, MAN); Sorong Distr., Warsamson, Warsamson R., [0°49'S 131°23'E], 0 m, 28 Jan. 2002, Heatubun 373 (K!, MAN); Tambrouw Regency, Fef Distr.; Forest above Fef, [0°48'S 132°27'E], 550 m, 26 Jan. 2013, Baker 1377 (BO, K!, L, MAN); Papua Province: Wandamen Mts, 1400 m, 8 July 1928, Mayr 253 (BO!); Kapaor, April 1872, Beccari s.n. (FI!, K!); Fak-Fak, Timika, Road to Kali Kopi from mile 38. Loc 35, [4°25'S 136°56'E], 100 m, 6 Feb. 1998, Baker et al. WJB 819 (BH, BO, K!, MAN); Fak-Fak, Timika, between Timika and port, peat lens (plot 22) Loc 20, [4°47'S 136°51'E], 0 m, 16 Feb. 1998, Witono JK 17 (BH, BO, K!, L, MAN); Jayawijaya, Snow Mts region, E of the Baliem Valley, Kab. Jayawijaya, Kec. Kurima, vicinity of Panggema village, [4°10'S 139°20'E], 1350 m, Oct. 1992, Milliken 1423 (K!); Mt Carstensz, [4°5'S 137°11'E], 12 Jan. 1912, Kloss s.n. (K!); Jayapura, North Cyclops Mts, [2°30'S 140°32'E], 150 m, 30 Jan. 2001, Desianto BD 03 (AAU!, K!, MAN); Cyclops Mts, on grassy slope above Ifar, [2°33'S 140°31'E], 400 m, 31 Dec. 1954, Mackee 1868 (L!); Fak Fak, Kaimana distr., Kroy village, [3°38'S 135°42'E], 100 m, 27 Nov. 2000, Mehen SM 08 (AAU, CANB, K!, LAE, MAN); 4 km SW of Bernhard Camp, Idenburg R., [3°29'S 139°6'E], 900 m, March 1939, Brass 13095 (A!, L!); Same locality as preceding, [3°29'S 139°6'E], 850 m, March 1939, Brass 13434 (A!); 6 km SW of Bernhard Camp, Idenburg R., [3°30’S 139°5’E], 1200 m, Feb. 1939, Brass 12872 (A!, L!); Same locality as preceding, [3°30'S 139°5'E], 1100 m, Feb. 1939, Brass 12953 (A!, L!); Same locality as preceding, [3°30'S 139°5'E], 1150 m, Feb. 1939, Brass 12998 (A!, L!); Same locality as preceding, [3°30'S 139°5'E], 1100 m, Feb. 1939, Brass 12952 (A!, L!); Same locality as preceding, [3°29'S 139°6'E], 1100 m, Feb. 1939, Brass 12997 (A!). papua new guinea. Central Province: Mafulu, [8°33'S 147°7'E], 1250 m, Sept. 1933, Brass 5299 (BRI!, NY!); Dieni, Ononge Road, [8°37'S 146°50'E], 500 m, April 1933, Brass 3998 (A!, BO!, BRI, NY); Veimauri logging site, [9°2'S 147°3'E], 5 Sept. 1984, Naoni UPNG 6146 (LAE!, US); Kubuna, [8°42'S 146°45'E], 100 m, Nov. 1933, Brass 5631 (A!, BO!, BRI, NY); Head of Goldie R. some 4 km from the National walking track entrance, [9°21'S 147°31'E], 400 m, Aug. 1981, Naoni UPNG EKN s.n. (LAE!); Kairuku Sub-distr., near Mipa Airtstrip, Maipa village, [8°20'S 146°33'E], 50 m, 18 Sept. 1962, Darbyshire 973 (CANB!); Sogeri Plateau, [9°25'S 147°26'E], 550 m, 1 Aug. 1962, Pullen 3460 (CANB!); Port Moresby Sub-distr., logging road between Kuriva & Veimauri Rs, [9°5'S 147°5'E], 50 m, 27 Feb. 1972, Essig LAE 55185 (LAE!); Port Moresby Sub-distr., Owers Corner, Kokoda Trail, [9°25'S 147°30'E], 1 March 1972, Essig LAE 55187 (BH, LAE!); Towards Mt Yule, 8 Dec. 1890, Mueller s.n. (FI!, MEL!); Chimbu Province: Crater Mt, Wildlife Management Area, near Haia village, Wara O., [6°43'S 145°0'E], 650 m, 3 March 1997, Takeuchi 11697 (A!, GH); East Sepik Province: Maprik Sub-distr., Prince Alexander Range, SE side of Mt Turu, near Yangoru Patrol Post, Wewak-Angoram Area, [3°37'S 143°22'E], 600 m, 19 Aug. 1959, Pullen 1504 (CANB!, LAE!); Ambunti Sub-distr., E ridge of Sumset (Mt Hunstein), [4°30'S 142°39'E], 1200 m, 17 Aug. 1966, Hoogland 11052 (CANB!, LAE!); Gulf Province: Purari Delta, 32.5 km E of Baimuru, [7°29'S 145°6'E], 0 m, 24 March 1974, Croft LAE 61091 (BH, L!, LAE!); Baimuru Sub-distr., Purari R., delta area 32.5 km E of Baimuru, [7°31'S 145°7'E], 0 m, 24 March 1974, Croft LAE 61085 (LAE!); Baimuru Sub-distr., Vailala R., 70 km at 110 degrees from Baimuru, 50 km at 318 degrees from Kerema, [7°38'S 145°28'E], 50 m, 4 April 1974, Croft LAE 61279 (BH, LAE!); Milne Bay Province: Raba Raba Sub-distr., Junction Ugat and Mayu Rs, near Mayu, [9°37'S 149°10'E], 350 m, 30 June 1972, Streimann NGF 28658 (LAE!); Same locality as preceding, [9°37'S 149°10'E], 350 m, 13 July 1972, Streimann NGF 28835 (BH, BRI, L!, LAE!); Maiyu R., E of Mt Suckling, [9°40'S 149°10'E], 450 m, 14 June 1972, Pullen 8296 (BH, CANB!, L!, LAE); N slopes of Mt Dayman, Maneau Range, [9°41'S 149°17'E], 700 m, 15 July 1953, Brass 23422 (A!); Raba Raba Sub-distr., Mayu camp site I, Mt Suckling, [9°37'S 149°10'E], 400 m, 9 June 1972, Leach NGF 33288 (BH, LAE!); Morobe Province: Sattelberg, Lauterbach 564 (B†, FI!); Sattelberg, [6°29'S 147°47'E], 17 Sept. 1935, Clemens 135 (L!); same locality as preceding, [6°29'S 147°47'E], 13 Jan. 1936, Clemens 1623 (L!); same locality as preceding, [6°29'S 147°47'E], June 1936, Clemens 298 (L!); Kwaimengu, Aseki patrol area, [7°21'S 146°12'E], 1150 m, 23 April 1966, Craven 1424 (CANB!, L!, LAE!); Kumbok Mt, N of Busu R., [6°33'S 146°59'E], 300 m, 26 Jan. 1993, Takeuchi 8691 (A!, L!, F); Same locality as preceding, [6°33'S 146°49'E], 400 m, 20 Jan. 1993, Takeuchi 8680 (A!); Between Gobadik and Gawam, [6°33'S 146°59'E], 350 m, 5 Feb. 1993, Takeuchi 8743 (A!); Lae Sub-distr., along Mo R., a few miles SW of Ana village, [7°49'S 147°32'E], 50 m, 29 Jan. 1972, Essig LAE 55167 (BH, LAE!); Lae Sub-Distr., along small stream, c. ½ mile NW of Ana Village, [7°48'S 147°33'E], 50 m, 27 Jan. 1972, Essig LAE 55155 (BH, LAE!); Menyamya Sub-distr., 2 miles SE of Aseki Patrol Post on Aseki-Koki Rd, [7°20'S 146°14'E], 1200 m, 9 Jan. 1972, Essig LAE 55140 (BH, LAE!); Huon Peninsula, Masba Creek area, c. 3 miles S of Pindiu, [6°27'S 147°31'E], 600 m, 22 May 1964, Hoogland 9030 (CANB!); Lae Distr., Lae Botanic Garden, [6°45'S 147°0'E], 0 m, 29 April 1974, Katik NGF 38000 (LAE!); Lae Subprovince, Lae Botanical Garden, [6°45'S 147°0'E], 0 m, 9 April 1978, Young LAE 74011 (BH, LAE!); Mumeng, Gurakor, along Bulolo Road, [6°45'S 147°0'E], 450 m, 15 Sept. 1971, Essig LAE 55005 (BH, LAE!); Oro Province: Managalase area, Near Pongani Falls SE of Sila, [9°5'S 148°23'E], 600 m, 21 Aug. 1964, Pullen 5767 (CANB!); Sandaun Province: Wutung Sub-province, Oenake Range, foothills of Mt Bougainville, [2°37'S 141°0'E], 550 m, 7 Sept. 1982, Kerenga LAE 56427 (L!, LAE); Telefomin, Delongkim Creek, Old village site on bank of Delongkim Creek near junction with Hak R., [5°8'S 141°35'E], 750 m, 16 Oct. 1993, Morren 3059 (K!); Telefomin Subprovince, Carpentaria Exploration Company, Frieda R. Camp, [4°40'S 139°55'E], 50 m, 26 April 1978, Essig LAE 74050 (BH, LAE!); Southern Highlands Province: Lake Kutubu, side of Mendi track, [6°9'S 143°39'E], 23 May 1956, Gray NGF 8135 (A, BRI!, LAE!); Mount Bosavi, Kolok, near Bona Village, WWF Integrated Conservation and Development Project Area, [6°26'S 142°47'E], 700 m, 2 Feb. 1996, Baker et al. WJB 616 (BH, K!, LAE); near Waro airstrip, 20 km SSW of Kutubu, [6°31'S 143°10'E], 500 m, 15 Oct. 1973, Jacobs s.n. (L!); Lake Kutubu, Wanunuku, near to Tugiri, WWF Integrated Conservation and Development Project Area, [6°21'S 143°13'E], 900 m, 12 Feb. 1996, Baker et al. 665 (BH, K!, LAE); Waro and Ubogo Villages, Kutubu distr., [6°32'S 143°12'E], 450 m, 14 Sept. 1993, Takeuchi 9242 (A, LAE!); Mt Bosavi, Northern side, [6°26'S 142°50'E], 1000 m, 23 Oct. 1973, Jacobs 9405 (L!, LAE!); Unknown Province: Mt Belford, 1100 m, Armit s.n. (MEL!); San Giuseppe, 10 Nov. 1892, Loria s.n. (FI!); Western Highlands Province: Kopiago Sub-distr., Batane, off Tari Rd, 9 miles from Kopiago, [5°22'S 142°33'E], 1350 m, 30 Oct. 1968, Womersley NGF 37233 (LAE!); Western Province: Palmer R., 2 miles below junction with Black R., [5°47'S 141°42'E], July 1936, Brass 7201 (A!, BRI); North Fly Distr., Junction of Harvey Creek and Ok Mani R., 10 km WNW of Tabubil, [5°14'S 141°8'E], 750 m, 14 Dec. 2000, Baker et al. 1137 (K!, LAE); Kiunga, Fly R. (Bulge), [7°2'S 140°2'E], 200 m, 28 March 1968, Millar NGF 35430 (A, BH, BO, BRI, CANB, K!, L!, LAE!, NSW); Nomad Subprovince, across R., 2 km from Nomad, [6°20'S 142°15'E], 18 April 1978, Essig LAE 74029 (BH, LAE!); Same locality as preceding, [6°20'S 142°15'E], 152 m, 18 April 1978, Essig LAE 74028 (BH, LAE!); Kiunga, 50 km NE of Ningerum, [7°22'S 141°28'E], 650 m, HYN 212 (LAE!). cultivated. indonesia: Bogor Botanic Garden ex New Guinea, X D 114 (FI!). united states. hawaiian islands: Kauai, 20 Aug. 2002, Chapin 80 (K!, PTBG).
habitat. The understorey or midstorey of lowland or premontane rainforest on well-drained to water-logged soil, sometimes on hill sides and ridge crests. This species grows in primary or secondary forests on a range of soil types including clay, limestone karst and volcanic soils; 0 – 1350 m elevation.
vernacular names and uses. A dozen local names are known for this species; tapolo (Pawaian), gilaia (Waskuk), kobu (Wagu), ugarreh (Daga), mara (Ambakanjah), kupal (Gal), tegradri (Irarutu), bim/kabim (Matbaat), yali (Biyal), seraach (Maibrat), sêméngbrè (Hattam). Names of unknown dialects from Papua New Guinea; manggam (Maprik), tooma (Western Province).
The stems are used for spears and spearheads (Sandaun and Chimbu Provinces, Papua New Guinea), for arrowheads (Papua Province, Indonesia) and for sewing thatch and as bow material (Papua Province, Indonesia). The foliage is utilised as roofing material and the palm as a whole, as an ornamental (Papua Barat Province, Indonesia). This palm is also used for magic (Papua Province, Indonesia).
conservation status. Least Concern (LC). This species has a large EOO (> 1,000,000 km2) and the relatively small AOO (300 km2) is thought to stem from the fact that Hydriastele pinangoides has still not been collected from many localities where it may very well occur.
notes. This variable species is distinguished from other members of the Nengella group mainly by its ruminate endosperm and spirally arranged triads with distinct alternating verticils of 3 in combination with broadly cuneate leaflets arranged in 2 – 4 distinct groups. Rarely, the leaflets can be ± regularly arranged and ± narrowly cuneate prompting comparison with Hydriastele divaricata and H. simbiakii, but H. divaricata has divaricate leaflets and inflorescences with 1 – 2 rachillae, whereas H. simbiakii has more numerous leaflets and flexible, often leaning, stems. Hydriastele flabellata can have similar looking foliage but this species has a spicate and relatively short inflorescence (more rarely with 2 rachillae or up to 3 rachillae known from one cultivated specimen) often with conspicuously cucullate rachilla bracts subtending spirally arranged triads with more or less well-defined verticils, and seeds with homogeneous endosperm.
We concur with the taxonomic changes for this species established by Essig & Young (1985), who formally synonymised multiple species thereby broadly circumscribing Hydriastele pinangoides. Here, we expand the delimitations of this variable species further by tentatively placing H. affinis, H. micrantha and H. cyclopensis in synonymy under H. pinangoides. We have opted for this solution because the morphology displayed by these species is consistent with H. pinangoides, as circumscribed here, although they are notable for their leaves which consist of more or less regularly arranged, narrowly cuneate leaflets. However, we have seen evidence of leaflet arrangement transitioning between forms collected from a single area (e.g. Brass 13095 [regularly arranged leaflets]; Brass 12952 and Brass 12998 [slightly irregularly arranged leaflets]), which supports the suggestion that leaflet arrangement can be plastic within this group. Two other collections (Essig LAE 55185; Koster BW 13879) further bridge the gap between the ± regularly arranged, narrowly cuneate leaflet form and the irregularly arranged, broadly cuneate leaflet form. Both specimens display variably cuneate leaflets with one group present in the middle of the rachis, but the grouping is less pronounced than that which is seen in a typical H. pinangoides specimen. On a nomenclatural point, we have inferred that the species epithet coined in the combination Gronophyllum cyclopense Essig & B. E. Young, a nomen novum to replace Nengella mayrii (Burret) Burret by Essig & Young (1985), appears to have been chosen in error, because the type of this name was collected in the Arfak Mountains, not the Cyclops Mountains. At the time, the name Gronophyllum mayrii (Burret) H. E. Moore already existed (based on a specimen that was collected in the Cyclops Mountains) and the recombination of Nengella mayrii in Gronophyllum required a nomen novum. In chosing the epithet cyclopense, Essig & Young appear to have confused the type localities of Gronophyllum mayrii and Nengella mayrii. Unfortunately, we have not been able to locate the type specimen of H. cyclopensis. We believe that it is extant at BO because it is recorded as such in the BO specimen database. However, the specimen could not be located by curators there. Its placement as a synonym of H. pinangoides is inferred from the species protologue (Burret 1933).
6. Hydriastele simbiakii Heatubun, Petoe & W. J. Baker sp. nov. Type: Indonesia, Papua Barat Province, Tambrauw Regency, Fef Distr., Sungai Sujak (Sungai Eyei), river bank near bridge, 23 Jan. 2013, Baker et al. 1365 (holotype K!, isotypes AAU!, BO!, L!, MAN!).
http://www.ipni.org/urn:lsid:ipni.org:names:77175855-1
Clustering, very slender rheophytic palm to 5 – 6 m tall, bearing c. 6 – 7 leaves per crown. Stem 1.5 – 2.5 cm in diam., shorter stems erect, longer stems leaning over river, clumps often including numerous stems; internodes 3.5 – 8 cm long, smooth, green turning brown. Leaves 64 – 104 cm long including petiole; sheath 26 – 36 cm long, clothed with small purple lacerate-peltate scales, crownshaft 34 – 44 × 3.5 – 4.5 cm; petiole 12 – 25 cm long, indumentum as sheath and additionally with patches of coarse grey to black scales; rachis c. 40 – 60 cm long, slightly arching, indumentum as petiole; leaflets 14 – 16 each side of rachis, regularly arranged, alternate to opposite, borne 0.5 – 3.5 cm apart, linear, obliquely praemorse apically, mid green adaxially and paler green abaxially, leathery; basal leaflets 15 – 20 × 0.8 – 1 cm, single-fold; middle leaflets 30 – 35 × 1 – 1.8 cm, single-fold; terminal leaflets 23 – 25 × 2 – 3 cm, comprising c. 5 folds. Inflorescences 13 – 17 cm long including 1.5 – 3 cm green to yellow peduncle, branched to 1 order; prophyll c. 17 × 3.5 cm, lepidote near apex; rachillae 7 – 16 × 0.2 – 0.3 cm, 3 – 4 per inflorescence, scarlet; triads spirally arranged with alternating verticils of 3. Staminate flowers 10 – 11 × 5 – 6 mm at anthesis, pink; calyx 1.5 – 2.6 × 1.7 – 2 mm, irregularly 3-lobed; petals 7.2 – 8 × 3.5 – 5 mm, lanceolate; stamens 6; filaments c. 0.8 – 1 × 1 mm; anthers 3.75 – 5 × 1.5 – 2 mm; pistillodes not seen. Pistillate flowers 4 – 5.5 × 2 – 3 mm after anthesis, sepals 2.5 – 3 × 2 – 3 mm, broadly imbricate, scarlet; petals 4 – 5.5 × 2 – 2.5 mm, broadly imbricate basally, conspicuously acute and valvate apically, white to pink; ovary c. 3 × 1.8 mm, ovoid; staminodes not seen. Fruits 8 – 12 × 4 – 5 mm, ellipsoid, red. Seeds 7 – 8 × 4 mm, ellipsoid; endosperm ruminate (Fig. 6).

Hydriastele simbiakii. A habit; B leaf diagram; C leaf apex; D leaf base; E inflorescence; F portion of rachilla with pistillate flowers; G portion of rachilla with fruits; H staminate flower whole and in longitudinal section; J pistillate flower whole and in longitudinal section; K fruit in longitudinal section. Scale bar A = 60 cm; B = 24 cm; C – D = 6 cm; E = 4 cm; F – H = 8 mm; J = 4 mm; K = 7 mm. All from Baker et al. 1365. drawn by lucy t. smith.
recognition. Distinguished from all other species in the Nengella group by forming clusters of pliable stems, the longer ones of which are leaning, by displaying 14 – 16 regularly arranged, linear leaflets per side, and by being restricted to growing on riverbanks within the flood-zone.
distribution. Known only from the type locality in the Tamrau Mountains in the central northern portion of the Bird’s Head Peninsula (Map 6).

Distribution map of Hydriastele simbiakii.
specimens cited. indonesia. Papua Barat Province: Tambrauw Regency, Fef Distr., Sungai Sujak (Sungai Eyei), river bank near bridge, [0°50'S 132°30'E], 400 m, 23 Jan. 2013, Baker et al. 1365 (AAU!, BO!, K!, L!, MAN!).
habitat. Steep riverbanks in the flood zone c. 1 – 2 m above fast flowing water, on rocky river margin over limestone; 400 m elevation.
vernacular names and uses. No local names are recorded for this species. The stems are used for making spears.
conservation status. Data deficient (DD). This species could be threatened as a new road has been constructed through the only known locality where it occurs, and there is a potentially increasing deforestation threat for the area in the future. Even so, more data are still needed about the distribution and abundance of this species before an assessment can be made.
notes. Hydriastele simbiakii is the only rheophytic species in the Nengella group, although a second rheophytic species in the genus, H. rheophytica, a member of the H. wendlandiana group (Dowe & Ferrero 2000b; Petoe et al. 2018), is known. Although closely related to H. pinangoides and other species with ruminate endosperm, it is immediately distinguished by its ecological preferences (scattered clumps of this species are restricted to a strictly riverine habitat) and by its leaves which have numerous regularly arranged, linear leaflets. Hydriastele pinangoides very rarely displays a regularly pinnate lamina.
The species is named for Victor I. Simbiak, botanist at Universitas Papua, who drew WJB’s attention to this palm in the field in the Tamrau Mountains.
7. Hydriastele splendida Heatubun, Petoe & W. J. Baker sp. nov. Type: Indonesia, Papua Province, Mimika Regency (Previously Fakfak Regency), Timika, PT. Freeport Indonesia Area of Work, Km 63 road to Tembagapura, 7 Feb. 1998, Baker et al. 820 (holotype K!, isotypes AAU!, BH!, BO!, L!, MAN!).
http://www.ipni.org/urn:lsid:ipni.org:names:77175863-1
Clustering or solitary, very slender palm to 4 m tall, bearing c. 4 – 5 leaves per crown. Stem 1.5 – 2 cm in diam.; internodes 6 – 15 cm long, smooth, green, with patches of small lacerate-peltate purple to black scales. Leaves c. 100 cm long including petiole, with conspicuous dense indumentum of scurfy, white, caducous scales (resembling vermiculite) throughout petiole and rachis, and extending on to adaxial and abaxial surface of lamina veins and throughout the sheath, scales blackening and sometimes lost with age; sheath c. 30 cm long, lepidote beneath the scurfy, white scales, crownshaft c. 50 cm long; petiole c. 20 cm long, indumentum as sheath; rachis c. 46 cm long, indumentum as sheath; lamina c. 80 × 40 cm, entire bifid with apical cleavage c. 30 cm deep, obovate, distal half of margin rounded and praemorse, leathery, bronze-pink or red when newly emerged. Inflorescences c. 18 cm long including c. 2 cm peduncle with indumentum as upper internodes, branched to 1 order; prophyll not seen; rachillae c. 11 – 16 × 0.2 cm, 3 – 4 per inflorescence, scarlet; rachilla bracts inconspicuous; triads spirally arranged with alternating verticils of 3. Staminate flowers not seen. Pistillate flowers c. 4 – 5 × 3 mm after anthesis (when dry), crimson; sepals c. 2 × 3 mm, rounded, imbricate; petals c. 4 × 2 mm, consisting of a basal rounded portion c. 2 mm long, imbricate, and an upper triangular portion c. 2 mm long, valvate; ovary c. 3.5 × 2 mm, ellipsoid; staminodes 3, c. 0.5 mm long triangular and very thin. Fruits c. 1 cm long, cylindrical, pink. Seeds c. 7 × 1.5 mm (when dry), turbinate, endosperm ruminate (Fig. 7).

Hydriastele splendida. A habit; B leaf; C detail of leaf indumentum; D portion of stem and sheath with attached infructescence; E portion of rachilla with pistillate flowers; F pistillate flower whole and in longitudinal section; G fruit. Scale bar A = 40 cm; B = 8 cm; C = 1.5 cm; D = 4 cm; E = 7 mm; F = 3.3 mm; G = 5 mm. All from Baker et al. 820. drawn by lucy t. smith.
recognition. This species is distinguished by its entire bifid leaves in combination with inflorescences with 3 – 4 rachillae bearing triads in alternating verticils of 3, and seeds with ruminate endosperm.
distribution. Known only from the type locality in Papua Province (Indonesia) on the lower slopes of the Mt Jaya region (Dransfield et al. 2000; Map 7).

Distribution map of Hydriastele splendida.
specimens cited. indonesia. Papua Province: Mimika Regency (Previously Fakfak Regency), Timika, PT. Freeport Indonesia Area of Work, Km 63 road to Tembagapura, [4°21'S 136°58'E], 435 m, 7 Feb. 1998, Baker et al. 820 (AAU!, BH!, BO!, K!, L!, MAN!).
habitat. The understorey of heath forest on outwashed sands and gravels on steep and mossy slopes with many fallen trees, in areas of high rainfall (Dransfield et al. 2000); 435 m elevation.
vernacular names and uses. None known.
conservation status. Data deficient (DD). More data are needed about the distribution and abundance of this species.
notes. Hydriastele splendida is a distinctive and exceptionally beautiful species (hence the species epithet). In habit and reproductive morphology, it is similar to H. pinangoides, to which it is undoubtedly closely related. However, H. splendida bears entire leaves, which are few in number and display a deep cleft in the broadly rounded, praemorse distal margin. The species can be confused with entire-leaved forms of H. flabellata, but H. flabellata differs significantly in its spicate inflorescences (occasionally with 2 rachillae or up to 3 rachillae known from cultivation) with spirally arranged triads with ± well-defined verticils and seeds with homogeneous endosperm.
Hydriastele splendida appears to be already well established in cultivation, often erroneously named as H. flabellata.
References
Baker, W. J. Maturbongs, R. A., Wanggai, J. & Hambali, G. G. (2000). Siphokentia. Palms 44: 175 – 181.
____ & Loo, A. H. B. (2004). A synopsis of the genus Hydriastele. Kew Bull. 59: 61 – 68.
____ & Couvreur, T. L. P. (2012). Biogeography and distribution patterns of Southeast Asian palms. In: D. J. Gower, K. G. Johnson, J. E. Richardson, B. R. Rosen, L. Rüber & S. T. Williams (eds), Biotic evolution and environmental change in Southeast Asia, pp. 164 – 190. Cambridge University Press.
Beccari, O. (1877). Raccolta di osservazioni botaniche intorno alle piante dell'arcipelago Indo-Malese e Papuano. Malesia 1: 28 – 34.
____ (1885). Reliquiae Schefferianae illustrazione di alcune palme viventi nel Giardino Botanico di Buitenzorg. Ann. Jard. Bot. Buitenzorg 2: 82.
____ (1905). Leptophoenix Becc. In: U. Martelli (ed.), Webbia; raccolta di scritti botanici 1: 298 – 299.
____ (1914). Neue Palmen Papuasiens. Bot. Jahrb. Syst. 52: 27 – 28.
____ (1923). Neue Palmen Papuasiens II. Bot. Jahrb. Syst. 58: 452 – 454.
____ (1934). Generi, specie e varietà nuove di palme gerontogee della tribù Arecaceae lasciate inedite dal Dr. Od. Beccari ed ordinate a cura di U. Martelli. Atti Soc. Tosc. Sc. Nat. Pisa, Mem. 44: 130 – 131.
Burret, M. (1933). Neue Palmen aus Neuguinea. Notizbl. Bot. Gart. Berlin-Dahlem 11: 709 – 711.
____ (1935). Neue Palmen aus Neuguinea II. Notizbl. Bot. Gart. Berlin-Dahlem 12: 339 – 343.
____ (1936). Die Palmengattungen Nengella Becc. und Leptophoenix Becc. Notizbl. Bot. Gart. Berlin-Dahlem 13: 314 – 316.
____ (1939). Burret, Palmae gesammelt in Neu Guinea von L.J. Brass. J. Arnold Arbor. 20: 206 – 207.
Dowe, J. L. (2017). Baron Ferdinand von Mueller, the “Princeps of Australian Botany,” and a Historical Account of his Australian Palms. Palms 61: 21 – 40.
____ & Ferrero, M. D. (2000a). Gronophyllum cariosum, an ornamental new species from Papua New Guinea. Palms 44: 161 – 165.
____ & Ferrero, M. D. (2000b). A new species of rheophytic palm from New Guinea. Palms 44: 194 – 197.
Dransfield, J., Baker, W. J., Heatubun, C. D. & Witono, J. (2000). The Palms of Mount Jaya. Palms 44: 202 – 208
____, Uhl, N. W., Asmussen, C. B., Baker, W. J., Madeline, M. H. & Lewis, C. E. (2008). Genera Palmarum: The Evolution and Classification of Palms. Kew Publishing.
Essig, F. B. & Young, B. E. (1985). A reconsideration of Gronophyllum and Nengella (Arecoideae). Principes 29: 129 – 137.
Govaerts, R., Dransfield, J., Zona, S., Hodel, D. R. & Henderson, A. (2017). World Checklist of Arecaceae. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet; http://wcsp.science.kew.org Retrieved 22 September 2017
Heatubun, C. D. (2000). In search of Caryota zebrina — A palm expedition to the Cyclops Mountains. Palms 44: 187 – 193.
____ (2005). The rediscovery of Beccari’s Nengella flabellata in Irian Jaya Barat. Folia Malaysiana 6: 27 – 34.
Loo, A. H. B., Dransfield, J., Chase, M. W. & Baker, W. J. (2006). Low-copy nuclear DNA, phylogeny and the evolution of dichogamy in the betel nut palms and their relatives (Arecinae; Arecaceae). Molec. Phylogenet. Evol. 39: 598 – 618.
Mueller, F. (1880). Select extra-tropical plants readily eligible for industrial culture or naturalisation, p. 163. Indian Ed. Calcutta.
Petoe, P., Cámara-Leret, R. & Baker, W. J. (2018). A monograph of the Hydriastele wendlandiana group (Arecaceae: Hydriastele). Kew Bull. 73 (1). https://doi.org/10.1007/S12225-018-9736-7
Ridley, H. N. (1916). Report on the Botany of the Wollaston Expedition to Dutch New Guinea, 1912 – 13. Trans. Linn. Soc. London, Bot. 9: 232
Scheffer, R. H. C. C. (1876). Sur quelques palmiers du groupe des Arécinées. Ann. Jard. Bot. Buitenzorg 1: 103 – 164.
Schumann, K. M. & Lauterbach, C. (1900). Die Flora der Deutschen Schutzgebiete in der Südsee, p. 208. Gebruder Borntraeger, Leipzig.
Wendland, H. & Drude, O. (1875). Palmae Australasicae, Praecedit dissertatio de Arecinarum generibus gerontogaeis. Linnaea 39: 182.
Young, B. E. (1985). A new species of Gronophyllum (Palmae) from Papua New Guinea. Principes 29: 138 – 141.
Acknowledgements
We acknowledge the support of the directors and curators of the following herbaria for providing access to specimens: A, AAU, BO, BRI, CANB, FI, K, L, LAE and MEL. Himmah Rustiami (Herbarium Bogoriense) provided valuable information on types at BO. We also thank John Dransfield for his support and advice. We would especially like to acknowledge the numerous institutions and counterparts in Indonesia and Papua New Guinea who have made our field work on New Guinea palms possible. Lucy Smith prepared the botanical illustrations for this work. This research was funded by a grant to the Palms of New Guinea Project through the BAT Biodiversity Partnership and was further facilitated by grants from the Bentham Moxon Trust and the International Palm Society Endowment Fund to CDH and WJB.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Heatubun, C.D., Petoe, P. & Baker, W.J. A monograph of the Nengella group of Hydriastele (Arecaceae). Kew Bull 73, 18 (2018). https://doi.org/10.1007/s12225-018-9743-8
Accepted:
Published:
DOI: https://doi.org/10.1007/s12225-018-9743-8