
Abstract
This review is partly composed of the presentation “Cytokine alteration and speculated immunological pathophysiology in silicosis and asbestos-related diseases” delivered during the symposium “Biological effects of fibrous and particulate substances and related areas” organized by the Study Group of Fibrous and Particulate Studies of the Japanese Society of Hygiene and held at the 78th Annual Meeting in Kumamoto, Japan. In this review, we briefly introduce the results of recent immunological analysis using the plasma of silica and asbestos-exposed patients diagnosed with silicosis, pleural plaque, or malignant mesothelioma. Thereafter, experimental background and speculation concerning the immunological pathophysiology of silica and asbestos-exposed patients are discussed.
Similar content being viewed by others


Introduction
This special issue of Environmental Health and Preventive Medicine is organized to introduce the contents of a symposium provided by the Study Group of Fibrous and Particulate Studies (SGFPS) of the Japanese Society of Hygiene (JSH) and held at the 78th Annual Meeting in Kumamoto, Japan, which was entitled “Biological effects of fibrous and particulate substances and related areas”. At this symposium, our research group presented a mini-lecture entitled “Cytokine alteration and speculated immunological pathophysiology in silicosis and asbestos-related diseases”. Although it would be beneficial to provide a summary here, our group has already presented reviews twice in Environmental Health and Preventive Medicine as parts of special issues. One review was entitled “Keynote lecture at the 13th Japanese Society of Immunotoxicology (JSIT 2006)––Pathophysiological development and immunotoxicology: what we have found from research related to silica and silicate such as asbestos”, and was issued in July, 2007 [1, 2]. This mini-review was part of “Report of the 13th annual meeting of the Japanese Society of Immunotoxicology (JSIT 2006)”, and a brief summary of our investigations regarding the immunological effects of silica and asbestos was presented [1, 2]. The other review was presented as “Immunological alterations found in mesothelioma patients and supporting experimental evidence”, published online in February and on paper in March, 2008 [3, 4]. In this mini-review, we presented immunological alterations found in patients with malignant mesothelioma (MM) and described the experimental background in which data were obtained from clinical samples [3, 4].
Thus, to avoid too much overlap of descriptions from the two reviews mentioned above, we briefly introduce recent and preliminary immunological results obtained from the plasma of silica and asbestos-exposed patients diagnosed with silicosis (SIL), pleural plaque (PP), or MM. Thereafter, experimental background and speculation concerning the immunological pathophysiology of silica and asbestos-exposed patients are discussed.
Cytokines in patients with SIL, PP or MM compared with healthy donors (HD)
To screen the immunological status of silica and asbestos-exposed patients diagnosed with SIL, PP, or MM, plasma cytokine levels were measured using the cytometric bead array of the human Th1/Th2 cytokine kit II (CBA, BD Bioscience, San Jose, CA, USA), and measurements were made using FACSCalibur flow-cytometry (BD Bioscience) according to the manufacturer’s instructions [5]. This kit measures interferon (IFN)-γ, tumor necrosis factor (TNF)-α, and interleukins (IL)-10, 6, 4, and 2. Data were analyzed using only patient samples with determined values that were more than the limit of detection of the measurement.
The following patients were included in the study: HD (n = 10), M:F = 7:3, age = 51.6 ± 10.6; SIL (n = 10), M:F = 10:0, age = 73.2 ± 7.4; PP (n = 15), M:F = 8:2, age = 70.2 ± 5.4; and MM (n = 8), M:F = 7:1, age = 67.4 ± 0.8.5. The SIL patients were brickyard workers from Bizen City, Okayama prefecture, Japan. The amount of free silica inhaled by these patients was estimated to be as high as 40–60% in their work environment. The subjects were diagnosed with pneumoconiosis according to the International Labor Office (ILO) 2000 [6]. They showed no clinical symptoms of autoimmune disorders. PP and MM patients were diagnosed in Okayama Rosai Hospital, Okayama, Japan. Three PP and five MM patients were working in the shipbuilding industry, five PP and one MM patient were making asbestos fibers, three PP patients were working in chemical industries, one PP patient was a construction worker, and two MM patients were working in furnace installation. Specimens were taken only after informed consent had been obtained. The study was approved by the Ethics Committee of Kawasaki Medical School, Kusaka Hospital, and Okayama Rosai Hospital.
As shown in Fig. 1, significant differences were found for IL-6 level. MM patients showed higher IL-6 levels than those of other groups.

Comparison of plasma cytokine levels in healthy donors (HD) and in patients with PP, malignant mesothelioma (MM), or silicosis (SIL). The following patients were included in the study: HD (n = 10), M:F = 7:3, age = 51.6 ± 10.6; SIL (n = 10), M:F = 10:0, age = 73.2 ± 7.4; PP (n = 15), M:F = 8:2, age = 70.2 ± 5.4; and MM (n = 8), M:F = 7:1, age = 67.4 ± 0.8.5. Plasma cytokines were measured using the cytometric bead array of the human Th1/Th2 cytokine Kit II, and measurements were made using FACSCalibur flow-cytometry according to the manufacturer’s instructions. Statistical analysis was performed using StatFlex version 5.0 software (Artech, Osaka, Japan) and StatView-J 5.0 software (SAS Institute, Cary, NC, USA)
In addition, it may be worth noting that the supportive experimental data regarding IFN-γ indicated that IFN-γ tended to be lower in MM than HD. A human T-cell leukemia virus type-1 (HTLV-1)-immortalized human polyclonal T cell line, MT-2, was used to establish a continuous exposure model of immune-competent cells to asbestos [7]. As previously reported [8–10], MT-2 cells died as a result of apoptosis mediated by production of reactive oxygen species (ROS) and activation of the mitochondrial apoptotic pathway following temporary exposure to chrysotile, an asbestos, at relatively high doses (25–50 μg/ml), which are similar to experimental doses that produced alveolar epithelial and pleural mesothelial cell apoptosis in vitro [11–15]. However, a low dose (5–10 μg/ml) and continuous exposure (more than 8 months) to chrysotile resulted in the acquisition of resistance to asbestos-induced apoptosis in MT-2 cells [9, 10, 16]. A comparison of cellular and molecular characteristics of original MT-2 cells and those of a continuously exposed subline, designated CB-1 to reflect exposure to chrysotile-B, revealed alterations of cytokine production (Fig. 2), activation of Src-family kinase, phosphorylation of signal transducers and activators of transcription (STAT)-3, and upregulation of Bcl-2 [9, 10, 16]. Figure 2 shows the excess production of IL-10 and reduced secretion of IFN-γ, TNF-α, and IL-6.

Comparison of cytokine secretion into culture supernatants between original MT-2 cells and subline CB-1, which was continuously exposed to chrysotile for more than eight months. Cytokines were measured using the cytometric bead array of the human Th1/Th2 cytokine kit II, and measurements were made using FACSCalibur flow-cytometry according to the manufacturer’s instructions. Statistical analysis was performed using StatFlex version 5.0 software (Artech) and StatView-J 5.0 software (SAS Institute)
Among the results obtained from the experimental model of continuous exposure of T cells to chrysotile, the reduction of IFN-γ was compatible with results shown in Fig. 1. MM patients showed decreased plasma IFN-γ compared with the other three patient groups. However, there were some differences in cytokine levels between data from the experimental model and clinical samples. We showed previously that the plasma level of IL-10 was higher in MM than HD [10], and the discrepancy between Fig. 1 and our previous findings might be because of the small number of subjects. The data presented here are somewhat preliminary and need to be augmented with measurements from more patients. At the very least, IL-10 may play an important role in the acquisition of resistance to asbestos-induced apoptosis by upregulation of Bcl-2 located downstream of activated STAT-3 driven by IL-10 [17, 18], and in the functional enhancement of the CD4+25+FoxP3+ regulatory T cell (Treg), as a main soluble factor, and transforming growth factor (TGF)-β [19–23]. The role of IL-10 in the immune system of patients exposed to silica/asbestos should be investigated with regard to the disruption of self-tolerance and modification of tumor immunity.
The decrease of IFN-γ was similar in the experimental model and clinical samples. Because disruption of signaling pathways related to IFN-γ was found in our experimental model (data not shown, paper in preparation), IFN-γ may play an important role in immunological alteration in patients with asbestos-related diseases. It will be necessary to clarify modifications of the IFN-γ-signaling pathway during cancer progression of asbestos-induced malignant tumors.
The higher IL-6 level in MM patients was not compatible with the experimental model. However, IL-6 is known to be altered when malignant tumors develop in patients [24–26]. Thus, we need to consider the alteration of IL-6 production due to the cancer-related change and the direct effect of asbestos on immune-competent cells. In addition, IL-6 should be evaluated as one of the plasma markers for MM [27, 28].
In contrast, SIL patients showed no particular alteration of measured cytokines. Although we previously reported an increase of anti-nuclear antibody (ANA)-titer and soluble Fas in SIL [29–33], no noteworthy changes of cytokines were found. It is well known that the typical immunological complications in SIL are associated with autoimmune diseases [34–37]. Generally, Th1 dominance is found in patients with generalized autoimmune diseases such as systemic lupus erythematosus (SLE) and systemic scleroderma (SSc) whereas Th2 dominance is present in patients with allergies and organ-specific autoimmune diseases such as Hashimoto’s disease and type I diabetes mellitus [38, 39]. Thus, SIL might show a tendency towards Th1 dominance as an initial stage of generalized autoimmune disorders, because complicated autoimmune diseases associated with SIL are usually a generalized type such as SSc or rheumatoid arthritis (known as Caplan’s syndrome [40–42]) [34–37]. Although IL-2 level did not differ from that in other groups measured, some markers such as soluble IL-2 receptor may be altered in SIL as pre-autoimmune disease status. It will be necessary to find some markers which indicate immune activation in SIL. In addition, we previously reported that silica slowly activates peripheral T cells in vitro regarding the expression of the early T cell activation marker CD69 [43]. Silica may activate the immune system chronically in SIL patients, and chronically activated cells may play some role in the development of disruption of self-tolerance.
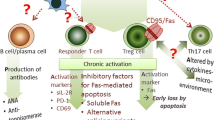
Speculative cellular mechanisms concerning autoimmune disorders in SIL
Treg cells have been widely investigated because of their critical role in the regulation of T cell homeostasis, as recorded for the autoimmune reaction, tolerance after a transplant, and the prevention of graft versus host diseases (GVHD), allergies, hypersensitivity, and tumor immunity. The reduction of the size and function of Treg cells is thought to result in an excess auto-reaction that causes autoimmune disorders [19–23]. From this viewpoint, we reported previously that the suppressive function in SIL of the peripheral CD4+25+ T cell fraction, in which functional Treg is located, was reduced compared with that of HD [44].
Speculation based on these data might suggest that:
-
1
silica affects CD4+25− responder T cells, resulting in their chronic and continuous activation;
-
2
these activated T cells change their phenotype towards CD4+25+ and substitute for Treg in the CD4+25+ fraction;
-
3
silica may affect CD4+25+FoxP3+ Treg, leading to their chronic and continuous activation;
-
4
these activated Treg exhibit a much higher change in CD95 (Fas; death receptor) expression; and
-
5
Fas-overexpressing Treg may be susceptible to Fas-mediated apoptosis.
During these cellular events, reduction of Treg size and function may occur in SIL to cause subsequent activation of autoimmunity, as shown in Fig. 3.

Schematic diagram of speculative cellular mechanisms relating to the disruption of self-tolerance and subsequent development of autoimmune disorders in SIL. PD-1 is the molecule that is expressed only in activated T cells, and not in Treg cells
Speculative cellular mechanisms concerning tumor immunity in MM
As mentioned above, MT-2 cells were used to create an in-vitro model of chronic and continuous exposure of T cells to asbestos [8–10, 16]. The continuously exposed subline CB-1 showed resistance to asbestos-induced apoptosis with activation of Src family kinase, upregulation of IL-10, phosphorylation of STAT-3, and upregulation of Bcl-2. Although it is not yet certain whether genetic alteration of CB-1 cells is caused by asbestos exposure, it is significant that although fewer cells obtained resistance to asbestos-induced apoptosis, gene expression level, at least, expanded during long-term culture with low-dose asbestos. In addition, TGF-β was also upregulated in CB-1 cells compared with original MT-2 cells, as shown in Fig. 4. In combination with data indicating that MT-2 shows a CD4+25+ phenotype and Treg is susceptible to HTLV-1 [45–48], the results suggest that MT-2 may be derived from immortalized Treg. Based on this speculation, upregulation of IL-10 and TGF-β is very important in consideration of the function of Treg, because both cytokines are typical soluble factors for Treg function and cell–cell contact [19–23]. If exposure to asbestos enhances the function of Treg, the mechanisms involved in the aggressive progression of mesothelioma after a very long latent period (approximately 40 years [49–51]) may be regarded the basis of immunological alteration in asbestos-exposed patients. At some point during the long latent period after initial exposure, disruption of tumor immunity in asbestos-exposed patients may occur, resulting in the rapid growth of tumor cells.

Schematic diagram of speculative cellular and molecular events involving MT-2 cells continuously exposed to the asbestos known as chrysotile
Conversely, inhibition of the disruption of tumor immunity may make tumor cells static and prevent rapid progression. This may be one approach for analysis of the immunological effects of asbestos. Of course, because there are many other immune-competent cell types involved in tumor immunity, for example natural killer (NK) cells [52, 53], CD8+ cytotoxic T cells [54, 55], and natural killer T (NKT) cells [56, 57], the effects of asbestos on these cells should also be analyzed to better recognize the occurrence of disrupted tumor immunity in patients with asbestos-related cancer such as MM. Particularly, regarding Nk cells, Froom et al. reported NK activity in asbestos-related workers [58]. They measured NK cell number and activity in 1,052 retired asbestos workers without symptomatic lung disease, lung cancer, or mesothelioma and with a long latency period from exposure; results were compared with those for 100 healthy age-matched controls. They reported the exposed workers showed a decreased NK cell activity and increased NK cell number, yielding a 10.8 higher odds ratio for low NK activity per cell compared with controls, which was due both to a decrease in NK cell activity and to an increase in NK cell number. Asbestos exposure of ten years or more increased the risk of low NK activity per cell. They conclude that exposure to asbestos is associated with diminished effectiveness of NK cells and a concomitant increase in the number of NK circulating cells. On the basis of these reports this should be investigated in the view of recent advances in the molecular and cellular biology of NK cells [53].
Although all these reports have not yet been completely confirmed, there is much literature showing the increased risk of nonthoracic cancers possibly resulting from asbestos exposure [59]. The effects of asbestos on immune-competent cells, described in this mini-review, may enhance hazardous cancer risks in the entire human body by alteration of tumor immunity.
References
Otsuki T. Immunotoxicological researches and preventive medicine. Foreword—the report from the 13th Annual Meeting of the Japanese Society of Immunotoxicology (JSIT 2006). Environ Health Prev Med. 2007;12:151–2.
Otsuki T, Miura Y, Maeda M, Hayashi H, Dong M, Nishimura Y. Keynote lecture in the 13th Japanese Society of Immunotoxicology (JSIT 2006)—pathophysiological development and immunotoxicology: what we have found from the researches related to the silica and silicate such as asbestos. Environ Health Prev Med. 2007;12:153–64.
Otsuki T. Reviews of a symposium entitled “malignant mesothelioma” organized by the Study Group on Fibrous and Particulate Substances of the 77th Annual Meeting of the Japanese Society of Hygiene, 2007. Environ Health Prev Med. 2008;13:53–4.
Miura Y, Nishimura Y, Maeda M, Murakami S, Hayashi H, Fukuoka K, et al. Immunological alterations found in mesothelioma patients and their experimental evidences. Environ Health Prev Med. 2008;13:55–9.
Takahashi K, Otsuki T, Mase A, Kawado T, Kotani M, Ami K, et al. Negatively-charged air conditions and responses of the human psycho-neuro-endocrino-immune network. Environ Int. 2008;34:765–72.
Guidelines for the Use of the ILO International Classification of Radiographs of Pneumoconioses, Revised Edition 2000, ILO (International Labour Organization), Geneva, Switzerland.
Miyoshi I, Kubonishi I, Yoshimoto S, Akagi T, Ohtsuki Y, Shiraishi Y, et al. Type C virus particles in a cord T-cell line derived by co-cultivating normal human cord leukocytes and human leukaemic T cells. Nature. 1981;294:770–1.
Hyodoh F, Takata-Tomokuni A, Miura Y, Sakaguchi H, Hatayama T, Hatada S, et al. Inhibitory effects of anti-oxidants on apoptosis of a human polyclonal T cell line, MT-2, induced by an asbestos, chrysotile-A. Scand J Immunol. 2005;61:442–8.
Otsuki T, Maeda M, Murakami S, Hayashi H, Miura Y, Kusaka M, et al. Immunological effects of silica and asbestos. Cell Mol Immunol. 2007;4:261–8.
Maeda M, Miura Y, Nishimura Y, Murakami S, Hayashi H, Kumagai N, et al. Immunological changes in mesothelioma patients and their experimental detection. Clin Med Circ Resp Pulm Med. 2008;2:11–7.
Saffiotti U. Respiratory tract carcinogenesis by mineral fibres and dusts: models and mechanisms. Monaldi Arch Chest Dis. 1998;53:160–7.
Kamp DW, Graceffa P, Pryor WA, Weitzman SA. The role of free radicals in asbestos-induced diseases. Free Radic Biol Med. 1992;12:293–315.
Brody AR, Liu JY, Brass D, Corti M. Analyzing the genes and peptide growth factors expressed in lung cells in vivo consequent to asbestos exposure and in vitro. Environ Health Perspect. 1997;105S5:1165–71.
Pass HI, Mew DJ. In vitro and in vivo studies of mesothelioma. J Cell Biochem Suppl. 1996;24:142–51.
Godleski JJ. Role of asbestos in etiology of malignant pleural mesothelioma. Thorac Surg Clin. 2004;14:479–87.
Miura Y, Nishimura Y, Katsuyama H, Maeda M, Hayashi H, Dong M, et al. Involvement of IL-10 and Bcl-2 in resistance against an asbestos-induced apoptosis of T cells. Apoptosis. 2006;11:1825–35.
Alas S, Bonavida B. Rituximab inactivates signal transducer and activation of transcription 3 (STAT3) activity in B-non-Hodgkin’s lymphoma through inhibition of the interleukin 10 autocrine/paracrine loop and results in down-regulation of Bcl-2 and sensitization to cytotoxic drugs. Cancer Res. 2001;61:5137–44.
Alas S, Bonavida B. Inhibition of constitutive STAT3 activity sensitizes resistant non-Hodgkin’s lymphoma and multiple myeloma to chemotherapeutic drug-mediated apoptosis. Clin Cancer Res. 2003;9:316–26.
Hori S, Takahashi T, Sakaguchi S. Control of autoimmunity by naturally arising regulatory CD4+ T cells. Adv Immunol. 2003;81:331–71.
Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T cells and immune tolerance. Cell. 2008;133:775–87.
Askenasy N, Kaminitz A, Yarkoni S. Mechanisms of T regulatory cell function. Autoimmun Rev. 2008;7:370–5.
Roncarolo MG, Gregori S, Battaglia M, Bacchetta R, Fleischhauer K, Levings MK. Interleukin-10-secreting type 1 regulatory T cells in rodents and humans. Immunol Rev. 2006;212:28–50.
Liu H, Leung BP. CD4+CD25+ regulatory T cells in health and disease. Clin Exp Pharmacol Physiol. 2006;33:519–24.
Kurebayashi J. Regulation of interleukin-6 secretion from breast cancer cells and its clinical implications. Breast Cancer. 2000;7:124–9.
Atreya R, Neurath MF. Involvement of IL-6 in the pathogenesis of inflammatory bowel disease and colon cancer. Clin Rev Allergy Immunol. 2005;28:187–96.
Hodge DR, Hurt EM, Farrar WL. The role of IL-6 and STAT3 in inflammation and cancer. Eur J Cancer. 2005;41:2502–12.
Upham JW, Garlepp MJ, Musk AW, Robinson BW. Malignant mesothelioma: new insights into tumour biology and immunology as a basis for new treatment approaches. Thorax. 1995;50:887–93.
Fitzpatrick DR, Peroni DJ, Bielefeldt-Ohmann H. The role of growth factors and cytokines in the tumorigenesis and immunobiology of malignant mesothelioma. Am J Respir Cell Mol Biol. 1995;12:455–60.
Tomokuni A, Aikoh T, Matsuki T, Isozaki Y, Otsuki T, Kita S, et al. Elevated soluble Fas/APO-1 (CD95) levels in silicosis patients without clinical symptoms of autoimmune diseases or malignant tumours. Clin Exp Immunol. 1997;110:303–9.
Otsuki T, Sakaguchi H, Tomokuni A, Aikoh T, Matsuki T, Kawakami Y, et al. Soluble Fas mRNA is dominantly expressed in cases with silicosis. Immunology. 1998;94:258–62.
Otsuki T, Ichihara K, Tomokuni A, Sakaguchi H, Aikoh T, Matsuki T, et al. Evaluation of cases with silicosis using the parameters related to Fas-mediated apoptosis. Int J Mol Med. 1999;4:407–11.
Tomokuni A, Otsuki T, Isozaki Y, Kita S, Ueki H, Kusaka M, et al. Serum levels of soluble Fas ligand in patients with silicosis. Clin Exp Immunol. 1999;118:441–4.
Otsuki T, Sakaguchi H, Tomokuni A, Aikoh T, Matsuki T, Isozaki Y, et al. Detection of alternatively spliced variant messages of Fas gene and mutational screening of Fas and Fas ligand coding regions in peripheral blood mononuclear cells derived from silicosis patients. Immunol Lett. 2000;72:137–43.
Uber CL, McReynolds RA. Immunotoxicology of silica. Crit Rev Toxicol. 1982;10:303–19.
Steenland K, Goldsmith DF. Silica exposure and autoimmune diseases. Am J Ind Med. 1995;28:603–8.
Shanklin DR, Smalley DL. The immunopathology of siliconosis. History, clinical presentation, and relation to silicosis and the chemistry of silicon and silicone. Immunol Res. 1998;18:125–73.
Otsuki T, Miura Y, Nishimura Y, Hyodoh F, Takata A, Kusaka M, et al. Alterations of Fas and Fas-related molecules in patients with silicosis. Exp Biol Med (Maywood). 2006;231:522–33.
Romagnani S. Type 1 T helper and type 2 T helper cells: functions, regulation and role in protection and disease. Int J Clin Lab Res. 1991;21:152–8.
Lucey DR, Clerici M, Shearer GM. Type 1 and type 2 cytokine dysregulation in human infectious, neoplastic, and inflammatory diseases. Clin Microbiol Rev. 1996:532–62.
Caplan A. Rheumatoid disease and pneumoconiosis (Caplan’s syndrome). Proc R Soc Med. 1959;52:1111–3.
Morgan WK. Caplan’s syndrome, An interesting clinico-pathological occurrence. Ann Intern Med. 1961;55:667–72.
Caplan A, Payne RB, Withey JL. A broader concept of Caplan’s syndrome related to rheumatoid factors. Thorax. 1962;17:205–12.
Wu P, Hyodoh F, Hatayama T, Sakaguchi H, Hatada S, Miura Y, et al. Induction of CD69 antigen expression in peripheral blood mononuclear cells on exposure to silica, but not by asbestos/chrysotile-A. Immunol Lett. 2005;98:145–52.
Wu P, Miura Y, Hyodoh F, Nishimura Y, Hatayama T, Hatada S, et al. Reduced function of CD4+25+ regulatory T cell fraction in silicosis patients. Int J Immunopathol Pharmacol. 2006;19:357–68.
Chen S, Ishii N, Ine S, Ikeda S, Fujimura T, Ndhlovu LC, et al. Regulatory T cell-like activity of Foxp3+ adult T cell leukemia cells. Int Immunol. 2006;18:269–77.
Matsubara Y, Hori T, Morita R, Sakaguchi S, Uchiyama T. Delineation of immunoregulatory properties of adult T-cell leukemia cells. Int J Hematol. 2006;84:63–9.
Shimauchi T, Kabashima K, Tokura Y. Adult T-cell leukemia/lymphoma cells from blood and skin tumors express cytotoxic T lymphocyte-associated antigen-4 and Foxp3 but lack suppressor activity toward autologous CD8+ T cells. Cancer Sci. 2008;99:98–106.
Abe M, Uchihashi K, Kazuto T, Osaka A, Yanagihara K, Tsukasaki K, et al. Foxp3 expression on normal and leukemic CD4(+)D25(+)T-cells implicated in human T-cell leukemia virus type-1 is inconsistent with Treg cells. Eur J Haematol. 2008, 28 May [Epub ahead of print].
Ilgren EB, Wagner JC. Background incidence of mesothelioma: animal and human evidence. Regul Toxicol Pharmacol. 1991;13:133–49.
Lanphear BP, Buncher CR. Latent period for malignant mesothelioma of occupational origin. J Occup Med. 1992;34:718–21.
O’Reilly KM, Mclaughlin AM, Beckett WS, Sime PJ. Asbestos-related lung disease. Am Fam Physician. 2007;75:683–8.
Burgess SJ, Maasho K, Masilamani M, Narayanan S, Borrego F, Coligan JE. The NKG2D receptor: immunobiology and clinical implications. Immunol Res. 2008;40:18–34.
Vivier E, Tomasello E, Baratin M, Walzer T, Ugolini S. Functions of natural killer cells. Nat Immunol. 2008;9:503–10.
Mescher MF, Popescu FE, Gerner M, Hammerbeck CD, Curtsinger JM. Activation-induced non-responsiveness (anergy) limits CD8 T cell responses to tumors. Semin Cancer Biol. 2007;17:299–308.
Kalinski P, Nakamura Y, Watchmaker P, Giermasz A, Muthuswamy R, Mailliard RB. Helper roles of NK and CD8+ T cells in the induction of tumor immunity. Polarized dendritic cells as cancer vaccines. Immunol Res. 2006;36(1–3):137–46.
Hong C, Park SH. Application of natural killer T cells in antitumor immunotherapy. Crit Rev Immunol. 2007;27:511–25.
Terabe M, Berzofsky JA. NKT cells in immunoregulation of tumor immunity: a new immunoregulatory axis. Trends Immunol. 2007;28:491–6.
Froom P, Lahat N, Kristal-Boneh E, Cohen C, Lerman Y, Ribak J. Circulating natural killer cells in retired asbestos cement workers. J Occup Environ Med. 2000;42:19–24.
Craighead JE. Nonthracic cancers possibly resulting from asbestos exposure. In: Craighead JE, Gibbs AR, editors. Asbestos and its diseases. New York: Oxford University Press; 2008. p. 230–52.
Acknowledgments
The authors thank Tamayo Hatayama, Shoko Yamamoto, Minako Kato, and Naomi Miyahara for technical assistance. Data obtained in the Department of Hygiene, Kawasaki Medical School, and published by the authors were supported by Special Coordination Funds for Promoting Science and Technology (H18-1-3-3-1), JSPS KAKENHI (19659153, 19790431, 19790411, 19890253 and 20390178), Kawasaki Medical School Project Grants (19-205Y, 19-506, 19-407 M and 19-603T), a Sumitomo Foundation Grant (053027), and a Yasuda Memorial Foundation Grant (H18).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Murakami, S., Nishimura, Y., Maeda, M. et al. Cytokine alteration and speculated immunological pathophysiology in silicosis and asbestos-related diseases. Environ Health Prev Med 14, 216–222 (2009). https://doi.org/10.1007/s12199-008-0063-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12199-008-0063-8