
Abstract
An effect of lithium (Li+) on growth, circumnutation, and glutamate-induced excitation in sunflower (Helianthus annuus L.) seedlings was investigated using time-lapse photography and extracellular electrical potential measurements. The seedlings were treated with a micro and millimolar concentration of lithium chloride (LiCl) both persistently in a hydroponic medium and acutely through Li+ injection. The length of hypocotyls, fresh weight of seedlings, intensity and period of circumnutation, and the number of action potentials (APs) after glutamate (Glu) injection were determined. It was found that the circumnutation intensity and period did not depend on hypocotyl length and fresh weight of seedlings. Under persistent Li+ treatment, the circumnutation intensity was constant at a concentration between 0.2 and 20 mM although the hypocotyls were significantly shorter in relation to the control, whereas at a concentration of 40 mM circumnutation intensity decreased without any changes in the hypocotyl length. Under persistent treatment with 0.5 and 20 mM Li+, the period of circumnutation was significantly prolonged. The number of APs in a Glu-induced series significantly increased in the seedlings exhibiting an Li+-induced decrease in circumnutation intensity (in 40 and 60 mM Li+). Additionally Li+ injection before Glu injection also augmented the series of APs in seedlings growing without Li+ in hydroponic medium. These Li+-sensitive responses demonstrated that circumnutation and growth are partly independent processes and reveal a relationship between circumnutation intensity and excitability in Helianthus annuus seedlings.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Plant shoots, stem, leaves, and roots move in a broad range of intensity and plants exhibit complex behaviour due to action of endogenous and environmental factors (Darwin and Darwin 1880). The plant endogenous movement named circumnutation is commonly considered as an inevitable consequence of organ growth (Engelmann and Johnsson 1998), but its quantitative relation to growth is still not well established (Johnsson 1997). The complex relations between circumnutation and growth have been reported for Oriza sativa (Hayashi et al. 2004; Yoshihara and Iino 2005), Arabidopsis thaliana (Schuster and Engelmann 1997), Phaseolus vulgaris (Millet and Badot 1996), Periploca graeca (Johnsson 1979), and Helianthus annuus (Zachariassen and Johnsson 1988; Stolarz et al. 2008). It is known that plants can grow without circumnutation but they can also grow and circumnutate simultaneously. Another not well-established process related to the movement is excitability. In animals, the relationship between the movement and excitability is apparent. In plants, a clear relation between rapid movement and action potential (AP) has been reported in sensitive Mimosa pudica and Dionaea muscipula. There are no reports concerning movement vs. excitability in ordinary plants that do not show spectacular rapid movements. In animals, both movements such as exploratory activity, hyper- and hypolocomotion, seizures (O’Donnell and Gould 2007; Ghasemi et al. 2010) and cortical excitability (Butler-Munro et al. 2010) are modulated by lithium (Li+) treatment. Lithium is a trace element naturally occurring in organisms. It occurs in soil and water resources and is taken up by all plants. It seems that Li+ is not required for plants’ physiological processes; however, the effect of Li+ on some of them has been reported (Aral and Vecchio-Sadus 2008). A micromolar Li+ treatment induces hypersensitivity-like response in Nicotiana tabacum (Naranjo et al. 2003) and metabolome and transcriptome changes in Brassica carinata seedlings (Li et al. 2009). In Phaseolus and Helianthus, circumnutation but not growth was affected by persistent Li+ application (Zachariassen and Johnsson 1988; Millet and Badot 1996). Lithium has also an effect on movements of Cassia fasciculata leaves (Gaillochet 1981) and Desmodium motorium lateral leaflets (Weber et al. 1992) and changes graviresponsiveness of roots in Pisum sativum (Belyavskaya 2001). Currently, it is postulated that Li+ acts via the inositol signalling pathway also involved in glutamate-receptor-, gravity-, and Ca2+-dependent plant responses (Stevenson et al. 2000). These responses together with Ca2+, K+ and Cl− ions and Ca2+, K+ and Cl− channels, and the H+ATP-ase actions are involved in the mechanism of growth, circumnutation, and excitability. The aim of our study was to characterise the effect of Li+ on growth, circumnutation, and series of glutamate-induced action potentials using a time-lapse photography method and extracellular electrical potential measurements in Helianthus annuus seedlings. To our knowledge, the influence of Li+ on plant excitability and its relation to plant endogenous movement have not been described so far. We have shown for the first time glutamate-induced plant excitability modulated by Li+—a neuroprotective agent present in environment.
Materials and methods
Experimental plants
Helianthus annuus L. seeds (PNOS, Ożarów Maz., Poland) were germinated on wet filter paper in a thermostated (24 ± 1 °C) darkened chamber. After 4 days, seedlings with 4.5 ± 0.5-cm-long hypocotyls were cultivated hydroponically (ten plants per pot) in aerated nutrient solution. The hydroponic culture was maintained for 3 days under constant illumination, 40 μmol m−2s−1 white light (Power Star HQT-T400 W/D OSRAM GmbH, Munich, Germany), at a temperature of 24 ± 1 °C and relative humidity 50–70 %. The seedlings were grown for 3 days in control medium or were treated with different LiCl or KCl concentrations and simultaneously filmed for circumnutation measurements. In seven-day-old seedlings, hypocotyl length and fresh weight of the seedlings were measured; and, electrophysiological measurements of the seedlings were performed. The growth rate (cm h−1) was calculated for hypocotyl growth during the 3 days of the experiment (hypocotyl length minus 4.5 cm divided by 72 h).
Circumnutation measurements
For circumnutation measurements, time-lapse photography recordings were made from 09:00 a.m. on the fourth day to 09:00 a.m. on the seventh day of seedling growth. A monochromatic camera (Mintron MTV-1368CD, Mintron Enterprise Co. Ltd, Taipei, Taiwan) was used to record the circumnutation trajectory of the hypocotyl apex. The plants were filmed from the top. Time-lapse images were recorded one frame per 5 min by Gotcha! Multicam software (Prescient System Inc., West Chester, PA, USA). The system was calibrated by a millimetre scale. The time-lapse images were digitised using Tracer (custom-made) and Microsoft Excel programs. Experimental points (coordinates x, y of the stem apex on the horizontal plane) were determined at 5-min intervals. The distance covered by the hypocotyls apex during 1 h was used to calculate the circumnutation rate. Circumnutation intensity was the rate of circumnutation divided by hypocotyl length. The circumnutation period was the duration of one circumnutation.
Electrophysiological measurements
The electrical measurements were carried out in a Faraday cage on 7- to 8-day-old seedlings. The changes in the electrical potential were measured with two extracellular Ag/AgCl electrodes (a silver wire, 0.2 mm diameter, World Precision Instruments, Sarasota, FL, USA) inserted across hypocotyls (Fig. 1). The electrical potential was recorded from tissues adjacent to the electrode, i.e. vascular bundles, parenchyma and epidermis. The reference electrode (Ag/AgCl) was placed in the hydroponic medium. Electrodes were interfaced with a data acquisition system Digidata 1322A (Axon Instruments, Foster City, CA, USA). The frequency of sample recording was 1 Hz. Electrophysiological experiments were carried out between 09:00 a.m. and 02:00 p.m. every day during the experimental time.

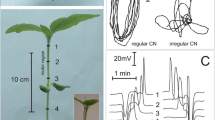
Scheme of a circumnutating Helianthus annuus seedling and electrode arrangement in extracellular electrical potential measurements. Two Ag/Cl electrodes (1, 2) were inserted across the hypocotyls. The reference electrode (ref) was placed in the hydroponic medium. i—The site of glutamate and lithium solution injection
Chemicals
The nutrient solution contained 4 mM Ca(NO3)2 × 4H2O, 5 mM KNO3, 1 mM NH4H2PO4, 2 mM MgSO4 × 7H2O; microelements: 0.085 mM Fe(III)citrate, 0.046 mM H3BO3, 0.0009 mM MnCl2 × 4H2O, 0.0003 mM CuSO4 × 5H2O, 0.0008 mM ZnSO4 × 7H2O, 0.0001 mM H2MoO4 × 2H2O; (pH 6.0). Additionally, 0.2, 0.5, 1, 5, 10, 20, 40, 60, 80 mM lithium chloride (MP Biomedicals, France) or 40 mM potassium chloride (POCH, Poland) in nutrient solution was used.
Injection Twenty microliters of a 50 mM Glu (l-glutamic acid, ICN Biomedicals, Germany) solution [pH adjusted to 7 by Tris/Mes buffer (Stolarz et al. 2010)], distilled water, LiCl (MP Biomedicals, France) or KCl (POCH, Poland) were injected with a syringe into the seedlings at the base of the hypocotyls, 1 cm above the root collar (Fig. 1). The injection of the solution lasted a few seconds.
Statistical analysis
The results obtained are presented as the mean ± SE in each experimental group. The results were estimated for significance by a t test at p < 0.05 and p < 0.01. The linear correlation coefficient was determined.
Results
Effect of lithium on growth and intensity of circumnutation of sunflower seedlings
The length of the hypocotyls of sunflower seedling growing in the control nutrient solution ranged between 9 and 19 cm and the mean was 13.8 ± 0.6 cm (n = 20) (Fig. 2a). The circumnutation rate of the hypocotyl was in the range from 0.1 to 4 cm h−1 and the mean was 1.7 ± 0.2 cm h−1. The correlation coefficient between the hypocotyl length and circumnutation rate was 0.73 (p < 0.01), as shown in Fig. 2a. Seedlings with longer hypocotyls exhibited a higher circumnutation rate, which shows that the circumnutation rate depends on the hypocotyl length. To distinguish the pure circumnutation vigour against the background of hypocotyl length changes, the circumnutation rate was divided by hypocotyl length and named circumnutation intensity (Fig. 2b). The mean circumnutation intensity for control seedlings was 0.12 ± 0.01 h−1. Circumnutation intensity does not depend in a statistically significant way either on hypocotyl length (correlation coefficient 0.54, p > 0.05) or fresh weight of seedlings (correlation coefficient 0.16, p > 0.05) (Fig. 2b, c).

The relationship between growth and circumnutation in one-week-old Helianthus annuus seedlings. a The circumnutation rate was the distance covered by the hypocotyl apex during 1 h; the mean hypocotyl length was 13.8 ± 0.6 cm; the mean circumnutation rate 1.7 ± 0.2 cm h−1. b Circumnutation intensity was the rate of circumnutation divided by the length of the hypocotyls; the mean circumnutation intensity 0.12 ± 0.01 h−1. c The mean fresh weight of the seedlings 1.16 ± 0.04 g. d The circumnutation period was the time between two nearest maxima of amplitude; the mean circumnutation period 167 ± 8 min. e The relation between the fresh weight of seedlings and the period of circumnutation; n = 20
The effect of permanent lithium treatment—lithium in the hydroponic nutrient solution—on hypocotyl length and circumnutation intensity is presented in Fig. 3. The hypocotyls were statistically significantly shorter in respect to the control at the sub- and low millimolar concentrations of LiCl (Fig. 3a). The hypocotyls were slightly shorter under 0.2, 0.5 (p < 0.05) and 5 and 10 mM LiCl (p < 0.01) in relation to the control seedlings. At the 20 and 40 mM LiCl concentrations, the hypocotyl length was similar to that in the control [13.8 ± 0.6 cm (n = 20)], but at 60 and 80 mM LiCl it drastically decreased to 9.1 ± 0.88 cm (n = 10) and 6.2 ± 0.22 cm (n = 10) (p < 0.01), i.e. it was reduced by 34 and 55 %, respectively (Fig. 3a). The 80 mM LiCl was highly toxic and caused withering of all plants after 3 days. Circumnutation intensity was also affected by Li+ treatment (Fig. 3b). Under 0.2–20 mM, circumnutation intensity was maintained at a constant level around 0.12 h−1, similar to the control experiment, and it drastically decreased under 40 and 60 mM LiCl (p < 0.05) (Fig. 3b). In 80 mM LiCl, circumnutation stopped completely. The statistically significant (p < 0.05) decrease in the hypocotyl length under 0.2–10 mM LiCl did not affect circumnutation intensity. Under 40 mM LiCl, circumnutation intensity significantly decreased without significant changes in hypocotyl length. LiCl at the concentration of 60 mM stopped circumnutation completely in 30 % of the seedlings and led to significant reduction of circumnutation intensity (p < 0.05) in 70 % of the seedlings. In this group, circumnutation was still maintained despite the strong decrease in growth. These results reveal that circumnutation intensity changes are independent of hypocotyl length changes under Li+ treatment in hydroponic medium in sunflower seedlings. The effect of Li+ on growth and circumnutation was compared with the KCl treatment (Fig. 3). In the 40 mM KCl solution, the hypocotyls were significantly shorter (p < 0.01) with respect to the control and to the 40 mM LiCl solution, and the intensity of circumnutation was similar to that in the control. This confirms possibility of independent regulation of growth and circumnutation by external factor.

Growth and circumnutation parameters of Helianthus annuus seedlings under lithium treatments. The seedlings were grown for 3 days in a nutrient solution with a lithium concentration from 0.2 to 80 mM; Values are mean ± SE; *p < 0.05; **p < 0.01 when compared with control. a Hypocotyl length changes. For the experiments, 4.5 ± 0.5 cm (dotted line) long hypocotyls of the seedlings were taken after 4 days germination. The hypocotyl growth rate (cm h−1) is shown on the bars. x—cessation of circumnutation. b Circumnutation intensity changes. c Circumnutation period changes
Effect of lithium on the period of circumnutation of sunflower seedlings
The mean circumnutation period in the control solution was 167 ± 13 min (n = 20) and did not depend on hypocotyl length and seedling fresh weight, as shown in Fig. 2d, e. The circumnutation period was affected by the Li+ treatment (Fig. 3c). It lengthened at the 0.5 and 20 mM LiCl concentrations in a statistically significant way (p < 0.05). At 0.5 mM, it was 192 ± 7 min (n = 10) and at 20 mM LiCl 194 ± 12 min (n = 10); thus the period was longer by approx. 30 min. At the higher concentration (40 mM LiCl and 40 mM KCl), the period was similar to that in the control. At 60 mM LiCl, the hypocotyl length and circumnutation intensity decreased but the period was still maintained around that of the control. This also confirms partial independence of the circumnutation mechanism on the hypocotyl growth, since both long and short hypocotyls exhibited the same period of circumnutation.
Glu-induced series of APs in sunflower seedlings
Injection of glutamate (Glu) into the hypocotyls of the sunflower seedlings growing in the control medium (n = 16) evoked series of APs (Table 1). In 50 % of the seedlings, the series were composed of 3.9 ± 0.6 APs and lasted 10 ± 3 min, n = 8. A typical recording is shown in Fig. 4a. The spikes were propagated, which is marked with an asterisk. In the remaining 50 % of the seedlings, one AP only or no APs were generated. To examine the effect of Li+ on Glu-induced excitation, Li+ was applied persistently through the hydroponic medium or acutely by injection 20 min before Glu injection.

Glutamate-induced excitation in Helianthus annuus seedlings. Example traces of a series of action potentials induced by 50 mM glutamate injection in the control experiment (a) and augmented series after lithium treatment (b). Propagation of action potentials is marked by asterisk. Electrode arrangement as in Fig. 1
Persistent lithium treatment—lithium in the hydroponic medium
The sunflower seedlings were growing at 5, 20, 40, and 60 mM LiCl concentrations for 3 days. In seedlings growing at the 5 and 20 mM LiCl, Glu-induced series were not significantly affected in relation to the control (Table 1). The injection of Glu into seedlings growing at the 40 and 60 mM LiCl concentrations evoked augmented excitation in 63 and 50 % of the seedlings. The series were composed of 11 ± 2 APs lasting 38 ± 6 min (n = 5), and 7.5 ± 1 APs lasting 24 ± 2 min (n = 4), respectively. This increase in the number of APs was statistically significant (p < 0.05). A typical recording is shown in Fig. 4b. The spike propagation is marked with an asterisk. In the remaining 37 and 50 % of the seedlings, one AP only or no APs were generated. Seedlings growing in 40 mM KCl showed Glu-induced series of APs similar to the control seedlings (Table 1).
Acute lithium treatment—lithium injection
To verify the effect of Li+ on Glu-induced series of APs, LiCl was injected 20 min before Glu injection (Table 1). The pre-injections were done in seedlings growing in the nutrient solution without Li+. Injection of 5, 10, 40 and 60 mM LiCl itself did not evoke series of APs. In the control experiments, distilled water was injected 20 min before Glu injection. Pre-injection of distilled water did not evoke series of APs and did not affect the Glu-induced series of APs (Table 1). In this experiment (n = 12), 58 % seedlings exhibited series composed of 4.4 ± 0.6 APs lasting 14 ± 2 min (n = 7), in the remaining 42 % of the seedlings, one AP only or no APs were generated. In the experiments with 5 and 10 mM LiCl pre-injection, no statistically significant increase in the number of Glu-induced APs was observed. In contrast, when 40 and 60 mM LiCl was pre-injected, a statistically significant (p < 0.01 and p < 0.001) increase in the number of Glu-induced APs was observed. In this case, the series were composed of 8.2 ± 1 APs and lasted 26 ± 5 min (n = 6) and 10.4 ± 0.9 APs and lasted 22 ± 4 min (n = 5), respectively. Pre-injection of 40 mM KCl into seedlings growing in the control medium solution did not affect the Glu-induced series of APs in a statistically significant way. In 58 % of the seedlings, they were composed of 5.5 ± 0.8 APs and lasted 11 ± 2 min (n = 6) in the remaining 42 % of the seedlings, one AP only or no APs were generated.
Discussion
Distinguishing between growth and circumnutating behaviour
Plant organs grow with a broad range of intensity. The values of the growth rate reported for Pinus silvestris hypocotyls were from 0.001 to 0.40 mm h−1 (Spurny 1975), for Helianthus annuus hypocotyls approx. 0.60 mm h−1 (Zachariassen and Johnsson 1988), and for three-week-old stems 0.06–0.36 mm h−1 (Stolarz et al. 2008). For shoots of Periploca graeca (Johnsson 1979) and hypocotyls of Arabidopsis thaliana (Schuster and Engelmann 1997), the threshold for circumnutation occurrence was approximately 0.5 and 0.05 mm h−1, respectively. The circumnutation vs. growth in one-week-old seedlings of Helianthus annuus has been presented herein. On average, the hypocotyl length was 6–14 cm, which corresponded to approximately 0.21–1.32 mm h−1 growth rate (Fig. 3a). First, we showed that the circumnutation rate in the control seedlings was correlated with the hypocotyl length in a linear way (Fig. 2a). This evidently supports the opinion that circumnutation is a growth-dependent movement. To analyse the pure circumnutation vigour, circumnutation intensity was calculated as the rate of circumnutation divided by hypocotyl length. The circumnutation intensity was the same in seedlings with long hypocotyls characterised by a high circumnutation rate and in seedlings with short hypocotyls with a lower circumnutation rate. The seedlings with equal-length hypocotyls exhibiting different circumnutation rates have different circumnutation intensities, therefore, this parameter characterises pure circumnutation vigour. Circumnutation intensity does not depend on the hypocotyl length, as it has been shown herein (Fig. 2b). Furthermore, it has also been shown that the Li+ persistent treatment modulates the hypocotyl length and circumnutation intensity but the circumnutation changes do not directly correspond to the changes in the hypocotyl length (Fig. 3a, b). Especially interesting was the application of 40 mM Li+ when the hypocotyl length did not change but circumnutation intensity decreased by half in respect to the control (Fig. 3a, b). A similar effect was observed in sunflower seedlings growing in red light in 20 mM Li+ (Zachariassen and Johnsson 1988). The other growth parameter assessed, i.e. the seedling fresh weight, was between 0.75 and 1.75 g (Fig. 2c). Our results showed that even a twofold increase in the seedling fresh weight did not influence circumnutation intensity so the latter does not depend on the seedling fresh weight. Zachariassen and Johnsson (1988) showed that circumnutation, but not growth, was inhibited by Li+ treatment in sunflower seedlings grown in red light. The same effect of Li+ was observed in circumnutating shoots of Phaseolus vulgaris (Millet and Badot 1996). Similar non-proportional changes in growth and circumnutation were observed in rice roots after aluminium treatment (Hayashi et al. 2004), in rice coleoptiles in a red light-induced response (Yoshihara and Iino 2005), and after ethylene stimulation in etiolated Col-0 Arabidopsis thaliana seedlings (Binder et al. 2006). Additionally, in Pisum sativum roots large circumnutations stopped completely although the growth was not stopped after the apex was removed (Spurny 1968). Our previous study of three-week-old sunflower showed a complex quantitative relation between the growth rate and the circumnutation rate in light–dark conditions (Stolarz et al. 2008). In the present study, a similar complex relationship has been shown in constant light under Li+ treatment of sunflower seedlings. Those literature examples together with our results presented above confirm the decoupling phenomenon between growth and circumnutation.
Lithium influences the biological clock in humans, rodents, unicellular organisms, and also in plants (Zachariassen and Johnsson 1988; Yin et al. 2006; O’Donnell and Gould 2007; Zarse et al. 2011; Hinrichsen et al. 2012). Here, we have shown that only 0.5 and 20 mM Li+ extended the circumnutation period by approx. 18 % (30 min). At higher concentrations, the circumnutation intensity and hypocotyl length decreased but the period was similar to that in the control experiments (Fig. 3d). Therefore, it is evident that the ultradian oscillator of circumnutation is independent of seedling growth but it is Li+ sensitive.
Based on the presented results, we postulate that circumnutation is partially independent of growth. There is evident autonomy of circumnutation in relation to elongation. The intensity and period of circumnutation were regulated without simple correspondence to hypocotyl length and fresh weight changes. Following Hayashi et al. (2004), we assume that circumnutation changes without growth inhibition, or growth inhibition without inhibition of circumnutation should be a part of early reaction to changes in the environmental conditions. Circumnutation can be disturbed until growth is disturbed, or growth can be disturbed without significant disturbance in the circumnutation intensity and period. The hypothesis about the partial autonomy of circumnutation in relation to growth contributes to explanation of a special ecological and physiological function of circumnutation being not only the way of growth (Darwin and Darwin 1880; Inoue et al. 1999; Larson 2000; Kosuge et al. 2013; Migliaccio et al. 2013).
Lithium augments glutamate-induced excitation of Helianthus annuus seedlings
Sunflowers with decreased intensity of circumnutation are more excitable (Fig. 3b; Table 1), as we have shown here. The essential role of K+, Cl− and Ca2+ in the mechanism of circumnutation is known; simultaneously, it is known that K+, Cl− and Ca2+ fluxes are essential for membrane potential maintenance and thus plant excitability. Excitability is an ability of an organism to generate spontaneous or stimulus-induced APs. Here, the ability to generate Glu-induced APs has been shown in one-week-old sunflower seedlings. In our previous study, we have demonstrated that injection of Glu induces series of APs and decreases circumnutation in three-week-old sunflower plants (Stolarz et al. 2010). In 75 % of plants, the series consisted of 6.9 ± 0.7 APs and lasted 15 ± 2 min. Here, in 58 % of the seedlings, Glu-induced series of APs were composed of 4.4 ± 0.6 APs and lasted 14 ± 2 min (Fig. 4a; Table 1). It is evident that seedlings exhibited a lower number of APs in relation to older plants; therefore, they were less excitable. We assume that this is connected with better vascular bundle maturation in older plants. This is the first time that we have shown that plant excitability is related to age. Similarly, Favre et al. (2011) have shown different excitability in different Arabidopsis thaliana accessions. Here, we found that persistent Li+ treatment (40 and 60 mM LiCl) augmented Glu-induced series of APs and decreased circumnutation intensity. The slowly circumnutating, smaller seedlings generated an augmented series composed even of 11 APs lasting about 30 min (Fig. 4b; Table 1), thus they had higher excitability than the control plants. The Li+ pre-injection confirmed this effect (Table 1). Here, we have shown lower and higher excitability in phenotypically different sunflower plants. A comparable approach was also presented by Ślesak (1996) when iron deficiency affected the motor and electrical activity of mimosa. To our knowledge, this is the first report showing Li+-modulated plant excitability. Micromolar to 100 mM LiCl concentrations were used in investigations with Li+ in plants (Gillaspy et al. 1995; Belyavskaya 2001; Bueso et al. 2007; Hawrylak-Nowak et al. 2012; Kalinowska et al. 2013). The micromolar Li+ concentration can affect biomass, leaf area, photosynthetic pigment accumulation, lipid peroxidation, and K+ content in sunflower and maize (Hawrylak-Nowak et al. 2012). Zachariassen and Johnsson (1988) and Millet and Badot (1996) showed that a micromolar LiCl concentration penetrated the root system of sunflower and the Li+ concentration inside of hypocotyls was tens of micromoles. Therefore, we assume that the active concentration of Li+ inside the hypocotyls in our study was micromolar as well. We assume that Li+ could act via signalling pathways. It is probable that Li+ acts by inhibition of inositol monophosphatase (Gillaspy et al. 1995) and disturbs the inositol triphosphate (IP3) level and thus deregulates the intracellular calcium levels [Ca2+] (Krol and Trebacz 2000; Stevenson et al. 2000; Krinke et al. 2007). Changes in the intracellular calcium concentration might affect the mechanism of circumnutation and action potential triggering. Another possible scenario is that Li+ (millimolar concentration) directly modulates transmembrane transport and thus affects the K+ status in plants (Gaillochet 1981; Millet and Badot 1996; Naranjo et al. 2003; Hawrylak-Nowak et al. 2012) as well as excitability and circumnutation behaviour. It is probable that the mechanism of Li+ and Glu action in plants shares some similarity with the Li+ and Glu action in animal cells (Lam et al. 1998; Davenport 2002; Forde and Lea 2007; Baluska 2010; Quiroz et al. 2010; Gardiner and Marc 2011).
In the present work, it has been shown that circumnutation and growth are partly independent. It was found that in sunflower seedlings that micro- and millimolar concentrations of Li+ decreased the growth and circumnutation but in a different way; furthermore, Li+ treatment augmented Glu-induced excitation. The persistent application and injections of neuroactive substances are standard methods in animal electrophysiology; here, the method has been successfully applied in vivo in a whole plant to study of plant behaviour. We are planning future behavioural, pharmacological, and electrophysiological investigations based on these Li+-dependent responses to understand better the role of the endogenous movement and excitability in plant growth.
Author contribution statement
Maria Stolarz designed and carried out the experiments, collected and analysed the results, and wrote the manuscript. Elzbieta Krol helped in the analysis of the results and editing the manuscript. Halina Dziubinska helped in the analysis of the results and editing the manuscript.
Abbreviations
- AP:
-
Action potential
- Glu:
-
Glutamate
- Li+ :
-
Lithium
References
Aral H, Vecchio-Sadus A (2008) Toxicity of lithium to humans and the environment—a literature review. Ecotox Environ Safe 70:349–356
Baluska F (2010) Recent surprising similarities between plant cells and neurons. Plant Signal Behav 5:87–89
Belyavskaya NA (2001) Lithium-induced changes in gravicurvature, statocyte ultrastructure and calcium balance of pea roots. In: Kiss JZ, Kern VD (eds) Space life sciences: gravity perception and transduction in plants, fungi and unicellular organisms. Elsevier Science Bv, Amsterdam, pp 961–966
Binder BM, O’Malley RC, Wang W, Zutz TC, Bleecker AB (2006) Ethylene stimulates nutations that are dependent on the ETR1 receptor. Plant Physiol 142:1690–1700
Bueso E, Alejandro S, Carbonell P, Perez-Amador MA, Fayos J, Belles JM, Rodriguez PL, Serrano R (2007) The lithium tolerance of the Arabidopsis cat2 mutant reveals a cross-talk between oxidative stress and ethylene. Plant J 52:1052–1065
Butler-Munro C, Coddington EJ, Shirley CH, Heyward PM (2010) Lithium modulates cortical excitability in vitro. Brain Res 1352:50–60
Darwin C, Darwin F (1880) The power of movement in plants. John Murray, London
Davenport R (2002) Glutamate receptors in plants. Ann Bot 90:549–557
Engelmann W, Johnsson A (1998) Rhythms in organ movement. In: Lumsden P, Millar A (eds) Biological rhythms and photoperiodism in plants. Bios Scientific Publishers, Oxford, pp 35–50
Favre P, Greppin H, Degli Agosti R (2011) Accession-dependent action potentials in Arabidopsis. J Plant Physiol 168:653–660
Forde BG, Lea PJ (2007) Glutamate in plants: metabolism, regulation, and signalling. J Exp Bot 58:2339–2358
Gaillochet J (1981) Effect of the lithium-chloride on the leaf movements of Cassia fasciculata. Planta 151:544–548
Gardiner J, Marc J (2011) Arabidopsis thaliana, a plant model organism for the neuronal microtubule cytoskeleton? J Exp Bot 62:89–97
Ghasemi M, Shafaroodi H, Nazarbeiki S, Meskar H, Heydarpour P, Ghasemi A, Talab SS, Ziai P, Bahremand A, Dehpour AR (2010) Voltage-dependent calcium channel and NMDA receptor antagonists augment anticonvulsant effects of lithium chloride on pentylenetetrazole-induced clonic seizures in mice. Epilepsy Behav 18:171–178
Gillaspy GE, Keddie JS, Oda K, Gruissem W (1995) Plant inositol monophosphatase is a lithium-sensitive enzyme encoded by a multigene family. Plant Cell 7:2175–2185
Hawrylak-Nowak B, Kalinowska M, Szymanska M (2012) A study on selected physiological parameters of plants grown under lithium supplementation. Biol Trace Elem Res 149:425–430
Hayashi Y, Nishiyama H, Tanoi K, Ohya T, Nihei N, Tanioka K, Nakanishi TM (2004) An aluminum influence on root circumnutation in dark revealed by a new super-HARP (high-gain avalanche rushing amorphous photoconductor) camera. Plant Cell Physiol 45:351–356
Hinrichsen RD, Belsky D, Jones LA, Mialki R (2012) The frequency of the spontaneous behavioral response in Paramecium tetraurelia is simultaneously modulated by both ultradian and circadian rhythms. Biol Rhythm Res 41:1–14
Inoue N, Arase T, Hagiwara M, Amano T, Hayashi T, Ikeda R (1999) Ecological significance of root tip rotation for seedling establishment of Oryza sativa L. Ecol Res 14:31–38
Johnsson A (1979) Circumnutation. In: Haupt W, Feinleib E (eds) Encyclopedia of plant physiology Physiology of Movements. Springer, Berlin, pp 627–646
Johnsson A (1997) Circumnutations: results from recent experiments on Earth and in space. Planta 203:147–158
Kalinowska M, Hawrylak-Nowak B, Szymanska M (2013) The Influence of two lithium forms on the growth, l-ascorbic acid content and lithium accumulation in lettuce plants. Biol Trace Elem Res 152:251–257
Kosuge K, Iida S, Katou K, Mimura T (2013) Circumnutation on the water surface: female flowers of Vallisneria. Sci Rep 3:1–7
Krinke O, Novotná Z, Valentová O, Martinec J (2007) Inositol trisphosphate receptor in higher plants: is it real? J Exp Bot 58:361–376
Krol E, Trebacz K (2000) ways of ion channel gating in plant cells. Ann Bot 86:449–469
Lam HM, Chiu J, Hsieh MH, Meisel L, Oliveira IC, Shin M, Coruzzi G (1998) Glutamate-receptor genes in plants. Nature 396:125–126
Larson KC (2000) Circumnutation behavior of an exotic honeysuckle vine and its native congener: influence on clonal mobility. Am J Bot 87:533–538
Li X, Gao P, Gjetvaj B, Westcott N, Gruber MY (2009) Analysis of the metabolome and transcriptome of Brassica carinata seedlings after lithium chloride exposure. Plant Sci 177:68–80
Migliaccio F, Tassone P, Fortunati A (2013) Circumnutation as an autonomous root movement in plants. Am J Bot 100:4–13
Millet B, Badot P (1996) The revolving movement mechanism in Phaseolus; New approaches to old questions. In: Greppin H, Degli Agosti R, Bonzon M (eds) Vistas on Biorhythmicity. University of Geneva, Geneva, pp 77–98
Naranjo MA, Romero C, Belles JM, Montesinos C, Vicente O, Serrano R (2003) Lithium treatment induces a hypersensitive-like response in tobacco. Planta 217:417–424
O’Donnell KC, Gould TD (2007) The behavioral actions of lithium in rodent models: leads to develop novel therapeutics. Neurosci Biobehav Rev 31:932–962
Quiroz JA, Machado-Vieira R, Zarate CA, Manji HK (2010) Novel insights into lithium’s mechanism of action: neurotrophic and neuroprotective effects. Neuropsychobiology 62:50–60
Schuster J, Engelmann W (1997) Circumnutations of Arabidopsis thaliana seedlings. Biol Rhythm Res 28:422–440
Ślesak E (1996) Effect of iron deficiency stress on leaves movements and electrical potentials in mimosa (Mimosa pudica L.). Acta Soc Bot Pol 65:283–289
Spurny M (1968) Effect of root tip amputation on spiral oscillations of growing hypocotyl with radicle of pea (Pisum sativum L.). Biol Plantarum 10:98–111
Spurny M (1975) Elongation and circumnutation oscillations of hypocotyl of pine seedlings (Pinus silvestris L.). Biol Plantarum 17:43–49
Stevenson JM, Perera IY, Heilmann I, Persson S, Boss WF (2000) Inositol signaling and plant growth. Trends Plant Sci 5:252–258
Stolarz M, Krol E, Dziubinska H, Zawadzki T (2008) Complex relationship between growth and circumnutations in Helianthus annuus stem. Plant Signal Behav 3:376–380
Stolarz M, Krol E, Dziubinska H, Kurenda A (2010) Glutamate induces series of action potentials and a decrease in circumnutation rate in Helianthus annuus. Physiol Plant 138:329–338
Weber U, Engelmann W, Mayer WE (1992) Effects of tetraethylammoniumchloride (TEA), vanadate, and alkali ions on the lateral leaflet movement rhythm of Desmodium motorium (Houtt) Merr. Chronobiol Int 9:269–277
Yin L, Wang J, Klein PS, Lazar MA (2006) Nuclear receptor Rev-erba is a critical lithium-sensitive component of the circadian clock. Science 311:1002–1005
Yoshihara T, Iino M (2005) Circumnutation of rice coleoptiles: its occurrence, regulation by phytochrome, and relationship with gravitropism. Plant, Cell Environ 28:134–146
Zachariassen E, Johnsson A (1988) Effects of lithum ions on the circumnutations of Helianthus hypocotyls. Physiol Plant 72:147–152
Zarse K, Terao T, Tian J, Iwata N, Ishii N, Ristow M (2011) Low-dose lithium uptake promotes longevity in humans and metazoans. Eur J Nutr 50:387–389
Acknowledgments
We thank Professor Kazimierz Trębacz for useful discussions and comments on the manuscript.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by A. Krolicka.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article

Cite this article
Stolarz, M., Król, E. & Dziubińska, H. Lithium distinguishes between growth and circumnutation and augments glutamate-induced excitation of Helianthus annuus seedlings. Acta Physiol Plant 37, 69 (2015). https://doi.org/10.1007/s11738-015-1814-y
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-015-1814-y