
Abstract
Three varieties (Alata, Pumila and Lanceolata) of Malaysian indigenous medicinal herb Labisia pumila Benth. grown in greenhouse and open field were tested to evaluate and compare their photosynthetic and maximal quantum efficiency of photosystem II (F v/F m) reactions. Every variety grown in greenhouse demonstrated higher light-saturated photosynthetic capacity than in the open field. The diurnal net photosynthesis (A) curve in the open field also displayed dual peaks with lower daily average A compared to the greenhouse. Varieties Alata and Pumila were found to acclimatize better under both growing conditions. The diurnal patterns of F v/F m indicated that plants grown under greenhouse encountered less photoinhibition than in open field condition. A decrease in chlorophyll (chl) a/b ratio in leaves of greenhouse plants with significant increase in chl b was observed. This study indicates that var. Alata and var. Pumila have the capacity to acclimatize to greenhouse growth condition.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Labisia pumila Benth. is a sub-herbacous plant with creeping stems from the family Myrsinaceae that is found widespread in Malaysian forest. It is an indigenous, understorey medicinal herb of Malaysia, popularly known as Kacip Fatimah, and sometimes also referred locally as Selusoh Fatimah, Rumput Siti Fatimah, Akar Fatimah, Tadah Matahari, Bunga Belangkas Hutan and Pokok Pinggang (Jamia et al. 2003). It has customarily been used by Malay women to induce and facilitate childbirth as well as a post-partum medicine (Burkill 1966). Stone (1988) categorized the three varieties of this herb in Malaysia as L. pumila var Alata, L. pumila var Pumila and L. pumila var Lanceolata. Each of the beneficial varieties has different usage. But the most universally utilized varieties by traditional healers are L. pumila var Alata and var Pumila. Other uses of this herb are for dysentery, dysmenorrhea, flatulence and gonnorhea treatments (Rozihawati et al. 2003). Due to the recent discovery of the estrogenic activities of this herb (Ezumi et al. 2006), the demand for this herb in the recent years has increased tremendously in the herbal and commercial industry (Jaafar et al. 2008).
Presently, L. pumila has been abundantly collected from the rainforest to suffice industry requirement. This system of harvest in Malaysia results in an unreliable supply and also inconsistent quality of this herb (Jaafar et al. 2008). With increasing demand for this herb over the years, the authors were concerned that it might endanger the species existence in nature and cause a loss of biodiversity to Malaysia. The effort to domesticate this herb commercially has not been too encouraging, due to the difficulty in raising the species away from its natural habitat. The plant is not only slow growing, but also, as highlighted by Jaafar et al. (2009), the species is very sensitive to changes in microclimate especially temperature, light and relative humidity. Therefore, the authors suggest that growing L. pumila under greenhouse could be one of the best ways to produce raw materials of this herb commercially, as the microenvironment of the growing plants can be controlled and manipulated. The productivity of L. pumila is a function of the production of assimilates by photosynthesis and translocation of assimilates to plant sinks (Jaafar et al. 2010). The alterations of photostynthesis rate (A) can also influence the plant growth and development processes (Lei et al. 1996). Under greenhouse condition, light is one of the important microclimate factors that influence plant productivity apart from temperature, relative humidity and CO2 concentrations. Despite the necessity of light for autotrophic organisms, no plant is capable of using 100% of solar irradiation for photosynthesis (Demmig et al. 1997). During plant acclimation to specific light environment, chlorophyll concentration, maximal quantum yield for photosystem II (F v/F m), chloroplast density, expression of antioxidant enzymes and Calvin cycle intermediate have been shown to fluctuate during this process (Demmig et al. 1997; Hernandez et al. 2004; Yoshimura et al. 2000). Nevertheless, little information is available on the photosynthetic acclimation responses of L. pumila either in the open field or greenhouse condition, which may be the basis for variety selection for commercial cultivation.
Plants grown in greenhouse and open field conditions vary their growth periods and growth responses very significantly, partly due to differences in the microclimates. In the open field, most plants receive more sunlight than they can actually use for photosynthesis, as a result photosynthesis produces more NADPH and ATP than required This accumulation of redox and energy will decrease the plastoquinone pool and inhibit the water splitting complex leading to PSII inactivation, the so-called photoinhibition (Karpinski et al. 1997). Under these circumstances, plants have to exhaust the excess light energy that had been absorbed. This will depend on a protective non-photochemical mechanism that quenches singlet-excited chlorophyll and harmlessly scour excess excitation energy as heat. The non-photochemical quenching (NPQ) process occurs in all photosynthetic eukaryotes and helps to control and shield photosynthesis in the environment, in which light energy absorption exceeds the capacity for light utilization (Patricia et al. 2001).
The main objective of this research was to determine the differences in photosynthetic responses of three varieties of L. pumila Benth. to different growing conditions (photosynthetic photon flux density, temperature, CO2 concentration) in the open field and greenhouse. We would also like to establish whether these varieties showed the ability to acclimatize under greenhouse conditions, which could ultimately be the basis for variety selection raised under greenhouses for commercial use.
Materials and methods
The site is situated at Field 2, University Agriculture Park, Universiti Putra Malaysia (longitude 101°44′N and latitude 2°58′S, 68 m above sea level) with a mean atmospheric pressure of 1.013 kPa. Three varieties of L. pumila used in this study, namely var Alata, Pumila and Lanceolata, were grown in a nursery plot in a greenhouse complex of the faculty of agriculture and measured during November of 2009. The seedlings were planted in soilless medium containing coco-peat, burnt paddy husk and well composted chicken manure in 5:5:1 (v/v) ratio in 25-cm diameter polyethylene bags. Day and night temperatures in the greenhouse were maintained at 27–30°C and 18–21°C, respectively, and relative humidity from 50 to 60%. Organic fertilizer was applied every 2 weeks using well-composted chicken manure at 20 g/plant. All the seedlings were irrigated using overhead mist irrigation given four times a day or when necessary. Each irrigation session lasted for 7 min.
A two-factor experiment was organized in a completely randomized design (CRD) with three varieties of L. pumila: var alata (Alata), var pumila (Pumila) and var lanceolata (Lanceolata) and two growing conditions (open field and greenhouse) replicated three times, and each treatment consisted of five plants. Standard error means (S.E.M.) was calculated and differences in mean values for each treatment were tested at p ≤ 0.05 according to Duncan multiple range test (DNMRT).
Total chlorophyll content was measured using a method from Hardwich and Baker (1973) based on fresh weight basis. Prior to each destructive harvest, seedling was analyzed for the leaf relative chlorophyll reading (SPAD meter 502, Minolta Inc, USA). The leaves of Labisia pumila with different greenness (yellow, light green and dark green) were selected for analysis and total leaf chlorophyll content was analyzed. For each type of leaf, the relative SPAD value was recorded (5 points/leaf) and sampled for chlorophyll content determination. Leaf disk 3 mm in diameter were obtained from leaf sample using a hole puncher. For each seedling, the measurements were conducted on the young fully expanded leaves; generally, on the second or third leaf from the tip of the stem was used. The leaf disks were immediately immersed in 20 ml of acetone in an aluminum foil-covered glass bottle for approximately 24 h at 0°C until all the green color had bleached out. Finally, 3.5 ml of the solution was measured using a spectrophotometer (UV-3101P, Labomed Inc, USA) at absorbances of 664 and 647 nm. Using the data obtained, the least squares regression was established to develop a predictive relation between pigment concentrations (mg/g fresh weight) attained from the destructive chlorophyll method and relative chlorophyll content from the SPAD meter readings.
Leaf gas exchange measurements were taken using closed system, infrared gas analyzer LICOR 6400 Portable Photosynthesis System (IRGA: LICOR Inc, Lincoln, NE, USA) set with optimal growth conditions by placing the cuvette head over fully expanded leaf supported by a tripod stand. Prior to the measurement, the maximum photosynthetic photon flux density for L. pumila was determined. The standard optimal conditions set for measurement of L. pumila were 800 μmol/m2/s photosynthetic photon flux density, 400 μmol/mol CO2, 30ºC leaf temperature and 60% relative humidity. Photosynthesis measurement was taken at 0800, 1000, 1200, 1400, 1600 and 1800 h, and the daily average photosynthesis was obtained by averaging the six measurements of net photosynthesis taken daily. Photosynthetic light response curves were measured at growth CO2 concentrations with an open flow infrared gas analyzer attached with a red LED light source (LI-6400, Li-Cor, Inc., Lincoln, NE, USA). Measurements began with saturating light (1,000 μmol/m2/s), followed by nine incremental light reductions (900, 800, 700, 600, 500, 400, 300, 200, 100 μmol/m2/s) until the final irradiance was 10 μmol/m2/s. Assimilation was recorded at each light level following a 15-min acclimation period. Use of decreasing light rather than increasing light reduces the equilibrium time required for stomatal opening and photosynthetic induction (Ibrahim et al. 2010; Kubiske and Pregitzer 1996). Photosynthetic light response curves were individually analyzed to determine the light-saturated net photosynthetic rate (A max), the light compensation point and light saturation point (Ibrahim et al. 2010; Givnish 1988).
Measurements of chlorophyll fluorescence were taken from fully expanded second leaf from the apex. Leaves were darkened for 20 min by attaching light-exclusion clips to the central region of the leaf surface. Chlorophyll fluorescence was measured using a portable chlorophyll fluorescence meter (Handy PEA, Hansatech Instruments Ltd, Kings, Lynn, UK). Measurements were recorded for 5 s (Philip et al. 2006). The fluorescence responses were induced by emitting diodes. Measurement of F o (initial fluorescence), F M (maximum fluorescence) and F V (variable fluorescence) were obtained from this procedure. F V is derived as the differences between F M and F o. The mean value of three representative plants was used to represent each sub-plot.
The microclimate characteristics outside and inside the glasshouse, particularly CO2 concentration, photosynthetic photon flux density and temperature, were observed to be largely different (Fig. 1). The CO2 concentration in the glasshouse was found to be higher than that outside at 0800 h (450 μmol/mol), but showed a big reduction than that outside at 1000 h (410 μmol/mol) and then started to increase again at 1800 h. Conversely, CO2 concentration outside was stable throughout the day and ranged from 419 to 425 μmol/mol (Fig. 1a). Photosynthesis photon flux density (PPFD) peaked at 1400 h in both the open and glasshouse conditions where PPFD for the former condition registered 50% higher PPFD than the glasshouse (1,215 vs. 606 μmol/m2/s; Fig. 1b). Similarly, the PPFD patterns follow the same trend as the diurnal pattern of temperature. The highest temperature recorded under both growing conditions peaked at 1400 h with open field and greenhouse conditions recording temperatures of 37 and 33°C, respectively (Fig. 1c).

Changes with time of the day in CO2 concentration (a), photosynthetic photon flux density (b), and temperature (c) of Labisia Pumila plants grown in the open field (open diamond with line) and greenhouse (closed square with line) growing conditions, n = 27
Results
Diurnal net photosynthesis
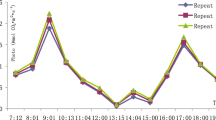
The different growing conditions had significant effects on the photosynthetic patterns of L. pumila. In greenhouse, the highest net photosynthesis (A) peak was achieved between 1200 and 1400 h (Fig. 2). For plant grown under open field, there were double peaks observed at 1000 and 1600 h in all the three varieties. Daily average A of open field plants were 4.10, 3.81, 3.51 μmol/m2/s and 5.05, 5.02, 4.71 μmol/m2/s in greenhouse for the varieties Alata, Pumila and Lanceolata, respectively. The plants daily A was comparatively higher under greenhouse condition compared to the open field by 29.3% (Table 1). Under both growing conditions var Lanceolata demonstrated significantly lower daily average as compared to the greenhouse (Fig. 2c), while A of var Alata recorded higher value (Fig. 2a) than var Pumila (Fig. 2b) although they were not significantly different (Table 1). It was also shown that var Alata and var Pumila acclimatized well under greenhouse condition where the interaction effects between varieties and growing conditions showed that var Alata and Pumila had the highest daily A when planted under greenhouse condition (Table 1).

Diurnal variations of net photosynthesis of three varieties of Labisia pumila viz var alata (a), var pumila (b), and var lanceolata (c) in the open field (open diamond with line) and greenhouse (closed square with line) growing conditions. Data are mean ± S.E., n = 9
Photosynthetic light response curves
Under greenhouse condition, all the three varieties recorded lower light saturation point than those grown in the open field (Table 1). In the latter condition, the light compensation point for var Lanceolata was found to be the highest (17.22 μmol/m2/s), and when treated with the greenhouse conditions the variety exhibited the lowest light compensation point compared to var Alata and var Pumila (8.22 vs. 9.12 and 9.72 μmol/m2/s, respectively). It was also observed that the light saturation point was highest in the open area compared to the glasshouse condition where there were no significant differences between all the three varieties. The A max measured in the open field condition for all the three varieties displayed lower values than those from the greenhouse with var Lanceolata exhibiting the lowest A max (6.01 μmol/m2/s). However, when exposed to greenhouse condition, var Alata registered a substantially (p < 0.05) higher A max (9.32 μmol/m2/s) than var Pumila (8.37 μmol/m2/s), followed by var Lanceolata (8.22 μmol/m2/s). Interaction effects between varieties and growing condition showed that Amax for var Alata and var Pumila was highest when planted under greenhouse condition (Table 1).
Chlorophyll fluorescence and chlorophyll content
The diurnal pattern of maximum photochemical efficiencies of PSII (F v/F m) was influenced by the growing conditions (p ≤ 0.01; Fig. 3). It was found that F v/F m ratio was higher in the greenhouse compared to the open field condition. For every variety, the F v/F m ratio ranged between 0.821 and 0.847 from morning to evening. For each variety, the F v/F m was almost similar in the morning and evening, and the values were close to 0.85. However, the lowest F v/F m ratio was observed around 1400 h under open field condition (0.626, 0.726, 0.526) for var. Alata, Pumila and Lanceolata, respectively. However, under greenhouse at 1400 h, the F v/F m ratio inside the greenhouse changed a little, had frail point and varied between 0.826 and 0.836. No statistical significance was observed in total chlorophyll content under different growth conditions, although chl b was higher from plants growing under greenhouse than open field that significantly showed lower chl a/chl b ratio in greenhouse than in open field (Table 2).

Diurnal variations of maximal photosystem II efficiency (F v/F m) of three varieties of Labisia pumila viz var alata (a), var pumila (b), and var lanceolata (c) in the open field (open diamond with line) and greenhouse (closed square with line) growing conditions. Data are mean ± S.E., n = 9
Discussions
The photosynthetic response of leaves to increased photon flux density (PPFD) depends on an intricate interaction between acclimation of photosynthetic machinery, photoinhibitory damage and repair (Adir et al. 2003; Pearcy and Sims 1994). The existence of the dual peaks of diurnal A curves exhibited by L. pumila plants exposed to open field condition might have resulted from photoinhibition under high PPFD condition between 800 and 1200 μmol/m2/s (Matos et al. 1998). The photosynthetic response of mature leaves to increased PPFD depends on the complex interactions between photoinhibitory repair, damage, acclimation and photoprotection of the photosynthetic machinery (Su and Liu 2005). It was also found that a decline in A for all the three varieties in open field condition occurred after 1000 h. The decrease might be due to an increase in PPFD that simultaneously escalated temperature outside the greenhouse (Fig. 1; Jaafar 2006). From the daily average of A (Table 1), each variety demonstrated similar acclimation trait in the open field as well as in the greenhouse. Var Alata and Pumila exhibited better adaptation in both open field and greenhouse growing conditions compared to var Lanceolata.
Labisia pumila seemed to be a shade adapted plant. In the present study, the Amax of L. pumila increased with simultaneous decrease in the compensation point and light saturation point when plants were exposed to greenhouse growing condition with low PPFD, suggesting that these plants were shade loving species (Lambers et al. 1998). According to Kitao et al. (2000) and Patakas et al. (2003), shade adapted plants have the ability to increase Amax and other energy dissipating mechanism when grown under low light condition compared to the high light condition. Var Alata and var Pumila were shown to acclimatize well under greenhouse condition. This is shown in Table 1, where interaction effects between growth conditions and varieties have shown that these varieties have the highest daily A and Amax under greenhouse condition, although there were no significant differences between their light compensation and light saturation point.
Chlorophyll fluorescence is frequently used to determine the state of energy distribution in the thylakoid membrane, photoinhibition and quantum efficiency of PSII (Maxwell and Johnson 2000; Mierowska et al. 2008). Under intense sunlight, photochemical impairment was reflected in either the increase in original fluorescence (F o), or in the decrease in maximum fluorescence (F m), or in the ratio of variable, F v (equals to F m − F o), to maximal fluorescence (F v/F m) (Thomas and Turner 2001). In the current study, the F v/F m ratio decreased as the PPFD increased under both growing conditions, with decrease in the open field being more acute (Fig. 3). The result signified that photoinhibition had occurred under high light state between 1200 and 1400 h reaching the peak at the latter hour regardless of plants’ growing conditions. In the open field, photoinhibition was related to the double peaked curves of diurnal A during daylight as a result of high PPFD (1200 μmol/m2/s). Subsequently, rapid recoveries of F v/F m were recorded from all the three varieties exposed to open field condition, which confirmed the presence of photo-defensive mechanism that might aid photosynthesis recuperation when favorable conditions are restored (Maxwell and Johnson 2000; Hernandez et al. 2004).
In the greenhouse condition, chl b was observed to be higher than chl a. Plants that have high chl b than a are generally able to accommodate to the low light situation (Lei et al. 1996). In this study, reduction in the ratio of chl a:b in the leaves of greenhouse plants with significant increase in chl b signified possible changes in the establishment of both light harvesting and electron transport components (Schiefthaler et al. 1997). The lowering of chl b could be an indication of chl destruction by excess irradiance under the open field condition (Jason et al. 2004). The result suggested that the high chl a to b ratio without significant changes in total chl content could be considered as a protecting mechanism due to the consequence of exposing plants to open field condition (Wang et al. 2007). It may be hypothesized that total chl content was high enough to enable the occurrence of high non-photochemical quenching (NPQ) of plants in open field and greenhouse conditions, and this could have facilitated efficient photo fortification of the photosynthetic apparatus against excess irradiation as established in the present study by the photosynthetic recovery observed after the 1400 h (Fig. 2; Thomas and Turner 2001; Patricia et al. 2001).
The current study had indicated that out of the three varieties investigated, var Alata and var Pumila displayed the highest ability to acclimatize to the greenhouse condition compared to var Lanceolata. Hence, they could be the best choice of selection for commercial planting under greenhouses. The approach of acclimation normally involves altered antenna size of photosystem II (Bailey et al. 2001), adjustment in reaction center stoichiometry (Walters and Horton 1994) and distorted levels of Rubisco (Seemann et al. 1987). Such acclimation is readily monitored through the consequent changes in the ratio of chl a to chl b, and in the light-saturated rate of photosynthesis (A max) that had also been established in the present experiment.
References
Adir N, Zer H, Ohad I (2003) Photoinhibition—a historical perspective. Photosynth Res 76:343–370
Bailey S, Walters RG, Jannsonn S, Horton P (2001) Acclimation of Arabidopsis thaliana to the light environment: the existence of separate low light and high light responses. Planta 213:794–801
Burkill IH (1966) A dictionary of the economic products of the Malay Peninsula, 2nd edn. Government of Malaysia and Singapore Publication, Kuala Lumpur
Demmig AB, Adam WW, Grace SC (1997) Physiology of light tolerance in plants. Hortic Rev 18:215–246
Ezumi MWH, Amrah SS, Suhaimi AWM, Mohsin SSJ (2006) Evaluation of the female reproductive toxicity of the aqueous extract of Labisia pumila var alata in rats. Ind J Pharmacol 38(5):355–356
Givnish TJ (1988) Adaptation to sun and shade: a whole-plant perspective. Aust J Plant Physiol 15:63–92
Hardwich K, Baker NR (1973) In vivo measurement of chlorophyll content of leaves. New Phytol 72:51–54
Hernandez JA, Rubio M, Olmos E, Ros BA, Martinez GP (2004) Oxidative stress induced by long term plum pox virus infection in peach (Prunus persica). Physiol Plant 122:486–496
Ibrahim MH, Jaafar HZE, Mohd Haniff H, Yusop R (2010) Changes in the growth and photosynthetic patterns of oil palm (Elaeis guineensis Jacq.) seedlings exposed to short term CO2 enrichment in a closed top chamber. Acta Physiol Plant 32(2):305–313. doi:10.1007/s11738-009-048-Y
Jaafar HZE (2006) Carbon dioxide enrichment technology for improved productivity under controlled environment system in the tropics. Acta Hortic 742:353–363
Jaafar HZ, Mohamed Haris NB, Rahmat A (2008) Accumulation and partitioning of total phenols in two varieties of Labisia pumila Benth. under manipulation of greenhouse irradiance. Acta Hortic 797:387–392
Jaafar HZE, Mohd Hafiz I, Philip E (2009) Leaf gas exchange properties of three varieties of Labisia pumila Benth. under greenhouse conditions. J Trop Plant Physiol 3:16–24
Jaafar HZE, Ibrahim MH, Por LS (2010) Effects of CO2 enrichment on accumulation of total phenols, flavonoid and chlorophyll content in two varieties of Labisia pumila Benth. exposed to different shade levels. In: Proceedings of international conference on balanced nutrient management for tropical agriculture, Kuantan, Pahang, Malaysia, 15–22 February 2010; UPM: Kuala Lumpur, Malaysia; pp 112–114
Jamia AJ, Ibrahim J, Khairana H, Juriyati J, Bukhari AB, Mohd CI, Kartini I (2003) In show directory: IPTA Research and Development Exposition, 9–12 October, Tun Razak Hall 1, PWTC, Kuala Lumpur
Jason JG, Thomas GR, Pharr DM (2004) Photosynthesis, chlorophyll fluorescence and carbohydrate content of Lilicium taxa grown under varied irradiance. J Am Soc Hortic Sci 129:486–495
Karpinski S, Reynolds H, Karpinska B (1997) Photosynthesis electron transport regulates the expression of cytosolic ascorbate peroxidase genes in Arabidopsis during excess light stress. Plant Cell 9:627–640
Kitao M, Lei TT, Koike T, Tobita H, Maruyama Y (2000) Susceptibility to photoinhibition of three deciduous broadleaf tree species with different successional traits raised under various light regimes. Plant Cell Environ 23:81–89
Kubiske ME, Pregitzer KS (1996) Effects of elevated CO2 and light availability on the photosynthetic response of trees of contrasting shade tolerance. Tree Physiol 16:351–358
Lambers H, Chapin FS, Pons TL (1998) Plant physiological ecology. Springer, New York
Lei TT, Tabuchi R, Kitao M, Koike T (1996) The functional relationship between chlorophyll content, leaf reflectance, and light capturing ability of Japanese forest species under natural shade and open light regimes. Physiol Plant 79:411–418
Matos MC, Matos AA, Mantas A, Cordeivero V, Viera JB (1998) Diurnal and seasonal changes in Prunus amygladus gas exchange. Photosynthetica 35:517–524
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence—a practical guide. J Exp Bot 51:659–668
Mierowska A, Keutgen N, Huysamer M, Smith V (2008) Photosynthetic acclimation of apple spur leaves to summer-pruning. Sci Hortic 102:9–27
Patakas A, Kofidis G, Bosabalis AM (2003) The relationship between CO2 transfer mesophyll resistance and photosynthetic efficiency in grapevine cultivars. Sci Hortic 97:255–263
Patricia M, Li XP, Krishna K (2001) Non-photochemical quenching a response to excess light energy. Plant Physiol 125:1558–1566
Pearcy RW, Sims DA (1994) Photosynthesis acclimation to changing light environment: scaling from leaf to whole plant. In: Caldweld MM, Pearcy RW (eds) Exploitation of environmental heterogeneity by plants: ecophysiological process above and below ground. Academic press, San Diego, pp 145–174
Philip E, Noor AY, Rizal MKM (2006) Growth and physiological evaluations of planted forest species along a recreational trail. J Trop Plant Physiol 1(1):57–65
Rozihawati Z, Aminah H, Lokman N (2003) Preliminary trials on the rooting ability of Labisia pumila cuttings. In: Malaysia science and technology congress 2003, Agricultural Sciences, Cititel Midvalley, Kuala Lumpur
Schiefthaler U, Russel W, Bolhar-Nordenkampf HR, Crithely C (1997) Photoregulation and photodamage in Schefflera arboricola leaves adapted to different light environment. Aust J Plant Physiol 26:485–494
Seemann JR, Sharkey TD, Wang JL, Osmond CB (1987) Environmental effects on photosynthesis, nitrogen-use efficiency, and metabolite pools in leaves of sun and shade plants. Plant Physiol 84:796–802
Stone BC (1988) Notes on the genus Labisia Lindyl (Myrsinaceae). Malayan Natl J 42:43–51
Su PX, Liu XM (2005) Photosynthetic characteristics of linze jujube in condition of high temperature and irridiation. Sci Hortic 104:339–350
Thomas DS, Turner DW (2001) Banana leaf gas exchange and chlorophyll fluorescence in response to soil drought, shading and lamina folding. Sci Hortic 90:218–228
Walters RG, Horton P (1994) Acclimation of Aradopsis thaliana to the light environment—changes in composition of the photosynthetic apparatus. Planta 195:248
Wang H, Wang F, Wang G, Mahourjat K (2007) The response of photosynthetic capacity, chlorophyll fluorescence and chlorophyll content of nectarine (Prunus persica var. Nectarina Maxim) to greenhouse and field grown conditions. Sci Hortic 112:66–72
Yoshimura K, Yabuta Y, Ishikawa Y (2000) Expression of spinach ascorbate peroxidase isoenzymes in response to oxidative stresses. Plant Physiol 123:223–233
Acknowledgments
The authors wish to acknowledge the financial support from the Ministry of Higher Education Malaysia and Research Management Center under the Research University Grant Scheme (Grant No.: 91007).
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by L. A. Kleczkowski.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Ibrahim, M.H., Jaafar, H.Z.E. Photosynthetic capacity, photochemical efficiency and chlorophyll content of three varieties of Labisia pumila Benth. exposed to open field and greenhouse growing conditions. Acta Physiol Plant 33, 2179–2185 (2011). https://doi.org/10.1007/s11738-011-0757-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11738-011-0757-1