
Abstract
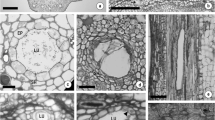
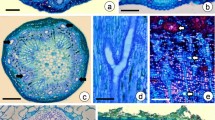
The structure, distribution and patterns of resin ducts in processes of its initiation and development were studied with the methods of thin section and ultrathin section. This paper emphasized the ultrastructural changes during canal development by a ring of the live epithelial cells, and the epithelial cells were usually surrounded with one or two layered sheath cells, which were normal parenchyma cells in some primary resin ducts and became dead cells with thick walls in other primary and secondary resin ducts. The resin ducts were found to occur in almost all organs, except in cotyledon. The resin ducts were formed by schizogeny and their development can be divided into three stages (e.g., initial stage, formation stage and mature stage). At the initial stage, the initial cells had many plastids without integral membrane structures, which contain one or two starch grains in them, and there are a few black osmiophilic droplets on the endoplasmic reticulum and membranes. A small number of osmiophilic droplets were present in the plastids. At the formation stage, the number of plastids, mitochondria and Golgi bodies in epithelial cells increased. The plastids were commonly surrounded by endoplasmic reticulum sheath. The larger osmiophilic droplets in cytoplasm and the smaller osmiophilic droplets on the plastids envelope, mitochondrion envelope and Golgi vesicles obviously increased in number during canal developing. At the mature stage, the cytoplasm of epithelial cells became thin with small nucleus. The number of mitochondria and Golgi body decreased, but numerous plastids still existed. Osmiophilic droplets were abundant in epithelial cells as in previous status. Taken together, the structures of plastids in epithelial cells gradually became well developed and the synthesis of resin was remarkably enhanced during resin duct formation and plastids should be the main site for resin synthesis.
Similar content being viewed by others

References
Berryman A A (1972). Resistance of conifers to invasion by bark beetle-fungus associations. BioScience, 22(10): 598–602
Boucher D, Lavall E R, Mauffette Y (2001). Biological performance of the white pine weevil in relation to the anatomy of the resin canal system of four different host species. Can J For Res, 31(11): 2035–2041
Chen T H, Ye Z H, Ge S (1985). Karyotype analysis in Pinus massoniana Lamb of different elevation populations. J Nanjing For Univ, 3: 132–148 (in Chinese)
Esau K (1965). Plant anatomy. 2nd ed. New York: John Wiley and Sons Press, 317–318
Fahn A (1979). Secretory tissues in plants. London: Academic Press, 185–209
Fang Y X, Liu J H, Chen Q (1983). Study of Pinus massoniana karyotype. Sci Silv Sin, 19(2): 212–216 (in Chinese)
Fang Y X, Lu Zhen (1990). Patterns of karyotype differentiation of different population of Pinus massoniana. Guihaia, 10(3): 201–207 (in Chinese)
Geng S L, Wu Y H, Zhao S, Wu H (2000). The initiation and development of resin ducts of the primary structure of stem in Pinus massoniana Lamb. J South China Agric Univ, 21(3): 44–47 (in Chinese)
Li A M, Wang Y R, Wu H (2004). Cytochemical localization of pectinase: the cytochemical evidence for resin ducts formed by schizogeny in Pinus massoniana. Acta Bot Sin, 46(4): 443–450
Ma X H (1988). Effects of rainfall on the nutrient cycling in man-made forests of Cunninghamia lanceolata and Pinus massoniana. For Res, 1(2): 123–131 (in Chinese)
Mo J M, Sandra B, Pang S L, Kong G H (2002). Role of understory plants on nutrient cycling of a restoring degraded pine forests in a MAB Reserve of Subtropical China. Acta Ecol Sin, 22(9): 1407–1413 (in Chinese)
National Cooperative Group of Pinus massoniana Provenance Tests (1986). The genetic variation of provenance and regional division of the seed origin of Pinus massoniana. Subtrop For Sci Technol, 2: 1–12 (in Chinese)
Phillips M A, Croteau R B (1999). Resin-based defenses in conifers. Trends Plant Sci, 4(5): 184–190
Qin G F, Zhou Z C, Wang P D, Jin G Q (1990). The genetic variation of natural population of Pinus massoniana. For Res, 3(2): 182–185 (in Chinese)
Song Z Q (2002). Fine chemical applications of pine resin (I)-the composition and properties of pine resin. J Chem Ind Fort Prod, 36(4): 29–33 (in Chinese)
Sun J J, Tang L Y, Tao L Q, Chen C C (2007). Research on Pinus massoniana geometry modeling based on morphological characters. Sci Silv Sin, 43(4): 71–76 (in Chinese)
Wu H (1990). The initiation and development of resin ducts in Pinus tabulaeformis Carr. J Wuhan Bot Res, 3(4): 311–316 (in Chinese)
Wu Z L (1963). Making great efforts on developing Pinus massoniana. For China, 63(1): 18–21 (in Chinese)
Xiang W H, Tian D L (2002). Nutrient cycling in Pinus massoniana stands of different age classes. Acta Phytoecol Sin, 26(1): 89–95 (in Chinese)
Xu J, Chen T H, Wang Z R (1998). Study on chromosome fluorescent banding pattern in Pinus massoniana. J Wuhan Bot Res, 16(2): 167–170 (in Chinese)
Xu J, Chen T H, Wang Z R (2001). Fluorescent banding pattern of the chromosomes of the 7 provenances of Pinus massoniana. J Nanjing For Univ (Nat Sci Ed), 25(5): 11–16 (in Chinese)
Xu L Y, Hu S Y (1986). Staining of epoxy thick sections. Chin Bull Bot, 4(1–2): 108–110 (in Chinese)
Yue S L, Rong W C (1994). Geographical variation of resin components between masson pine provenances. For Res, 7(4): 431–436 (in Chinese)
Zhao Y, Zhou Z C, Wu J F, Lan Y Z (2007). Growth response of superior provenance of masson pine to phosphorus supply and persistence of phosphorus effect. Sci Silv Sin, 43(10): 64–70 (in Chinese)
Zhou Z X (2001). Pinus massoniana in China. Beijing: China Forestry Publishing House, 14 (in Chinese)
Author information
Authors and Affiliations
Corresponding author
Additional information
__________
Translated from Scientia Silvae Sinicae, 2008, 44(9): 36–40 [译自: 林业科学]
About this article
Cite this article
Li, A., Wang, Y. & Wu, H. Initiation and development of resin ducts in the major organs of Pinus massoniana . Front. For. China 4, 501–507 (2009). https://doi.org/10.1007/s11461-009-0061-z
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11461-009-0061-z