
Abstract
As PM2.5 affect human health, it is important to target tree planting in the role of reducing air pollution concentrations. PM2.5 capture capability of greening trees is associated with leaf morphology, while quantitative research is scanty. In this paper, the PM2.5 capture capability of 25 species in Beijing and Chongqing were examined by a chamber device. Groove proportion, leaf hair, stomatal density, and stomata size were selected as indexes of leaf morphology. Results showed that groove proportion and stomata size significantly relate to PM2.5 capture quantity, while no significantly positive correlations were found for leaf hairs and stomatal density. Broadleaf species are conducive to PM2.5 capture for their rich leaf morphology and have an edge over coniferous in PM2.5 capture per leaf area. However, coniferous had a larger PM2.5 capture capability per tree due to the advantage of a large leaf area. Significant difference existed between the species in Beijing and Chongqing due to the different leaf morphology. Urban greening trees are diverse and the structures are complicated. Complex ecological environment may lead to different morphology characteristics. Climate and pollution conditions should be considered when greening.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Increasing PM2.5 concentrations have become the primary pollutants in many densely populated cities. In China, lots of cities are experiencing serious air pollution and bearing heavy burden of respiratory diseases (Shu et al. 2015; Fu et al. 2015). It has been documented that the incidence of lung cancer in Beijing was 1.055 for men and 1.149 for women due to 10 mg m−3 increase of PM2.5 concentration (Guo et al. 2014, 2016). Since 2001, the estimated average total mortality due to PM2.5 was approximately 5100 a year until 2012, and the unit capital mortality for all ages was around 15 in 10,000. Increasing PM2.5 concentration is the primary environmental problem, leading to an urgency of implementing air pollution abatement (Zheng et al. 2015).
Phytoremediation can clean air to a great extent depending on its capability of reducing speed velocity and capturing particles (Popek et al. 2015). The effectivity of trees to capture PM2.5 has been addressed a lot. Some authorities proposed tree planting as a dominant measure to alleviate airborne fine particulate matter. These plants commonly have a large surface area to filter PM out of the air by their removing or capturing capability derived from the leaf surface (Nowak et al. 2013; Chen et al. 2015). Understanding the PM2.5 capture capability of trees is crucial to assess the role of urban forest construction policies in reducing PM2.5 concentrations.
Many attempts have been made to quantify the effectiveness of urban trees in capturing PM2.5 (Dzierżanowski et al., 2011; Gromke and Ruck 2012; Sæbø et al. 2012; Speak et al. 2012; Popek et al. 2013). Although high effectiveness of trees has been demonstrated, significant differences between species were recorded. Both species and location to the pollution source are critical in determining the effectiveness (Pullman, 2009; Mori et al. 2015). Previous studies suggested that urban planting in the future should focus on the utility of conifers (Beckett et al. 1998; Beckett et al., 2000a, b). Species-specific features, such as leaf surface, leaf type, leaf area index, and leaf morphology, act as the main structures and are important factors affecting capture capability. Broadleaf species with rough leaf surfaces can capture more PM2.5 than those with smooth leaf surfaces (Nguyen et al. 2015), indicating that plant choices are important because proper or reasonably planned layout of them can maximize the efficiency in air pollution abatement (Fowler et al. 2004; Räsänen et al. 2013).
Pervious works qualitatively examined on the leaf roughness, leaf hair, stomatal density, and stomata size (Sæbø et al. 2012). However, a detailed quantitative research is necessary to analyze the influence of leaf morphology on PM capture capability. In this paper, the effectiveness of 25 tree species in capturing PM2.5 was examined using a chamber device. Tree leaves were sampled in growing season (e.g., from May to September), then dried and exposed to NaCl aerosol particles in the chamber device. Groove proportion, leaf hair, stomatal density, and stomata size were quantified using scanning electron microscopy (SEM). Capturing efficiencies were measured at controlled NaCl concentrations and duration and analyzed regarding leaf morphology.
Materials and methods
Study site and plant material
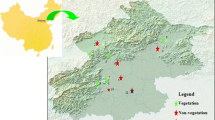
Fifteen species in Chongqing and 10 in Beijing were tested (Table 2). Beijing is located in the northwest of China and is a densely populated city with monsoon climate and poor air quality (39.54° N, 116.23° E) (Fig. 1). Previous documents revealed that PM2.5 concentrations in Beijing have increased in recent years (the annual PM2.5 concentration in 2014 was 85.9 μg m−3, data from Environmental Protection Administration, China). Chongqing (29.59° E, 106.54° N) has a relatively clean and humid subtropical monsoon climate. Local annual PM2.5 concentration in Chongqing 2014 was 62.8 μg m−3 (data from Environmental Protection Administration, China). In addition, there are 5 common species among the total 25 species including Platanus orientalis, Broussonetia papyrifera, Ginkgo biloba, Magnolia soulangeana, and Pinus massoniana. Besides, 20 species are broadleaf species, others are conifer.

Location of the two sampling sites
Sampling
All trees distributed along urban main roads where heavy traffic pollution dominate. Daily meteorological data were obtained by an automatic weather station at a height of 1.5 m above the ground in both cities. Temperature, humidity, wind speed, and precipitation were recorded. Meteorological data and PM2.5 concentrations (Environmental Protection Agency, China) during May to September 2014 were shown in Table 1.
Four trees of each species were sampled twice a month from May to September. These trees were grown well, with similar age, and a diameter at breast height (DBH). Note that the first year twigs can provide better information on pollution than perennial twigs. We sampled the first year twigs from four directions at three heights (low, middle, and high layer of canopy), which were at a 1.0–2.0-m height above ground level depending on the tree structure. Twelve twigs in each tree were sampled with weight ranging from 300 to 500 g, then, washed with 500 mL deionized water in the laboratory. Lastly, 12 sampled twigs of each tree were dried in an incubator and exposed to PM2.5 in a chamber device.
We assumed that all leaves of a tree are comparable to sampled ones and exposed to the same pollution concentration in field conditions. In order to investigate the PM2.5 capture capability of a tree, we multiplied the masses of PM2.5 per unit leaf area by total leaf area. Leaf area index (LAI) and vertical projection area of the crown (S t ) were applied to estimate total leaf area. A tape measure and DBH ruler were used in the field to measure the DBH, tree height (H), and crown diameter (C).
Chamber device
Particles gradually accumulate on the surfaces of tree leaves until there is a dynamic equilibrium between deposition and loss (Mitchell et al. 2010). Aerosol particles with controlled concentration and size can be generated by an aerosol generator. In this research, sodium chloride (NaCl) solution with a concentration of 0.1 mol L−1 was chosen as the PM2.5 source (Freer-Smith et al. 2004) because the component of PM2.5 in Beijing and Chongqing mainly consists of sulfate, nitrate, black carbon, and organic pollutants (Guo et al. 2014), while Cl− was less. Thus, there is no background interference (Beckett et al., 2000a, b; Freer-Smith et al. 2004). Pressure of aerosol generator was set to 25 psi, ensuring aerosol particle diameters were under 2.5 μm. PM2.5 concentration at downstream of the vacuum pump was measured 5 min at a time by DustMate (Turnkey Instruments, UK). Each test lasted about half an hour when PM2.5 concentrations remain stable. An average PM2.5 concentration of 550 μg m−3 was determined. After being exposed to PM2.5 in a sedimentation chamber (Fig. 2), twigs were washed by 500 mL deionized water and the filtrates were collected. Ion chromatograph (DionexICS-1600, USA) were used to detect the concentration of Cl−(mg mL−1).

Components of chamber device (Vacuum pump, Tai guan JP-40V; Aerosol generator, SH600; sedimentation chamber and air mixing chamber, a cube of side length is 400 mm, which consistent with Hwang et al. (2011), made from acrylic board. Twigs were placed in the center of the sedimentation chamber)
Statistics and classification of leaf morphology
Leaves were dried under air temperature of 80 °C in an oven. Two small square samples with a length of 5 mm and 1–2 cm to the central vein were cut from both leaf sides. Then, the samples were stick to the observation platform and plated with gold by ion-plating apparatus of Scan Electron Microscopy (SEM, HitachiS-3400 N, Japan). Afterward, we magnified them to 150–2000 times to observe the leaf morphology.
Sampled 25 species have distinctive grooves (Fig. 3a–d), such as stripe, net, corrugated, nodular, or verruca, where PM2.5 is often deposited. Proportion of groove (G) is expressed as follows:

Typical leaf morphology of the tested trees (a–d represents net, nodular or verrucous, stripe and corrugated groove, respectively. e–g embodies the leaf hair difference which represents hairless, sparse hair and hairy, respectively. h–k showed different stomatal type. h and i represent the stomatal that major semi-axis greater than 20 μm and less than 20 μm, respectively. j and k refer to the stomatal density greater than 100 mm−2 and less than 100 mm−2, respectively)
where A g (μm2) represents the projection area of groove, A t represents the area of the leaf sample (μm2).
Leaf hair per unit area (1 mm2) on both leaf sides was counted (Fig. 3e–g). It is noted that leaf hairs of most sampled trees are longer than 150 μm, while leaf hairs of some trees are shorter than 150 μm. A correction factor (c1 = 0.4) was introduced for the leaf hair shorter than 150 μm. The length of the major long axis was considered as the stoma size. Stoma number per unit leaf area was counted to calculate the stoma density (Fig. 3h–k).
Data analysis
Leaves were dried by airing after washing. For broadleaf species, leaf area was obtained by the leaf area meter (YMJ-B). For needle leaves, an equation was used (Li et al. 2001). Captured PM2.5 per unit area (APM 2.5 ) is expressed as:
where C i represents Cl − concentration in the filtrate and V represents the volume of the filtrate (500 mL). A i represents the leaf areas. Total leaf area(Y) is represented as (Yang. 2011):
where LAI represents leaf area index, S t represents vertical projection area of the crown, and C represents the average crown diameter. Then, captured PM2.5 per tree (T, mg) is calculated as follows:
Results
Leaf area per tree
Particle capture capability positively relate to the total leaf area per tree (Song et al. 2015). The difference in total leaf area per tree was examined by one-way ANOVA analysis and K-means clustering analysis. One-way ANOVA analysis showed that the total leaf area had significant difference among the 25 species (P < 0.01). Then, all the species were divided into four clusters (Fig. 4). The first cluster includes P. orientalis, G. biloba, Symplocos setchuensis, and Platycladus orientalis. Their leaf areas range from 21 to 37 m2. Species of the second cluster includes P. massoniana. Ficus microcarpa, Erythrina variegate, Magnolia soulangeana, and Pinus armandii, and have leaf areas ranging from 63 to 77 m2. The third cluster contains Cunninghamia lanceolata, Populus tomentosa and Fraxinus pennsylvanica, with leaf areas ranging from 87 to 102 m2. Others belong to the fourth cluster with a leaf area between 42 and 61 m2. We found that the total leaf area was superior for conifer (except P. orientalis) and lower for broadleaf species.

Cluster analysis of the total leaf areas per tree
Outcome of leaf morphology
Groove proportion ranges from 3 to 25 % and mostly between 10 and 20 %. There are about 50 % trees without leaf hair. Leaf hair of B. papyrifera, Litsea elongata in CQ, and B. papyrifera, P. tomentosa in BJ are 63.33 ± 11.60, 60.66 ± 7.14, 71.41 ± 9.28, 47.19 ± 9.41 mm−2, respectively, which is larger than others. P. orientalis in both cities and F. microcarpa in CQ have larger stomatal density and stomata size. Cinnamomum camphora in CQ and M. soulangeana in BJ have bigger stomata size.
APM2.5 at different classified levels is shown in Fig. 5. In total, APM2.5 increase as the groove proportion and leaf hair rise. The APM2.5 is less than 1 mg m−2 when groove proportion is lower than 10 %. However, it sharply increases to 3 mg m−2 when groove proportion exceeds 20 %. The APM2.5 at different classified levels of leaf hair shows that (1) species without leaf hair seems to capture the lowest APM2.5 (1.20 mg m−2), (2) significant increase in APM2.5 (1.85 mg m−2) when leaf hair ranges from 0 to 50 mm−2, (3) largest APM2.5 exists when leaf hair exceed 50 mm−2. In spite of the little increase of APM2.5 with stomata size and stomatal density, APM2.5 concentrate at a high level when stomata size and stomatal density exceed 20 μm, 100 mm−2, respectively.

APM2.5 at different classified level (The central rectangle spans the first quartile to the third quartile and the segment inside the rectangle shows the median, while the small squares represent the average. The triangle above and below or overlapping the dash show the maximum and minimum value)
Difference in capturing efficiency among the trees
After sedimentation, elution, and sample detection, captured PM2.5 per unit leaf area and per tree of each species were calculated (Table 2). A significant difference exists between species (P < 0.01). For the captured PM2.5 per unit leaf area, C. lanceolate, and Grevillea robusta in Chongqing had the highest PM2.5 capturing capability. However, G. biloba, Cinnamomum japonicum, and M. soulangeana shows the lowest PM2.5 capture capability. Captured PM2.5 of G. biloba is merely 10 % of C. lanceolata. Others range from 1.5 to 2.5 mg m−2. The results in Beijing indicate that the most efficient species are B. papyrifera and Sophora japonica. Besides, PM2.5 captured by conifer is lower than the actual, which can be explained by the fact that we ignored the wax layer and sticky secretions (Table 3).
For the captured PM2.5 per tree, C. lanceolata captures the largest PM2.5. G. robusta, P. massoniana, and Litsea elongate are efficient in Chongqing. B. papyrifera is the most inefficient species due to the lower leaf area. Others range from 14.00 to 123.00 mg per tree. In Beijing, P. tomentosa shows the highest PM2.5 capture capability, while P. massoniana, B. papyrifera, and P. armandii are the intermediate ones among the 25 tree species. G. biloba, P. orientalis, and M. soulangeana belong to the lowest species. B. papyrifera and P. massoniana have comparatively high PM2.5 capture capability. However, the same species in Beijing capture more PM2.5 than those in Chongqing.
All the leaf characteristics data were normalized using SPSS (vision 19.0) to obtain dimensionless data including captured PM2.5 per leaf area (ZM), groove proportion (ZGP), leaf hairs (ZLH), stomata size (ZSS), and stomatal density (ZSD). Results found that there were significantly positive correlation between ZM and ZGP, ZSS (p < 0.01). A significant correlation between ZM, ZLH, and ZSD were not found (p > 0.05).
Discussion
The PM2.5 capture capability of 25 species in Beijing and Chongqing were tested by a chamber device. We obtained PM2.5 capture capability per unit leaf area and per tree, which can be helpful for selecting proper vegetation in urban settings. We tested the same species in Chongqing and Beijing at the same environment (same pollution level, temperature, and humidity). Surprisingly, there was a difference between the two sites. It can be seen that the APM2.5 in Beijing were larger, possibly due to the rich leaf morphology.
Leaf features of coniferous species can cause high air turbulence inside the tree crowns, leading to an increase in the interception capacity of contaminants (Bunzl et al. 1989). In this study, conifers did not show a significant advantage to capture APM2.5 compared with broadleaf species. However, PM2.5 accumulation capacity of conifers was superior to most broadleaf species for larger leaf areas per tree.
Leaf morphology appears to be a dominant factor in particle deposition (Mitchell et al. 2010). In previous studies, leaf morphology was qualitatively analyzed, while few quantified results were made (Chai et al. 2002). More detailed classifications and quantification of leaf morphology need to be further refined. In our study, groove proportions were quantified to evaluate the roughness. Meanwhile, leaf hair and stomatal density were quantified by counting them in a fixed leaf area. In addition, some studies revealed that particulate matter can get into leaves through the stomata, where fine particulates often crowded (Song et al. 2015; Lehndorff et al. 2006). Therefore, stomata size was also quantified. Our main results reveal that grooves are the main parts of a blade that capture PM2.5. A strong correlation between the PM2.5 accumulation and groove proportion proves that leaf surface roughness is a facilitator for PM2.5 capture (Fig. 6a). In addition, stomata size is an important influence factor for PM2.5 capture capability.

Correlation analysis of normalized leaf morphology and captured PM2.5 per unit leaf area (yb represents the species in Beijing; yc represents the species in Chongqing; yt represents the species in both Chongqing and Beijing. Each colored small square represents a tree species)
In this research, NaCl was used as a PM2.5 source, thus the mass of NaCl is lower than the natural PM2.5 source. The mass of captured PM2.5 per tree in our study was lower than the results in Song et al. (2015). Overall, the method used in this study is suitable for comparing the difference between species qualitatively merely.
Leaf characteristic and leaf morphology
Though all sampled species can capture PM2.5 and leaf surfaces have a considerable capacity (Wang et al. 2015), the amount differ significantly depending on leaf morphology. B. papyrifera is the most hairy for both lower and upper sides of the leaf and has the high groove proportion. The stomata size and stomatal density of B. papyrifera are not counted because leaf hair covered the stoma on leaf surface. Koelreuteria bipinnat has the highest groove proportion and stomata size, while the leaf hair and stomatal density of K. bipinnat are low. All of them do not show the highest PM2.5 capture capability. G. robusta is the most efficient broadleaf species and C. lanceolate is the most efficient conifer with high groove proportion and low stomata size. Species with high groove proportion and low stomata size is most effective at capturing PM2.5. Earlier studies also reported that mounts of PM2.5 captured on rough tree leaves with low stomatal density were high (Hwang et al. 2011; Räsänen et al. 2013).
Nguyen et al. (2015) found that trees with leaf hairs have high PM2.5 capture capability. Species with densely haired leaves were most effective at capturing PM (Dzierżanowski et al., 2011; Weber et al. 2014). However, we found no significant correlation between PM2.5 capture capability and leaf hair (Fig. 6d). This may attribute to differed methodological approaches and limited tree species in this research. A large number of tree species need to be studied in the future.
The role of stoma activity in particle deposition is ambiguous. On one hand, transpiration of water through stomata cools the surface which is conducive to attracting PM2.5; on the other hand, transpired water repels PM2.5 due to diffusiophoresis (Hinds 1999). No statistically significant correlations are found between PM2.5 capture capability and stomata size (Fig. 6b), stomatal density (Fig. 6c) in Chongqing and Beijing. For the stomata size, significant correlation with PM2.5 accumulation exists when tree species are classified into two groups according to stoma size:(1) The bigger one: PO, MS, CC, FM, KB, SS, PT, FP, which average stomata size are 111.19 μm. (2) The smaller one: CJ, BP, GB, FS, GR, LE, CL, PM, PA, EV, SJ, PLO, which average stomata size are 44.41 μm (Fig. 7). It can be seen that the correlation of the smaller one is higher than the bigger one, which can be explained by the restrain effect due to small stomata size. When the stomata size grow, the restrain effect become smaller.

Correlation analysis of normalized stomata size and captured PM2.5 per unit leaf area (y1: CJ, BP, GB, FS, GR, LE, CL, PM, PA, EV, SJ, PLO; y2: PO, MS, CC, FM, KB, SS, PT, FP)
Difference between coniferous and broadleaf species
Conifers shows the highest particle capture efficiency of tested tree species, which coincides with previous studies. Among the conifers, pines captured significantly more PM2.5 than cypresses (Beckett et al., 2000a, b). In this study, C. lanceolata is the most effective species in PM2.5 accumulation. Nevertheless, P. orientalis belongs to cypresses and has the least efficiency of PM2.5 accumulation, which coincides with the results of Song et al. (2015). It may be due to that pine trees deposited more PM2.5 than cypress ones.
More complex structure of the foliage of the conifers explained their greater effectiveness at capturing particles (Beckett et al., 2000a, b). However, in this study, conifers did not show a significant advantage to capture APM2.5 comparing with broadleaf species (Fig. 8a), which may attribute to that the structure of the conifer crowns were not considered. The total leaf area per tree of the conifer is higher than broadleaf species. Therefore, PM2.5 accumulation capacity per tree of conifers are superior to most of broadleaf species (Fig. 8b).

Captured PM2.5 of different species in Beijing and Chongqing
In addition, some trees are better able to survive in smoky and polluted conditions due to differences in physiological mechanisms of varied species. All in all, the best choices for pollution-control plantings are coniferous and broadleaved species with rough leaf surfaces and high adaptability (Beckett et al. 1998, 2000a, b; Silli et al. 2015).
Difference between two sites
A number of studies have demonstrated the effects of pollution on tree leaves. Deposition of PM was responsible for the change on leaf surface morphology (Gupta et al. 2015). Furthermore, it was found that the effects of PM2.5 on leaves relate to their acidity, salinity, and trace metal content properties (Grantz et al. 2003). Leaf density and thickness are altered when exposed to pollution environment and higher levels of NOX (Jochner et al. 2015). Pääkkönen et al. (1997) found that higher stomatal density and thicker leaves result in a greater tolerance to pollution. In addition, it was likely that PM2.5 might have an indirect effect via altering soil chemistry, which is also believed to be the major effect of PM on trees (Grantz et al. 2003). Trees strengthened the characteristics of their leaf structures under polluted conditions, which are regarded as adaptive and compensative to the adverse effects of air pollution (Chaturvedi et al. 2013). Studies also showed that trees develop different morphologies under polluted conditions (Karenlampi 1986; Veselkin 2004).
In addition, the chemical composition and wax structure may also be different in Beijing and Chongqing, which are significant for PM2.5 capture (Burkhardt 2010). Therefore, it is worthy of further study regarding the effect of PM2.5 pollutions on leaf morphology, including chemical composition, wax structure, groove proportion, leaf hair, stomatal density, and stomata size.
Summary and conclusion
This study revealed that broadleaf species with rich leaf morphology, namely, leaf groove, leaf hair and stomata, can capture more PM2.5 per leaf area than coniferous. However, coniferous captured larger PM2.5 per tree due to their large leaf area per tree. Among coniferous, C. lanceolata, P. orientalis, P. armandii were most efficient in capturing PM2.5. G. robusta, Erythrina variegata, K. bipinnata, P. tomentosa, F. pennsylvanica showed relatively high PM2.5 accumulations. A difference exists between the trees in Beijing and Chongqing due to the environment and leaf characteristic difference. Groove proportion and stoma size positively relate to PM2.5 accumulations, while there is no significant correlation between PM2.5 capture and stomatal density and leaf hair. Efficiency of PM2.5 capture capability in this study was examined by gas chamber, which is worthy of further analysis in field conditions. Thus, climate conditions, urban planning and management, and advantage tree species should be considered when greening. Urban greening trees are diverse and the structures are complicated. Complex ecological environment may lead to different physiological characteristics. Hence, it is valuable to examine the PM2.5 accumulation capability in specific ecological environment and different growth stages of trees. In addition, physiological indicators such as leaf photosynthetic rate, transpiration rate and stomatal conductance should be considered.
References
Beckett KP, Freer-Smith PH, Taylor G (1998) Urban woodlands: their role in reducing the effects of particulate pollution. Environ Pollut 99:347–360
Beckett KP, Freer Smith PH, Taylor G (2000a) Particulate pollution capture by urban trees: effect of species and wind speed. Glob Chang Biol 6:995–1003
Beckett KP, Freer Smith PH, Taylor G (2000b) Effective tree species for local air quality management. J Arboric 26:12–19
Bunzl K, Schimmack W, Kreutzer K, Schierl R (1989) Interception and retention of Chernobyl-derived 134 Cs, 137 Cs and 106 Ru in a spruce stand. Sci Total Environ 78:77–87
Burkhardt J (2010) Hygroscopic particles on leaves: nutrients or desiccants? Ecol Monogr 80:369–399
Chai YX, Zhu N, Han HJ (2002) Dust removal effect of urban tree species in Harbin. J Appl Eco l13:1121–1126
Chaturvedi RK, Prasad S, Rana S, Obaidullah SM, Pandey V, Singh H (2013) Effect of dust load on the leaf attributes of the tree species growing along the roadside. Environ Monit Assess 185:383–391
Chen X, Zhou Z, Teng M, Wang P, Zhou L (2015) Accumulation of three different sizes of particulate matter on plant leaf surfaces: effect on leaf traits. Arch Biol Sci 00:102–102
Dzierżanowski K, Popek R, Gawrońska H, Sæbø A, Gawroński SW (2011) Deposition of particulate matter of different size fractions on leaf surfaces and in waxes of urban forest species. Int J Phytoremediat 13(10):1037–1046
Fowler D, Skiba U, Nemitz E, Choubedar F, Branford D, Donovan R, Rowland P (2004) Measuring aerosol and heavy metal deposition on urban woodland and grass using inventories of 210Pb and metal concentrations in soil. Water Air Soil Pollut 4:483–499
Freer-Smith PH, El-Khatib AA, Taylor G (2004) Capture of particulate pollution by trees: a comparison of species typical of semi-arid areas (Ficus nitida and Eucalyptus globulus) with European and North American species. Water Air Soil Pollut 155:173–187
Fu J, Jiang D, Lin G, Liu K, Wang Q (2015) An ecological analysis of PM2.5 concentrations and lung cancer mortality rates in China. BMJ Open 5:e009452
Grantz DA, Garner JHB, Johnson DW (2003) Ecological effects of particulate matter. Environ Int 29:213–239
Gromke C, Ruck B (2012) Pollutant concentrations in street canyons of different aspect ratio with avenues of trees for various wind directions. Bound-lay meteoro l144:41–64
Guo S, Hu M, Zamora ML, Peng J, Shang D, Zheng J, Molina MJ (2014) Elucidating severe urban haze formation in China. Proc Natl Acad Sci 111:17373–17378
Guo Y, Zeng H, Zheng R, Li S, Barnett AG, Zhang S, Williams G (2016) The association between lung cancer incidence and ambient air pollution in China: a spatiotemporal analysis. Environ Res 144:60–65
Gupta GP, Kumar B, Singh S, Kulshrestha UC (2015) Urban climate and its effect on biochemical and morphological characteristics of Arjun (Terminalia arjuna) plant in National Capital Region Delhi. Chem Eco l31:524–538
Hinds WC (1999) Aerosol technology: properties, behavior, and measurement of airborne particles. Wiley, New York.
Hwang HJ, Yook SJ, Ahn KH (2011) Experimental investigation of submicron and ultrafine soot particle removal by tree leaves. Atmos Environ 45:6987–6994
Jochner S, Markevych I, Beck I, Traidl-Hoffmann C, Heinrich J, Menzel A (2015) The effects of short- and long-term air pollutants on plant phenology and leaf characteristics. Environ Pollut 206:382–389
Karenlampi L (1986) Relationships between macroscopic symptoms of injury and cell structural changes in needles of ponderosa pine exposed to air pollution in California. Ann Bot Fenn 23:255–264
Lehndorff E, Urbat M, Schwark L (2006) Accumulation histories of magnetic particles on pine needles as function of air quality. Atmos Environ 40:7082–7096
Li J, Zhou P, Zhao L (2001) Influence of drought stress on transpiring water-consumption of seedlings. Acta Ecol Sin 22:1380–1386
Mitchell R, Maher BA, Kinnersley R (2010) Rates of particulate pollution deposition onto leaf surfaces: temporal and inter-species magnetic analyses. Environ Pollut 158:1472–1478
Mori J, Hanslin HM, Burchi G, Sæbø A (2015) Particulate matter and element accumulation on coniferous trees at different distances from a highway. Urban For Urban Gree 14:170–177
Nguyen T, Yu X, Zhang Z, Liu M, Liu X (2015) Relationship between types of urban forest and PM2.5 capture at three growth stages of leaves. J Environ Sci-China 27:33–41
Nowak DJ, Hirabayashi S, Bodine A, Hoehn R (2013) Modeled PM2.5 removal by trees in ten US cities and associated health effects. Environ Pollut 78:395–402
Pääkkönen E, Holopainen T, Kärenlampi L (1997) Variation in ozone sensitivity among clones of Betula pendula and Betula pubescens. Environ Pollut 95:37–44
Popek R, Gawrońska H, Wrochna M, Gawroński SW, Sæbø A (2013) Particulate matter on foliage of 13 woody species: deposition on surfaces and phytostabilisation in waxes—a 3-year study. Int J Phytoremediat 15:245–256
Popek R, Gawrońska H, Gawroński SW (2015) The level of particulate matter on foliage depends on the distance from the source of emission. Int J Phytoremediat 17:1262–1268
Pullman MR (2009) Conifer PM2.5 deposition and re-suspension in wind and rain events. Cornell University
Räsänen JV, Holopainen T, Joutsensaari J, Ndam C, Pasanen P, Rinnan Å, Kivimäenpää M (2013) Effects of species-specific leaf characteristics and reduced water availability on fine particle capture efficiency of trees. Environ Pollut 183:64–70
Sæbø A, Popek R, Nawrot B, Hanslin HM, Gawronska H, Gawronski SW (2012) Plant species differences in particulate matter accumulation on leaf surfaces. Sci Total Environ 427:347–354
Shu Y, Zhu L, Yuan F, Kong X, Huang T, Cai YD (2015) Analysis of the relationship between PM2.5 and lung cancer based on protein-protein interactions. Comb Chem high T Scr 19:1–9
Silli V, Salvatori E, Manes F (2015) Removal of airborne particulate matter by vegetation in an urban park in the city of Rome (Italy): an ecosystem services perspective. Annali di Botanica 5:53–62
Song Y, Maher BA, Li F, Wang X, Sun X, Zhang H (2015) Particulate matter deposited on leaf of five evergreen species in Beijing, China: source identification and size distribution. Atmos Environ 105:53–60
Speak AF, Rothwell JJ, Lindley SJ, Smith CL (2012) Urban particulate pollution reduction by four species of green roof vegetation in a UK city. Atmos Environ 61:283–293
Veselkin D (2004) Anatomical structure of Ectomycorrhiza in Ledeb. And Ledeb. Under conditions of Forest ecosystems polluted with emissions from copper-smelting works. Russ J Ecol 2:71–78
Wang L, Gong H, Liao W, Wang Z (2015) Accumulation of particles on the surface of leaves during leaf expansion. Sci Total Environ 532:420–434
Weber F, Kowarik I, Säumel I (2014) Herbaceous plants as filters: immobilization of particulates along urban street corridors. Environ Pollut 186:234–240
Yanglili (2011) Research on the Related indicators of Green Quantity for 8 kinds of Landscape Plants. Southwest university
Zheng S, Pozzer A, Cao CX, Lelieveld J (2015) Long-term (2001–2012) concentrations of fine particulate matter (PM2.5) and the impact on human health in Beijing, China. Atmos Chem Phys 15:5715–5725
Acknowledgments
The research is supported by Forestry Public Welfare Project of China (Grant No. 201304301), Fundamental Research Funds for the Central Universities (Grant Nos. 201304301 BLX2014-12, 2015ZCQ-SB-01, and 2016ZCQ06).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Philippe Garrigues
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Liang, D., Ma, C., Wang, Yq. et al. Quantifying PM2.5 capture capability of greening trees based on leaf factors analyzing. Environ Sci Pollut Res 23, 21176–21186 (2016). https://doi.org/10.1007/s11356-016-7687-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-016-7687-9