
Abstract
Ionotropic P2X and metabotropic P2Y purinergic receptors are expressed in the central nervous system and participate in the synaptic process particularly associated with acetylcholine, GABA, and glutamate neurotransmission. As a result of activation, the P2 receptors promote the elevation of free intracellular calcium concentration as the main signaling pathway. Purinergic signaling is present in early stages of embryogenesis and is involved in processes of cell proliferation, migration, and differentiation. The use of new techniques such as knockout animals, in vitro models of neuronal differentiation, antisense oligonucleotides to induce downregulation of purinergic receptor gene expression, and the development of selective inhibitors for purinergic receptor subtypes contribute to the comprehension of the role of purinergic signaling during neurogenesis. In this review, we shall discuss the participation of purinergic receptors in developmental processes and in brain physiology, including neuron-glia interactions and pathophysiology.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
During the last two decades, evidence for the participation of ATP as neurotransmitter in neuronal signaling was collected by Drs. Surprenant [1] and Silinsky [2]. Purine-sensitive receptors were first classified as P1 G-coupled receptors which are activated by adenosine and P2 receptors, responding to stimulation of ATP [3]. Based on receptor cloning and studying of receptor-induced signal transduction, P2 receptors were divided into P2X receptors as ATP-gated ion channels and P2Y G protein-coupled receptors [4].
The expression of purinergic receptors has been identified during development and differentiation processes [5–10]. Nucleotides exert a synergic effect on cell proliferation in association with growth factors, chemokines, or cytokines in early stages of development [11–13] by parallel activation of the MAP kinase pathway and/or by transactivation of growth factor receptors [14, 15].
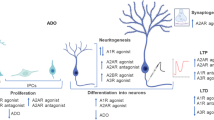
The complete role of ATP action in developmental processes still needs to be elucidated. It is known that ATP activates purinergic receptors resulting in many cases in increases of intracellular free calcium concentration [Ca2+]i. Changes in [Ca2+]i are involved in several events of differentiation and the embryogenesis process [16, 17]. Spitzer et al. [18] showed that naturally occurring patterns of Ca2+ transients encoded neuronal differentiation. Distinct frequency patterns of [Ca2+]i elevations were sufficient to promote neuronal differentiation, including physiological neurotransmitter receptors expression [19]. ATP and UTP are the main purinergic agonists activating P2X or P2Y receptors. These nucleotides can be rapidly degraded in the extracellular space by ectoenzymes to ADP or UDP, subsequently activating distinct P2Y receptors, or be finally degraded to adenosine, which is known to induce physiological responses via activation of P1 G protein-coupled receptors [20] (Fig. 1).

Purine-induced signaling pathway involves the activation of P1 adenosine and P2 purinergic receptors and purine hydrolysis by ectonucleotidases. The scheme demonstrates purinergic receptor activity present in glia-glia, neuron-glia, and neuron-neuron interaction during neurogenesis as well as in the metabolism of the adult brain
In this review article, we shall discuss the roles of purinergic signaling in neurogenesis such as cell cycle control during neural progenitor proliferation and differentiation as well as in maintaining physiology of neurons and glial cells and the involvement of purinergic receptors in pathophysiology. In addition, we shall outline state-of-the-art approaches used in investigation of P2 receptor function in physiological processes such as the use of antisense oligonucleotides, generation of knockout animals, and identification of new purinergic receptor subtype-selective drugs.
Study of purinergic receptor function during in vitro differentiation
During the development of the mammalian nervous system, neural stem cells and their derivative progenitor cells generate neurons by asymmetric and symmetric divisions [21]. P2 receptors were shown to be one of the first functionally active membrane receptors in chick embryo cells during gastrulation, in which ATP caused rapid accumulation of inositol triphosphate and Ca2+ mobilization in a similar way as acetylcholine (Ach) did via activation of muscarinic acetylcholine receptors, whereas other endocrine-acting substances such as insulin and noradrenaline (NA) induced much weaker effects in terms of intracellular calcium signaling [22, 23]. The induction of transient fluctuation in [Ca2+]i also denominated as calcium wave signaling allows for a coupling of spatial and temporal information. Thus, calcium waves have been proposed to play a role in mapping of neuronal networks [24] and to modulate neurogenesis during embryonic cortical development [25].
Neurotransmitters are prominent candidates for transcellular signals that could influence the development of embryonic neurons as they surround neural cells throughout brain development [26–29]. In addition, functional ligand-gated ionic channel receptors have been identified in neural progenitor cells prior to establishing cortical and subcortical synapses [30, 31]. In this context, the extracellular signaling mechanisms controlling the various transition steps involved in adult neurogenesis are still poorly understood. One approach used to identify the function of P2 receptors during development and differentiation is the use of in vitro models for neuronal and glial differentiation such as embryonic and adult neural progenitor cells (NPC), also known as neural stem cells (NSC), embryonic stem (ES), and embryonal carcinoma (EC) cells.
ES cells are obtained from the inner mass cell of the blastocyst. The differentiation of these cells closely resembles the in vivo process and, therefore, provides stable models for embryonic growth and development [32, 33]. ATP promotes cell proliferation acting through P2X3, P2X4, P2Y1, and P2Y2 receptors in murine ES cells [34]. Tissue-nonspecific alkaline phosphatase (TNAP) was also detected in these cells and used as a marker for their undifferentiated stage [35].
The neuronal differentiation of EC cells, originated from irradiated embryo cells [36], also resembles early neuronal development in vivo. P19 mouse EC cells express stem cell-specific marker proteins and their phenotypic changes in specific differentiation stages are similar to those of stem cells [37]. Recently, our laboratory [38] has determined gene and protein expression of P2 receptor subtypes throughout in vitro neuronal differentiation of P19 cells as well as in the undifferentiated cell stage suggesting the participation of purinergic signaling in initiating and directing differentiation. Differential expression and activity of P2Y1, P2Y2, P2Y4, P2X2 subtypes and P2X6 subunits were reported during neuronal maturation of P19 cells [38, 39]. As direct evidence for participation of purinergic receptors in neuronal differentiation, the presence of the antagonists pyridoxalphosphate-6-azophenyl-2′,4′-disulfonic acid (PPADS), reactive blue 2, or suramin during differentiation of P19 neural progenitor cells (NPC) to P19 neurons resulted in reduced activity of cholinergic and glutamate NMDA receptors in differentiated P19 cells, pointing at a participation of P2Y1, P2Y2, and P2X2 receptors.
Other in vitro neuronal and glial differentiation models used to understand the purinergic signaling are neural stem cells or progenitor cells which are isolated from the subventricular region (SVZ) located in the lateral ventricles (type B cells) or in the subgranular region of the gyrus dentatus of the hippocampus (residual radial glia) or even from the subcortical parenchyma of the cerebral cortex of embryonic and adult brain [40–42]. These regions in the adult brain act as neural stem cell reservoirs. These cells are already advanced in their differentiation stage when compared to ES or EC cells. Since NSC and NPC are capable of differentiating in both functional neurons and glial cells, they possess potential therapeutic applications such as ES cells in regeneration therapy following neuronal loss.
These NPC differentiate into olfactory, cerebellar, and retinal neurons [40] in the presence of growth factors, neurotransmitters, vasoactive peptides in vivo [43], and growth factors such as epidermal growth factor (EGF), fibroblast growth factor 2 (FGF-2), and leukemia inhibitory factor (LIF) in vitro. When exposed to a high concentration of FGF-2 in suspension, proliferating NPC form tridimensional cell aggregates denominated as neurospheres, which following induction to differentiation express neuronal marker proteins such as β-III-tubulin, microtubule-associated protein-2 (MAP-2), and synaptophysin [44] and express P2X3 and P2X7 receptors which may contribute to early [Ca2+]i transients as prerequisites for further differentiation [41]. Shukla et al. [45] identified functional P2 receptors in adult mouse hippocampal progenitors in situ and the nucleoside triphosphate-hydrolyzing ectoenzyme (NTPDase) in type B cells of the SVZ [46] and in hippocampal progenitor cells. In adult murine NPC of SVZ, P2Y1 receptor activity mainly contributes to [Ca2+]i transients with some participation of P2Y2 receptors. The presence of the specific P2Y1 receptor antagonist MRS 2179 resulted in diminished cell proliferation in neurospheres due to reductions of [Ca2+]i transients. Similar results were obtained with NPC from SVZ of P2Y1 receptor knockout mice [47]. P2Y1 receptor-deficient mice are viable; however, they have deficits in platelet aggregation [48]. It is suggested that the purine signaling underlies autocrine or paracrine mechanisms and P2Y1 and P2Y2 receptors are important for NPC differentiation [47]. These models are useful tools to study the roles of P2 receptor signaling in early stages of development and differentiation. The importance of ATP release and purinergic signaling has not only been demonstrated in developmental progenitor cell expansion and neurogenesis, but also in persistent progenitor cells of the adult brain [49].
Expression of purinergic receptors during development of the central nervous system
Purinergic signaling pathways are also involved in embryonic neurogenesis in much the same way as already discussed for in vitro differentiation models. ATP mediates elevation of [Ca2+]i and proliferation of immortalized human stem cells from embryonic telencephalon and mouse embryonic neurospheres [50, 51]. Ca2+ waves through radial glial cells in slices of the embryonic rat ventricular zone are mediated by P2Y1 receptors. Disrupting Ca2+ waves between embryonic NPC reduced ventricular zone cell proliferation during the peak of embryonic neurogenesis [25].
ATP directly contributes to modulate network-driven giant depolarizing potentials in the rat hippocampus during early stages of postnatal development [52]. In the developing hippocampal system a trophic role of ATP and the involvement of P2 receptor subtypes in shaping interneuronal connections during neuronal differentiation have been suggested [53]. Alterations of the regulation of embryonic growth by purinergic receptors might be involved in the onset of morphological malformations [54]. During rat postnatal development ectonucleotidase activity in the cerebral cortex steadily increases, reaching maximum values at 21 days of age [55]. Several P2Y and P2X receptors were shown to be dynamically expressed in the pre- and postnatal central and peripheral nervous system [56–59]. ATP inhibited motor axon outgrowth during early embryonic neurogenesis, most likely through the P2X3 receptor, and it was speculated that P2X7 receptors might be involved in programmed cell death during embryogenesis [58].
From all of the studied P2X receptors, homomeric P2X2 receptors were the first expressed in the rat central nervous system (CNS) on embryonic day 14 (E14) [56]. On E14, heteromeric receptors were formed by P2X2/3 receptor subunits. P2X3 receptor immunoreactivity was detected in cranial motor neurons as early as on E11, when neurons exited the cell cycle and started axon outgrowth, as well as postnatally on days 7 and 14 (P7 and P14) [56, 60]. Moreover, expression of P2X3-containing heteromeric receptors and other subunits was developmentally regulated in nucleus ambiguous motoneurons [61]. From E14 onwards P2X7 receptors were also expressed in the embryonic brain. For instance, in primary cultures of human fetal astrocytes basal levels of P2X7 receptor mRNA transcription and protein expression were detected [62]. Sperlágh et al. [63] have demonstrated that ATP regulates glutamate release via activation of P2X7 receptors. P2X7 receptor-induced excessive glutamate release alters Ca2+ homeostasis, subsequently resulting in activation of the apoptosis-related caspase cascade [64].
P2X receptor expression was downregulated in Purkinje cells and deep cerebellar nuclei at P21 and P66 rat embryonic stages, with the exception of P2X5 receptors whose immunoreactivity in granular cells was increased [65]. Evidence for participation of P2X receptors in different developmental processes such as neurite outgrowth (involving P2X3 receptors), postnatal neurogenesis (related to P2X4 and P2X5 receptor expression), and cell death (possibly involving P2X7 receptors) was collected. However, P2X1 and P2X6 receptor subunits may not play a role in neuronal development [58].
Neocortical neurons from 2-week-old rats possess a quite elaborated purine-triggered signaling system which includes both P2Y and P2X receptor activation [66]. Weissman et al. [25] showed that [Ca2+]i waves and subsequent ATP release, with consequent P2Y1 receptor activation, accompanied radial glial cell-derived neurogenesis in cultured slices of the developing rat forebrain, as mentioned above. Moreover, the importance of calcium signaling for differentiation of NPC has been studied [67, 68], and direct evidence for the participation of P2Y1 receptor-activated pathways in the early development has been provided by Scemes et al. [69]. P2Y receptors (particularly the P2Y1 subtype) were widely expressed in the embryonic rat brain as early as on E11 [57]. There was a marked decrease in the concentration of mRNA coding for P2Y1 receptors and upregulation of mRNA transcription coding for P2Y2 receptors in freshly isolated astrocytes of developing rat hippocampus [57].
Functional interactions between neurons and glia: a physiological overview
An increasing amount of evidence, initiated by the neuron-glia unit idea proposed by Hyden [70], indicates that glial cells, once referred to as a simple support portion in the CNS, are now considered indispensable functional partners of neurons [71], both in physiological and pathological conditions. However, many questions remain unanswered: (1) how glia detects and interacts with neural function; (2) does neuron-glia signaling play a significant role in synaptic transmission and plasticity; and (3) how glial cells can communicate with other glial cells.
Another important subject related to the interaction between glia and neurons emerges in neurogenesis. There is now a general agreement that the adult mammalian nervous system possesses many characteristics of astrocytes. The importance of glia in neuronal development was confirmed in a recent study showing that the number of GFAP (glial fibrillary acidic protein)-containing cells was reduced following transgenic targeting of adult mouse subependymal and subgranular zones, resulting in an almost complete loss of neurogenesis [72, 73]. In addition to assisting migration of neurons to their correct position and managing neurite outgrowth to their final communication targets [74, 75], glial cells have become an essential key for understanding neuronal differentiation by promoting initial stem cell proliferation and instructing undifferentiated cells to adopt a neuronal fate [76, 77].
In the mature brain, the proximity of astrocytes to neuronal synapses or to the blood-brain barrier makes these cells appropriate to control water diffusion and ion concentration in extracellular spaces [71, 78]. In particular, astrocytes regulate homeostatic environment and neurotransmitter levels by functional syncytium, in which gap junctions and specific membrane carriers play an important role [79–81]. In addition, glial cells produce and release a vast number of neurotrophins, including fibroblast growth factor, nerve growth factor, and transforming growth factor, which directly interfere in neuron physiology and coordinate developmental processes [71, 82–85].
ATP release and degradation, connecting adenosinergic and purinergic systems
As already mentioned, it is well documented that glial cells may directly alter neuronal activity by releasing neurotrophins and consequently modulating neurotransmitter release in the synapse [86, 87]. One of the main mechanisms connecting the neuron-glia system is believed to be mediated by the release of glutamate from glial cells [88, 89]. In this context, growing evidence indicates that purinergic receptor ligands are widely involved in the cell-cell signaling mechanism by acting as neurotransmitters or neuromodulators released by glial cells to control synaptic transmission in the CNS, as part of multiple functions of astrocytes [22, 90, 91] (Fig. 1).
ATP is an ideal molecule for cell signaling due to its intrinsic properties such as its small size, diffusing molecule rate, instability and low concentration in the extracellular environment, and impossibility to cross the plasma membrane [92, 93]. These properties imply the presence of particular pathways for ATP release that could be associated with cellular excitation/response and cell-cell signaling [94, 95]. First, ATP may be stored in synaptic vesicles alone or with other neurotransmitters and then released, as a classic synaptic mechanism in the peripheral or central nervous system [96, 97]. Second, a nonvesicular mechanism of ATP release could be observed through gap junction hemichannels, ATP-binding cassette proteins, P2X7 receptor pores in glial cells, and via chloride channels [98–101]. Third, ATP could be released due to cytolysis or cell damage. While this is not a physiological mechanism, it takes place following biological trauma and contributes to pathological conditions [102].
Subsequent to these mechanisms, the metabolism of the released ATP is regulated by a vast number of different families of ectonucleotidases in the synaptic cleft, including the ectonucleoside triphosphate diphosphohydrolase (E-NTPDase) and the ectonucleotide pyrophosphatase phosphodiesterase (E-NPP) which catalyze the degradation of ATP to ADP or AMP. The degradation to adenosine is mediated by ecto-5′-nucleotidase (E-5′-NT) and alkaline phosphatase [91, 103] (Fig. 1). Consequently, the reaction products resulting from the ATP hydrolysis may bind to P2 receptors, in the case of ADP, or to P1 receptors in the case of adenosine [104].
The adenosinergic receptor ligand adenosine is recognized as an important regulator of cellular homeostasis in the CNS and may be involved in the prevention or induction of apoptosis [105]. The reduction of ectonucleotidase activity in certain pathological conditions provided additional evidence for the accumulation of ATP in the extracellular environment [20]. Therefore, the complexity of the communication of neural and nonneural cells expands the functional significance by the interaction of the purinergic receptors in association with a variety of neurotransmitter systems.
ATP-mediated neuron-glia signaling
Novel studies in the purinergic field began to converge with glial research as it became more widely accepted that ATP is released through synaptic vesicles and thus accessible to perisynaptic glial cells, allowing them to detect neuronal activity. In particular, glial cells are responsive to ATP, as all types of glia, such as astrocytes, oligodendrocytes, microglia, and Schwann cells, express purinergic receptors [91]. In Schwann cells and oligodendrocytes, ATP-mediated signaling predominantly occurs through P2Y receptors, which in turn trigger intracellular Ca2+ release [106, 107]. However, the function of P2X1–6 receptors in astrocytes remains unclear, although P2X-mediated currents could be detected in astrocyte cells in culture, and P2X7 receptors are widespread in these cells with possible contribution to pathological conditions [108, 109].
Glial cells express many types of neurotransmitter receptors and conventionally are considered to be nonexcitable [110, 111]. However, a surprising observation was reported by Dani et al. [112] that synaptic transmission may propagate to glial cells as calcium waves, inducing membrane depolarization and regulating neurotransmitter release. These properties of glial cells suggest possible rapid communications between neurons and glia during synaptic transmission. This glial communication mechanism allows the released ATP to act onto adjacent astrocytes and neurons, thus supporting the propagation of Ca2+ waves in glial syncytium [113]. For example, in neuronal-glial cocultures prepared from hippocampus, ATP secreted by astrocytes was shown to inhibit glutamatergic synapses through activation of P2Y receptors [114].
The glial communication mechanism based on Ca2+ wave propagation could be inhibited by P2 receptor blockers or enzymes that rapidly hydrolyze extracellular ATP [115]. A stimulation applied to a single astrocyte in cocultures of rat forebrain astrocytes and associated neurons caused an elevation of [Ca2+]i and induced Ca2+ wave propagation in dorsal spinal cord through P2Y1 receptor activation [116] and glutamate release [117]. This new finding provided a parallel mechanism of intercellular communication that could allow astrocytes to detect synaptic function, propagate the information through neighboring glial cells, and then influence synaptic function in a distant part of the nervous system.
Purinergic receptor-calcium signaling in glial cells plays important roles during CNS development. P2X1, P2X4, and P2X7 receptors were expressed in microglia at rat embryonic stage (E16) [59]. Moreover, changes in P2X4 receptor expression in microglial cells during postnatal development of the rat cerebellum have been reported. P2X5 receptor immunoreactivity was also upregulated in microglia and granular cells. Both P1 and P2 receptors contribute to the modulation of oligodendrocyte (OP) development, since they have been shown to exert similar effects on OP proliferation and differentiation [118].
The majority of the studies of ATP action have been concerned with the short-term P2 receptor signaling that occurs in neurotransmission and in secretion [119]. Furthermore, there is increasing evidence that purines and pyrimidines can have trophic roles in neuritogenesis [120, 121], regeneration [122], and proliferation [123]. However, some purines by themselves have limited trophic effects in a few types of cells; they appear to be much more effective as neuritogenic agents when they are combined with other trophic factors, such as NGF. For instance, inosine and 5′AMP alone do not elicit neurite extensions in PC12 cells [124].
Heine et al. [53] demonstrated that P2 receptor activation induced fiber outgrowth in organotypic cocultures in rat hippocampus. Fiber outgrowth was inhibited in the presence of the purinergic antagonist PPADS, suggesting the involvement of P2 receptors. In another study, the synergistic interaction between bFGF and ATP was reported on DNA synthesis in primary cultures of rat cortical astrocytes. ATP and bFGF induced a twofold and tenfold incorporation of [3H]thymidine into astrocytes, respectively, but when ATP and bFGF were added at the same time a 50-fold increase in [3H]thymidine incorporation was observed [12].
Neuroprotection
ATP can activate P2X7 receptors in astrocytes to release glutamate, GABA, and also ATP which might regulate the excitability of neurons in certain pathological conditions [125]. It has been suggested that astrocytes can sense the severity of damage in the CNS by the amount of ATP released from damaged cells and that extracellular ATP concentration and the corresponding subtype of activated astrocytic P2 receptor modulate the tumor necrosis factor-α (TNF-α)-mediated inflammatory response [126]. After mechanical brain injury, the administration of PPADS facilitated the recovery of pathologically changed electroencephalograms [127]. These results suggest that interference with the ATP-induced excitatory responses could provide neuroprotection and possible therapeutic consequences.
Evidence for a neuroprotective role was also found for the adenosine A1 receptor in hippocampus. This cerebral region is highly sensitive to hypoxia and ischemia. The study of the action of hypoxia on synaptic transmission in hippocampal slices has suggested that substances being released during hypoxia, such as GABA, ACh, and even glutamate, may also play neuroprotective roles. However, the actions of these neurotransmitters become evident only when activation of P1 receptors is impaired, suggesting a critical role for this receptor during hypoxic events. These substances can operate in a redundant or even overprotective manner, acting as a substitute for some adenosine actions when the nucleoside is not operative [128].
Neuroimmune interactions
Microglia, the immune cells of the CNS, can be activated by purines and pyrimidines to release inflammatory cytokines such as IL-1, IL-6, and TNF-α. However, hyperstimulation of the immune reaction in the brain may accelerate neuronal damage. The P2X7 receptor is considered to have a potentially pivotal role in the regulation of various inflammatory conditions. ATP selectively suppresses the synthesis of the inflammatory protein microglial response factor through calcium influx via P2X7 receptors in microglia [129], which also leads to enhancement of interferon-γ (IFN-γ)-induced type II nitric oxide synthase (NOS) activity [130, 131]. P2X7 receptor activity also participated in ATP-induced IL-1 release from macrophages and microglia that had been primed with substances such as bacterial endotoxin [132] and was shown to stimulate the transcription of nuclear factor κB, TNF-α [133], the stress-activated protein kinases (SAPK)/JNK pathway [134], and the production of 2-arachidonoylglycerol, which is also involved in inflammation induction by microglial cells.
P2Y rather than P2X7 receptors seem to have a major role in the IL-6 production by microglial cells [135]. ATP evoked the release of plasminogen [136] and IL-6 [135]. The stimulation of microglia by either ATP or BzATP revealed neurotoxic properties and the involvement of the P2X7 receptor has been reported in excitotoxic/necrotic and apoptotic degeneration [109].
Neurological disorders
Epilepsy
Several anti-epileptic agents reduce the ability of astrocytes to transmit Ca2+ waves, raising the possibility that blockade of ATP-induced [Ca2+]i transients in astrocytes by purinergic receptor antagonists could offer new treatments for epileptic disorders. Antiepileptic effects of adenosine are mostly due to the well-known inhibitory actions of P1 receptors on synaptic transmission in the hippocampus. However, as recently pointed out, adenosine actions are not limited to presynaptic actions on glutamate release [137]. The intraventricular injection of high doses of ATP in rats evoked severe chronic-tonic convulsions, whereas lower doses of ATP or adenosine elicited a kinetic state with muscle weakness [138]. P2X2 and P2X4 receptor expression in the hippocampus of seizure-prone gerbils was significantly reduced compared with that of normal gerbils [139]. GABAA receptors mediated modulation of expression of both P2X2 and P2X4 receptors, which may play an important role in the regulation of seizure activity in the gerbil hippocampus [139]. P2X7 receptors are thought to play a definite, but not yet well defined role in epilepsy. Treatment with the GABAB receptor agonist baclofen and antagonist phaclofen resulted in increased and decreased P2X7 receptor expression in hippocampus, respectively [140]. These purinergic receptor responses were interpreted as compensatory responses to the modulation of GABAB receptor function [140]. It is noteworthy to mention that this positive relationship between P2X and GABAA receptors was also reported for the spinal cord [141] and dorsal root ganglia (DRG) [142]. In these populations of neurons, ATP-mediated P2X receptor function may participate in neuronal transmission accompanied by GABA-mediated actions [139].
Pain
The heteromeric channel comprised of P2X2 and P2X3 subunits was expressed almost exclusively in a subset of primary afferents implicated in nociception [143–145]. It has been observed that mechanical allodynia is reduced in mice with deleted P2X3 receptor genes [146, 147] in agreement with data obtained in rats that have been treated with intrathecal antisense oligonucleotides reducing expression of P2X3 receptors [148] or with the selective antagonist for P2X3 and P2X2/3 receptors A-317491 [148, 149]. P2X3 receptor knockout mice showed additional defects in afferent pathways.
The P2X4 receptor is also implicated in pain sensation. Activation of dorsal horn microglia and tactile allodynia developing several days after ligation of a spinal nerve were greatly reduced when gene expression of P2X4 receptor in the dorsal horn had been inhibited by the presence of intrathecal antisense oligonucleotides [150]. Accordingly, intraspinal administration of microglia following induction of expression and activity of P2X4 receptors produced tactile allodynia in naive rats. Intrathecal administration of cultured brain microglia produced allodynia, but only when the cells had been pretreated with ATP [150]. The inhibition of P2X4 receptor activity in microglia might be a new therapeutic strategy for pain induced by nerve injury.
Alzheimer’s disease
Alzheimer’s disease (AD) is caused by extracellular deposition of amyloid β-peptide, which can damage neurons, leading to their dysfunction and death [151]. ATP and, in particular, aluminum-ATP promoted the formation of thioflavin T-reactive fibrils of β-amyloid and an unrelated amyloidogenic peptide, which could be blocked by suramin [152].
Microglial cells are believed to contribute to the progression of AD and are known to release proinflammatory neurotoxic substances. Extracellular ATP, acting through the P2X7 receptor, can alter β-amyloid peptide-induced cytokine secretion from human macrophages and microglia and thus may be an important modulator of neuroinflammation in AD [153]. P2X7 receptors mediate superoxide production in primary microglia, and the expression of this receptor subtype was specifically upregulated around β-amyloid plaques in a transgenic mouse model of AD [154].
In contrast to the control human brain, the P2Y1 receptor was colocalized with a number of characteristic AD structures such as neurofibrillary tangles, neuritic plaques, and neuropil threads in the hippocampus and cortex [155]. In general, control brain tissue exhibited a greater and more abundant level of P2Y1 receptor immunostaining than AD tissue did, probably due to severe neuronal cell degeneration in most AD brains. The intense P2Y1 receptor staining observed over pathological AD structures might imply that this receptor is involved either directly or indirectly in signaling events mediating neurodegeneration of pyramidal cells. Alternatively, P2Y1 receptors might have other diverse signaling roles, possibly involved in the production of intracellular tau deposits or might even serve to stabilize these tangle structures in some way [156].
Ischemia/hypoxia
Under pathological conditions of hypoxia or ischemia, extracellular purine nucleotides leak from damaged cells and thereby may reach high concentrations in the extracellular space [157]. A direct participation of extracellular ATP and P2 receptors in ischemic stress has been reported in various cellular systems [157–160]. For example, P2X2 and P2X4 receptor expression in neurons and microglia, respectively, in the hippocampus of gerbils was upregulated following transient global ischemia [161]. Increased P2X7 receptor expression in astrocytes, microglia, and neurons appears to contribute to the mechanisms of cell death caused by in vivo and in vitro ischemia [162, 163]. Following induction of ischemia P2X7 receptor mRNA transcription and protein expression were elevated in cultured cerebellar granule neurons and organotypic hippocampal cultures [163]. Hence, the P2X7 receptor is apparently an important element in the mechanisms of cellular damage induced by hypoxia/ischemia. In many cell types, the activation of the P2X7 receptor led to rapid cytoskeletal rearrangements, such as membrane blebbing and cell lysis [164]. P2Y1 receptors are intensely expressed in Purkinje cells in deep layers of the cerebral cortex and in ischemia-sensitive areas of the hippocampus [165]. In conclusion, extensive evidence demonstrates a postischemic time- and region-dependent upregulation of P2X2,4,7 and P2Y1 receptor subtypes in neurons and glial cells and suggests a direct role of P2 receptors in the pathophysiology of cerebral ischemia in vitro and in vivo.
Trauma and axotomy
P2 receptors are suggested to be involved in neuronal reactions after axotomy. Colocalization and temporal coactivation of purinergic and nitrergic markers support this idea, indicating possible interactions between these two systems [166]. Following peripheral nerve lesions, P2X3 receptor expression in DRG neurons was changed [167]. The increased expression of P2X3 receptor mRNA in intact neurons indicates a role of this subtype in the post-injury pathomechanism in primary sensory neurons [167]. After spinal cord injury, large regions of the peritraumatic zone were characterized by a sustained process of pathologically high ATP release [168]. Spinal cord neurons express P2X7 receptors, and exposure to ATP led to high-frequency spiking, irreversible increases in [Ca2+]i and cell death. The administration of P2 receptor antagonists (PPADS, oxATP) after acute impact injury significantly improved functional recovery and diminished cell death in the peritraumatic zone [168]. The involvement of P2X1 and P2X2 receptors in neuronal reactions after hemicerebellectomy was also described [169]. Furthermore, neuronal NOS and P2 receptors were colocalized and showed temporal coactivation after cerebellar lesions, indicating a close relationship between these two systems [166]. In addition, in this mixed model of differentiation and axotomy, the colocalization of ataxin-2 (ax-2, involved in resistance to degeneration phenomena, which may be lost after mutation)-immunopositive cells and P2X2 receptors was demonstrated in neurons, and post-lesional induction of P2X1 receptor and ax-2 immunoreactivity was reported as well [170]. In vivo treatment of P2Y2 receptor-expressing sciatic nerves with ATP-γS increased expression levels of the growth-associated protein 43 (GAP-43) as a marker for axonal growth in wild-type but not in P2Y2 −/− mice [171].
Possible therapeutic manipulations to modulate astrocytic proliferation and to diminish glial scar formation in the adult brain and during development include the use of drugs known to interfere with nucleotide synthesis. Pekovic et al. [172] showed that treatment with the purine nucleoside analogue ribavirin (Virazole; 1-β-D-ribofuranosyl-1,2,4-triazole-3-carboxamide) downregulates the process of reactive gliosis after sensory motor cortex lesion of the adult brain and facilitates re-establishing synaptic connections with the denervated cells at the lesion site. This may be a useful approach for improving neurological recovery from brain damage. The antiproliferative effect of ribavirin is due to the inhibition of de novo nucleic acid synthesis after depletion of GTP and dGTP pools with consequent impairment of specific transduction pathways.
ATP-induced effects on cell cycle progression
There is evidence showing that extracellular ATP enhances the expression of cell cycle regulating proteins [173, 174]. Progression of the cell cycle is highly controlled. Cyclins are synthesized and degraded in a synchronous way due to changing transcription or proteolysis rates, thereby directing the periods of the cellular cycle. Cyclins interact with cyclin-dependent kinases (cdks) resulting in activation of their kinase activity, phosphorylating their targets and themselves, and regulating the specific progression of the cell cycle through checkpoints [175].
Proliferation rates in mammalians are largely determined during the G1 phase of the cell cycle. The relevant proteins include three D-type cyclins (D1, D2, and D3) that, in different combinations, bind to and allosterically regulate one of two cdk subunits, cdk4 and cdk6, as well as the E-type cyclins (E1 and E2), which govern the activity of a single catalytic subunit, cdk2 [176]. Various combinations of D-type cyclins are expressed in different cell types, whereas cyclin E-cdk2 complexes are ubiquitously expressed [177].
Two families of cdk inhibitors regulate the activity of G1-type cyclins-cdks complexes: the Ink4 family (p16, p15, p18, and p19), which blocks the activity of cyclin D-cdk4-6 complexes, and the Cip/Kip family (p21, p27, and p57), which preferentially inhibits cyclin E-cdk2 complexes and also acts as a scaffold for the catalytically active cyclin D-cdk4-6 complexes. In addition to cyclins and cdks, mitogen-activated protein kinase (MAPK) is also believed to have a role in induction of cell proliferation. Therefore, cyclin D-dependent kinases may play a role in controlling the cell cycle of embryonic and maybe neural progenitor cells. In addition MAPK is also believed to have a role in induction of cell proliferation. Extracellular ATP induces Ca2+-dependent MAPK activation via stimulation of P2 receptors in neonatal rat astrocytes [178]. On the other hand, cell proliferation is associated with activation of diverse proteins. Positive regulators include cyclins and their partners with catalytic activity (cdks), which are essential for progression of the cells through each phase of the cell cycle and various cell cycle checkpoints [179, 180]. The regulation of cyclin D1 expression is also mediated by the Ras/ERK signaling pathways [181, 182]. Raf/MEK/ERK and PI3-K/Akt signaling pathways can act in synergy to promote the G1-S phase cell cycle progression in both normal and cancer cells [183, 184]. The promoter for cyclin D1 contains an AP-1 site, and the ectopic expression of either c-fos or c-jun induces cyclin D1 mRNA expression [185, 186]. In many cell types, phosphatidylinositol (PI)-3-kinase-dependent signaling pathways also regulate cyclin D1 expression [187]. It was also reported that the control of the cell cycle regulatory proteins was dependent on PI3-kinase and p44/42 MAPK pathways, indicating that extracellular ATP alone is sufficient to induce cell cycle progression beyond the G1 phase of the cell cycle. These findings also suggest that, once P2 receptors are activated, protein kinase C (PKC) transmit signals to the nucleus through one or more of the MAPK cascades, which may include Raf-1, MEK, and ERK, and stimulate transcription factors such as myc, max, fos, and jun. Moreover, MAPKs are upstream regulators of cdk2 and cdk4 expression. It has been reported that p44/42 MAPK phosphorylation is essential and sufficient for the increase in cdk2 [188, 189] and decrease in p27Kip1 expression [190, 191]. However, Delmas et al. [192] provided evidence that p44/p42 MAPK activation triggers p27Kip1 degradation independently from cdk2/cyclin E in NIH 3T3 cells. As described above, ATP regulation of the MAPK and cdk-cyclin complex has not been elucidated in other types of cells [193].
It is documented in the literature that purinergic receptor inhibitors interfere with the S phase of the cell cycle. Neurospheres treated with the purinergic receptor antagonists reactive blue 2 or suramin are mostly in S phase (5.7 ± 0.3% or 8.4 ± 2.3%) when compared to untreated control neurospheres with 16.4 ± 1.8% of the cells being in S phase. Moreover, neurosphere cultures treated with suramin or reactive blue 2 showed an increase in the expression of the tumor suppressor p27 as a strong regulator of cell division [49].
The discussed findings led to the suggestion that extracellular ATP plays an important physiological role during mammalian embryonic development by stimulating proliferation of ES cells, and therefore P2 receptor agonists and antagonists might provide novel and powerful tools for modulating embryonic cell functions. In conclusion, P2X and P2Y purinergic receptors can promote proliferation of ES cells as well as of progenitor cell types by a mechanism by that ATP induces increases in [Ca2+]i, leading to activation of PKC, PI3-kinase/Akt, p38, and p44/42 MAPK, followed by an alteration in the cdk-cyclin complex with p21 and p27, which are involved in stimulation of cell proliferation.
Pharmacological approaches
Most purinergic receptors do not have specific inhibitors. Therefore, P2 receptor agonists and antagonists acting on most of the purinergic receptor subtypes are widely used in experimental approaches to study biological functions of these receptors. Such approaches are feasible, since these compounds mostly have higher affinities to some P2 receptor subtypes than to other ones. As an example, we have used suramin, PPADS, and reactive blue 2 to study the participation of P2Y1, P2Y2, and P2X2 receptors in neuronal differentiation of P19 EC cells [38].
One possible approach towards a subtype-specific inhibitor would be based on results from P2 receptor structure determination. Using site-directed mutagenesis it has been possible to understand which amino acids are involved in ATP binding and to identify allosteric sites in purinergic receptors. The knowledge obtained on location and structural features of ligand and inhibitor binding sites is used in rational based drug design of selective purinergic subtype antagonists. Alternatively, combinatorial libraries formed by vast amounts of possible ligands can be employed for discovery of subtype-specific inhibitors.
A-317491 was identified as a specific inhibitor for P2X2/3 and P2X3 receptors. In the presence of A-317491 both thermal hyperalgesia and mechanical allodynia were attenuated after chronic nerve constriction injury in which P2X3 homomeric and P2X2/3 heteromeric receptor activities were involved. Although active in chronic pain models, A-317491 was ineffective in reducing nociception in animal models of acute postoperative pain and visceral pain indicating that P2X3 and P2X2/3 receptor activation may not be a major mediator of acute postoperative or visceral pain [149]. MRS 2179 (2′-deoxy-N 6-methyladenosine 3′,5′-bisphosphate) was discovered as a specific inhibitor of P2Y1 receptor activity [194]. This compound has an efficient antithrombotic action in which P2Y1 receptors are involved [195].
Based on structure design or combinatorial library approaches specific agonists or antagonists may be discovered for other purinergic receptor subtypes. For instance, the SELEX (systematic evolution of ligands by exponential enrichment) technique provides a particularly promising approach for the discovery of such compounds. This technique is based on the reiterative presentation of a partial random RNA or single-stranded DNA library to a protein preparation containing a particular purinergic receptor subtype. RNA or DNA molecules bound to a target site on the receptor are displaced from the receptor and eluted by addition of an excess concentration of an unspecific purinergic receptor antagonist and amplified by reverse transcription polymerase chain reaction (PCR) or PCR to restore the library used for the next in vitro selection cycle. Using this approach, it was possible to identify inhibitors specific for isoforms of a target protein [196]. Our group prepared membrane protein fractions of 1321N1 cells stably transfected with rat P2X2 receptors and coupled them onto an immobilized artificial membrane (IAM) as matrix for affinity chromatography. The equilibrium binding to the receptor and competition between ATP and the purinergic antagonists suramin and 2′3′-O-(2,4,6-trinitrophenyl) adenosine 5′-triphosphate (TNP-ATP) were analyzed by a chromatographic assay using [α-32P]-ATP as a radioligand. Our data indicate that suramin does not compete with ATP for the ligand binding site and TNP-ATP is a competitive antagonist, as already shown by Trujillo et al. [197]. Moreover, this chromatographic assay can be used in in vitro selection procedures for RNA aptamers binding to P2X2 receptors from a combinatorial SELEX RNA library [198]. The development of a subtype-specific P2X receptor antagonist by using the SELEX technique or another combinatorial library-based approach shall serve as proof of principle and encourage further works to obtain such specific antagonists for all P2 receptor subtypes as tools for elucidating their biological functions and for possible therapeutic applications.
Conclusion
P2 receptor function is involved in most physiological processes and participates in neurotransmission in the CNS. Results obtained with mouse ES and P19 EC and neural progenitor cells suggest an important role of purinergic signaling in early embryogenesis, especially in cell proliferation, migration, and differentiation, with different subtypes of receptors participating in these processes. Our understanding of the biological functions of specific P2 receptor subtypes during CNS development and in the adult brain has increased due to the availability of knockout animals and specific inhibition of gene expression or activity of purinergic receptor subtypes. The importance of P2 receptor signaling in neuroprotection, neuroimmunity, and guiding neuronal differentiation, especially in glial and microglial cells, has been related to purinergic receptor expression. Most importantly, specific agonists and antagonists for individual P2 receptor subtypes are both needed for studying their involvement in biological processes. The discovery of such selective compounds will elucidate yet unknown biological functions of P2 receptor subtypes as well as open new avenues for therapeutic approaches to disease states in which purinergic receptor activity is involved.
Abbreviations
- ACh:
-
acetylcholine
- AD:
-
Alzheimer’s disease
- AP:
-
alkaline phosphatase
- ax-2:
-
ataxin-2
- cdks:
-
cyclin-dependent kinases
- CNS:
-
central nervous system
- DRG:
-
dorsal root ganglia
- EC:
-
embryonal carcinoma
- NTPDase2:
-
ectonucleoside triphosphate diphosphohydrolase 2
- E-NPP:
-
ectonucleotide pyrophosphatase phosphodiesterase
- EGF:
-
epidermal growth factor
- E-NTPDase:
-
ectonucleoside triphosphate diphosphohydrolase
- ES cells:
-
embryonic stem cells
- E-5′-NT:
-
ecto-5′-nucleotidase
- FGF-2:
-
fibroblast growth factor 2
- GABA:
-
γ-aminobutyric acid
- GFAP:
-
glial fibrillary acidic protein
- IFN-γ:
-
interferon-γ
- LIF:
-
leukemia inhibitory factor
- LTP:
-
long-term potentiation
- MAP-2:
-
microtubule-associated protein-2
- MAPK:
-
mitogen-activated protein kinase
- MRF:
-
microglial response factor
- MRS 2179:
-
2′-deoxy-N 6-methyladenosine 3′,5′-bisphosphate
- NPC:
-
neural progenitor cells
- NPC:
-
neural stem cells
- NA:
-
noradrenaline
- NOS:
-
nitric oxide synthase
- NTPDase:
-
ectonucleoside triphosphate diphosphohydrolase
- NGF:
-
nerve growth factor
- OP:
-
oligodendrocyte
- oxATP:
-
oxidized ATP
- PPADS:
-
pyridoxalphosphate-6-azophenyl-2′,4′-disulfonic acid
- SVZ:
-
subventricular zone
- TGF:
-
transforming growth factor
- TNF-α:
-
tumor necrosis factor-α
- GAP-43:
-
growth-associated protein 43
- TNAP:
-
tissue-nonspecific alkaline phosphatase
- TNP-ATP:
-
2′3′-O-(2,4,6-trinitrophenyl) adenosine 5′-triphosphate
References
Evans RJ, Derkach V, Surprenant A (1992) ATP mediates fast synaptic transmission in mammalian neurons. Nature 357:503–505
Silinsky EM, Gerzanich V, Vanner SM (1992) ATP mediates excitatory synaptic transmission in mammalian neurones. Br J Pharmacol 106:762–763
Burnstock G (1978) A basis for distinguishing two types of purinergic receptor. In: Straub RW, Bolis L (eds) Cell membrane receptors for drugs and hormones: a multidisciplinary approach. Raven, New York, pp 107–118
Burnstock G, Kennedy C (1985) Is there a basis for distinguishing two types of P2 purinoceptor? Gen Pharmacol 16:433–440
Brandle U, Zenner H-P, Ruppersberg JP (1999) Gene expression of P2X receptors in the developing inner ear of the rat. Neurosci Lett 273:105–108
Adrian K, Bernhard MK, Breitinger H-G et al (2000) Expression of purinergic receptors (ionotropic P2X1-7 and metabotrophic P2Y1-11) during myeloid differentiation of HL60 cells. Biochim Biophys Acta 1492:127–138
Orriss, IR, Knight GE, Ranasinghe S et al (2006) Osteoblast responses to nucleotides increase during differentiation. Bone 39:300–309
Berchtold S, Ogilvie ALL, Bogdan C et al (1999) Human monocyte derived dendritic cells express functional P2X and P2Y receptors as well as ecto-nucleotidases. FEBS Lett 458:424–428
Ryten M, Dunn PM, Neary JT et al (2002) ATP regulates the differentiation of mammalian skeletal muscle by activation of a P2X5 receptor on satellite cells. J Cell Biol 158:345–355
Wang L, Jacobsen SE, Bengtsson A et al (2004) P2 receptor mRNA expression profiles in human lymphocytes, monocytes and CD34+ stem and progenitor cells. BMC Immunol 5:16
Huang N, Wang DJ, Heppel LA (1989) Extracellular ATP is a mitogen for 3T3, 3T6, and A431 cells and acts synergistically with other growth factors. Proc Natl Acad Sci U S A 86:7904–7908
Neary JT, Whittemore SR, Zhu Q et al (1994). Synergistic activation of DNA synthesis in astrocytes by fibroblast growth factor and extracellular ATP. J Neurochem 63:490–494
Lemoli RM, Ferrari D, Fogli M et al (2004) Extracellular nucleotides are potent stimulators of human hematopoietic stem cells in vitro and in vivo. Blood 104:1662–1670
Neary JT, Zhu Q (1994) Signaling by ATP receptors in astrocytes. Neuroreport 5:1617–1620
Lenz G, Gottfried C, Luo ZJ et al (2000) P2Y purinoceptor subtypes recruit different Mek activators in astrocytes. Br J Pharmacol 129:927–936
Salter M, Hicks JL (1995) ATP causes release of intracellular Ca2+ via the phospholipase C beta/IP3 pathway in astrocytes from the dorsal spinal cord. J Neurosci 15:2961–2971
Miranov SL (1994) Metabotropic ATP receptor in hippocampal and thalamic neurones: pharmacology and modulation of Ca2+ mobilizing mechanisms. Neuropharmacology 33:1–13
Spitzer NC, Root CM, Borodinsky LN (2004) Orchestrating neuronal differentiation: patterns of Ca2+ spikes specify transmitter choice. Trends Neurosci 27:415–421
Gu X, Spitzer NC (1997) Breaking the code: regulation of neuronal differentiation by spontaneous calcium transients. Dev Neurosci 19:33–41
Zimmermann H (1996) Biochemistry, localization and functional roles of ecto-nucleotidases in the nervous system. Prog Neurobiol 49:589–618
Götz M, Huttner WB (2005) The cell biology of neurogenesis. Nat Rev Mol Cell Biol 6:777–788
Abbracchio MP, Burnstock G (1998) Purinergic signaling: pathophysiological roles. Jpn J Pharmacol 78:113–145
Laasberg T (1990) Ca2+-mobilizing receptors of gastrulating chick embryo. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol 97:9–12
Katz LC, Shatz CJ (1996) Synaptic activity and the construction of cortical circuits. Science 274:1133–1138
Weissman TA, Riquelme PA, Ivic L et al (2004) Calcium waves propagate through radial glial cells and modulate proliferation in the developing neocortex. Neuron 43:647–661
Cicirata F, Meli C, Castorina C et al (1991) Neurotransmitter amino acid levels in rat thalamus and cerebral cortex after cerebellectomy. Int J Dev Neurosci 9:365–369
Miranda-Contreras L, Mendoza-Briceno RV, Palacios-Pru EL (1998) Levels of monoamine and amino acid neurotransmitters in the developing male mouse hypothalamus and in histotypic hypothalamic cultures. Int J Dev Neurosci 16:403–412
Miranda-Contreras L, Benitez-Diaz PR, Mendoza-Briceno RV et al (1999) Levels of amino acid neurotransmitters during mouse cerebellar neurogenesis and in histotypic cerebellar cultures. Dev Neurosci 21:147–158
Miranda-Contreras L, Ramirez-Martens LM, Benitez-Diaz PR et al (2000) Levels of amino acid neurotransmitters during mouse olfactory bulb neurogenesis and in histotypic olfactory bulb cultures. Int J Dev Neurosci 18:83–91
Flint AC, Liu X, Kriegstein AR (1998) Nonsynaptic glycine receptor activation during early neocortical development. Neuron 20:43–53
Haydar TF, Wang F, Schwartz ML et al (2000) Differential modulation of proliferation in the neocortical ventricular and subventricular zones. J Neurosci 20:5764–5774
Evans MJ, Kaufman M (1981) Establishment in culture of pluripotent cells from mouse embryos. Nature 292:154–156
McWhir J, Schnieke AE, Ansell R et al (1996) Selective ablation of differentiated cells permits isolation of embryonic stem cell lines from murine embryos with a non-permissive genetic background. Nat Genet 14:223–226
Heo SJ, Han HJ (2006) ATP stimulates mouse embryonic stem cell proliferation via protein kinase C, phosphatidylinositol 3-kinase/Akt, and mitogen-activated protein kinase signaling pathways. Stem Cells 24:2637–2648
Anneren C, Cowan CA, Melton DA (2004) The Src family of tyrosine kinases is important for embryonic stem cell self-renewal. J Biol Chem 279:31590–31598
Martin GR (1981) Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc Natl Acad Sci U S A 78:7634–7638
Ulrich H, Majumder P (2006) Neurotransmitter receptor expression and activity during neuronal differentiation of embryonal carcinoma and stem cells: from basic research towards clinical applications. Cell Prolif 39:281–300
Resende RR, Majumder P, Gomes KN et al (2007) P19 embryonal carcinoma cells as in vitro model for studying purinergic receptor expression and modulation of N-methyl-d-aspartate-glutamate and acetylcholine receptors during neuronal differentiation. Neuroscience 146:1169–1181
da Silva RL, Resende RR, Ulrich H (2007) Alternative splicing of P2X6 receptors in developing mouse brain and during in vitro neuronal differentiation. Exp Physiol 92:139–415
Gage FH, Coates PW, Palmer TD et al (1995) Survival and differentiation of adult neuronal progenitor cells transplanted to the adult brain. Proc Natl Acad Sci U S A 92:11879–11883
Hogg RC, Chipperfield H, Whyte KA et al (2004) Functional maturation of isolated neural progenitor cells from the adult rat hippocampus. Eur J Neurosci 19:2410–2420
Pluchino S, Zanotti L, Deleidi M et al (2005) Neural stem cells and their use as therapeutic tool in neurological disorders. Brain Res Rev 48:211–219
Cameron HA, Tanapat P, Gould E (1998) Adrenal steroids and N-methyl-D-aspartate receptor activation regulate neurogenesis in the dentate gyrus of adult rats through a common pathway. Neuroscience 82:349–354
Svendsen CN, ter Borg MG, Armstrong RJ et al (1998) A new method for the rapid and long term growth of human neural precursor cells. J Neurosci Methods 85:141–152
Shukla V, Zimmermann H, Wang L et al (2005) Functional expression of the ecto-ATPase NTPDase2 and of nucleotide receptors by neuronal progenitor cells in the adult murine hippocampus. J Neurosci Res 80:600–610
Braun N, Sevigny J, Mishra SK et al (2003) Expression of the ecto-ATPase NTPDase2 in the germinal zones of the developing and adult rat brain. Eur J Neurosci 17:1355–1364
Mishra SK, Braun N, Shukla V et al (2006) Extracellular nucleotide signaling in adult neural stem cells: synergism with growth factor-mediated cellular proliferation. Development 133:675–684
Leon C, Hechler B, Freund M et al (1999) Defective platelet aggregation and increased resistance to thrombosis in purinergic P2Y(1) receptor-null mice. J Clin Invest 104:1731–1737
Lin JH, Takano T, Arcuino G et al (2007) Purinergic signaling regulates neural progenitor cell expansion and neurogenesis. Dev Biol 302:356–366
Ryu JK, Choi HB, Hatori K et al (2003) Adenosine triphosphate induces proliferation of human neural stem cells: role of calcium and p70 ribosomal protein S6 kinase. J Neurosci Res 72:352–362
Tran PB, Ren D, Veldhouse TJ et al (2004) Chemokine receptors are expressed widely by embryonic and adult neural progenitor cells. J Neurosci Res 76:20–34
Safiulina VF, Kasyanov AM, Sokolova E et al (2005) ATP contributes to the generation of network-driven giant depolarizing potentials in the neonatal rat hippocampus. J Physiol 565:981–992
Heine C, Heimrich B, Vogt J et al (2006) P2 receptor-stimulation influences axonal outgrowth in the developing hippocampus in vitro. Neuroscience 138:303–311
Burnstock G (1996) Development and perspectives of the purinoceptor concept. J Auton Pharmacol 16:295–302
Torres IL, Battastini AM, Buffon A et al (2003) Ecto-nucleotidase activities in spinal cord of rats changes as function of age. Int J Dev Neurosci 21:425–429
Cheung KK, Burnstock G (2002) Localization of P2X3 receptors and coexpression with P2X2 receptors during rat embryonic neurogenesis. J Comp Neurol 443:368–382
Cheung KK, Ryten M, Burnstock G (2003) Abundant and dynamic expression of G protein-coupled P2Y receptors in mammalian development. Dev Dyn 228:254–266
Cheung KK, Chan WY, Burnstock G (2005) Expression of P2X purinoceptors during rat brain development and their inhibitory role on motor axon outgrowth in neural tube explant cultures. Neuroscience 133:937–945
Xiang Z, Burnstock G (2005) Expression of P2X receptors on rat microglial cells during early development. Glia 52:119–126
Kidd EJ, Miller KJ, Sansum AJ et al (1998) Evidence for P2X3 receptors in the developing rat brain. Neuroscience 87:533–539
Brosenitsch TA, Adachi T, Lipski J et al (2005) Developmental downregulation of P2X3 receptors in motoneurons of the compact formation of the nucleus ambiguous. Eur J Neurosci 22:809–824
Narcisse L, Scemes E, Zhao Y et al (2005) The cytokine IL-1beta transiently enhances P2X7 receptor expression and function in human astrocytes. Glia 49:245–258
Sperlágh B, Köfalvi A, Deuchars D et al (2002) Involvement of P2X7 receptors in the regulation of neurotransmitter release in the rat hippocampus. J Neurochem 81:1196–1211
Feuvre RL, Brough D, Rothwell N (2002) Extracellular ATP and P2X7 receptors in neurodegeneration. Eur J Pharmacol 447:261–269
Xiang Z, Burnstock G (2005) Changes in expression of P2X purinoreceptor in rat cerebellum during postnatal development. Dev Brain Res 156:147–157
Lalo U, Voitenko N, Kostyuk P (1998) Iono- and metabotropically induced purinergic calcium signalling in rat neocortical neurons. Brain Res 799:285–291
Ciccolini F, Collins TJ, Sudhoelter J et al (2003) Local and global spontaneous calcium events regulate neurite outgrowth and onset of GABAergic phenotype during neural precursor differentiation. J Neurosci 23:103–111
Maric D, Maric I, Chang YH et al (2003) Prospective cell sorting of embryonic rat neural stem cells and neuronal and glial progenitors reveals selective effects of basic fibroblast growth factor and epidermal growth factor on self-renewal and differentiation. J Neurosci 23:240–251
Scemes E, Duval N, Meda P (2003) Reduced expression of P2Y1 receptors in connexin43-null mice alters calcium signaling and migration of neural progenitor cells. J Neurosci 23:11444–11452
Hyden H (1961) Satellite cells in the nervous system. Sci Am 205:62–70
Vernadakis A (1996) Glia-neuron intercommunications and synaptic plasticity. Prog Neurobiol 49:185–214
Imura T, Kornblum HI, Sofroniew MV (2003) The predominant neural stem cell isolated from postnatal and adult forebrain but not early embryonic forebrain expresses GFAP. J Neurosci 23:2824–2832
Garcia AD, Doan NB, Imura T et al (2004) GFAP-expressing progenitors are the principal source of constitutive neurogenesis in adult mouse forebrain. Nat Neurosci 7:1233–1241
Powell EM, Geller HM (1999) Dissection of astrocyte-mediated cues in neuronal guidance and process extension. Glia 26:73–83
Merkle FT, Alvarez-Buylla A (2006) Neural stem cells in mammalian development. Curr Opin Cell Biol 18:704–709
Song H, Stevens CF, Gage FH (2002) Astroglia induce neurogenesis from adult neural stem cells. Nature 417:39–44
Svendsen CN (2002) The amazing astrocyte. Nature 417:29–32
Montgomery DL (1994) Astrocytes: form, functions, and roles in disease. Vet Pathol 31:145–167
Rose CR, Ransom BR (1997) Gap junctions equalize intracellular Na+ concentration in astrocytes. Glia 20:299–307
Cotrina ML, Lin JH, Alves-Rodrigues A et al (1998) Connexins regulate calcium signaling by controlling ATP release. Proc Natl Acad Sci U S A 95:15735–15740
Cotrina ML, Lin JH, Nedergaard M (1998) Cytoskeletal assembly and ATP release regulate astrocytic calcium signaling. J Neurosci 18:8794–8804
Ciccarelli R, Ballerini P, Sabatino G et al (2001) Involvement of astrocytes in purine-mediated reparative processes in the brain. Int J Dev Neurosci 19:395–414
Fields RD, Stevens-Graham B (2002) New insights into neuron-glia communication. Science 298:556–562
Hansson E, Ronnback L (2003) Glial neuronal signaling in the central nervous system. FASEB J 17:341–348
Hagg T (2005) Molecular regulation of adult CNS neurogenesis: an integrated view. Trends Neurosci 28:589–595
Araque A, Perea G (2004) Glial modulation of synaptic transmission in culture. Glia 47:241–248
Lin SC, Bergles DE (2004) Synaptic signaling between neurons and glia. Glia 47:290–298
Araque A, Carmignoto G, Haydon PG (2001) Dynamic signaling between astrocytes and neurons. Annu Rev Physiol 63:795–813
Hertz L, Zielke HR (2004) Astrocytic control of glutamatergic activity: astrocytes as stars of the show. Trends Neurosci 27:735–743
Cotrina ML, Lin JH, Lopez-Garcia JC et al (2000) ATP-mediated glia signaling. J Neurosci 20:2835–2844
Fields RD, Burnstock G (2006) Purinergic signalling in neuron-glia interactions. Nat Rev Neurosci 7:423–436
Chaudry IH (1982) Does ATP cross the cell plasma membrane. Yale J Biol Med 55:1–10
Fields RD, Stevens B (2000) ATP: an extracellular signaling molecule between neurons and glia. Trends Neurosci 23:625–633
Vizi ES, Burnstock G (1988) Origin of ATP release in the rat vas deferens: concomitant measurement of [3H]noradrenaline and [14C]ATP. Eur J Pharmacol 158:69–77
Bodin P, Burnstock G (2001) Purinergic signalling: ATP release. Neurochem Res 26:959–969
Edwards FA, Gibb AJ, Colquhoun D (1992) ATP receptor mediated synaptic currents in the central nervous system. Nature 359:144–146
Mori M, Heuss C, Gahwiler BH et al (2001) Fast synaptic transmission mediated by P2X receptors in CA3 pyramidal cells of rat hippocampal slice cultures. J Physiol 535:115–123
Reisin IL, Prat AG, Abraham EH et al (1994) The cystic fibrosis transmembrane conductance regulator is a dual ATP and chloride channel. J Biol Chem 269:20584–20591
Schwiebert EM (1999) ABC transporter-facilitated ATP conductive transport. Am J Physiol 276:C1–C8
Stout CE, Costantin JL, Naus CC et al (2002) Intercellular calcium signaling in astrocytes via ATP release through connexin hemichannels. J Biol Chem 277:10482–10488
Suadicani SO, Brosnan CF, Scemes E (2006) P2X7 receptors mediate ATP release and amplification of astrocytic intercellular Ca2+ signaling. J Neurosci 26:1378–1385
Franke H, Krugel U, Illes P (2006) P2 receptors and neuronal injury. Pflugers Arch 452:622–644
Zimmermann H (2000) Extracellular metabolism of ATP and other nucleotides. Naunyn Schmiedebergs Arch Pharmacol 362:299–309
Ralevic V, Burnstock G (1998) Receptors for purines and pyrimidines. Pharmacol Rev 50:413–492
Ribeiro JA, Sebastiao AM, de Mendonca A (2003) Participation of adenosine receptors in neuroprotection. Drug News Perspect 16:80–86
Bennett MR, Buljan V, Farnell L et al (2006) Purinergic junctional transmission and propagation of calcium waves in spinal cord astrocyte networks. Biophys J 91:3560–3571
Butt AM (2006) Neurotransmitter-mediated calcium signalling in oligodendrocyte physiology and pathology. Glia 54:666–675
Walz W, Gimpl G, Ohlemeyer C et al (1994) Extracellular ATP-induced currents in astrocytes: involvement of a cation channel. J Neurosci Res 38:12–18
Franke H, Illes P (2006) Involvement of P2 receptors in the growth and survival of neurons in the CNS. Pharmacol Ther 109:297–324
Deitmer JW, Verkhratsky AJ, Lohr C (1998) Calcium signalling in glial cells. Cell Calcium 24:405–416
Vesce S, Bezzi P, Volterra A (1999) The active role of astrocytes in synaptic transmission. Cell Mol Life Sci 56:991–1000
Dani JW, Chernjavsky A, Smith SJ (1992) Neuronal activity triggers calcium waves in hippocampal astrocyte networks. Neuron 8:429–440
Guthrie PB, Knappenberger J, Segal M et al (1999) ATP released from astrocytes mediates glial calcium waves. J Neurosci 19:520–528
Zhang JM, Wang HK, Ye CQ et al (2003) ATP released by astrocytes mediates glutamatergic activity-dependent heterosynaptic suppression. Neuron 40:971–982
Innocenti B, Parpura V, Haydon PG (2000) Imaging extracellular waves of glutamate during calcium signaling in cultured astrocytes. J Neurosci 20:1800–1808
Fam SR, Gallagher CJ, Salter MW (2000) P2Y(1) purinoceptor-mediated Ca(2+) signaling and Ca(2+) wave propagation in dorsal spinal cord astrocytes. J Neurosci 20:2800–2808
Domercq M, Brambilla L, Pilati E et al (2006) P2Y1 receptor-evoked glutamate exocytosis from astrocytes: control by tumor necrosis factor-alpha and prostaglandins. J Biol Chem 281:30684–30696
Agrestia C, Meomartinib ME, Amadiob S et al (2005) ATP regulates oligodendrocyte progenitor migration, proliferation, and differentiation: involvement of metabotropic P2 receptors. Brain Res Rev 48:157–165
Burnstock G (2006) Pathophysiology and therapeutic potential of purinergic signaling. Pharmacol Rev 58:58–86
Gysbers JW, Rathbone MP (1992) Guanosine enhances NGF-stimulated neurite outgrowth in PC12 cells. Neuroreport 3(11):997–1000
D’Ambrosi N, Cavaliere F, Merlo D et al (2000) Antagonists of P2 receptor prevent NGF-dependent neuritogenesis in PC12 cells. Neuropharmacology 39:1083–1094
D’Ambrosi N, Murra B, Cavaliere F et al (2001) Interaction between ATP and nerve growth factor signalling in the survival and neuritic outgrowth from PC12 cells. Neuroscience 108:527–534
Sanches G, de Alencar LS, Ventura AL (2002) ATP induces proliferation of retinal cells in culture via activation of PKC and extracellular signal-regulated kinase cascade. Int J Dev Neurosci 20:21–27
Braumann T, Jastorff B, Richter-Landsberg C (1986) Fate of cyclic nucleotides in PC12 cell cultures: uptake, metabolism, and effects of metabolites on nerve growth factor-induced neurite outgrowth. J Neurochem 47:912–919
Wang CM, Chang YY, Kuo JS et al (2002) Activation of P2X(7) receptors induced [(3)H]GABA release from the RBA-2 type-2 astrocyte cell line through a Cl(-)/HCO(3)(-))-dependent mechanism. Glia 37:8–18
Kucher BM, Neary JT et al (2005) Bi-functional effects of ATP/P2 receptor activation on tumor necrosis factor-α release in lipopolysaccharide-stimulated astrocytes. J Neurochem 92:525–535
Krügel U, Kittner H, Franke H et al (2001) Accelerated functional recovery after neuronal injury by P2 receptor blockade. Eur J Pharmacol 420:R3–R4
Sebastião AM, de Mendonça A, Ribeiro JA et al (2001) Neuroprotection during hypoxic insults: role of adenosine. Drug Dev Res 52:291–295
Kaya N, Tanaka S, Koike T et al (2002) ATP selectively suppresses the synthesis of the inflammatory protein microglial response factor (MRF)-1 through Ca2+ influx via P2X7 receptors in cultured microglia. Brain Res 952:86–97
Gendron FP, Chalimoniuk M, Strosznajder J et al (2003) P2X7 nucleotide receptor activation enhances IFN gamma-induced type II nitric oxide synthase activity in BV-2 microglial cells. J Neurochem 87:344–352
Ohtani Y, Minami M, Satoh M et al (2000) Expression of inducible nitric oxide synthase mRNA and production of nitric oxide are induced by adenosine triphosphate in cultured rat microglia. Neurosci Lett 293:72–74
Ferrari D, Chiozzi P, Falzoni S et al (1997) Purinergic modulation of interleukin-1β release from microglial cells stimulated with bacterial endotoxin. J Exp Med 185:579–582
Hide I, Tanaka M, Inoue A et al (2000) Extracellular ATP triggers tumor necrosis factor alpha release from rat microglia. J Neurochem 75:965–972
Humphreys BD, Rice J, Kertesy SB et al (2000) Stress-activated protein kinase/JNK activation and apoptotic induction by the macrophage P2X7 nucleotide receptor. J Biol Chem 275:26792–26798
Shigemoto-Mogami Y, Koizumi S, Tsuda M et al (2001) Mechanisms underlying extracellular ATP-evoked interleukin-6 release in mouse microglial cell line, MG-5. J Neurochem 78:1339–1349
Inoue K, Nakajima K, Morimoto T et al (1998) ATP stimulation of Ca2+-dependent plasminogen release from cultured microglia. Br J Pharmacol 123:1304–1310
Khakh BS (2001) Molecular physiology of P2X receptors and ATP signalling at synapses. Nat Rev Neurosci 2:165–174
Knutsen LJ, Murray TF et al (1997) Adenosine and ATP in epilepsy. In: Jacobson KA, Jarvis (eds) Purinergic approaches in experimental therapeutics. Wiley-Liss, Danvers, MA
Kang TC, An SJ, Park SK et al (2003) P2X2 and P2X4 receptor expression is regulated by a GABA(A) receptor-mediated mechanism in the gerbil hippocampus. Mol Brain Res 116:168–175
Kang TC, Park SK, Hwang IK et al (2004) GABA(B) receptor-mediated regulation of P2X7 receptor expression in the gerbil hippocampus. Mol Brain Res 121:12–18
Stoeckel ME, Uhl-Bronner S, Hugel S et al (2003) Cerebrospinal fluid-contacting neurons in the rat spinal cord, a gamma-aminobutyric acidergic system expressing the P2X2 subunit of purinergic receptors, PSA-NCAM, and GAP-43 immunoreactivities: light and electron microscopic study. J Comp Neurol 457:159–174
Labrakakis C, Tong CK, Weissman T et al (2003) Localization and function of ATP and GABAA receptors expressed by nociceptors and other postnatal sensory neurons in rat. J Physiol 549:131–142
Chen CC, Akopian AN, Sivilotti L et al (1995) A P2X purinoceptor expressed by a subset of sensory neurons. Nature 377:428–431
Lewis CJ, Neidhart S, Holy C et al (1995) Coexpression of P2X2 and P2X3 receptor subunits can account for ATP-gated currents in sensory neurones. Nature 377:432–434
Khakh BS, Humphrey PP, Surprenant A (1995) Electrophysiological properties of P2X-purinoceptors in rat superior cervical, nodose and guinea-pig coeliac neurones. J Physiol 484:385–395
Cockayne DA, Hamilton SG, Zhu QM et al (2000) Urinary bladder hyporeflexia and reduced pain-related behaviour in P2X3-deficient mice. Nature 407:1011–1015
Souslova V, Cesare P, Ding Y et al (2000) Warm-coding deficits and aberrant inflammatory pain in mice lacking P2X3 receptors. Nature 407:1015–1017
North RA (2003) The P2X3 subunit: a molecular target in pain therapeutics. Curr Opin Investig Drugs 4:833–840
Jarvis MF, Burgard EC, McGaraughty S et al (2002) A-317491, a novel potent and selective non-nucleotide antagonist of P2X3 and P2X2/3 receptors, reduces chronic inflammatory and neuropathic pain in the rat. Proc Natl Acad Sci U S A 99:17179–17184
Tsuda M, Shigemoto-Mogami Y, Koizumi S et al (2003) P2X4 receptors induced in spinal microglia gate tactile allodynia after nerve injury. Nature 424:778–783
Haughey NJ, Mattson MP (2003) Alzheimer’s amyloid beta-peptide enhances ATP/gap junction-mediated calcium-wave propagation in astrocytes. Neuromol Med 3:173–180
Exley C, Korchazhkina OV (2001) Promotion of formation of amyloid fibrils by aluminium adenosine triphosphate (AlATP). J Inorg Biochem 84:215–224
Rampe D, Wang L, Ringheim GE et al (2004) P2X7 receptor modulation of beta-amyloid- and LPS-induced cytokine secretion from human macrophages and microglia. J Neuroimmunol 147:56–61
Parvathenani LK, Tertyshnikova S, Greco CR et al (2003) P2X7 mediates superoxide production in primary microglia and is up-regulated in a transgenic mouse model of Alzheimer’s disease. J Biol Chem 278:13309–13317
Moore D, Chambers J, Waldvogel H et al (2000) Regional and cellular distribution of the P2Y1 purinergic receptor in the human brain: striking neuronal localisation. J Comp Neurol 421:374–384
Moore D, Iritani S, Chambers J et al (2000) Immunohistochemical localization of the P2Y1 purinergic receptor in Alzheimer’s disease. Neuroreport 11:3799–3803
Braun N, Zhu Y, Krieglstein J et al (1998) Upregulation of the enzyme chain hydrolyzing extracellular ATP after transient forebrain ischemia in the rat. J Neurosci 18:4891–4900
Melani A, Turchi D, Vannucchi MG et al (2005) ATP extracellular concentrations are increased in the rat striatum during in vivo ischemia. Neurochem Int 47:442–448
Phillis JW, O’Regan MH, Perkins LM et al (1993) Adenosine 5′-triphosphate release from the normoxic and hypoxic in vivo rat cerebral cortex. Neurosci Lett 151:94–96
Juranyi Z, Sperlagh B, Vizi ES et al (1999) Involvement of P2 purinoceptors and the nitric oxide pathway in [3H]purine outflow evoked by short-term hypoxia and hypoglycemia in rat hippocampal slices. Brain Res 823:183–190
Cavaliere F, Florenzano F, Amadio S et al (2003) Up-regulation of P2X2, P2X4 receptor and ischemic cell death: prevention by P2 antagonists. Neuroscience 120:85–98
Cavaliere F, Amadio S, Sancesario G et al (2004) Synaptic P2X7 and oxygen/glucose deprivation in organotypic hippocampal cultures. J Cereb Blood Flow Metab 24:392–398
Franke H, Günther A, Grosche J et al (2004) 2X7 receptor expression after ischemia in the cerebral cortex of rats. J Neuropathol Exp Neurol 63:686–699
Kim M, Jiang LH, Wilson HL et al (2001) Proteomic and functional evidence for a P2X7 receptor signaling complex. EMBO J 2:6347–6358
Moran-Jimenez MJ, Matute C et al (2000) Immunohistochemical localization of the P2Y1 purinergic receptor in neurons and glial cells of the central nervous system. Mol Brain Res 78:50–58
Viscomi MT, Florenzano F, Conversi D et al (2004) Axotomy dependent purinergic and nitrergic co-expression. Neuroscience 123:393–404
Tsuzuki K, Kondo E, Fukuoka T et al (2001) Differential regulation of P2X3 mRNA expression by peripheral nerve injury in intact and injured neurons in the rat sensory ganglia. Pain 91:351–360
Wang X, Arcuino G, Takano T et al (2004) P2X7 receptor inhibition improves recovery after spinal cord injury. Nat Med 10:821–827
Florenzano F, Viscomi MT, Cavaliere F et al (2002) Cerebellar lesion up-regulates P2X1 and P2X2 purinergic receptors in precerebellar nuclei. Neuroscience 115:425–434
Viscomi MT, Florenzano F, Amadio S (2005) Partial resistance of ataxin-2-containing olivary and pontine neurons to axotomy-induced degeneration. Brain Res Bull 66:212–221
Arthur DB, Akassoglou K, Insel PA et al (2005) P2Y2 receptor activates nerve growth factor/TrkA signaling to enhance neuronal differentiation. Proc Natl Acad Sci U S A 102:19138–19143
Peković S, Filipovic R, Subasic S et al (2005) Downregulation of glial scarring after brain injury: the effect of purine nucleoside analogue ribavirin. Ann N Y Acad Sci 1048:296–310
Pines A, Bivi N, Vascotto C et al (2006) Nucleotide receptors stimulation by extracellular ATP controls Hsp90 expression through APE1/Ref-1 in thyroid cancer cells: a novel tumorigenic pathway. J Cell Physiol 209:44–55
Gerasimovskaya EV, Ahmad S, White CW et al (2002) Extracellular ATP is an autocrine/paracrine regulator of hypoxia-induced adventitial fibroblast growth. Signaling through extracellular signal-regulated kinase-1/2 and the Egr-1 transcription factor. J Biol Chem 277:44638–44650
Murray AW (2004) Recycling the cell cycle: cyclins revisited. Cell 116:221–234
Morgan DA, Class R, Soslau G et al (1997) Cytokine-mediated erythroid maturation in megakaryoblastic human cell line HU-3. Exp Hematol 25:1378–1385
Sherr CJ, Roberts JM (2004) Living with or without cyclins and cyclin-dependent kinases. Genes Dev 18:2699–2711
Neary JT, Kang Y, Willoughby KA et al (2003) Activation of extracellular signal-regulated kinase by stretch-induced injury in astrocytes involves extracellular ATP and P2 purinergic receptors. J Neurosci 23:2348–2356
Budagian V, Bulanova E, Brovko L et al (2003) Signaling through P2X7 receptor in human T cells involves p56lck, MAP kinases, and transcription factors AP-1 and NF-kappa B. J Biol Chem 278:1549–1560
Ahmed-Choudhury J, Russell CL, Randhawa S et al (2003) Differential induction of nuclear factor-kappaB and activator protein-1 activity after CD40 ligation is associated with primary human hepatocyte apoptosis or intrahepatic endothelial cell proliferation. Mol Biol Cell 14:1334–1345
Krejci P, Bryja V, Pachernik J et al (2004) FGF2 inhibits proliferation and alters the cartilage-like phenotype of RCS cells. Exp Cell Res 297:152–164
Ahamed S, Foster JS, Bukovsky A et al (2001) Signal transduction through the Ras/Erk pathway is essential for the mycoestrogen zearalenone-induced cell-cycle progression in MCF-7 cells. Mol Carcinog 30:88–98
Cordova-Alarcon E, Centeno F, Reyes-Esparza J et al (2005) Effects of HRAS oncogene on cell cycle progression in a cervical cancer-derived cell line. Arch Med Res 36:311–316
Hideshima T, Nakamura N, Chauhan D et al (2001) Biologic sequelae of interleukin-6 induced PI3-K/Akt signaling in multiple myeloma. Oncogene 20:5991–6000
Glauser DA, Schlegel W (2007) Sequential actions of ERK1/2 on the AP-1 transcription factor allow temporal integration of metabolic signals in pancreatic (beta) cells. FASEB J (in press)
Chu TS, Wu MS, Wu KD et al (2007) Endothelin-1 activates MAPKs and modulates cell cycle proteins in OKP cells. J Formos Med Assoc 106:273–280
Migliaccio A, Castoria G, Di Domenico M et al (2002) Sex steroid hormones act as growth factors. J Steroid Biochem Mol Biol 83:31–35
Lents NH, Keenan SM, Bellone C et al (2002) Stimulation of the Raf/MEK/ERK cascade is necessary and sufficient for activation and Thr-160 phosphorylation of a nuclear-targeted CDK2. J Biol Chem 277:47469–47475
Reshetnikova G, Barkan R, Popov B et al (2000) Disruption of the actin cytoskeleton leads to inhibition of mitogen-induced cyclin E expression, Cdk2 phosphorylation, and nuclear accumulation of the retinoblastoma protein-related p107 protein. Exp Cell Res 259:35–53
Bhatt KV, Spofford LS, Aram G et al (2005) Adhesion control of cyclin D1 and p27Kip1 levels is deregulated in melanoma cells through BRAF-MEK-ERK signaling. Oncogene 24:3459–3471
Lefevre G, Calipel A, Mouriaux F et al (2003) Opposite long-term regulation of c-Myc and p27Kip1 through overactivation of Raf-1 and the MEK/ERK module in proliferating human choroidal melanoma cells. Oncogene 22:8813–8822
Delmas C, Manenti S, Boudjelal A et al (2001) The p42/p44 mitogen-activated protein kinase activation triggers p27Kip1 degradation independently of CDK2/cyclin E in NIH 3T3 cells. J Biol Chem 276:34958–34965
Zhang W, Lee JC, Kumar S et al (1999) ERK pathway mediates the activation of Cdk2 in IGF-1-induced proliferation of human osteosarcoma MG-63 cells. J Bone Miner Res 14:528–535
Boyer JL, Mohanram A, Camaioni E et al (1998) Competitive and selective antagonism of P2Y1 receptors by N6-methyl 2′-deoxyadenosine 3′,5′-bisphosphate. Br J Pharmacol 124:1–3
Baurand A, Gachet C (2003) The P2Y(1) receptor as a target for new antithrombotic drugs: a review of the P2Y(1) antagonist MRS-2179. Cardiovasc Drug Rev 21:67–76
Ulrich H, Trujillo CA, Nery AA et al (2006) DNA and RNA aptamers: from tools for basic research towards therapeutic applications. Comb Chem High Throughput Screen 9:619–632
Trujillo CA, Nery AA, Martins AH et al (2006) Inhibition mechanism of the recombinant rat P2X(2) receptor in glial cells by suramin and TNP-ATP. Biochemistry 45:224–233
Trujillo CA, Majumder P, Gonzalez FA et al (2007) Immobilized P2X(2) purinergic receptor stationary phase for chromatographic determination of pharmacological properties and drug screening. J Pharm Biomed Anal 44:701–710
Acknowledgments
H.U. and L.R.G.B. are grateful for grant support from FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo) and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico), Brazil. K.N.G.’s Ph.D. thesis is supported by fellowship from CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior). C.L.G.’s undergraduate research, C.A.T.’s and K.K.Y.’s Ph.D. theses and R.R.R.’s postdoctoral research are supported by fellowships from FAPESP. P.M.’s Ph.D. thesis was supported by fellowship from FAPESP.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License ( https://creativecommons.org/licenses/by-nc/2.0 ), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Majumder, P., Trujillo, C.A., Lopes, C.G. et al. New insights into purinergic receptor signaling in neuronal differentiation, neuroprotection, and brain disorders. Purinergic Signalling 3, 317–331 (2007). https://doi.org/10.1007/s11302-007-9074-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11302-007-9074-y