
Abstract
To provide new insights into microbial functions in the Kushiro Wetland, Japan, we measured vertical profiles of fluorescein diacetate hydrolysis (total microbial activity), β-glucosidase and xylosidase (organic matter decomposition), acid phosphatase (phosphate production) and potential denitrifying (denitrification) activities as microbial enzyme activities in soil to depths of approximately 1.5 m from two sites with different vegetation in November 2008 (winter) and August 2009 (summer). Active organic matter decomposition, phosphate production and denitrification were evident in shallow litter and peat layers, and total microbial activity was high. Almost no differences in decomposition and total microbial activity were observed between seasons, whereas phosphate production and denitrification were higher in summer. All activities were low in mid-depth volcanic-ash and clay layers because of low carbon, nitrogen and phosphorus levels. Surprisingly, the total microbial activity and decomposition in deep clay & peat and peat layers were the same as or higher than in shallow layers. However, denitrification was limited, probably because dissolved organic matter containing humic-like substances was unsuitable as a substrate. Moreover, total soil phosphorus levels, acid phosphatase activity and multiple linear regression analysis revealed that the soil in the Kushiro Wetland is likely P limited.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Wetlands are transitional ecosystems found between terrestrial and aquatic ecosystems that are subjected to loading of organic matter and nutrients from their catchment areas. Wetland soils generally consist of accreted peat and are rich in organic matter. The behavior of organic matter and nutrients in wetlands can therefore influence various ecosystems such as eutrophication and acidification in aquatic ecosystems or changes in greenhouse gases in the atmosphere. Various microbial communities found in wetland soil can play roles in the transformation and storage of organic matter and nutrients (D’Angelo and Reddy 1994; McLatchey and Reddy 1998; Prenger and Reddy 2004). Thus, wetland soil is the main site for microbial functions involving the transformation and storage of organic matter and nutrients (Martens 1995). However, the specific functions that occur in wetland soil are still unclear.
These microbial functions of wetland soils are poorly understood because they have rarely been investigated in deep soil. Indeed, most soil microbiology studies have focused on the top 25 cm of soil, where the densities of microorganisms are highest (Fierer et al. 2003). However, large numbers of microbes have been found well below the surface (Fritze et al. 2000; Blume et al. 2002; Fierer et al. 2003). For example, Fierer et al. (2003) reported that approximately 35 % of the total microbial biomass in the top 2 m of terrestrial soil was found at depths >25 cm. Accordingly, a substantial microbial presence is also expected in deep layers of wetland soils.
Wetland soil forms when the rate of organic matter production exceeds the rate at which organic matter is decomposed. Over time, the accreted soil in wetlands becomes very thick. Because groundwater in such wetland soils undergoes a complex migration and upwells infrequently, even deep soil is likely to be subjected to loading of organic matter and nutrients. Therefore, the microbial function of deep wetland soil is likely as important as that in shallow soil. However, there is little information available regarding the transformation and storage of organic matter and nutrients in deep wetland soils.
The freshwater Kushiro Wetland in northeastern Hokkaido Prefecture, Japan, which is the largest wetland in Japan, provides a variety of wetland habitats that support a diversity of wildlife. In 1980, the wetland was registered under the Ramsar Convention on Wetlands, and in 1987 it officially became a national park in Japan. However, there have recently been reports of increased sedimentation and nutrient loading into the Kushiro Wetland in response to changes in land use, such as an increase in agricultural land and timber harvesting, and river channelization (Nakamura et al. 2002, 2004; Ahn et al. 2008). Increased sedimentation and nutrient loading affect all living organisms in a wetland and can cause drastic changes in wetland structure and function.
Microbial function, such as the transformation and storage of organic matter and nutrients, is determined by the level of extracellular enzymes produced by microbes to facilitate degradation of organic matter and the resultant acquisition of limiting nutrients (Sinsabaugh and Foreman 2001; Hill et al. 2010). Assays of such microbial enzyme activities have become useful techniques for monitoring microbial activity and identifying the mechanisms that underlie microbial processes (Sinsabaugh et al. 2002). Glycosidases are linked to organic carbon decomposition, while esterases are associated with phosphorus acquisition and denitrifying enzymes are important to nitrogen removal. Therefore, we measured the activities of these enzymes as indicators of microbial function in the soil of Kushiro Wetland.
In this study, we examined the vertical profiles of (1) soil chemistry, (2) pore-water chemistry and (3) fluorescein diacetate hydrolysis (total microbial activity), β-glucosidase and xylosidase activity (organic matter decomposition), acid phosphatase activity (phosphate production) and potential denitrifying activity (denitrification) as microbial functions using soil collected to a depth of approximately 1.5 m from two sites with different vegetation in winter 2008 and summer 2009. Furthermore, we explored the usefulness of multiple linear regression analysis as a predictive tool to clarify the effects of soil and pore-water chemistry on these microbial enzyme activities. Specifically, we tested the following hypotheses: (1) microbial enzymes are vertically active, resulting in high microbial functions even in deep soil layers, and (2) microbial functions differ seasonally.
Sampling and methods
Study sites and sampling
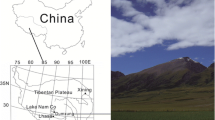
Two sites with different vegetation types in the Kushiro Wetland were selected (Fig. 1). Site 1 was dominated by Phragmites australis (Cav.) and Calamagrostis langsdorffii, while site 2 was dominated by C. langsdorffii. The two sites were about 20 m from an embankment and about 45 m apart. A sample of wetland soil approximately 1.5 m deep was collected from each site on 19 November 2008 (winter) and 11 August 2009 (summer) using a Handy Geoslicer sampling tool (Fukken Co. Ltd, Hiroshima, Japan). The instrument is made of stainless steel and comprises a sample box and a shutter (Takada et al. 2002). We observed the sedimentary facies in 5–6 layers in the soils collected (Table 1). The pH in each soil layer was measured on 11 August 2009 using a pH meter (IQ160, IQ Scientific Instruments, Carlsbad, CA, USA). To determine the vertical profiles of the microbial enzyme activities, soil samples were collected from each layer with a spatula while avoiding soil at the boundaries between layers and then analyzed for soil and pore-water chemistry. Each sample was transferred into a nylon bag and stored in a cooler for return to the laboratory. At the laboratory, soil samples were stored in a refrigerator at 4 °C until further processing.

Locations of Kushiro Wetland, northeastern Hokkaido, Japan (a), and sampling sites 1 and 2 in the wetland (b)
During summer sampling (9–11 August 2009), we monitored temporal changes in soil temperature at soil depths of 0, 50, 100, 150 and 200 cm at site 1 (TidbiT v2, Onset Computer Corporation, Bourne, MA, USA).
Soil and pore-water chemistry
To determine the soil water content (WC), about 5 g of wet soil was weighed (wet weight), dried at 105 °C to a constant weight, and weighed again. The dry soil was then homogenized and combusted at 450 °C for 12 h to determine the loss on ignition (IL) as a measure of organic matter content. The total carbon (TC) and total nitrogen (TN) contents of soils were determined using an NC analyzer (Sumigraph NC90A, Sumika Chemical Analysis Service, Ltd., Tokyo, Japan). Total phosphorous (TP) contents were determined using the method described by O’Halloran (1993). For this analysis, a 0.5-g sub-sample of dried soil was weighed in a porcelain crucible and combusted in a muffle furnace at 550 °C for 2 h. After cooling, the ash in the crucible was transferred to a 50-mL polyethylene tube with 25 mL of 0.5 M H2SO4. The tube was then placed on a shaker for 16 h, after which the phosphate concentration in the supernatant was determined by the colorimetric method (Murphy and Riley 1962). During winter, there was insufficient material in the litter layers at both sites to measure IL or TP.
Pore water was extracted from a subsample from each layer by centrifugation at 3000×g for 30 min. The supernatant was filtered (pore size: 0.2 μm) and analyzed for NH4 +, NO2 −, NO3 −, dissolved organic carbon (DOC) and the absorbance of humic-like substances. Concentrations of NH4 +, NO2 − and NO3 − were determined using an auto-analyzer (swAAt, BL TEC K.K., Tokyo, Japan), while concentrations of DOC were determined by using a total organic carbon (TOC) analyzer (TOC-500A, Shimadzu, Kyoto, Japan). Pore water could not be collected from the volcanic-ash layer at site 2 during winter; therefore, there were no data for NH4 +, NO2 −, NO3 − or DOC concentrations for this period. The pore water for measurement of humic-like substances was diluted by a factor of 3 with ultrapure water, after which the absorbance of humic-like substances was measured at 240 and 260 nm using a spectrophotometer (UVmini-1240, Shimadzu). We could not measure the absorbance in the upper peat, volcanic-ash or clay layers at either site during either season because there was insufficient sample material.
Measurement of microbial enzyme activities
Fluorescein diacetate hydrolysis (FH) activity was determined by spectrophotometric analysis (Schnürer and Rosswall 1982). About 4 g of wet soil and 0.25 mL of fluorescein diacetate solution (10 mg L−1) were added to 50 mL of sterile, 60 mM sodium phosphate buffer (pH 7.6), and the mixture was then incubated at 24 °C for 2 h on a rotary shaker. Two subsamples were taken from the mixture periodically to determine the absorbance at 490 nm. To follow the abiotic degradation of fluorescein diacetate, controls were incubated without soil and the increase in absorbance was determined. The FH activity was expressed as the increase in absorbance per gram of soil per hour.
The activities of β-glucosidase (GL), xylosidase (XY) and acid phosphatase (AP) were measured by fluorometric techniques (Freeman et al. 1995) using 4-methylumbelliferyl-β-d-glucopyranoside, 4-methylumbelliferyl-β-d-xylopyranoside and 4-methylumbelliferyl-phosphate, respectively, as substrates. About 0.4 g of wet soil, 2.0 mL of sodium phosphate buffer (pH 5.0) and 1.0 mL of the appropriate 3 mM substrate solution were added to a test tube and incubated for 2 h at 30 °C. At the end of the incubation period, 8 mL of ethanol was added to the test tubes to stop the reactions and the amount of 4-methylumbelliferone produced was determined from the fluorescence of the filtrate (excitation/emission [Ex/Em] wavelengths, 365/455 nm). For AP activity, modified universal buffer (MUB, pH 5.5) was used instead of sodium phosphate buffer.
Potential denitrifying (PD) activity was examined using the acetylene blockage technique (Yoshinari and Knowles 1976; Yoshinari et al. 1977). Briefly, about 3 g of wet soil and 20 mL of KNO3 solution (0.5 mg-N L−1) were added to a 70-mL glass vial. The vial was then sealed with a butyl-rubber stopper and aluminum cap, after which the air in the headspace was replaced with nitrogen gas. The vials were then incubated under anaerobic conditions for 5 h at 20 °C in the dark after C2H2 was added to a final pressure of 6 kPa. See Senga et al. (2001, 2006) for a more detailed description of these procedures.
The FH, GL, XY and AP activities were measured in duplicate during winter 2008 and in triplicate in summer 2009. The XY activity in the litter layer of site 1 in winter could not be measured due to a lack of residual sample. PD activity was measured in triplicate in winter and summer.
Statistical analyses
Statistical analyses was performed using the R 2.15.0 software (R Development Core Team 2012). Statistically significant correlations among microbial activities were determined by Pearson’s product–moment correlation. Multiple linear regression (MLR) was used to determine the effects of soil and pore-water chemistry parameters on the respective microbial enzyme activities. The Akaike Information Criterion (AIC), which balances the fit of a model against the number of parameters, was used to select the best fit model (Anderson and Burnham 2001). The equations with the lowest AIC determined by stepwise regression were accepted as the best equations for the data. The adjusted R 2 value reported is a modified R 2 that compensates for the number of terms in the equation. To minimize problems associated with multicollinearity (highly correlated variables), single correlation analysis was carried out prior to the regression analysis. This correlation analysis showed that TC was highly correlated with TN (R = 0.95, P < 0.001), and that it was also correlated with WC (R = 0.69, P < 0.001), IL (R = 0.79, P < 0.001), C/N (R = 0.69, P < 0.001) and C/P (R = 0.83, P < 0.001). Therefore, TC was not included in the regression analysis.
Results
The soil temperature at 0 cm at site 1 ranged from 18 °C to 21 °C from 9 to 11 August 2009, and was higher during daytime than at night, while soil temperature below 50 cm remained almost constant. The mean soil temperatures at soil depths of 50, 100, 150 and 200 cm were 13, 11, 9 and 8 °C, respectively.
There was no upper peat layer observed at site 1 during winter, but the other sedimentary facies and their approximate depths were almost the same at both sites in both seasons (Table 1). The shallow layers consisted almost entirely of un-decomposed litter or peat and therefore had high WC, IL, TC, TN and TP values. The layers from 5 to 26 cm contained volcanic ash derived from the eruption of Mt. Tarumae in 1667, while the layers from about 20–100 cm were predominantly clay or a mixture of clay and peat (hereafter referred to as clay & peat). The clay may have come from the nearby river during periods of overflow. WC, IL, TC, TN and TP were low in the clay layers. The deepest layers consisted of peat, with increased WC, IL, TC, TN and TP compared with the clay and clay & peat layers. The C/N and C/P ratios were higher in the litter and peat layers than in the other layers. The pH in the volcanic-ash layers at both sites was slightly higher than in the other layers.
The concentrations of NH4 + in pore water increased with depth at sites 1 and 2 during winter 2008 (Fig. 2a) and summer 2009 (Fig. 2b). NH4 + concentrations were higher in summer than in winter in all layers. Peaks in NO2 − concentration were observed in the clay & peat layers of both sites during winter (Fig. 2c). During summer, the NO2 − concentrations were very low, but it was detected in almost all layers (Fig. 2d). There were high levels of NO3 − in the clay and clay & peat layers during winter (Fig. 2e), whereas concentrations were lower in summer, although it was detected in all layers of both sites (Fig. 2f).

Vertical profiles of pore-water concentrations of NH4 + in winter (a) and summer (b), NO2 − in winter (c) and summer (d), and NO3 − in winter (e) and summer (f) at sites 1 (gray bars) and 2 (white bars). The upper and lower labels “Peat” on the y-axis represent the shallow (approx. 5–20 cm) and deep (approx. 70–150 cm) peat layers, respectively. There are no winter data for the volcanic-ash layer at site 2. “Winter” and “summer” refer to the sampling dates of 19 November 2008 and 11 August 2009, respectively
DOC concentrations increased with depth at sites 1 and 2 during winter and summer (Fig. 3a, b). There were no large differences in DOC concentrations between sites or seasons.

Vertical profiles of dissolved organic carbon (DOC) concentrations in pore water at sites 1 (gray bars) and 2 (white bars) in winter (a) and summer (b). The upper and lower labels “Peat” on the y-axis represent the shallow (depth approx. 5–20 cm) and deep (approx. 70–150 cm) peat layers, respectively. There are no winter data for the volcanic-ash layer at site 2. “Winter” and “summer” refer to the sampling dates of 19 November 2008 and 11 August 2009, respectively
The absorbance of humic-like substances was greatest in the bottom peat layers at both sites in both seasons (Table 2). The absorbance values in the litter layers were lower in winter than in summer, whereas they were slightly higher during winter than summer in the bottom peat layers. We found no large differences in absorbance values between sites 1 and 2.
FH activities were lower in the clay layers, except at site 2 in winter (Fig. 4a, b). There were no large differences in FH activities between summer and winter. GL activity was detected in all layers at both sites during both seasons (Fig. 5a, b). No large differences in GL activities were observed between summer and winter, except for in the clay & peat and peat layers at site 2. Relatively high XY activities were also detected in all layers at both sites in both seasons (Fig. 6a, b). We found no large differences in XY activities between summer and winter. Surprisingly, FH, GL and XY activities in deep clay & peat and peat layers were the same as or higher than those in shallow layers. AP activity was detected in all layers at both sites in both seasons, with higher activities observed in the litter and peat layers (Fig. 7a, b). There was also high AP activity in the bottom peat layers of site 2 during summer. AP activities tended to be higher in summer than in winter. PD activities at both sites in both seasons were greatest in the litter layers, while they were almost undetectable in the clay layers (Fig. 8a, b). The bottom peat layers showed low levels of denitrifying activity. PD activities were greater in summer than in winter, except in the clay layers.

Vertical profiles of fluorescein diacetate hydrolysis (FH) activity at sites 1 (gray bars) and 2 (white bars) in winter (a) and summer (b). The upper and lower labels “Peat” on the y-axis represent the shallow (depth approx. 5–20 cm) and deep (approx. 70–150 cm) peat layers, respectively. The activity measurements were performed in duplicate in winter and in triplicate in summer. Error bars for summer data indicate the standard deviations of triplicate measurements. “Winter” and “summer” refer to the sampling dates of 19 November 2008 and 11 August 2009, respectively

Vertical profiles of β-glucosidase (GL) activity at sites 1 (gray bars) and 2 (white bars) in winter (a) and summer (b). The upper and lower labels “Peat” on the y-axis represent the shallow (approx. 5–20 cm) and deep (approx. 70–150 cm) peat layers, respectively. The activities were measured in duplicate in winter and in triplicate in summer. Error bars for summer data indicate the standard deviations of triplicate measurements. “Winter” and “summer” refer to the sampling dates of 19 November 2008 and 11 August 2009, respectively

Vertical profiles of xylosidase (XY) activity at sites 1 (gray bars) and 2 (white bars) in winter (a) and summer (b). The upper and lower labels “Peat” on the y-axis represent the shallow (depth approx. 5–20 cm) and deep (approx. 70–150 cm) peat layers, respectively. There are no winter data for the litter layer at site 1. The activities were measured in duplicate in winter and in triplicate in summer. Error bars for summer data indicate the standard deviations of triplicate measurements. “Winter” and “summer” refer to the sampling dates of 19 November 2008 and 11 August 2009, respectively

Vertical profiles of acid phosphatase (AP) activity at sites 1 (gray bars) and 2 (white bars) in winter (a) and summer (b). The upper and lower labels “Peat” on the y-axis represent the shallow (depth approx. 5–20 cm) and deep (approx. 70–150 cm) peat layers, respectively. The activities were measured in duplicate in winter and in triplicate in summer. Error bars for summer data indicate the standard deviations of triplicate measurements. “Winter” and “summer” refer to the sampling dates of 19 November 2008 and 11 August 2009, respectively

Vertical profiles of potential denitrifying (PD) activity at sites 1 (gray bars) and 2 (white bars) in winter (a) and summer (b). The upper and lower labels “Peat” on the y-axis represent the shallow (depth approx. 5–20 cm) and deep (approx. 70–150 cm) peat layers, respectively. Error bars for summer data indicate the standard deviations of triplicate measurements. “Winter” and “summer” refer to the sampling dates of 19 November 2008 and 11 August 2009, respectively
There were strong positive correlations between GL and XY activities, and between AP and PD activities (Table 3), while there were weaker positive correlations between FH and AP, XY and PD, and XY and AP activities. Best subsets regression indicated that the best MLR model to explain GL, XY, AP and PD activities included parameters for the soil and pore-water chemistry (Table 4). FH activity only showed a positive correlation with TP.
Discussion
The vertical profiles of soil microbial enzyme activities (FH, GL, XY, AP and PD) in the Kushiro Wetland provided substantial information. In the surface wetland soil there is active organic matter decomposition and nutrient cycling through microbial processes (Hakulinen et al. 2005; Senga et al. 2011). In this study, we found relatively high FH, GL, XY, AP and PD activities (Figs. 4, 5, 6, 7, 8, respectively) in the shallow litter and peat layers. However, these microbial enzyme activities were also considerable in the deep clay & peat and peat layers, indicating that they warrant attention.
Fluorescein diacetate hydrolysis has been used to estimate total microbial activity in soil and litter (Schnürer and Rosswall 1982). MLR analysis showed a positive correlation between FH activity and TP (Table 4). Fluorescein diacetate is hydrolyzed by nonspecific esterases, including phosphatase (Meyer-Reil 1991). We found a weak, but significant positive correlation between FH and AP activities (Table 3), indicating that phosphate production may be an important process of total microbial activity in the Kushiro Wetland.
Beta-glucosidase and β-xylosidases decompose the cellulose and hemicellulose of plant litter, producing soluble saccharides such as glucose and xylose. Therefore, GL and XY activities are linked to the process of organic matter decomposition. GL and XY activities in all layers at both sites in both seasons were relatively high (Figs. 5, 6). In the deep clay & peat and peat layers there were substantial GL and XY activities, suggesting that organic matter was being actively decomposed by microbes in deep soil layers as well as in surface soil layers. In addition, there were only small differences between winter and summer GL and XY activities; thus, active and constant decomposition can be expected in the wetland soil throughout the year. These findings were supported by the FH activity observed in all layers during both seasons.
Almost all of the phosphorus was likely in the form of organic phosphorus because there was a positive correlation between TP and IL (R = 0.54, P = 1.10 × 10−2). These results show that the shallow and deep soil was rich in organic phosphorus as a source for the regeneration of inorganic phosphorus, but that inorganic phosphorus was limited. AP activity was detected in all layers at both sites in both seasons (Fig. 7a, b). Exogenous phosphatases are central to the regeneration of inorganic phosphorus, particularly in P-limited aquatic ecosystems (Prenger and Reddy 2004; Hill et al. 2006, 2010). An increase in AP activity in response to declining or limiting inorganic phosphorus has been demonstrated in numerous aquatic environments (Kang and Freeman 1999; Wright and Reddy 2001; Hill et al. 2006), and it is likely that the soil in the Kushiro Wetland is also P limited.
AP activities tended to be higher in the litter and upper peat layers, and were enhanced in summer (Fig. 7a, b). During summer, the plant uptake of inorganic phosphorus would increase; thus, it is assumed that the litter and upper peat layers in the Kushiro Wetland are under strong P limitation, especially in summer. Phosphatases are not only produced by microbes, but also by higher plants (Tarafdar and Claassen 1988); accordingly, the roots of such higher plants may be another source of the high AP activity in the surface layers during the summer growing season.
In general, rapid denitrification was observed in the surface soils (White and Reddy 2000, Dodla et al. 2008). In this study, PD activities in the surface litter and peat layers were also higher than those in the bottom clay & peat and peat layers (Fig. 8a, b). In addition, PD activities in the surface litter and peat layers at sites 1 and 2 were higher in summer than in winter, suggesting that denitrification in the surface layers of the Kushiro Wetland was enhanced by warming in summer. Another possible source of high PD activity in the surface layers during summer may be the labile dissolved organic matter and oxygen released through plant roots. PD activities were positively correlated with AP activities (Table 3). As described above, high AP activity in the surface layers in summer would include the activity of phosphatases from plant roots. Live plant roots exude a range of dissolved organic compounds, including sugars, organic acids and amino acids (Lambers and Poorter 1992). These could enhance denitrification because denitrifying bacteria are heterotrophs (Karjalainen et al. 2001). Additionally, aeration of soil by some aquatic plants is known to support strengthened nitrification–denitrification (Risgaard-Petersen and Jensen 1997; Karjalainen et al. 2001). Nitrification occurs under oxic and suboxic conditions (Jørgensen et al. 1984; Senga et al. 2002) and supplies NO3 − from NH4 +. There should be active denitrification in the surface soil, where the anoxic environment is adjacent to the oxic environment (Horiuchi and Tsuchiya 1999). In the upper layers at sites 1 and 2, summer NH4 + concentrations would be low because of NH4 + consumption through nitrification or plant uptake (Fig. 2b), whereas low, but measurable NO3 − was used in active denitrification or plant uptake (Fig. 2f).
We previously reported that NO3 − and DOC accumulated in the deeper soil during the winter of 2008 because most of the DOM was comprised of humic-like substances that cannot be used as a substrate for denitrification, the result being a suppression of denitrifying activity (Senga et al. 2011). In the present study, we found the same trends in absorbance of humic-like substances; specifically, low PD activities, accumulations of NO3 − and DOC, and high absorbance values in the clay & peat and peat layers (Figs. 2f, 3b; Table 2). Regardless of the season, there were larger amounts of humic-like substances in the clay & peat and peat layers than in the litter. As described above, heterotrophic denitrification requires organic matter as an electron donor; therefore, DOM can limit the denitrification rate (Payne, 1973).
We expected the activities of GL and XY to be important for denitrification because β-glucosidase and β-xylosidases decompose the cellulose and hemicellulose of plant litter, producing soluble saccharides such as glucose and xylose. We found a weak, but significant relationship between PD and XY activities, but no relationship between PD and GL activities, despite the strong, significant relationship between GL and XY activities (Table 3). These results may indicate that denitrifying bacteria preferentially use DOM through hydrolysis to xylosides rather than to glycosides.
MLR analyses suggested that the variability in GL, XY, AP and PD activities could be explained by using soil and pore-water chemistry parameters as predictive variables (Table 4). These findings show that microbial enzymatic processes are complicated and could be limited by many environmental factors. The levels of these activities were related to factors associated with soil organic matter; namely, IL, TN, TP, C/N, or C/P. Additionally, the activities of XY, AP and PD were inversely related to DOC concentrations. These relationships could potentially explain the significance of organic matter for decomposition, phosphate production and denitrification. Moreover, TP exhibited a positive correlation with all activities except that of GL, whereas TN was negatively correlated with GL and PD. These results suggest that the soil in the Kushiro Wetland was P limited. As mentioned above, TP (Table 1) and AP (Fig. 7) activity also showed evidence of P limitation.
Many researchers have reported that microbial enzyme activity varies according to vegetation (e.g., Prenger and Reddy 2004; Lü et al. 2013); therefore, we expected vegetative litter to have a distinct effect on microbial enzyme activity. Although we often observed differences among the activities at the two sites (e.g., FH activity in the shallow layers or GL activity in the deep layers during winter), it was hard to determine if these differences were influenced by vegetation. Unfortunately, we only sampled one site in areas dominated by each vegetation type; therefore, additional studies are needed to address the influence of vegetation.
In support of our hypothesis, there was considerable microbial function in the deep soil (to 1.5-m depth) that cannot be ignored. Accordingly, evaluation of the microbial function in a wetland it is necessary to estimate microbial activity in both the shallow and deep layers. In fact, it is not known whether the inclusion of soil to a depth of 1.5 m was sufficient to clarify the microbial function in the Kushiro Wetland; therefore, future studies are needed to assess the activities of microbial enzymes at greater depths.
References
Ahn YS, Nakamura F, Mizugaki S (2008) Hydrology, suspended sediment dynamics and nutrient loading in Lake Takkobu, a degrading lake ecosystem in Kushiro Mire, northern Japan. Environ Monit Assess 145:267–281
Anderson D, Burnham K (2001) Commentary on models in ecology. Bull Ecol Soc Am 82:160–161
Blume E, Bischoff M, Reichert JM, Moorman T, Konopka A, Turco RF (2002) Surface and subsurface microbial biomass, community structure and metabolic activity as a function of soil depth and season. Appl Soil Ecol 592:1–11
D’Angelo EM, Reddy KR (1994) Diagenesis of organic matter in a wetland receiving hypereutrophic lake water: II. Role of inorganic electron acceptors in nutrient release. J Environ Qual 23:937–943
Dodla SK, Wang JJ, DeLaune RD, Cook RL (2008) Denitrification potential and its relation to organic carbon quality in three coastal wetland soils. Sci Tot Environ 407:471–480
Fierer N, Schimel JP, Holden PA (2003) Variations in microbial community composition through two soil depth profiles. Soil Biol Biochem 35:167–176
Freeman C, Liska G, Ostle NJ, Jones SE, Lock MA (1995) The use of fluorogenic substrates for measuring enzyme activity in peatlands. Plant Soil 175:147–152
Fritze H, Pietikainen J, Pennanen T (2000) Distribution of microbial biomass and phospholipid fatty acids in Podzol profiles under coniferous forest. Eur J Soil Sci 51:565–573
Hakulinen R, Kähkönen MA, Salkinoja-Salonen M (2005) Vertical distribution of sediment enzyme activities involved in the cycling of carbon, nitrogen, phosphorous and sulphur in three boreal rural lakes. Water Res 39:2319–2326
Hill BH, Elonen CM, Jicha TM, Cotter AM, Trebitz AS, Danz NP (2006) Sediment microbial enzyme activity as an indicator of nutrient limitation in Great Lakes coastal wetland. Freshw Biol 51:1670–1683
Hill BH, Elonen CM, Jicha TM, Bolgrien DW, Moffett MF (2010) Sediment microbial enzyme activity as an indicator of nutrient limitation in the great rivers of the upper Mississippi River basin. Biogeochem 97:195–209
Horiuchi E, Tsuchiya T (1999) Effects of emergent plants on gas emission from soil surface in water. Jpn J Limnol 60:291–297
Jørgensen KS, Jensen HB, Sørensen J (1984) Nitrous oxide production from nitrification and denitrification in marine sediment at low oxygen concentrations. Can J Microbiol 30:1073–1078
Kang H, Freeman C (1999) Phosphatase and arylsulphatase activities in wetland soils: annual variation and controlling factors. Soil Biol Biochem 31:449–454
Karjalainen H, Stefansdottir G, Tuominen L, Kairesalo T (2001) Do submersed plants enhance microbial activity in sediment? Aquat Bot 69:1–13
Lambers H, Poorter H (1992) Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences. Adv Ecol Res 23:187–261
Lü Y, Wang C, Jia Y, Du J, Ma X, Wang W, Pu G, Tian X (2013) Responses of soil microbial biomass and enzymatic activities to different forms of organic nitrogen deposition in the subtropical forests in East China. Ecol Res 28:447–457
Martens R (1995) Current methods for measuring microbial biomass C in soil: potentials and limitations. Biol Fertil Soils 19:87–99
McLatchey GP, Reddy KR (1998) Regulation of organic matter decomposition and nutrient release in a wetland soil. J Environ Qual 27:1268–1274
Meyer-Reil LA (1991) Ecological aspects of enzymatic activity in marine sediments. In: Chróst RJ (ed) Microbial enzymes in aquatic environments. Springer, New York, pp 84–95
Murphy J, Riley JP (1962) A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta 27:31–36
Nakamura F, Jitsu M, Kameyama S, Mizugaki S (2002) Changes in riparian forest in the Kushiro Mire, Japan, associated with stream channelization. River Res Appl 18:65–79
Nakamura F, Kameyama S, Mizugaki S (2004) Rapid shrinkage of Kushiro Mire, the largest mire in Japan, due to increased sedimentation associated with land-use development in the catchment. Catena 55:213–229
O’Halloran IP (1993) Total and organic phosphorus. In: Carter MR (ed) Soil sampling and methods of analysis. Lewis Publishers, Boca Raton, pp 213–229
Payne WJ (1973) Reduction of nitrogenous oxides by microorganisms. Bacteriol Rev 37:409–452
Prenger JP, Reddy KR (2004) Microbial enzyme activities in a freshwater marsh after cessation of nutrient loading. Soil Sci Soc Am J 68:1796–1804
R Development Core Team (2012) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria, ISBN 3-900051-07-0, URL http://www.R-project.org
Risgaard-Petersen N, Jensen K (1997) Nitrification and denitrification in the rhizosphere of the aquatic macrophyte Lobelia dormanna L. Limnol Oceanogr 42:529–537
Schnürer J, Rosswall T (1982) Fluorescein diacetate hydrolysis as a measure of total microbial activity in soil and litter. Appl Environ Microbiol 43:1256–1261
Senga Y, Seike Y, Mochida K, Fujinaga K, Okumura M (2001) Nitrous oxide in brackish Lakes Shinji and Nakaumi, Japan. Limnology 2:129–136
Senga Y, Mochida K, Okamoto N, Fukumori R, Seike Y (2002) Nitrous oxide in brackish Lake Nakaumi, Japan II: the role of nitrification and denitrification in N2O accumulation. Limnology 3:21–27
Senga Y, Mochida K, Fukumori R, Okamoto N, Seike Y (2006) N2O accumulation in estuarine and coastal sediments: the influence of H2S on dissimilatory nitrate reduction. Est Coast Shelf Sci 67:231–238
Senga Y, Hiroki M, Nakamura Y, Watarai Y, Watanabe Y, Nohara S (2011) Vertical profiles of DIN, DOC, and microbial activities in the wetland soil of Kushiro Mire, northeastern Japan. Limnology 12:17–23
Sinsabaugh RL, Foreman CM (2001) Activity profiles of bacterioplankton in a eutrophic river. Freshw Biol 46:1239–1249
Sinsabaugh RL, Carreiro MM, Alvarez S (2002) Enzyme and microbial dynamics of litter decomposition. In: Burns RG, Dick RP (eds) Enzymes in the environment: activity ecology & applications. Marcel Dekker, New York, pp 249–265
Takada E, Nakata T, Miyagi T, Haraguchi T, Nishitani Y (2002) Handy geoslicer - new soil sampler for Quaternary geologist (in Japanese). Chishitu News 579:12–18
Tarafdar JC, Claassen N (1988) Organic phosphorus compounds as a phosphorus source for higher plants through the activity of phosphatases produced by plant roots and microorganisms. Biol Fertil Soils 5:308–312
White JR, Reddy KR (2000) The effects of phosphorus loading on organic nitrogen mineralization of soils and detritus along a nutrient gradient in the northern Everglades, Florida. Soil Sci Soc Am J 64:1525–1534
Wright AL, Reddy KR (2001) Heterotrophic microbial activity in northern Everglades wetland soils. Soil Sci Soc Am J 65:1856–1864
Yoshinari T, Knowles R (1976) Acetylene inhibition of nitrous oxide reduction by denitrifying bacteria. Biochem Biophys Res Commun 69:705–710
Yoshinari T, Hynes R, Knowles R (1977) Acetylene inhibition of nitrous oxide reduction and measurement of denitrification and nitrogen fixation in soil. Soil Biol Biochem 9:177–183
Acknowledgments
We thank G. Morimoto and Y. Osawa for their helpful suggestions regarding statistical analysis. This study was supported by the Pollution Control Research Fund of the Japanese Ministry of the Environment, and a Grant-in-Aid for Exploratory Research (No. 23710018) from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Senga, Y., Hiroki, M., Terui, S. et al. Variation in microbial function through soil depth profiles in the Kushiro Wetland, northeastern Hokkaido, Japan. Ecol Res 30, 563–572 (2015). https://doi.org/10.1007/s11284-015-1257-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11284-015-1257-3