
Abstract
The disruptive effects of tertiary species on otherwise positive pairwise species interactions (e.g. context-dependent parasitism in pollinator syndromes) is well-known. However, few—if any—studies have investigated how invasive plants affect interactions between facilitative plants and their native plant communities. Further, if tertiary invasive species can change interactions among native species from positive to negative, then a tertiary native should be capable of the same phenom for pairwise interactions between natives and invasives. Our previous research indicates invasive black mustard (Brassica nigra) changes interaction signs for otherwise positive species interactions between the dominant, native facilitator California buckwheat (Eriogonum fasciculatum) and its co-dominant beneficiary California sagebrush (Artemisia californica) in semi-arid California coastal sage scrub habitat. Here, E. fasciculatum and A. californica seedlings increased B. nigra shoot growth in pairwise species interactions in the greenhouse. However, in three-way species interactions, E. fasciculatum and A. californica together reduced B. nigra SLA, height, and reproductive potential while not increasing shoot DW. In three-way species interactions, B. nigra did not significantly reduce E. fasciculatum facilitation of A. californica. Also surprisingly, light competition with B. nigra resulted in an increase in A. californica height, which reduced the negative effects of A. californica light competition on shade-intolerant E. fasciculatum. In an additive field experiment, A. californica protected E. fasciculatum from facilitating germination and growth of B. nigra when water competition was minimized. Taken together, this study demonstrates the importance of studying species interactions between competitive, native perennials in the current ecological context of invaded ecosystems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Tertiary species can alter pairwise species interactions (e.g. Wooton 1994; Chamberlain et al. 2014), even in obligatory mutualisms, such as context-dependent parasitism observed in pollinator syndromes (see Palmer et al. 2003 for further explanation). Another positive but highly context-dependent species interaction is plant facilitation, in which interactions shift to competition as alleviated stressors weaken along spatial and temporal gradients (Bertness and Callaway 1994). However, the literature has yet to illuminate how non-native tertiary species affect pairwise species interactions in plant facilitation systems, despite the well-documented occurrences of facilitative plants recruiting harmful invasives (e.g. Rodríguez-Buriticá and Miriti 2009). Conversely, if a tertiary species can alter interactions between native species (Chamberlain et al. 2014), a tertiary native could theoretically change pairwise interaction signs between native and invasive plants, but to our knowledge, this has never been demonstrated.
Invasive plants fundamentally alter ecological processes, both directly through exclusion of native flora (D’Antonio and Vitousek 1992) and indirectly through changes to soil properties and disturbance regimes (Stylinski and Allen 1999). For example, in semi-arid California coastal sage scrub (CSS), fast-growing invasive annuals have competitively excluded nearly all native annual and perennial grasses and forbs (Bowler 2000). At the community level, the loss of native plant diversity degrades habitat quality for animals (Bowler 2000). At the ecosystem level, invasive annuals reduce overall plant productivity (Goldstein and Suding 2014) and increase wildfire frequency by senescing easily ignitable shoot tissues early in the growing season (Minnich and Dezzani 1998). The decrease in fire intervals degrades habitat heterogeneity by inhibiting native perennials from reestablishing and reaching late post-fire successional stages (Minnich and Dezzani 1998). As fires occur more frequently, long-lived shrubs and trees may never reach late growth stages. Therefore, species interactions among long-lived perennials and other shorter-lived species during early growth stages may become more determinant to the distribution of all plant species.
Our previous work indicates the CSS dominant native facilitatory perennial California buckwheat (Eriogonum fasciculatum) benefits from the competitive effects of its nearly exclusive beneficiary, the codominant native perennial California sagebrush (Artemisia californica) (Schlau et al. 2021). Greenhouse and field experiments suggest E. fasciculatum acidifies CSS’s calcareous soils and alleviates alkalinity stress for A. californica. Positive interactions begin as early as the seedling stage and result in nearly exclusive positive spatial associations maintained into late growth forms when canopies are > 1 m in diameter. These previous findings led to a question as to whether E. fasciculatum’s positive spatial association with A. californica conveys the benefits of complimentary recruitment inhibition of other plant species, especially harmful invasives. Eriogonum fasciculatum and A. californica co-dominate a majority of remaining CSS habitat patches (e.g. Eliason and Allen 1997). However, slower-growing, long-lived perennial seedlings compete poorly for light and water with faster-growing invasive annuals (e.g. Eliason and Allen 1997; Cione et al. 2002). Indeed, an exploratory study suggested the highly invasive, allelopathic annual forb black mustard (Brassica nigra) disrupts otherwise positive species interactions in a plant facilitation system with E. fasciculatum and A. californica at early growth stages. Co-occurring E. fasciculatum and A. californica in late growth stages (canopies > 1 m diameter) are expected to not facilitate—and even inhibit recruitment of B. nigra, but recruitment outcomes at the seedling-to-juvenile growth stages remains far more unclear and is the primary focus of the study presented here.
This study asked the following questions: (1) Does B. nigra affect interactions between E. fasciculatum and A. californica at the seedling-to-juvenile growth stages? (2) At late growth stages, does A. californica prevent E. fasciculatum from recruiting B. nigra? (3) At any growth stage, do E. fasciculatum and A. californica synergistically harm B. nigra? (4) Finally, in an investigation into mechanisms of facilitation, does reducing soil pH in A. californica monocultures mimic facilitative effects of E. fasciculatum on A. californica? The dynamics of invaded facilitative systems, or invasions of any positive interactions between native species that enhance primary productivity, biodiversity, resistance to invasion, and habitat quality, demand extensive investigation if biologists are to enhance biological communities’ potential to adapt to changes in water availability, temperature, wildfire severity, eutrophication, invasive species, and geographic range (Brooker 2006). Furthermore, as increasing temperatures and invasives increase the frequency of wildfires in various ecosystems, studying species interactions between newly establishing seedlings of competitive, native perennials can further inform long-term land management decisions. In other words, improving our understanding of facilitative plant systems in the Anthropocene requires studying facilitation within the greater ecological context of intensely invaded plant communities.
Methods
Study system
California coastal sage scrub (CSS) is a Mediterranean-type ecosystem that occupies < 10–15% of its pre-colonial range (Westman 1981). White flight, the resulting poorly managed urban sprawl, and soil N-loading (soil eutrophication), primarily from heavy diesel truck emissions and heavy construction equipment, continue to facilitate the establishment of non-native plants into increasingly isolated habitat patches (Talluto and Suding 2008). CaCO3 soil deposits (2600 μg Ca2+ g−1 soil; Westman 1981) from ancient seashells uplifted with marine sediment produce multifaceted nutrient stress for local plant communities. As a mean 250–450 mm of annual precipitation falls during short winter rainy seasons, calcareous soils “harden” soil water up to pH 8.2 (Irvine Ranch Water District Comprehensive Financial Annual Report 2018). As CaCO3 disassociates, Ca2+ ions bond to PO43− and NO3−, making them unavailable to plants (Strom et al. 2005). By pulling H3O+ out of the rhizosphere, CO32− ions degrade proton gradients that roots establish to uptake nutrients (e.g. Fuglsang et al. 2007).
Target species
Eriogonum fasciculatum is a long-lived (> 25 year) perennial, facultatively evergreen and shade-intolerant shrub. Eriogonum fasciculatum facilitates A. californica via acidification of alkaline soils (presumably via mobilize of nutrients; Schlau et al. 2021), but outcompetes most other plant species for water with woody roots that spread > 3 m laterally (Kirkpatrick et al. 1977) and up to 2.5 m deep (Kummerow et al. 1977; Hellmers et al. 1995).
Co-dominant A. californica similarly competitively excludes most other species (Kirkpatrick et al. 1977). Terpene leachates inhibit germination and growth of several species (Halligan 1973, 1975; Karban 2007). Fibrous, shallow roots quickly absorb soil water from morning dew and offseason rain events to outcompete most other plants for water (Gray 1982). Both A. californica and E. fasciculatum canopies exhibit lateral dominant growth forms > 1 m wide to outcompete heterospecific seedlings for light.
Brassica nigra was chosen as the tertiary invasive due to its dominance of disturbed habitat and high fecundity in CSS, as well as its potential to allelopathically alter facilitatory interactions between E. fasciculatum and A. californica. Unlike most Angiosperms, B. nigra does not form mutualistic interactions with mycorrhizae, and instead, releases fungicidal root exudates, which indirectly harms other plant species by preventing associations with mycorrhizae (Schreiner and Koide 1993). When B. nigra germinates or leaches isothiocyanate into soils, the forb directly inhibits germination and growth of at least some other plant species (Turk and Tawha 2003; Tawaha and Turk 2003; Oduor et al. 2020).
Greenhouse competition-facilitation experiment
To test how B. nigra seedlings affect pairwise species interactions between E. fasciculatum and A. californica seedlings, the three species were grown in 7 L pots in the UC Irvine greenhouse utilizing a full factorial design (n = 15 ea. treatment). An additional 15 replicates of A. californica monocultures with 36 g elemental sulfur soil amendments were used to mimic plant-soil feedback of E. fasciculatum and test a hypothesis that E. fasciculatum alleviates alkalinity stress for A. californica with acidic root exudates. As a control for plant-soil feedback effects on soil pH, 12 more replicates were filled with soil and watered, but contained no plants.
Genetically local E. fasciculatum and A. californica seeds were furnished by the Irvine Ranch Conservancy’s Irvine Ranch Natural Landmarks in Irvine, CA. Brassica nigra seeds were harvested from the UC Irvine CSS Ecological Preserve. All seeds germinated for greenhouse experiment were collected after the above average 2018–2019 winter rainy season. Plants were started in germination trays under misting benches, then replanted in 7 L pots following growth of first true leaves.
Each 7 L pot contained four plants. Pairwise species interactions were tested with two of each target species. Three-way species interactions were tested with two of the treated species and one each of the other two species. For example, in order to test the effects of B. nigra and A. californica on E. fasciculatum growth, two E. fasciculatum grew with one B. nigra and one A. californica.
The 7 L pots contained UCI native CSS soil mix (1.5 pumice:1 sand:1 redwood mulch:1 peat moss by volume). Previous research indicated E. fasciculatum facilitation of A. californica was conditional on alkaline soil (Schlau et al. 2021). Soil pH levels were recorded for all pots using 1:1 slurry method with a HANA field pH meter prior to planting but after conditioning the soil with local “hard” tap water (28–90 mg Ca2+⋅L−1; pH 7.8–8.2; IRWD 2018) for 8 days to raise the pH to a closer approximation of field conditions (mean pH 7.8) observed in previous experiments. All plants were continuously watered with “hard” tap water. Soil pH was remeasured again prior to destructive harvest for analysis of plant-soil feedback effects. All pot positions were rerandomized weekly.
To test how pairwise and three-way species interactions affected overall shoot growth after 89 days in the greenhouse, log response ratios (mean effect size) of mean shoot DW (g) with pooled standard deviation were analyzed. Log response ratios in pairwise species interactions were derived from the natural log of mean shoot DW for target species in pairwise species interaction treatments divided by the mean shoot DW of the target species in monoculture controls. Pooled standard deviations were used to calculate 95% confidence intervals (see Online Appendix for full description of calculations).
As indicated from a previous study (B. Schlau unpublished), E. fasciculatum was expected to facilitate shoot growth of the other target species. Brassica nigra was expected to decrease shoot DW of both E. fasciculatum and A. californica. Artemisia californica was expected to not affect E. fasciculatum shoot DW and decrease B. nigra shoot DW (see Fig. 1 for graph of proposed species interactions).

Conceptual model of hypothesized system of pairwise and three-way species interactions between E. fasciculatum (EF), A. californica (AC), and B. nigra (BC). Outer arrows represent pairwise species interactions. Internal radiating triangles represent effect of two species on the third. Interaction signs represent (+) positive/facilitation, (−) negative; competition, and (0) neutral species interactions
A primary objective of this study was to test the effects of a tertiary, invasive species on positive pairwise species interactions between codominant, native species. To test if the tertiary species affected species interactions between the two native species, three-way species interaction effect sizes were analyzed with expected values for additive three-way species interactions. To predict additive three-way species interaction effects, means of the respective pairwise species interaction effect log response ratios with 95% confidence intervals derived from pooled standard deviations (again, see Online Appendix for full description of equations). Standard deviations were determined with pairwise effect means. Deviation of observed from predicted three-way species interactions were considered significant, and therefore non-additive interactions, if the predicted effect’s 95% confidence interval lied outside the observed effect’s 95% confidence interval. Synergistic three-way species interactions were indicated by significantly stronger observed three-way species interaction effects than predicted. Antagonistic three-way species interactions were indicated by significantly weaker effects than predicted.
Log response ratios showed a significantly antagonistic three-way species interaction that weakened negative effects on E. fasciculatum shoot DW observed with E. fasciculatum in pairwise species interactions with B. nigra and A. californica (see “Results” section, Fig. 2). Thus, relationships of E. fasciculatum shoot DW to the shoot DWs of A. californica and B. nigra in pairwise species interactions as well as E. fasciculatum shoot DW versus the sum of A. californica and B. nigra shoot DWs in three-way species interactions were analyzed with linear regressions. Negative relationships in pairwise species interactions were interpreted as competition and positive relationships in three-way species interactions were interpreted as an interaction between A. californica and B. nigra that alleviated negative effects of B. nigra and A. californica on E. fasciculatum observed in pairwise species interactions.

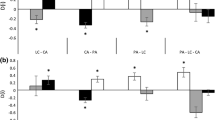
Mean effect size (response ratios) of target species (underlined species name) shoot DW in pair-wise species interactions (species name), three-way species interactions (× 3), expected three-way species interactions (Exp. X3), and reduced pH for A. californica (Low pH). Bars around means display 95% CI. Mean effect size is significantly different from zero when 95% CIs do not include zero
In order to investigate processes behind observed patterns in species interactions, shoot DW (covariable) was analyzed with ANCOVAs for relationships to specific leaf area (SLA), soil pH, and shoot height (dependent variables) in each species interaction treatment (independent variables). If an ANCOVA suggested a significant difference among slopes, linear regressions were used to test if any particular treatment effected a significant relationship between the dependent variable (e.g. SLA) and the covariable (shoot DW). For example, if pairwise species interactions with B. nigra resulted in an observed significant positive relationship with linear regression analysis between A. californica shoot height and shoot DW, the result was interpreted as A. californica growing taller due to interactions, such as light competition, with B. nigra.
Another of this study’s objectives was to test for interactions between E. fasciculatum and A. californica that increase resistance and resilience to invasion by B. nigra and potentially other invasives. Plants harm other plant species in ways that may not be detected from analyzing total shoot DW alone. Therefore, B. nigra leaf mass and inflorescence count (dependent variables) were compared among species treatments (independent variables) with ANOVAs and analyzed in conjunction with results from ANCOVA and linear regression analysis for indications of competition-driven tradeoffs among SLA, shoot height, and reproductive potential (i.e. inflorescence count). Aggregated results were interpreted with Westoby’s Leaf-Height-Seed Hypothesis (Westoby 1998).
Root DW and root:shoot ratios were not included in analysis due to entanglement of belowground tissues. The entangled roots that were impossible to separate while avoiding extensive root tissue damage.
At a desnity of two shrubs per 7 L pot in a previous experiment, E. fasciculatum facilitated A. californica shoot growth with no loss in its own shoot DW (Schlau et al. 2021). However, at four shrubs per treatment in the greenhouse here, A. californica appeared to shade E. fasciculatum seedlings. The shading led to an impromptu hypothesis that A. californica may harm its facilitator E. fasciculatum with light competition at this experiment’s planting density. As a test of this impromptu hypothesis, a linear regression was used to test for a relationship between E. fasciculatum shoot DW versus A. californica distal canopy width and using a prediction that A. californica distal canopy width would have a significantly inverse relationship to E. fasciculatum shoot DW.
Field recruitment facilitation-inhibition experiments
To test whether A. californica inhibits E. fasciculatum from recruiting B. nigra when the native shrubs are in late growth stages, an additive field experiment was conducted in the UC Irvine Coastal Sage Scrub Ecological Preserve. Long-lived, perennials that benefit from facilitation by a heterospecific tend to eventually outcompete their facilitator once the beneficiary establishes and is less vulnerable to previously alleviated stressors. Unlike other plant facilitation systems, a significant, positive spatial association between E. fasciculatum and A. californica persists into late growth stages when canopies are > 1 m diameter (Schlau et al. 2021). The persistence of mutual tolerance instead of competitive exclusion indicates positive or, at a minimum, neutral species interactions persist into later growth stages. This previous study led to a hypothesis that A. californica can protect E. fasciculatum from facilitating recruitment of harmful invasive species and E. fasciculatum facilitation of A. californica enhances the competitive effects of A. californica recruitment inhibition.
To tease apart facilitation from water competition, 30 genetically local B. nigra seeds were sown in 12 × 12 × 10cm buried pots of field soil and adjacent 12 cm2 plots of tilled bare ground. Topsoil was removed to a depth of 8–10 cm to minimize the possibility of B. nigra seedbanks from previous years. Ten replicates of each paired treatment (i.e. buriend pots and adjancet tilled ground) were positioned beneath south-facing canopy edges of E. fasciculatum, an adjacent A. californica, where the two species’ canopies crossed, and under shaded controls isolated by 0.5–1.0 m from the nearest plant (n = 80; n = 10 replicates per treatment). All shrub canopies were > 1 m diameter to better ensure each shrub exhibited the full metabolism of a perennial in late growth stages. The experiment was initially setup in the second half of December 2017. The germinations were counted × 3 a week between March 1st and April 20th 2018. In February and March of 2018, the study area experienced record low rainfall. Starting March 14th, seeds were watered with 200 mL of tap water × 3 a week until April 24th, which was the 14th day following the last observed germination. All B. nigra died within 2 weeks after germination, presumably due to water-stress. Field treatments were left in place for the following growing season and reseeded to 30 B. nigra in October before the 2018–2019 winter rainy season began. After the second rainy season, several B. nigra had established and destructively harvested after the first flowers closed. Brassica nigra reproductive potential was estimated with inflorescence count as a proxy. Root and shoot tissue were dried in ovens at 65 °C for 72 h. Shoot DW was analyzed as a metric of growth performance and productivity. Root:shoot ratios were analyzed for indications of alleviation of nutrient stress. Mean germinations (dependent variable) were compared with one-way and two-way fixed effect ANOVAs. A one-way ANOVA was utilized to compare germinations with canopy treatment as the independent variable. The two-way ANOVA was utilized to test for interactive effects between canopy treatment and tilled bare ground or buried pots (independent variables). The three dependent variables (shoot DW, root DW, and root:shoot ratios) were compared with one-way fixed-effects ANOVAs with canopy treatment as the independent variable. Residuals were not normally distributed for any dependent variables, so type 3 SS were used in analysis.
Statistical analyses were performed with R(v0.98.1062). Graphs were designed with MS Office Suite 2016.
Results
Log response ratios (mean effect sizes) showed E. fasciculatum increased A. californica mean shoot DW 101.9% in pairwise species interactions and an intermediant 85.3% in three-way species interactions compared to A. californica monoculture controls (Fig. 2). Brassica nigra did not significantly affect A. californica shoot DW in pairwise species interactions (Fig. 2).
Log response ratios of E. fasciculatum shoot DW indicated both A. californica and B. nigra significantly reduced E. fasciculatum shoot DW in pairwise species interactions (Fig. 2). In three-way species interactions, A. californica and B. nigra significantly decreased E. fasciculatum shoot DW, but was significantly less than the expected additive effect (Fig. 2).
Both E. fasciculatum and A. californica significantly increased B. nigra shoot DW in pairwise species interactions compared to monoculture controls (Fig. 2). However, three-way species interactions of A. californica and E. fasciculatum on B. nigra produced an antagonistic effect that did not affect B. nigra shoot DW compared to B. nigra monocultures (Fig. 2).
Finally, reducing soil pH with elemental sulfur significantly increased A. californica shoot DW by 40.0% (Fig. 2).
Artemisia californica Log-transformed shoot height to Log-transformed shoot DW showed a significant relationship between pairwise treatments with B. nigra and monoculture controls (ANCOVA: Est. Std. − 0.54828, p = 0.0391, Fig. S2, Table S1). Artemisia californica Log-transformed shoot height increased with Log-transformed shoot DW in pairwise species interactions with E. fasciculatum in a moderately significant relationship (Linear Regression: R2 = 0.22, p = 0.0765, Fig. S2, Table S1). Artemisia californica Log-transformed shoot height increased significantly with Log-transformed shoot DW in pairwise species interactions with B. nigra (R2 = 0.65, p = 0.0008, Fig. S2, Table S1) and in three-way species interactions (R2 = 0.48, p = 0.0063, Fig. S2, Table S1).
Artemisia californica distal canopy width significantly decreased E. fasciculatum shoot DW in pairwise species interactions (Linear Regression: R2 = 0.59, p = 0.00586; Fig. S1). Subsequent linear regression analysis of E. fasciculatum shoot DW versus the shoot DW of A. californica, B. nigra, and the sum of A. californica and B. nigra suggest competitive effects between A. californica and B. nigra weakened both species competitive effects on E. fasciculatum. In pairwise species interactions, increasing A. californica shoot DW decreased E. fasciculatum shoot DW significantly (R2 = 0.33, p = 0.0521; Fig. 3A), and B. nigra shoot DW had no significant effect on E. fasciculatum shoot DW (R2 = 0.06, p = 0.384, Fig. 3B). However, as the sum of A. californica and B. nigra shoot DW increased, E. fasciculatum shoot DW significantly increased in three-way species interactions (R2 = 0.41, p = 0.026, Fig. 3C).

E. fasciculatum shoot DW (g) versus a A. californica shoot DW (g), b B. nigra shoot DW (g), and c the sum of A. californica and B. nigra shoot DW (g). Linear regressions showed A. californica shoot DW had a moderately significant, inverse relationship to E. fasciculatum shoot DW (n = 12, p = 0.0521⬩), B. nigra shoot DW had no affect (n = 14, p = 0.384), and the sum of A. californica and B. nigra shoot DW had a significant, positive relationship to E. fasciculatum shoot DW (n = 12, p = 0.026*)
A series of ANCOVAs were used to tease apart the processes by which A. californica and E. fasciculatum did not increase B. nigra shoot DW in three-way species treatments. Log-transformed B. nigra SLA versus log-transformed B. nigra shoot DW had moderately significant, negative effects in Monocultures (Est. Std. = − 1.229685, p = 0.0575) and in three-way species interactions (Est. Std. = − 0.768761, p = 0.0783; Table S3). In pairwise species interactions with E. fasciculatum and A. californica, linear regression analysis (Fig. S4, Table S3) showed significant, inverse relationships (with E. fasciculatum: R2 = 0.28, p = 0.0308; and with A. californica: R2 = 0.39, p = 0.013). In three-way species interaction, the antagonistic, inverse relationship was moderately significant (R2 = 0.27, p = 0.0858). A second ANCOVA showed three-way species treatments affected the relationship of B. nigra Log-transformed shoot height versus shoot DW (Est. Std. = 0.29053, p = 0.00394; Fig. S5, Table S4). B. nigra shoot height increased with shoot DW in a significantly positive relationship in pairwise species interactions with A. californica (Linear Regression: R2 = 0.50, p = 0.00656) or in three-way species interactions (R2 = 0.34, p = 0.0343).
ANOVAs of three-way species interactions saw total B. nigra leaf mass was equal among treatments (\({F}_{\mathrm{3,50}}=1.366, p=0.265;\text{data not shown}\)). However, shoot height (\({F}_{\mathrm{3,50}}=6.203, p=0.0015;\text{data not shown}\)) and inflorescence count (\({F}_{\mathrm{3,50}}=2.895, p=0.0452;\text{data n}\text{ot shown}\)) varied significantly among species interaction treatments. Tukey HSD post hoc analysis of pairwise species interactions suggested A. californica and E. fasciculatum had no effect on B. nigra shoot height or inflorescence count. However, in three-way species interactions A. californica and E. fasciculatum together decreased B. nigra shoot height 33.1% (p = 0.0009, data not shown) and reduced inflorescence count 66.8% (p = 0.0452, data not shown) compared to B. nigra monocultures.
Additive field experiment
Among canopy treatments, B. nigra germination differed significantly in the field (\({F}_{\mathrm{3,72}}=8.381, p<0.0001;\) Fig. 4a) and between pots and tilled bare ground (\({F}_{\mathrm{1,72}}=17.473, p<0.0001;\) Fig. 4b), but with no interactive effect (TWO-WAY ANOVA: \({F}_{\mathrm{3,72}}=2.053, p=0.114;\) Fig. 4b). Shoot DW (ANOVA: \({F}_{\mathrm{3,24} }=9.972, p=0.0034\)), root DW (ANOVA: \({F}_{\mathrm{3,24} }=10.03, p=0.0002\)), and root:shoot ratios (ANOVA\(: {F}_{\mathrm{3,24} }=4.775, p=0.007\)) differed significantly among canopies (Fig. 5a–c, respectively). Tukey HSD post hoc analysis showed that germination, shoot growth, and root growth under A. californica, mixed canopies, and shaded bare ground treatments were equal, but E. fasciculatum increased mean B. nigra germination by 103.2% (p = 0.0043; Fig. 4a), shoot DW 433.3% (p = 0.0012; Fig. 5a), root DW 160.7% (p = 0.0112; Fig. 5b), and decreased root:shoot ratios by 44.5% (p = 0.0149; Fig. 5c) compared to B. nigra under shaded controls. Eriogonum fasciculatum recruitment of B. nigra only occurred in buried pots (Fig. 4b) as no B. nigra survived on tilled bare ground under E. fasciculatum.

a Mean B. nigra germinations under E. fasciculatum, A. californica, mixed canopies, and shaded controls. b Mean B. nigra germinations in buried pots (light gray with dots) and tilled bare ground (dark grey with waves) under E. fasciculatum, A. californica, mixed canopies, and shaded controls. Letters indicate significant differences from Tukey HSD post hoc analysis of permutation ANOVAs

a B. nigra mean shoot DW (g) (F3,24 = 9.972, p = 0.0034; b B. nigra mean root DW (g) (F3,24 = 10.03, p = 0.0002; and c B. nigra mean root:shoot ratios (F3,24 = 4.775, p = 0.007) under E. fasciculatum, A. californica, mixed canopies, and shaded controls. Letters indicate significant difference between means with Tukey HSD post hoc test on Permutation ANOVAs.
Discussion
This study tested (1) how a harmful invasive annual affects facilitative species interactions between two dominant, native perennials from seedlings to juveniles, and (2) can a competitive native perennial protect its facilitator from facilitating recruitment of a harmful invasive. While E. fasciculatum facilitation of B. nigra seedlings was expected in pairwise interactions, it was not predicted that A. californica would also facilitate B. nigra (Fig. 2). Once these observations are considered, the most surprising results were that in three-way species interactions among seedlings, the combined effects of E. fasciculatum and A. californica on B. nigra reversed the facilitation observed in pairwise interactions with B. nigra (Fig. 2). As seedlings, B. nigra does not significantly reduce E. fasciculatum facilitation of A. californica, and three-way species interactions lessen harmful competition with shade-intolerant E. fasciculatum for light. In three-way species interactions among seedlings, E. fasciculatum and A. californica also decrease B. nigra SLA, height, and inflorescence count. During later growth stages in the field, A. californica inhibits E. fasciculatum from recruiting B. nigra. Finally, results support the hypothesis that E. fasciculatum facilitates A. californica by acidification of alkaline soils. However, the presence of B. nigra inhibits E. fasciculatum soil acidification. Considered together, results presented here demonstrate significant effects of a tertiary invasive species on interactions between two ecologically important native plant species and demonstrate the potential for native perennials to increase resistance and resilience to biological invasion.
If pairwise interactions with A. californica and E. fasciculatum free B. nigra from intraspecific competition, B. nigra shoot DW could increase, but this scenario does not explain all the results. Three-way species interactions did not affect B. nigra shoot DW (Fig. 2), but SLA decreased as B. nigra shoot DW increased (Fig. S4, Table S3), and total leaf mass remained the same in all treatments (data not shown). When total leaf mass remains constant as SLA decreases, then a plant is growing its leaves thicker with less leaf area. Thicker leaves with reduced surface area is a common growth response of plants experiencing N-stress and/or water-stress (Violle et al. 2009). Field results below indicate E. fasciculatum is highly efficient at water competition. Further, B. nigra height (data not shown) and inflorescence (data not shown) also decreased in three-way species interactions compared to B. nigra monoculture controls. Three-way species interactions did not change B. nigra shoot DW. However, E. fasciculatum and A. californica together decreased B. nigra shoot height while significantly increasing the ratio of B. nigra shoot height to shoot DW (Fig. S5, Table S4). Between three-way species interactions and B. nigra monocultures, pairwise species interactions between B. nigra and either A. californica and E. fasciculatum were intermediate for SLA v. shoot DW in an ANCOVA as well as ANOVAs of B. nigra height and inflorescence count. Westoby’s Leaf-Height-Seed Strategy (1998) hypothesizes plants have finite resources to allocate to leaf growth, shoot growth, or reproduction. Resources allocated to shoot height per unit shoot DW would then be at the coast of leaf growth (reduced SLA per unit shoot DW) and reproduction (reduced inflorescence count). Scaled up to population level in the field, the effects of E. fasciculatum and A. californica together could reduce B. nigra fitness.
Indeed, field results suggest when E. fasciculatum cannot exclude B. nigra through water competition, A. californica can inhibit its codominant native facilitator from recruiting B. nigra. Eriogonum fasciculatum reduced germination (Fig. 4b) and growth of B. nigra on tilled bare ground treatments (Fig. 5a–c). The shallow lateral roots of E. fasciculatum could outcompete B. nigra seeds for limited water and prevent germination in CSS’s well-drained soils during the short rainy season. When sufficient precipitation allows B. nigra to establish, E. fasciculatum tap roots (up to 2.5 m soil depth) could outcompete B. nigra for water at greater soil depths during CSS’s long dry season. When buried pot treatments minimized water competition—or possibly when sufficient precipitation reduces the significance of E. fasciculatum water competition—field results suggest A. californica can prevent E. fasciculatum from facilitating germination and growth of B. nigra (Figs. 4b, 5a–c).
As nutrient availability increases, competition over N and P can be less deterministic than competition for light as fast-growing species produce more biomass than slower-growing heterospecifics (Bobbink et al. 2010). In pairwise interactions with Eriogonum fasciculatum, A. californica shoot DW more than doubled (Fig. 2), but competition and facilitation can occur concurrently (Bertness and Callaway 1994). The mobilization of nutrients otherwise unavailable in calcareous soils by E. fasciculatum would explain why A. californica shaded E. fasciculatum (Fig. S1) and reduced E. fasciculatum shoot DW 74.2% (Fig. 2) at densities of four shrubs per 7 L pot. At lower densities of two shrubs per 7 L pot—tested in a previous experiment—E. fasciculatum facilitates A. californica shoot growth and A. californica does not affect E. fasciculatum shoot DW (Schlau et al. 2021). Results here also align with a previous field study (Schlau et al. 2021), in which the two shrubs shared a positive spatial association in late growth stages at 1 m2 and 1 m scales, but not at 0 m (point-intercepts). In order for E. fasciculatum to maintain positive spatial associations with A. californica into late growth stages, the two shade-intolerant perennial species likely may have to partition space to access sunlight.
Negative effects on E. fasciculatum shoot DW were significantly antagonistic in three-way species interactions compared to expected additive effects from pairwise interactions (Fig. 2). Brassica nigra appears to trigger a plasticity growth response in A. californica to grow taller (Fig. S2). Eriogonum fasciculatum shoot DW’s significantly positive linear relationship with the sum of A. californica and B. nigra shoot DW in three-way interactions provides further evidence that interactions between A. californica and B. nigra diminish both species competitive effects on E. fasciculatum (Fig. 3C). When B. nigra (or possibly any vertical growth-dominant species) effects taller growth in A. californica, the shading effects of A. californica on E. fasciculatum seen in pairwise interactions (Fig. S1) may weaken.
Despite B. nigra competitive effects on E. fasciculatum in pair-wise interactions, in three-way interactions, B. nigra did not significantly reduce facilitation of A. californica by E. fasciculatum (Fig. 2) and mean soil pH was not reduced in pots with B. nigra, even when E. fasciculatum was also present (Fig. S5). This suggests the possibility that facilitation is not driven by E. fasciculatum acidifying soils as hypothesized here and previously shown (Schlau et al. 2021). Data presented here does not offer an explanation of alternate means of facilitation. However, B. nigra may inhibit soil acidification by E. fasciculatum after a critical growth period. Indeed, Rangkadilok et al. (2002) found that secondary metabolite production in B. nigra changes with the plant’s growth stages. The latter explanation could be answered with subsequent tests where soil pH is monitored at regular time intervals following planting. A third possibility is that soil acidification is the mechanism of facilitation in pairwise interactions between A. californica and E. fasciculatum, but not in three-way species interactions. Instead, if and when B. nigra triggers the growth plasticity response in A. californica to grow taller, A. californica may devote more resources to shoot tissues to be more competitive for light and increase its shoot DW (Fig. S2).
Another goal of this study was to test a hypothesis that E. fasciculatum acidifies CSS’s calcareous soils and alleviates soil alkalinity stress for A. californica. In pairwise species interactions, E. fasciculatum reduced soil pH and increased A. californica shoot DW compared to A. californica monocultures (Fig. S3, Table S2). In pairwise interactions with E. fasciculatum, the y-intercept and slope differed significantly, suggesting the relationship between soil pH and A. californica shoot DW was not driven by competition with E. fasciculatum. Coinciding with these results, utilizing elemental sulfur to reduce soil pH resulted in a weaker but still significant increase in A. californica shoot DW (Fig. 2). While soil pH did not have a significant relationship to A. californica shoot DW (data not shown), the slopes for A. californica shoot DW versus soil pH converged at pH 6.2 in both pairwise interactions with E. fasciculatum and in three-way species interactions (Fig. S3). What’s more, a subsequent polynomial regression of A. californica shoot DW versus soil pH in sulfur-treated pots has a significant relationship with an optimal soil pH at approximately 6.2 (data not shown). Mean soil pH in treatments with E. fasciculatum and A. californica was also 6.2 (Fig. S5). Further testing with a controlled gradient of soil pH is needed to adequately address the question of optimal soil pH for A. californica shoot DW as well as if E. fasciculatum reduces soil pH to that optimal range, but data here suggests E. fasciculatum reduces soil pH to an optimal range for A. californica growth and is the means of facilitation.
Conclusions
The experiments presented here emphasize the significance of studying positive pairwise species interactions within their greater ecological context. In the Anthropocene, this means considering species interactions within intensely invaded habitats. Most surprisingly, the tertiary species here—a fast-growing, allelopathic invasive (B. nigra)—appears to reduce competitive effects of a heterospecific beneficiary (A. californica) upon its facilitator (E. fasciculatum) during seedling-to-juvenile growth stages. During late growth stages, the competitive beneficiary protects the facilitative plant from recruiting the harmful invasive. The codominance of A. californica and E. fasciculatum once reduced native biodiversity in CSS (see collective works of Whittaker et al.), but today, competitive native species can enhance resistance and resilience to invasion (e.g. Maron and Marler 2007). If this is true of E. fasciculatum and A. californica, as results presented here suggest, then the niche of the two native species may shift from suppressor to protector of native biodiversity. In more applicable terms, restoration of disturbed CSS habitat may benefit from cultivated co-disperal of these native perennials, especially into re-occurring populations of B. nigra—and likely any invasive annual. Indeed, as fire intervals decrease and habitat becomes continually isolated, species interactions between seedlings of dominant perennials will likely become even more determinant to post-fire vegetative succession.
Data availability
All data was collected by Benjamin Schlau and data is available with the following link: https://docs.google.com/spreadsheets/d/1r_yA4ZtozkPcgaabkXk_MWSzEqGleE_g/edit?usp=sharing&ouid=109694860468006015664&rtpof=true&sd=true.
References
Bertness MD, Callaway R (1994) Positive interactions in communities. Trends Ecol Evol 9:191–193. https://doi.org/10.1016/0169-5347(94)90088-4
Bobbink R et al (2010) Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecol Appl 20:30–59. https://doi.org/10.1890/08-1140.1
Bowler P (2000) Ecological restoration of coastal sage scrub and its potential role in conservation plans. Environ Manag 26:85-S96. https://doi.org/10.1007/s002670010064
Brooker R (2006) Plant–plant interactions and environmental change. New Phytol 171:271–289. https://doi.org/10.1111/j.1469-8137.2006.01752.x
Chamberlain S, Bronstein J, Rudgers J (2014) How context dependent are species interactions? Ecol Lett 17:881–890. https://doi.org/10.1111/ele.12279
Cione N, Padgett P, Allen E (2002) Restoration of native shrubland impacted by exotic grasses, frequent fire, and nitrogen deposition in Southern California. Restor Ecol 10:376–384. https://doi.org/10.1046/j.1526-100X.2002.02038.x
D’Antonio C, Vitousek PM (1992) Biological invasions by exotic grasses, grass/fire cycle, and global change. Annu Rev Ecol Syst 23:63–87. https://doi.org/10.1146/annurev.es.23.110192.000431
Eliason S, Allen E (1997) Exotic grass competition in suppressing native shrubland re-establishment. Restor Ecol 5:245–255. https://doi.org/10.1046/j.1526-100X.1997.09729.x
Fuglsang A, Guo Y, Cuin T, Qiu Q, Song C et al. (2007) Arabidopsis protein kinase PKS5 inhibits the plasma membrane H+-ATPase by preventing interaction with 14-3-3 Protein. Plant Cell 19:1617–1634. https://doi.org/10.1105/tpc.105.035626
Goldstein L, Suding K (2014) Intra-annual rainfall regime shifts competitive interactions between coastal sage scrub and invasive grasses. Ecology 95:425–435. https://doi.org/10.1890/12-0651.1
Gray J (1982) Community structure and productivity in ceanothus chaparral and coastal sage scrub of southern California. Ecol Mon 52:415–434. https://doi.org/10.2307/2937353
Halligan J (1973) Bare areas associated with shrub stands in grassland: the case of Artemisia californica. Bioscience 23:429–432. https://doi.org/10.2307/1296544
Halligan J (1975) Toxic terpenes from Artemisia californica. Ecology 56:999–1003. https://doi.org/10.2307/1936312
Hellmers H (1955) Root systems of some chaparral plants in southern California. Ecol. 36: 667-678 doi: https://doi.org/10.2307/1931305
Irvine Ranch Water District (2018) Water Quality Report. Irvine Ranch Water District. Irvine, CA
Karban R (2007) Experimental clipping of sagebrush inhibits seed germination of neighbours. Ecol Lett 9:791–797. https://doi.org/10.1111/j.1461-0248.2007.01068.x
Kummerow J, Krause D, Jow W (1977) Root systems of chaparral shrubs. Oecologia 29:163–177. https://www.jstor.org/stable/4215454.
Maron J, Marler M (2007) Native plant diversity resists invasion at both low and high resource levels. Ecology 88:2651–2661. https://doi.org/10.1890/06-1993.1
Minnich RA, Dezzani RJ (1998) Historical decline of coastal sage scrub in the Riverside-Perris Plain, California. Western Birds. https://doi.org/10.1890/06-1993.1
Oduor A, Kleunen M, Stift M (2020) Allelopathic effects of native and invasive Brassica nigra do not support the novel-weapons hypothesis. Am J Bot 107:1106–1113. https://doi.org/10.1002/ajb2.1516
Palmer T, Stanton M, Young T (2003) Competition and coexistence: exploring mechanisms that restrict and maintain diversity within mutualist guilds. Am Nat 162:63–79. https://doi.org/10.1086/378682
Rangkadilok N, Nicolas ME, Bennett RN, Premier RR, Eagling DR, Paul W, Taylor J (2002) Developmental changes of sinigrin and glucoraphanin in three Brassica species (Brassica nigra, Brassica juncea and Brassica oleracea var. italica). Sci Hortic 96:11–26. https://doi.org/10.1016/S0304-4238(02)00118-8
Rodríguez-Buriticá S, Miriti M (2009) Biting the hand that feeds: the invasive grass Schismus barbatus (Poaceae) is facilitated by, but reduces establishment of, the native shrub Ambrosia dumosa (Asteraceae). J Veg Sci 20:241–250. https://doi.org/10.1111/j.1654-1103.2009.05588.x
Schlau B, Huxman TE, Mooney KA, Pratt JD (2021) Facilitation at early growth stages results in spatial associations and stable coexistence in late growth stages of two long-lived, dominant shrubs. Oikos 130:2182–2190. https://doi.org/10.1111/oik.08603
Schreiner P, Koide R (1993) Mustards, mustard oils and mycorrhizas. New Phytol 123:107–113. https://doi.org/10.1111/j.1469-8137.1993.tb04536.x
Stylinski C, Allen E (1999) Lack of native species recovery following severe exotic disturbance in southern California shrublands. J Appl Ecol 36:1–12. https://doi.org/10.1046/j.1365-2664.1999.00423.x
Strom L, Owen AG, Godbold DL, Jones DL (2005) Organic acid behaviour in a calcareous soil implications for rhizosphere nutrient cycling. Soil Biol Biochem 37:2046–2054. https://doi.org/10.1016/j.soilbio.2005.03.009
Talluto M, Suding K (2008) Historical change in coastal sage scrub in southern California, USA in relation to fire frequency and air pollution. Landsc Ecol 23:803–815. https://doi.org/10.1007/s10980-008-9238-3
Tawaha AM, Turk MA (2003) Allelopathic effect of black mustard (Brassica nigra) on wild barley (Hordeum spontaneum). J Agron Crop Sci 189:298–303. https://doi.org/10.1046/j.1439-037X.2003.00047.x
Turk MA, Tawha AM (2003) Allelopathic effect of black mustard (Brassica nigra) on germination and growth of wild oat (Avena fatua L.). Crop Prot 22:673–677. https://doi.org/10.1046/j.1439-037X.2003.00047.x
Violle C, Garnier E, Lecoeur J, Roument C, Pedeur C, Blanchard A, Navas ML (2009) Competition, traits and resource depletion in plant communities. Oecologia 160:747–755. https://doi.org/10.1007/s00442-009-1333-x
Westman W (1981) Diversity relations and succession in Californian coastal sage scrub. Ecology 62:439–455. https://doi.org/10.2307/1936680
Westoby M (1998) A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 199:213–227. https://doi.org/10.1023/A:1004327224729
Wooton J (1994) The nature and consequences of indirect effects in ecological communities. Annu Rev Ecol Syst 25:443–466. https://doi.org/10.1146/annurev.es.25.110194.002303
Funding
Open access funding provided by SCELC, Statewide California Electronic Library Consortium. Not Applicable.
Author information
Authors and Affiliations
Contributions
BS conceived, setup, and ran experiments, collected data, primary analyzer of data, wrote the manuscript and prepared figures. TH conceived application of ANCOVAs. TH and KM contributed mightily to experimental design and statistical analysis. TH, KM, and JP reviewed the manuscript and gave informative advice that lead to constructive revisions.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
Not Applicable.
Additional information
Communicated by Ling Zhang.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Schlau, B.M., Huxman, T.E., Mooney, K.A. et al. Three-way species interactions reverse the positive pairwise effects of two natives on an exotic invader. Plant Ecol 224, 349–359 (2023). https://doi.org/10.1007/s11258-023-01304-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-023-01304-6