
Abstract
This is our follow-up study carried out in an order to collect experimental evidence regarding the impact of heavy water (D2O) on the spontaneous oscillatory peptidization of l-proline (l-Pro). Our earlier studies have been focused on the two sulfur-containing proteinogenic α-amino acids, i.e., l-cysteine (l-Cys) and l-methionine (l-Met), and it seemed interesting to assess the effect induced by D2O on one more proteinogenic α-amino acid, i.e., l-Pro. It needs to be added that unlike l-Met, but similar to l-Cys, the oscillatory peptidization of l-Pro dissolved in the organic-aqueous solvent characterizes with the circadian rhythm. As analytical techniques, we used high-performance liquid chromatography with the evaporative light-scattering detection (HPLC-ELSD), mass spectrometry (MS), scanning electron microscopy (SEM), and turbidimetry. The obtained results can in certain sense be viewed as analogous to those earlier reported for l-Cys and l-Met by demonstrating that heavy water considerably hampers the oscillatory peptidization of l-Pro. However, an unexpected observation was also made than unlike the cases with l-Cys and l-Met, the observed hampering effect of D2O on the oscillatory peptidization of l-Pro is not monotonously dependent on the concentration of D2O in the system, but it is the strongest pronounced for 10% (v/v) D2O in the employed binary methanol–water solvent (with the investigated proportions of D2O in this solvent changing from 0 to 30%). Although we have no rational explanation for this striking effect, we believe that it should not pass unnoticed and therefore it is emphasized in this study. Maybe this firm quantitative result will prove an inspiration for future researchers interested in getting a deeper insight into the role of D2O in life processes, and more specifically in the kinetic and the mechanistic aspects thereof.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The impact of D2O on living organisms is of a considerable interest for biology and life sciences in general, both due to its documented lethal effects on higher organisms and particularly on those of the mammals [1,2,3,4,5], and to its possible curative potential through hampering proliferation of cancer cells [6,7,8,9,10,11]. This hampering effects and induction of apoptosis could be of a cytotoxic and/or a cytostatic nature, although neither well defined molecular targets of D2O, nor detailed mechanisms of such processes are clear. One reason is that no extensive studies have been so far undertaken to make us better understand the molecular level of such interactions, although certain researchers speculate about possible interactions between D2O and DNA of the investigated species, inhibiting the protein synthesis (e.g., [12,13,14]).
Our own research interests have been focused for over a decade now on spontaneous behavior of proteinogenic α-amino acids in the abiotic aqueous liquid systems and we were the first ones to report on spontaneous oscillatory chiral inversion [15,16,17,18] and spontaneous oscillatory peptidization [19,20,21,22,23,24,25] of a number of amino acids. Based on the results of these studies and aiming to gain deeper understanding of molecular mechanisms governing the aforementioned oscillatory phenomena, we decided to scrutinize an impact of heavy water (D2O) upon them. The results of our preliminary investigations were published in [26] and they remain in agreement with the results of the other research groups examining an impact of heavy water upon the two best known oscillatory reactions, i.e., the Belousov–Zhabotinsky (BZ) [27,28,29] and the Bray–Liebhafsky (BL) reaction [30, 31]. The most up-to-the-date information on the impact of D2O on the BZ reaction provided in paper [29] points out to an increased induction period in a cerium-catalyzed BZ reaction, in pace with progressive substitution of the deuterated reactants. Moreover, a delayed onset and reduced amplitude of oscillations were also observed. In earlier papers [30, 31], the authors reported on a considerable perturbation of the mechanism and kinetics of the BL reaction, caused by replacement of bulk H2O as a reaction medium with D2O. Thus it seems evident that heavy water perturbs both kinetics and mechanism of the oscillatory reactions with an evident slow-down of these processes.
Oscillatory chiral inversion and oscillatory peptidization of proteinogenic α-amino acids has been observed by us in abiotic systems only and it remains unknown, if such processes can occur in living organisms as well. Such possibility can be anticipated keeping in mind that all biological processes take place in an aqueous environment. If so, the question arises as to a possible impact of D2O on α-amino acids and on spontaneously formed oligopeptides, which might be competitive to or exerted in the parallel with interactions between D2O and DNA. Although our research does not aspire to providing an answer regarding an impact of D2O on the processes running in living organisms, we decided to undertake a systematic investigation of its impact on spontaneous peptidization of selected endogenous and exogenous α-amino acids in the abiotic aqueous media. In our earlier papers [32, 33], we presented the results of such investigations with two proteinogenic and sulfur atom containing α-amino acids, i.e., l-cysteine (l-Cys) and l-methionine (l-Met), respectively. These investigations were carried out with use of high-performance liquid chromatography with the evaporative light scattering detection (HPLC-ELSD), scanning electron microscopy (SEM), mass spectrometry (MS) and turbidimetry. An unequivocal outcome of these studies pointed out to hampering of spontaneous oscillatory peptidization with l-Cys and l-Met by heavy water.
This is our follow-up study carried out in an order to collect experimental evidence regarding an impact of heavy water (D2O) on spontaneous oscillatory peptidization of l-proline (l-Pro), a very important endogenous α-amino acid playing many vital roles in human and animal organisms. For example, it is essential for the synthesis of proteins and its deficiency can result in lowered levels of collagen, problems with wound healing, or an improper structure of blood arteries [34]. On the other hand, the perturbed metabolism of l-Pro can result in hereditary disorders characterized by an excessive level of l-Pro in the blood, known as hyperprolinemia type I (due to the deficiency of the enzyme proline oxidase) or type II (due to its complete lack) [35, 36]. Based on clinical results, certain researchers relate hyperprolinemia to schizophrenia [37, 38].
Experimental
Reagents and samples
l-Pro was of analytical purity, purchased from Sigma-Aldrich (St Louis, MO, USA; cat. # P0380). Methanol (MeOH) was of HPLC purity (Merck KGaA, Darmstadt, Germany). Heavy water (D2O) was acquired from the Cambridge Isotopic Laboratories (Andover, MA, USA; 99% purity), and water (H2O) was de-ionized and double distilled in our laboratory by means of the Elix Advantage model Millipore System.
The l-Pro sample prepared for the HPLC-ELSD experiment was dissolved at a concentration of 1 mg mL−1 (i.e., 8.69 × 10−3 mol L−1) in MeOH + H2O, 70:30 (v/v). The chromatographic measurements of the concentration changes of the monomeric l-Pro were carried out in the 17-min intervals for the period of 65 h at 21.0 ± 0.5 °C.
All the l-Pro solutions used for mass spectrometry (MS), scanning electron microscopy (SEM) and turbidimetry were prepared at a concentration of 1 mg mL−1 in the binary liquid mixtures MeOH + X, 70:30 (v/v), where X: the binary mixture of H2O + D2O in the changing volume proportions: 30:0, 25:5, 20:10, 10:20 and 0:30. The mass spectrometric, scanning electron microscopic and turbidimetric measurements were also carried out for the l-Pro sample prepared at a concentration of 1 mg mL−1 in pure heavy water (D2O).
High-performance liquid chromatography with evaporative light scattering detection (HPLC-ELSD)
High-performance liquid chromatography with evaporative light scattering detection (HPLC-ELSD) was employed to separate the oligopeptides from the monomeric amino acid which is l-Pro. The analysis of the freshly prepared L-Pro solution using a Varian model 920 liquid chromatograph (Varian, Harbor City, CA, USA) equipped with a Varian 900-LC model autosampler, a gradient pump, a Varian 380-LC model ELSD detector, Pursuit 5C18 (5 μm particle size) column (250 mm × 4.6 mm i.d.; Varian; cat. no. A3000250C046), and a Galaxie software. The chromatographic column was thermostated at 30 °C in a Varian ProStar 510 model column oven. The chromatographic analyses were carried out for the 20 μL sample aliquots, using methanol–water (80:20, v/v) as mobile phase at the flow rate of 0.6 mL min−1 in the isocratic mode.
Mass spectrometry (MS)
Mass spectrometric detection was carried out to detect the presence of oligopeptides in the amino acid solution. The mass spectra were recorded for the soluble peptide fraction (as the insoluble microparticle suspensions self-separated by sedimentation) of the six investigated l-Pro samples straight away after 7 days storage in the measuring cell of turbidimeter. Mass spectra were recorded in the positive ionization mode with use of the Varian MS-100 mass spectrometer (the extended ESI–MS scan, positive ionization, the spray chamber temperature 50 °C, the drying gas temperature 250 °C, the drying gas pressure 25 psi, the capillary voltage 50 V, and the needle voltage 5 kV).
Scanning electron microscopy (SEM)
Visualization of nano- and microparticles was performed with a JEOL JSM-7600F model scanning electron microscope (SEM). Micrographs were recorded for the six investigated l-Pro samples after 1 month of aging at 22.0 ± 0.5 °C. Visualization was performed for microparticles obtained from the respective solutions evaporated to dryness.
Turbidimetry
Turbidity measurements were performed for six investigated l-Pro samples. To monitor the turbidity in continuous mode we used the turbidity sensor (TRB-BTA, Vernier Software & Technology, Beaverton, OR, USA). For the experiments 15 mL aliquots of the l-Pro solutions were freshly prepared and placed in the instrument vials. Turbidity changes were continuously registered for 7 days, in the 1-min intervals, under the thermostatic conditions at 25.0 ± 0.5 °C. To confirm the qualitative reproducibility of the results, the turbidity measurements were repeated twice.
The stability of turbidity was controlled for acetonitrile, water, heavy water and 70% aqueous acetonitrile as the reference solvents over the course of 1 day. In each case, the turbidity was quite stable and amounted accordingly 80.1 NTU (nephelometric turbidity units) for ACN, 0.0 NTU for H2O, 91.8 NTU for D2O and 40.1 NTU for the 70% aqueous ACN.
Results and discussion
The supplementary material contains schematic representation of the processes of: spontaneous oscillatory chiral inversion (Fig. S1a), spontaneous oscillatory peptidization (Fig. S1b) and of these two processes running in the parallel (Fig. S3c). Schemes given in Fig. S1 illustrate an up-to-the-date understanding of the molecular-level mechanisms of the aforementioned processes upon an example of l-Pro. It is worth noting that all elementary steps given in Figs. S1a) and S1b are largely based on transfer of the hydrogen cation which is particularly important in view of the fact that each subsequent part of this publication will focus on tracking an effect of D2O on the dynamics of peptidization of l-Pro using a variety of analytical techniques.
High-performance liquid chromatography with evaporative light scattering detection (HPLC-ELSD)
The raw HPLC-ELSD data given in this paper were first published by Sajewicz et al. [23] in 2014. Based on these results, new and more accurate calculations have been made and the results obtained, which are going to be presented in this section.
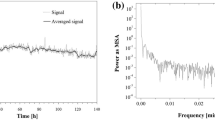
To perform the HPLC-ELSD experiment, the achiral HPLC mode was used to separate the monomeric l-Pro from the spontaneously produced peptides and to check changing amounts of the monomer. A chromatogram obtained with use of the ELSD detector for l-Pro dissolved in 70% aqueous MeOH remained qualitatively unchanged for 65 h of sample storage and the retention time of the monomeric l-Pro was equal to tR = 5.46 min. Fig. S2a (given in the supplementary material) demonstrates the time series of the changing L-Pro peak heights in the function of the sample storage period [23]. The non-linear changes in signal intensity may be observed, which are equivalent to quantitative changes of the monomeric l-Pro. To assess whether the HPLC signal recorded for the monomeric l-Pro contains a significant periodic component, a Fourier transform was performed on the data shown in Fig. S2a and the power spectrum calculated for the peak of l-Pro is plotted in Fig. S2b. In Fig. S2b, the predominant peak can be seen at 0.0008 min−1, which implies periodicity of ca. 20.8 h. Owing to a more accurate handling of the experimental data in this study (originally published in paper [23]), the period of the oscillatory peptidization of l-Pro comes out somewhat longer, as compared with our previous result (ca. 20 h, [23]). Periodicity of spontaneous peptidization of l-Pro (equal either to ca. 20 h, or ca. 20.8 h) unequivocally points out to the circadian rhythm of this process. The total length of the HPLC-ELSD data was limited to 65 h by experimental stability of the chromatographic system and certainly a longer time series would be desirable to even better assess the obtained periodicity.
For the sake of comparison, we also stored the l-Pro sample dissolved in pure D2O for the period of 48 h and practically no considerable quantitative changes of the amino acid signal were observed within this span of time (excluding an insignificant instrumental noise). Thus a conclusion was drawn that ageing of l-Pro in heavy water did not result in measurable oscillations originating from spontaneous oscillatory peptidization process.
Mass spectrometric (MS) tracing of spontaneous peptidization of l-Pro
The peptidization process of each of the six l-Pro samples was running spontaneously for 7 days at 25.0 ± 0.5 °C in the darkness. Then for each sample, the mass spectrum was recorded to give evidence of peptides formed in the course of the sample storage. Obviously, it is possible to register mass spectra for the monomeric l-Pro and for the soluble peptides only and in our case, registration was carried out in the range up to m/z 3500 (Fig. 1a–f). Higher insoluble peptides self-separated from the solution by sedimentation and their presence could only be documented with use of the scanning electron microscopy (SEM), as it will be demonstrated in the subsequent section.

Mass spectra recorded for l-Pro dissolved in MeOH + X, 70:30 (v/v), where X: the binary mixture of H2O + D2O in the changing volume proportions; a 0% D2O; b 5% D2O; c 10% D2O; d 20% D2O; e 30% D2O and fl-Pro dissolved in pure D2O
With the l-Pro sample containing no heavy water (D2O), mass spectrum shows quite considerable numbers of high intensity signals which appear mostly in the m/z range below 2500, with the highest signal intensities up to 80 kCounts (Fig. 1a). Mass spectrum obtained for the l-Pro sample containing 5% D2O reveals certain decrease in signal intensity, with the highest signal intensities below 60 kCounts (Fig. 1b). Moreover, in the mass spectrum treated as a fingerprint, a shift of the signals is observed towards the lower m/z values (below m/z 2000). With the l-Pro sample containing 10% D2O, the intensity of the registered signals is even lower, with the highest intensity values falling below 10 kCounts. Moreover, the mass spectrum treated as a fingerprint shows further shift of the signals toward the range below m/z 1000 (Fig. 1c). Interestingly, the mass spectrum recorded for the l-Pro sample with a still higher contribution of heavy water (20% D2O) again shows the high numbers of the high intensity signals (up to 100 kCounts), spread across an entire m/z range (Fig. 1d). A similar trend is maintained with the l-Pro solution containing 30% D2O (Fig. 1e). The mass spectrum recorded for l-Pro dissolved in pure D2O is characterized with very high signal intensities (up to 1400 kCounts), specifically in the m/z range below 2500 (Fig. 1f).
In conclusion, the results of mass spectrometry show an interesting and at the same time a rather striking pattern. Starting from the l-Pro sample that contains no heavy water up to that containing 10% D2O, gradual decrease is observed in the amounts and intensities of signals derived from soluble peptides. Beginning from the l-Pro sample with 20% D2O addition, high intensity signals originating from soluble peptides are again observed, with their yields and intensities gradually growing toward the highest numbers valid for the 100% content of D2O as a monocomponent solvent.
Scanning electron microscopic (SEM) tracing of spontaneous peptidization of l-Pro
After one month of storage the l-Pro samples, all of them were subjected to examination by means of SEM. For this purpose, drops of individual solutions along with the respective sediments were deposited on the measuring pin of the microscope and evaporated to dryness. This allowed registering the lower and soluble peptides (also recorded by mass spectrometry), but also the higher insoluble ones, appearing in the form of a sediment. Selected micrographs which illustrate the observed regularities are given in Fig. 2 and Fig. S3 (supplementary material).

Scanning electron micrographs recorded for the l-Pro-derived peptides retrieved from the samples dissolved in MeOH + X, 70:30 (v/v), where X: the binary mixture of H2O + D2O in the changing volume proportions; a 0% D2O, × 1900; b 10% D2O, × 750; c 20% D2O, × 2500; d 100% D2O, × 1500
Fig. 2a shows a micrograph recorded for the l-Pro sample stored in complete absence of heavy water (D2O). Peptides formed under such conditions give extensive, spatial and star-like structures. The addition of 5% D2O to the l-Pro sample causes visible difference in the obtained peptide structure (Fig. S3b; supplementary material). This time the elongated, streaked and branched structures appear that are no longer star-like. Texture of the peptide matter formed in the presence of 10% D2O is quite different from the previous two. Now we observe single, rather spherical peptides that are not organized into more extensive structures (Fig. 2b). Ten percent D2O in the l-Pro sample significantly inhibits growth of soluble peptide structures (as demonstrated by mass spectrometry), but it also results in small amounts of insoluble peptides present in the sediment (as demonstrated by means of SEM). The 20% proportion of D2O in the l-Pro solution again results in a granular structure of the resulting peptides. One can notice spherical clusters of peptide structures and small peptide fragments scattered across an entire field of view of the microscope (Fig. 2c). Peptides formed in the presence of 30% D2O (Fig. S3e; supplmentary material) appear as increasingly smaller clusters. Larger peptide clusters appear only sporadically and the high numbers of small peptides predominate. The micrograph recorded for l-Pro dissolved in pure D2O shows the smallest peptide aggregations from all those presented in this study. In fact, it shows sparse and loosely distributed peptide structures (Fig. 2d). Based on the results of scanning electron microscopy, it can be assumed that the large star- and band-like structures originate from insoluble particles of higher peptides, while small, spherical and largely dispersed structures are derived from the lower and most probably soluble peptides. In the other words, an increased content of D2O inhibits formation of the higher insoluble peptides.
Last not least, it needs to be frankly admitted that microscopic images depend on the place of focusing the microscope, which may lead to arbitrary choices, misinterpretations of the results obtained, or in extreme cases, even to some manipulations. To avoid such criticism, we added an overview of the micrographs valid for three different concentration of D2O in solution (0, 10 and 30% D2O) in Fig. S4 and recorded from three different places of focusing the microscope for one and the same sample. From these data it is evident that in despite of different place of focusing the microscope, basic characteristics of the D2O concentration impact on peptide yields (discussed in the preceding paragraph) remains unchanged.
In conclusion, the results of mass spectrometry combined with those derived from the scanning electron microscopy present an interesting picture, never observed in our earlier studies (l-Cys [32] and l-Met [33]) and showing an impact of heavy water on peptidization of proteinogenic α-amino acids. In the case of the l-Pro samples stored in an absence of heavy water, high yields of both the lower soluble peptides (MS) and the higher insoluble ones (SEM) are confirmed. An addition of 5% D2O equally reduces the yields of soluble and insoluble peptides. However, addition of 10% D2O not only slows down these two processes, but it virtually inhibits them. In this case, only a small number of soluble peptides is registered with MS and visualized by means of SEM. The results valid for the l-Pro sample containing 20% D2O indicate a return to high yields of soluble peptides (MS) that appear as fine structures in SEM. The l-Pro samples containing 30 and 100% D2O demonstrate an “awaited” tendency, observed in our earlier studies [32, 33]. This tendency can be described in the following way: With the growing proportions of heavy water in solution, the yields of soluble peptides increase and those of insoluble peptides decrease.
Turbidimetric tracing of spontaneous peptidization of l-Pro
The plots of turbidity changes were recorded in the 1-min intervals for the period of 7 days for one l-Pro sample dissolved in pure D2O and five l-Pro samples dissolved in the mixture of MeOH + X, 70:30 (v/v), where X: the binary mixture of H2O + D2O in the changing volume proportions: 30:0, 25:5, 20:10, 10:20 and 0:30. The obtained data reveal differences in peptidization dynamics, depending on the amount of heavy water contained in each of the samples. Selected results well reflecting the trend of the observed changes are presented in Fig. S5 (supplementary material).
A sudden initial increase in signal intensity visible in both turbidimetric plots is an effect of adjusting the temperature of the samples to the temperature of the turbidity measuring device set at 25.0 ± 0.5 °C (as turbidity is strongly temperature-dependent). For l-Pro dissolved in MeOH + H2O (in an absence of D2O), the turbidity values start step-wise dropping after a few hours from the beginning of the experiment (as indicated with an arrow in Fig. S5a). A systematic decline in the range of two turbidity units (from 46 to 44 NTU) lasts until the third day of sample storage, and then a rather stagnant plot is observed, with insignificant turbidity changes until the end of the measuring period. The 20% content of D2O results in an extension of the initiation period to 3 days (as indicated with an arrow in Fig. S5b), and then a step-wise and short-lived increase in turbidity of the sample starts, lasting approx. half a day. Then up to day 6, quite insignificant turbidity changes (within the range of ca. 0.1 NTU) can be noticed, after which time the turbidity starts gradually dropping. Summing up, it can be concluded that the presence of D2O in solution clearly extends the initiation period of turbidity changes.
It should not be forgotten that due to the lack of standardization in turbidity units, measuring devices and calibration techniques, the use of turbidimetry for analytical determinations can only be empirical and rather qualitative. However, the results and discussion provided in this section confirm our earlier observations on hampering effect of D2O on the process of spontaneous peptidization of proteinogenic α-amino acids, reported in papers [32, 33] and also based on turbidimetric results.
Conclusion
When considering spontaneous peptidization of l-Pro with different contributions of D2O, we can distinguish two characteristic ranges 1 and 2 of the D2O proportions in solution. The results originating from range 1 with lower proportions (5 and 10%) of D2O characterize with strong hampering of formation of the lower (soluble) and the higher (insoluble) peptides, as documented with use of two analytical techniques, i.e., mass spectrometry (MS) and scanning electron microscopy (SEM). Almost full inhibition of peptidization was eventually observed for l-Pro dissolved in the mixture containing 10% D2O. The results valid for higher proportions (20 and 30%) of D2O in the liquid system, and also for pure D2O used as a solvent belong to range 2. These results characterize with monotonously increasing yields of lower (soluble) peptides and monotonously decreasing yields of higher (insoluble) peptides. A general statement can be made that characteristic chemical structure of the l-Pro molecule incorporating the pyrrolidine ring and the secondary amino group can be responsible for these two different patterns of peptidization.
References
Kredel FE, Harkins HN, Harkins WD (1934) Toxicity of heavy water. Proc Soc Exp Biol Med 32:5
Katz JJ, Crespi HL, Czajka DM, Finkel AJ (1962) Course of deuteriation and some physiological effects of deuterium in mice. Am J Physiol 203:907–913
Bachner P, McKay DG, Rittenberg D (1964) The pathological anatomy of deuterium intoxication. Proc Natl Acad Sci USA 51:464–471
Richter CP (1976) A study of taste and smell of heavy water (98%) in rats. Proc Soc Exp Biol Med 152:677–684
Kanto U, Clawson AJ (1980) Use of deuterium oxide for the in vivo prediction of body composition in female rats in various physiological states. J Nutr 110:1840–1848
Laissue JA, Burki H, Berchtold W (1982) Survival of tumorbearing mice exposed to heavy water or heavy water plus methotrexate. Cancer Res 42:1125–1129
Altermatt HJ, Gebbers JO, Laissue JA (1988) Heavy water delays growth of human carcinoma in nude mice. Cancer 62:462–466
Takeda H, Nio Y, Uegaki K, Hirahara Sasaki S, Tamura K, Ohtani H (1998) Mechanism of cytotoxic effects of heavy water (deuterium oxide: D2O) on cancer cells. Anticancer Drugs 9:715–725
Kushner DJ, Baker A, Dunstall TG (1999) Pharmacological uses and perspectives of heavy water and deuterated compounds. Can J Physiol Pharmacol 77:79–88
Hartmann J, Bader Y, Horvath Z, Saiko P, Grusch M, Illmer C, Madlener S, Fritzer-Szekeres M, Alken R-G, Szekers T (2005) Effects of heavy water (D2O) on human pancreatic tumor cells. Anticancer Res 25:3407–3412
Kalkur RS, Ballast AC, Triplett AR, Spendier K (2014) Effects of deuterium oxide on cell growth and vesicle speed in RBL-2H3 cells. PeerJ 2:e553. https://doi.org/10.7717/peerj.553
Lamprecht J, Schroeter D, Paweletz N (1990) Mitosis arrested by deuterium oxide. Light microscopic, immunofluorescence and ultrastructural characterization. Eur J Cell Biol 51:303–312
Lamprecht J, Schroeter D, Paweletz N (1989) Disorganization of mitosis in HeLa cells by deuterium oxide. Eur J Cell Biol 50:360–369
Schroeter D, Lamprecht J, Eckhardt R, Futterman G, Paweletz N (1992) Deuterium oxide (heavy water) arrests the cell cycle of PtK2 cells during interphase. Eur J Cell Biol 58:365–370
Sajewicz M, Kronenbach D, Gontarska M, Kowalska T (2008) TLC and polarimetric investigation of the oscillatory in vitro chiral inversion of l-alanine. J Planar Chromatogr 21(1):43–47
Sajewicz M, Gontarska M, Wojtal Ł, Kronenbach D, Leda M, Epstein IR, Kowalska T (2008) Experimental and model investigation of the oscillatory transenantiomerization of l-α-phenylalanine. J Liq Chromatogr Relat Technol 31:1986–2005
Sajewicz M, Kronenbach D, Staszek D, Wróbel M, Grygierczyk G, Kowalska T (2008) Experimental investigation of the oscillatory transenantiomerization of l-tyrosine. J Liq Chromatogr Relat Technol 31:2006–2018
Sajewicz M, Gontarska M, Kowalska T (2014) HPLC/DAD evidence of the oscillatory chiral conversion of phenylglycine. J Chromatogr Sci 52:329–333
Sajewicz M, Gontarska M, Kronenbach D, Kowalska T (2009) On the spontaneous abiotic peptization of phenylglycine in an aqueous medium. Acta Chromatogr 21:151–160
Sajewicz M, Matlengiewicz M, Leda M, Gontarska M, Kronenbach D, Kowalska T, Epstein IR (2010) Spontaneous oscillatory in vitro chiral conversion of simple carboxylic acids and its possible mechanism. J Phys Org Chem 23:1066–1073
Sajewicz M, Gontarska M, Kronenbach D, Leda M, Kowalska T, Epstein IR (2010) Condensation oscillations in the peptidization of phenylglycine. J Syst Chem. https://doi.org/10.1186/1759-2208-1-7
Sajewicz M, Matlengiewicz M, Juziuk M, Penkala M, Weloe M, Schulz M, Kowalska T (2013) Thin-layer chromatographic evidence of proline peptidization in solution and its thin-layer chromatographic enantioseparation. J Liq Chromatogr Relat Technol 36:2497–2511
Sajewicz M, Dolnik M, Kowalska T, Epstein IR (2014) Condensation dynamics of l-proline and l-hydroxyproline in solution. RSC Adv 4:7330–7339
Godziek A, Maciejowska A, Talik E, Wrzalik R, Sajewicz M, Kowalska T (2016) On spontaneously pulsating proline-phenylalanine peptide microfibers. Curr Protein Pept Sci 17:106–116
Maciejowska A, Godziek A, Talik E, Sajewicz M, Kowalska T, Epstein IR (2016) Spontaneous pulsation of peptide microstructures in an abiotic liquid system. J Chromatogr Sci 54:1301–1309
Godziek A, Łągiewka A, Kowalska T, Sajewicz M (2018) The influence of heavy water as a solvent on the spontaneous oscillatory reactions of α-amino acids. Reac Kinet Mech Cat 123:141–153
Kreuels T, Martin W, Brinkmann K (1980) Influence of D2O on the Belousov-Zhabotinsky reaction. Ber Bunsenges Phys Chem 84:411–412
Karavaev AD, Kazakov VP, Tolstikov GA (1986) Deuteration effect in auto-oscillation chemiluminescencje of the Belousov-Zhabotinskii reaction. Reac Kinet Catal Lett 32:21–26
Rossi F, Rustici M, Rossi C, Tiezzi E (2007) Isotopic effect on the kinetics of the Belousov-Zhabotinsky reaction. Int J Mol Sci 8:943–949
Stanisavljev D, Begović N, Vukojević V (1998) Influence of heavy water in the Bray-Liebhafsky oscillating reaction. J Phys Chem A 102:6887–6891
Stanisavljev DR, Vukojević VB (2002) Investigation of the influence of heavy water on kinetic pathways in the Bray-Leiebhafsky reaction. J Phys Chem A 106:5618–5625
Fulczyk A, Łata E, Dolnik M, Talik E, Kowalska T, Sajewicz M (2018) Impact of D2O on peptidization of L-cysteine. Reac Kinet Mech Cat 125:555–565
Fulczyk A, Łata E, Talik E, Kowalska T, Sajewicz M (2019) Impact of D2O on peptidization of l-methionine. Reac Kinet Mech Cat 126:939–949
Barbul A (2008) Proline precursors to sustain mammalian collagen synthesis. J Nutr 138:2021S–2024S
Pavone P, Pratico AD, Sorge G, Meli C, Ruggieri M, Rizzo R, Fiumara A (2017) Hyperprolinemia Type IA: benign metabolic anomaly or a trigger for brain dysfunction? J Inborn Errors Metab Screen 5:1–3
Mitsubuchi H, Nakamura K, Matsumoto S, Endo F (2014) Biochemical and clinical features of hereditary hyperprolinemia. Pediatr Int 56:492–496
Clelland CL, Read LL, Baraldi AN, Bart CP, Pappas CA, Panek LJ, Nadrich RH, Clelland JD (2011) Evidence for association of hyperprolinemia with schizophrenia and a measure of clinical outcome. Schizophr Res 131:139–145
Duarte M, Alonso J, Moreira A, Antunes D, Ferreira C, Correia H, Marques M, Sequeira S (2017) Hyperprolinemia as a clue in the diagnosis of a patient with psychiatric manifestations. Brain Dev 39:539–541
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Fulczyk, A., Łata, E., Talik, E. et al. Impact of D2O on peptidization of l-proline. Reac Kinet Mech Cat 128, 599–610 (2019). https://doi.org/10.1007/s11144-019-01681-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11144-019-01681-y