
Abstract
Purpose
Flower strips have been shown to increase insect biodiversity and improve agricultural yields through increased pollination and pest predation. Less is known about their potential to increase soil organic carbon (SOC). We aimed to investigate the biomass production and SOC sequestration potential of flower strips as a sustainable management option of temperate agricultural soils.
Methods
23 flower strips across varying soil types and climatic regions in Germany were sampled for aboveground and belowground peak biomass in order to estimate the annual carbon input to the soil. Those were used as 23 scenarios to model the potential SOC sequestration of the flower strips compared to a business-as-usual scenario for 1533 sites of the German Agricultural Soil Inventory using the RothC model.
Results
On average, flower strips sequestered 0.48 ± 0.36 Mg C ha−1 year−1 in the initial 20-year period after establishment. Converting 1 % of the total German cropland area into flower strips would thus lead to a mitigation of 0.24 Tg CO2 year−1, which equals 0.4 % of current agricultural greenhouse gas emissions in Germany.
We found a negative correlation between C sequestration rate and the number of plant species in the flower strips, mainly related to grasses outcompeting herbaceous species.
Conclusion
Flower strips are one overlooked option for increasing SOC stocks of croplands that has multiple benefits for agro-ecosystems. However, within a flower strip it might not be possible to maximise both plant biodiversity and SOC sequestration.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Soil organic carbon (SOC) sequestration is acknowledged as an essential element of achieving carbon–neutral societies and mitigating greenhouse gas emissions (Freibauer et al. 2004; Jacobs et al. 2020; Sanderman et al. 2017). Therefore, so-called carbon farming practices are being actively incorporated into political strategies, including the European Union’s “Farm to Fork Strategy” and the Common Agricultural Policy (European Commission 2022). Additionally, a voluntary carbon market, in which CO2 certificates are traded based on SOC sequestration is emerging (Jacobs et al. 2020).
Historical land use changes and long-term agricultural management is estimated to have caused a loss of 116 Pg C from soils to the atmosphere, which amounts to about 17 % of total atmospheric CO2 (Sanderman et al. 2017). Cropland soils are often depleted in SOC (Prout et al. 2021). One of the major drivers of these historic and ongoing SOC losses is a reduction in carbon input, due to a high human appropriation of the net primary production (NPP) in agricultural systems (Haberl et al. 2007). Increases in SOC can therefore only be achieved by a higher return of biomass to the soil, which should be aimed at by e.g. maximising the photosynthetic activity on the field (Janzen et al. 2022). Measures like cover cropping, agroforestry, hedgerows or increasing the proportion of permanent crops such as grasses are currently considered as most efficient for increasing SOC stocks in cropland soils (Bellassen et al. 2022; Drexler et al. 2021; Lugato et al. 2014). All of these measures have in common that they not only increase SOC, but also create synergies such as increasing biodiversity, reducing soil erosion, or taking up excess nutrients (Blanco-Canqui et al. 2015; Fan et al. 2015).
Such synergistic effects also apply to flower strips, as a feature of a more sustainable agriculture in several countries that exercise intense, industrialised agricultural production. In these production systems, flower strips, i.e. parts of agricultural fields that are seeded with mostly diverse flower mixes, are a broadly recognised, effective method for increasing biodiversity of both plants and animals, particularly insects, in the agricultural landscape (Haaland et al. 2011; Nicholls and Altieri 2013; Pfiffner et al. 2019; Sutter et al. 2017; Thomas and Marshall 1999). Furthermore, they have been shown to increase agricultural yield of both cereal crops (Carreck and Williams 2002; Tschumi et al. 2015) and fruit orchards (Foutain 2022; Garibaldi et al. 2013) through increased pollination (Buhk et al. 2018; Feltham et al. 2015) as well as through increased predation of harmful insects (Tschumi et al. 2016). Moreover, they have been shown to provide additional ecosystem services such as animal habitats, particularly in winter (Gontijo 2019; Haaland et al. 2011; Harwood et al. 1994), and improved landscape aesthetics (Westphal et al. 2015). Finally, flower strips are likely to produce large amounts of biomass that is not removed from the field at harvest, thereby increasing not only the photosynthetic carbon input into agricultural soils during growth, but also via incorporation of aboveground biomass during regular tillage, suggesting a potential for increasing SOC stocks and soil carbon sequestration. Thus, flower strips provide multiple ecosystem functions in agriculture systems; increasing biodiversity, and through that improving yields, as well as potentially mitigating greenhouse gas emissions through increased carbon sequestration.
Although the effect of flower strips on insect biodiversity and agricultural yields have been studied thoroughly, the characterization of flower strips themselves is rarely the focus of examination. Numerous aspects of the flower strip itself remain unknown, including the biomass production potential and their effect on SOC stocks. The aim of this study was to i) comprehensively measure aboveground and belowground peak biomass under various pedo-climatic conditions, ii) assess if short-term effects on SOC stocks are detectable and iii) estimate the SOC sequestration potential of flower strips using a SOC turnover model in combination with observed biomass and estimated C input data using 1533 points from the German Soil Inventory. The study was performed on German agricultural soils as a model region for temperate, intensive agriculture.
Materials and methods
Study sites and sampling procedure
In this study, 23 flower strips located adjacent to cropland in Germany were investigated. The sampling sites were selected in order to cover the diverse pedo-climatic conditions of Germany (Fig. 1, S. Tab 1). Flower strips were already established but not yet mulched down at the time of sampling (July—September 2021). The different site characteristics (clay content, silt content, pH, age of the flower strip, mean annual temperature, mean total annual precipitation) are provided in S. Tab 1. Flower strips were not fertilised nor harvested, so that the entire biomass remained on the flower strip. There were both annual (i.e. reseeding each year) and perennial flower strips. The annual flower strips were mulched, tilled and reseeded every year with an annual flower mix, while few sites (GG1, Ha, HB and Lue) were seeded once and not disturbed afterwards.

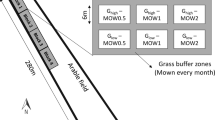
Diagram of the experimental setting, sampling design and modelling process. The location of the 23 flower strips are shown as green circles on the map of Germany in the top left-hand side. The sampling design for the soil cores (brown circles) are shown in the cropland and flower strip at each flower strip location, as well as the sampling areas for the aboveground and belowground biomass sampling in the flower strips (green squares). Created with Biorender.com
In each flower strip, five plots were established for sampling the aboveground and belowground biomass. These plots were 50 × 50 cm in size, 12 m apart from each other and at least 12 m from the top and bottom edges of the flower strip in order to avoid edge effects. No specific distance to the cropland was maintained because the flower strips were of different widths ranging from 6 to 30 m. The flower strips were sampled between July and September 2021, at the expected time of greatest standing biomass (Ram et al 1989; Wang et al. 2020).
The aboveground biomass (AGB) growing within the sampling plot was harvested by cutting it 1 cm above the soil. The individual plants cut from the sampling plot were identified to species-level, and the number of each observed species was recorded. Afterwards, the total AGB sample was air-dried to weight constancy.
The belowground biomass (BGB) was sampled in the upper topsoil layer (0–10 cm) and the lower topsoil layer (10–30 cm). In the upper topsoil layer, the total soil mass was excavated and spread on a canvas and all bigger central roots were picked out manually. To determine the mass of the smaller roots, a representative composite sample of the upper topsoil layer was taken.
In the lower soil layer (10–30 cm), three samples per plot were collected true to volume using a root corer (4.7 cm diameter). The collected BGB, was washed to separate it from soil and other organic materials. Before washing, the composite soil sample from the 0–10 cm layer with the finer roots was weighed and analysed for water content. Root samples were washed with a root washer (GVF, Benzonia, MI, USA). Each root sample was collected on a 0.63 mm sieve. Any other litter material (e.g. straw) was sorted out by hand. After washing, the roots were dried at 75 °C until constant weight and then weighed. While the 10–30 cm root biomass and the 0–10 cm coarser central root biomass were calculated per volume (all roots sampled per given volume), the 0–10 cm finer root biomass was calculated based on a root:soil weight ratio in combination with soil bulk density. We chose that for this complex and unreferenced root sampling setup, due to the fact that the diversity and patchiness of plants in the flower strips made representative core sampling almost impossible. At the same time, excavating and washing the whole 0–30 cm layer at an area of 50 × 50 cm was also not feasible. A compromise of manually picking out coarser topsoil roots from the whole soil volume plus representative sampling of finer roots was thus attempted.
For the chemical analysis of the soil (C, N, pH, soil texture), five disturbed samples were taken with a thin auger (2 cm diameter) and pooled according to the depth intervals 0–10 cm (upper topsoil) and 10–30 cm (lower topsoil) in the flower strip and in the adjacent cropland. The soil samples were dried at 60 °C until constant weight, and then sieved at 2 mm and milled in a ball mill. Afterwards, the soil samples were analysed for organic carbon and total nitrogen contents by dry combustion in an elemental analyser (LECO TruMac, St. Joseph, MI, USA).
Whole soil bulk density was determined by volumetric samples using steel cylinders (n = 5, 250 cm3) in the flower strips and the adjacent cropland for each layer. The rock fragment fraction was neglected in this study. The bulk density samples were dried at 105 °C for one week and then weighed. SOC stocks (Mg ha−1) were calculated by multiplying bulk density with sampling depth and SOC content. SOC stocks were compared based on equivalent soil mass (Wendt and Hauser 2013).
Carbon input calculation and RothC modelling
Due to the low age of the flower strips (on average three years since establishment), we were not able to base any SOC sequestration estimates on measured SOC stock changes alone. Therefore, we used the measured flower strip biomass data in a comprehensive modelling approach to estimate the carbon sequestration potential of flower strips in German cropland topsoils (0–30 cm). Flower strip carbon input was calculated using measured above- and below-ground biomass with established conversion factors (0.45 for above- and 0.4 for below-ground respectively; Kätterer et al. 2011). The latter value originates from an earlier root washing study and is lower than for aboveground biomass due to a slight dilution with mineral particles (Poeplau et al. 2018). To account for root exudates, 31 % of belowground biomass C was added to the total belowground C input (Pausch and Kuzyakov 2018).
Each set of C inputs (AGC and BGC) from the sampled flower strips was used as an input scenario. All 23 flower strip scenarios were used as C inputs on a total of 1533 selected sites from the German Agricultural Soil Inventory. In the German Agricultural Soil Inventory, a grid of 8 × 8 km covering a total of 3104 sampling points (2234 on croplands) was sampled with the primary purpose of used to estimating a baseline SOC stocks of German agricultural soils to a depth of 1 m (Poeplau et al. 2020). From these data, 1533 cropland sites were selected from the complete inventory of agricultural soils based on availability of management data and soil type criteria; a minimum of six consecutive years of reliable management data (main crop type and yield, fertiliser and manure application) was required, and organic soils (> 87 g kg−1 SOC) were excluded. The available management including information on crop yields, cover crops and organic fertilisers were used to calculate OC inputs for a business-as-usual (BAU) scenario with allocation coefficients (Riggers et al. 2019). The input estimation as described by (Franko et al. 2011) was applied here, since it gave the best fit to 139 German long-term monitoring sites (Riggers et al. 2019). All 23 flower strip scenarios as well as the BAU scenario were run on all 1533 points, so that a total of 36,792 model runs were completed using the RothC model (Coleman and Jenkinson 1996) as modified by (Dechow et al. 2019).
RothC is a model describing soil carbon decomposition using first order kinetics (Eq. 1) and a system of four interacting SOC pools each characterised by a specific reference decomposition rate (kref) (Coleman and Jenkinson 1996). These decomposition rates are modulated by rate modifying factors considering the effects of temperature (a), soil water deficit (b) and soil cover (c). This reference decomposition rate is applied to calculate the change in SOC stock (Y) over time (Eq. 2).
Monthly climate data required for RothC modelling was retrieved from DWD (German meteorological service) for the period 2001–2017, for each of the individual Inventory points. Both climate and management time series were repeated for 400 years, to run the model into equilibrium.
The ratio of decomposable plant material (DPM) and resistant plant material (RPM) (DPM/RPM ratio), as a parameter for organic matter decomposability, was assumed to be equal for cereal crops and flower strips (1.44).
In the flower strips scenarios, it was assumed that the soil was covered by vegetation the whole year, while fallow periods occurred between consecutive crops in crop rotations of the BAU scenario. In RothC, crop coverage has implications for the decomposition rate modifying factor c (Coleman and Jenkinson 1996) that is used to differentiate between SOC decomposition for periods with (c = 0.6) and without vegetation cover (c = 1.0) on a monthly basis, so that in the end the actual decomposition rates are slightly lower under flower strips compared to fallow periods in the BAU scenario. Vegetation cover has a strong effect on soil moisture, which has been shown to slow down organic matter decomposition as compared to bare soil (Sparling et al. 1982). This is incorporated into the RothC model by Coleman and Jenkinson (1996).
The SOC sequestration potential of flower strips was estimated for each individual inventory site and for each scenario as follows:
where the flower strip scenario SOC stocki (Mg C ha−1) is the modelled soil C stock at time i for a given combination of inventory site and flower strip scenario C inputs, and BAU SOC stock (Mg C ha−1) is the modelled soil C stock at time i with the corresponding inventory C inputs. Flower strip NPP was not directly associated with soil properties or climate (Fig. 6). We thus ran all possible combinations and argue that the range of estimated sequestration rates will reflect the variability of the potential C sequestration of flower strips.
Statistics
Correlations between the measurements of the flower strips (AGB, BGB, total biomass, root:shoot ratio, age of the flower strip, total number of plant species), soil parameters (sand, silt and clay content, pH), climatic parameters (mean temperature and total precipitation of the vegetation period (April—September)) as well as the Shannon index, Pielou index and the average C sequestration rate for the initial 20 years were assessed using Spearman’s correlation.
Shannon Index, quantifying plant species diversity, was calculated according to (Shannon and Weaver 1964):
where S is the total number of species observed in the flower strip, and pi is the proportion of plants of species i out of the total number of plants in the flower strip.
Pielou Index, referring to plant species evenness, was calculated according to (Pielou 1966):
where H’ and S are as defined above.
Paired t-tests were used to determine the significance of the differences in SOC stocks between flower strips and cropland.
Modelling and statistical analyses were performed in R 4.1.1 (R Core Team 2020). RothC was run using the SoilR package (Sierra et al. 2012), and analysis of the modelling results was done with linear models. Differences in C input between the BAU scenario and the 23 flower strip scenarios were assessed with paired Sign-tests from the BSDA package (Arnholt and Evans 2021) as the distributions of the variables were skewed. Confidence intervals for median values were calculated using the DescTools package (Signorell, et mult. al. 2022).
Results
Flower strip biomass and carbon stocks
In the flower strips, the mean total biomass was 9.8 ± 3.3 Mg ha−1. The mean AGB production was 7.4 ± 3.4 Mg ha−1, which corresponds to 75 % of the total biomass. BGB contributed a smaller share of the total biomass production (2.4 ± 1.5 Mg ha−1; 25 %). The coefficient of variation between sites for BGB (62.5) was larger than that of AGB (33.7).
The mean root:shoot ratio was 0.4 ± 0.3 (Fig. 2B) with two flower strips having root:shoot ratios greater than 1, meaning a greater biomass belowground compared to aboveground.

Boxplots of a) dry matter (Mg ha−1) of the flower strips (n = 23); AGB (aboveground biomass), BGB (belowground biomass), Total Biomass (AGB + BGB) and b) root:shoot ratio (unitless). Observations shown as dots; outliers signified by crosses
The SOC stocks for the sampled sites and for each soil layer are shown in Fig. 3A. The upper topsoil layer showed significantly higher mean values of SOC stocks in the flower strips (25.9 ± 6.7 Mg ha−1) compared to cropland (23.0 ± 7.2 Mg ha−1). In the lower topsoil layer, the flower strips showed a slightly higher mean value of SOC stocks (42.6 ± 15.8 Mg ha−1) compared to cropland (40.5 ± 13.3 Mg ha−1), but these differences were not significant.

SOC stocks (Mg ha−1) of the cropland (blue) and adjacent flower strips (green) for the upper (0–10 cm), lower (10–30 cm) and complete topsoil layer (0–30 cm) a) in boxplots (n = 23), and B) the difference between the flower strip SOC stock and the cropland SOC stock (Mg ha−1) for the individual flower strip locations. Flower strip “Siggen” (Si) has very high SOC stock compared to the cropland, and has therefore been excluded from b) to increase readability of the graph
The mean stocks for the complete soil depth of 0–30 cm were 68.4 ± 20.9 Mg ha−1 for the flower strips and 63.5 ± 18.4 Mg ha−1 for cropland, but these values were also not significantly different (P > 0.05). For 15 out of 23 sites, the mean value of SOC stocks of the complete topsoil profile in the flower strips was higher compared to that of the cropland (Fig. 3B).
RothC modelling
Across all combinations of inventory sites and flower strip scenarios, the mean SOC sequestration potential of the flower strips after 50 years was 16.5 ± 12.9 Mg C ha−1 (Fig. 4). 92 % of all model runs of the flower strip scenarios result in a gain of soil C after 50 years compared to the BAU scenario.

Combined violin plot and scatter plot of the change in SOC (Mg ha−1) of flower strips across 50 years, with a fitted GAM (generalised additive model) line (black), where the potential total sequestered C is the difference between the BAU scenario and the flower strip scenario. Years 10, 20, 30, 40, and 50 are included in the figure, and the black GAM-line is fitted to the same years as well as through 0. A linear model (green) is fitted to years 0, 10 and 20. For each year, n = 35,259
After the first 20 years, a mean of 9.6 ± 7.3 Mg C ha−1 has been accumulated in the soils under flower strips compared to the BAU scenario. This corresponds to an average yearly sequestration rate of 0.48 ± 0.36 Mg C ha−1 year−1.
The modelled rate of change in SOC stock within the initial 20 years of flower strip establishment varies greatly between flower strip scenarios, and scales with total biomass (Fig. 5). Only the GG 2 and TE 2 scenarios appear to have a considerable number of model runs with a negative rates of change in SOC stocks after 20 years, both of which have relatively low flower strip biomass.

Distribution of the modelled rate of change in SOC stock (Mg C ha−1 year−1) of the initial 20 years for each inventory site (n = 1533) for each of the 23 flower strip scenarios. The colour scales with the total biomass in the flower strip scenario (Mg ha−1). A difference of 0 is shown by a grey line, and the striped line signifies the mean rate of change in SOC stock across all combinations of inventory sites and flower strip scenarios
The sequestration rate of the flower strips correlated significantly and positively with both aboveground biomass, belowground biomass and total biomass. Simultaneously, there was a significantly negative correlation between the mean C sequestration rate and the number of species observed in the flower strip as well as Shannon index and Pielou index. Generally, total biomass was also significantly negatively correlated with the number of observed plant species, as well as the Shannon and Pielou indices, albeit not statistically significant at P < 0.05 (Fig. 6).

Correlogram of influences of soil characteristics, weather data and count of different species on biomass production, root:shoot ratio, C sequestration rate, age and diversity parameters in the flower strips. The colour indicates the Spearman’s correlation coefficient p and the asterisks indicate the significance level (* < 0.05, ** < 0.01, *** < 0.001). PrecipVeg21 is the precipitation of the vegetation period (April to September) 2021, TempVeg21 is the mean temperature of the vegetation period (April to September) 2021. C seq rate, 20 years is the mean C sequestration rate for the initial 20 years of that flower strip scenario compared to the BAU scenario. The plant names (e.g. Sunflower, Buckwheat) is the count of the individual plant species
No individual plant species correlated significantly with aboveground biomass with the exception of grasses (as an order), where a negative correlation was observed. However, a significant positive correlation between grasses and belowground biomass was also observed, resulting in no significant correlation between grasses and total biomass.
Both the median aboveground C inputs and the median belowground C inputs were significantly greater (P < 0.001) in the flower strip scenarios than for the BAU scenario (Tab. 1). Further, the BAU scenario had a greater BGC:AGC ratio compared to the flower strip scenarios (Fig. 7, Table 1, P < 0.001), which is related to the fact that a significant amount of the AGC was exported from croplands as yield. The median values of AGC, BGC and BGC:AGC ratios were compared, as the distributions of observations for both the BAU scenario and the flower strip scenarios were positively skewed. In addition to being skewed, all three C inputs in the flower strip scenarios had multiple unevenly distributed peaks (Fig. 7), which may be caused by the relatively low number of observations and great variation between flower strips (n = 23).

Probability density (%) graph of the a) aboveground C (AGC, Mg C ha−1) input, excluding organic fertiliser, b) belowground C (BGC, Mg C ha−1) input and c) BGC:AGC ratio (unitless) for the BAU scenario (blue) and the flower strip scenarios (green). In C, 28 inventory sites (1.8 % of 1533 sites) have been excluded to increase readability of the figure, as their BGC:AGC ratio is greater than 3. The dashed blue line represents the median value of the BAU scenario input (n = 1533), while the solid green line depicts the median of the flower strip scenarios (n = 23)
In the BAU scenario, organic fertiliser was part of the C input to the soil while in the flower strip scenario, we assumed that no organic fertiliser was applied. Even though the median C input from organic fertilisers across all inventory sites was 0.36 Mg C ha−1, the median total C input from all sources is still significantly greater in the flower strip scenarios compared to the BAU scenarios (Table 1).
The sites with the greatest mean sequestration rate for flower strips are located in Central and Eastern Germany (Fig. 8, darkest blue colour). The median sequestration rate is negative at 49 sites (3.2 % of all inventory sites) across all flower strip scenarios, of which one site has a negative sequestration rate for all flower strip scenarios. The sites that have near-zero or negative mean sequestration rates are primarily located in the North-Western areas of Germany as well as the Southernmost regions (Fig. 8).

Map of the mean rate of change in SOC (Mg C ha−1 year−1) for all of the flower strip scenarios of the initial 20 years, for the 1533 Inventory sites. Red signifies loss of SOC compared to the BAU scenario, while blue signifies increases in SOC compared to the BAU scenario
In Germany, 12.67 mio hectares of mineral soil (< 87 g C kg−1) are currently used as cropland (Federal Environmental Agency 2021). Converting 1 % of the German cropland area with mineral soils (approximately 0.13 mio ha) to flower strips with an average sequestration rate of 0.48 Mg C ha−1 year−1 in the first 20 years, will result in a yearly increase of 0.065 Tg C in the top 30 cm of soil per year.
Discussion
Carbon sequestration of flower strips
Our estimated sequestration rate of the first 20 years of flower strips on German croplands was 0.48 Mg C ha−1 year−1. The mean sequestration rate for the first 50 years was 0.33 Mg C ha−1 year−1 which is similar to that of cover crops, estimated by Seitz et al. (2022) to be 0.28—0.33 Mg C ha−1 year−1 for Germany in the initial 50 years, as well as the mean estimate of 0.32 Mg C ha−1 year−1 from a global review study by (Poeplau and Don 2013).
In addition to cover crops, other proposed carbon sequestration measures related to potentially increased NPP in situ include land use conversion to grasslands as well as agroforestry, such as hedgerows (Bellassen et al. 2022).
Estimates of the C sequestration potential of agricultural land converted to grasslands range from 0.4—1.9 Mg C ha−1 year−1 for a 20-year period, with an average of 0.85 Mg C ha−1 year−1 (Freibauer et al. 2004; Hu et al. 2019; Johnston et al. 2009; Poeplau and Don 2013). The estimated SOC sequestration potential of agroforestry in Europe is 0.22 Mg C ha−1 year−1 (Bellassen et al. 2022; Pellerin et al. 2020), while Kay et al. (2019) estimates the potential of various combinations of agroforestry practices in Europe to range between 0.09—7.29 Mg C ha−1 year−1 in soil and woody biomass combined.
The carbon sequestration rate of hedgerows has been estimated to be 2.1—5.2 Mg C ha−1 year−1 for the whole ecosystem including woody biomass (Drexler et al. 2021), and ranging between 0.3—1.5 Mg C ha−1 year−1 of C sequestration in the 0–30 cm layer of the soil (Biffi et al. 2022; Drexler et al. 2021). Hedgerows can serve similar biodiversity functions in an agricultural landscape as flower strips (Montgomery et al. 2020; Thiel et al. 2015), but flower strips have a more positive effect on pollinators and yield improvements through predation (Albrecht et al. 2020).
Thus, the C sequestration potential of measures with permanent woody biomass on croplands is certainly higher than that of flower strips. However, the implementation of agroforestry requires structural and system-wide changes to agricultural management practices, which make these measures difficult and expensive for farmers, and may additionally result in a net loss of income through lost productivity. Similarly, hedgerows can be immense commitments for farmers, as they are established long-term in one site and need regular care with cutting. In comparison, flower strips are relatively simple to implement, inexpensive and do not require long term commitment from the farmers. However, the benefits in terms of C sequestration of both measures require continued long-term commitment since the sequestered C can be released back to the atmosphere when the measure is stopped.
Generally, an increase in C input to the soil has the potential to increase the SOC stock (Janzen et al. 2022; Riggers et al. 2019). Overall, the flower strip scenarios create greater total C inputs to the soil, particularly from aboveground biomass compared to the BAU scenario, (Fig. 7, Table 1), which can be explained by the harvest and subsequent export of biomass from croplands which does not occur on flower strips. Additionally, the C sequestered by application of manure in the BAU scenario does not count towards net C sequestration. Since the flower strips do not receive manure, the net C sequestration of flower strips as compared to the BAU scenario was thus even slightly underestimated. The flower strip scenarios also had a 3 % higher median belowground C input, which can be caused by the presence of perennial plants (grasses, clovers), particularly in more established (older) flower strips as the plant species composition may shift towards grasses (Fig. 6). Added C from fertiliser and manure accounted for 12 % (median) of total added C in the BAU, but this did not compensate for the differences in plant biomass C input between the flower strip scenarios and the BAU scenario.
As climatic variables and soil parameters were identical for the BAU scenario and flower strip scenarios, the differences in C input can drive the observed C sequestration effect of the flower strips. In addition to differences in C input, the flower strips scenarios also assume full soil coverage across the year, while the BAU scenario can have fallow periods between annual crops in accordance with the management information. Thus, the decomposition rates of the modelled SOC pools in RothC were slower for the flower strips, as soil cover reduces the decomposition rate of a pool.
The greatest effect on SOC stock occured when the yearly aboveground C input of the flower strip was much larger than the BAU scenario (> 5 Mg C ha−1, S. Figure 1). However, significant positive changes in SOC stock were also observed when the total biomass C or AGC input of the flower strip scenarios were similar to or slightly less than that of the BAU scenario (S. Figure 1). In these cases, the BGC:AGC ratio was relatively high, so that the acknowledged and implemented relatively higher stabilisation efficiency of root-derived C may have outweighed the lower total C input.
The fact that the difference in amount of C input was the major driver of difference in SOC stock between the BAU scenario and the flower strip scenarios is also delineated in the spatial pattern of C sequestration through flower strip implementation across Germany (Fig. 8). The greatest C inputs from crop residues and organic fertilisers as well as the NPP production patterns were found in North-West and South-East Germany (Jacobs et al. 2020). This pattern is reflected in where the effect of flower strip implementation was observed to be smallest (Fig. 8). The highest SOC sequestration potential of flower strips might thus be found in Central and North-Eastern Germany, assuming that the pedo-climatic conditions are suitable everywhere to reach comparable flower strip NPP as measured in the present study. Our assumption that the pedo-climatic conditions allow for a similar NPP production across Germany can be questioned, particularly for the drier and less fertile areas of Eastern Germany. However, i) the obtained dataset of flower strip biomass in this study was not correlated to climate variables and only weakly correlated with soil properties (Fig. 6) and ii) the flower strips sampled in exactly this rather dry part of Germany had the fourth, seventh and eighteenth highest total biomass, which suggests that these flower strips do not produce consistently less biomass than those sampled in other regions of Germany. Thus, we have no observations that indicate that the flower strips should not be representative for various climate and soil conditions within Germany.
As presented above, the modelled SOC sequestration rates in this study appear reasonable and are well in the range of comparable measures. We also detected short-term significant differences in SOC stocks in 0–10 cm and a positive tendency also in the 10–30 cm layer (Fig. 3). We interpret this as an indication that flower strips increase SOC stocks when kept for a certain time. However, i) the missing significant difference in 0–30 cm, ii) the low average flower strip age of 3.2 years, as well as iii) the absent correlation of SOC stock change and age of the flower strip prevented us from deriving sequestration rates based on measured SOC stocks. This also highlights the difficulty of detecting changes in SOC stocks after only a few years (Smith 2004), which is, however, commonly attempted by upcoming carbon certification schemes.
The dynamics of other greenhouse gases such as methane and nitrous oxide have not been investigated for flower strips. Other carbon sequestration methods, such as cover cropping, have been shown to increase e.g. nitrous oxide emissions under certain circumstances (Basche et al. 2014), and the total effect of flower strips on climate change requires these estimates to be thorough. It is however likely that nitrous oxide emissions will drop significantly upon flower strip establishment due to the cessation of fertilisation (Shcherbak et al. 2014).
Currently, about 117.057 ha of flower strips are established in Germany as of 2019 (Fritz 2019), which corresponds to approximately 1 % of the cropland area. If an additional 1 % of cropland on mineral soils in Germany was converted to flower strips, 0.065 Tg C year−1 (0.24 Tg CO2 year−1) could potentially be sequestered annually in the top 30 cm of the soil. This corresponds to a small proportion of the annual greenhouse gas emissions from agriculture in Germany (0.4 %, NIR 2021). However, i) SOC sequestration is only one positive feature of flower strips, ii) the likely reduction in N2O emissions would increase the climate mitigation potential of flower strips and iii) on the path towards climate neutrality, even options with relatively limited potential should be considered if they do not interfere with or prevent options of greater impact. The goal of a 1 % conversion of cropland to long-term flower strips is ambitious, yet not unrealistic.
Uncertainties
The estimated C inputs from above- and below-ground biomass from flower strips come with uncertainties. Firstly, the biomass was sampled only once, at the estimated overall peak biomass. Secondly, no estimates for flower strip biomass, from which the C inputs are derived, have previously been published, and thus there are no references to directly compare and evaluate our observations with. However, within a multi-species flower strip as well as across flower strips of varying species composition, it is unlikely that we actually caught this exact moment of peak biomass at all sampled sites. Some of the unexplained variation across flower strip biomass might thus arise from the differences in sampling dates. It is likely that we underestimated total biomass due to the fact that NPP was either ongoing or concluded weeks ago for some species at the chosen sampling dates between July and September. At the same time, belowground C inputs could be overestimated, since we converted the full root biomass into C inputs, while this would only be the case for annual plants. Perennial plants like grasses maintain a proportion of their roots so that root biomass does not equal C inputs to the soil (Gill and Jackson 2000). However, it was not possible to differentiate between annual and perennial roots in the samples. It could not be assessed, which effect caused a larger error or whether they cancel each other out.
Further uncertainties arise from the modelling exercise from various sources. On the one hand, the strengths of the used set of inventory points are that i) they originate from a systematic random sample (monitoring grid) that is considered as representative for German croplands and that ii) quality checked management information that directly correspond to measured SOC stocks could be used for the BAU scenario, which reduce the uncertainty. On the other hand, it has been shown that SOC turnover models are very sensitive to C input estimates. These estimates can be derived by many different sets of allocation coefficients, which have been found to result in a wide range of average C inputs to the soil (Keel et al. 2017; Riggers et al. 2019). Still, the chosen combination of model and input estimation approach was found to perform well on 139 long-term monitoring sites in Germany (Riggers et al. 2019). Furthermore, we are confident that using the recalibrated version of the RothC model (Dechow et al. 2019), which is also capable of distinguishing between different types of input (AGC/BGC, organic fertiliser), reduced the model-error compared to the more widely used standard version. The magnitude and range of modelled SOC sequestration in this study appeared reasonable, although we were not able to directly validate the model results due to the lack of long-term SOC timeseries under flower strips.
Synergies, trade-offs and potential leakage effects
Flower strips contribute with numerous ecosystem services, including increased plant and insect biodiversity, provide nesting and foraging habitats for animals, reduce soil erosion and minimise nutrient leaching, as well as improve landscape aesthetics and can be used for human recreational purposes (Haddaway et al. 2018; Marshall and Moonen 2002; Westphal et al. 2015). There was a significant positive correlation between the mean C sequestration rate of the initial 20 years and both the total biomass as well as belowground biomass of the flower strip (Fig. 6), suggesting that highly productive flower strips have higher sequestration potential. Further, we found that flower strips with many different plant species and evenly distributed and more diverse plant populations had lower C sequestration rates. Thus, in contrast to a previous study showing that cover crop diversity may have a positive effect on C inputs (Gentsch et al. 2020), the present study suggests a potential trade-off between C sequestration and plant species diversity in the flower strips.
Flower strips are not always resown in spring so the plant species composition can shift from deliberately sown species (sunflowers, buckwheat, phacelia) towards spontaneously occurring plant species, some of which are not included in typical flower strip mixtures (grasses, thistle) (Fig. 6). If flower strips are not re-established after a few years, it is possible that the intentionally sown flower species may be outcompeted by grasses (Piqueray et al. 2019), which can result in lower plant biodiversity than intentionally diverse flower strips. However, the diversity of multi-species grassland has also been shown to support high pollinator biodiversity (Cong et al. 2020). In this study, no direct correlation between C sequestration rate and presence of grass in the flower strip was found (Fig. 6). However, the abundance of grasses was positively correlated to the root:shoot ratio of the flower strip, which may successively translate into a higher sequestration efficiency (Kätterer et al. 2011; Poeplau et al. 2021), which may influence the lack of correlation between species diversity and C sequestration of the flower strips.
Unlike cover crops, it is not feasible to implement flower strips on large proportions of the cropland area, and the total potential effect for climate change mitigation is thereby comparatively lower. However, converting edges of agricultural fields to flower strips can be done independently of the crop and the related management, and thus, flower strips are a more versatile management tool for increasing the total SOC stock in cropland areas.
Additionally, Klimm (2020) has shown that flower strips need to be tailored to the regional environmental conditions to have significant positive impacts on ecosystem services, including provision of food and shelter for native insects.
Conversion of cropland to flower strips reduces the area under cultivation, potentially compromising the production of agricultural goods. This could cause land-use spillover where land use change in one region causes land use change in another region (Meyfroidt et al. 2020); if the agricultural yield in Germany decreases due to implementation of flower strips, it is possible that a comparable area will be converted to agricultural land somewhere else to sustain food, feed and fibre production on a global scale. Leakage effects should thus be considered when the overall benefits of flower strips are discussed. However, obligations for European farmers to set cropland aside are increasing and flower strips seem one promising option to do so. Furthermore, they may even have positive effects on per area yield (Pywell et al. 2015), which might compensate for losses of cropping area.
Conclusion
For the first time, biomass of flower strips has been determined and analysed in relation to plant biodiversity. We found a significant negative correlation between the total number of unique plant species in the flower strip and its total biomass production, suggesting that highly diverse flower strips are less productive. The biomass of the flower strip is not harvested or removed, and thus flower strips have a much greater input of C to the soil compared to croplands.
Additionally, the C sequestration potential of flower strips has also been modelled for the first time. We estimate that flower strips can sequester 0.48 Mg ± 0.36 C ha−1 year−1 in the initial 20 years after establishment in temperate, intensively used mineral soils. This rate is comparable to other C sequestrations methods, including cover crops and land use conversion to grassland. As with the total biomass, there is a negative correlation between C sequestration rate and biodiversity; thus there may be a trade-off between maximising plant biodiversity and C sequestration in flower strips.
If an additional 1 % of German cropland on mineral soils was converted to flower strips, an additional 0.065 Tg C would be sequestered in the soil, which corresponds to 0.4 % of all GHG emissions from the German agricultural sector. Converting cropland to flower strips is not a fail-safe C sequestration method. Areas with already-high inputs of organic C from organic fertilisers, including manure, and biomass will see a lower potential C sequestering effect of flower strips. Furthermore, only a permanent flower strip can act as a significant C sink for climate mitigation. However, flower strips often improve agricultural yields through increased pollinator attraction and pest predation and have synergistic relations with overall ecosystem biodiversity. Therefore, implementing flower strips as a simple and easy-to-implement management option by farmers can improve multiple critical agro-environmental issues simultaneously.
Data availability
The datasets generated during and analysed during the current study are available in the Zenodo repository “Flower strip data Germany 2021” and through https://doi.org/10.5281/zenodo.7077659.
References
Albrecht M, Kleijn D, Williams NM, Tschumi M, Blaauw BR, Bommarco R, Campbell AJ, Dainese M, Drummond FA, Entling MH, Dominik Ganser G, de Groot A, Goulson D, Grab H, Hamilton H, Herzog F, Isaacs R, Jacot K, Jeanneret P, Jonsson M, Knop E, Kremen C, Landis DA, Loeb GM, Marini L, McKerchar M, Morandin L, Pfister SC, Potts SG, Rundlöf M, Sardiñas H, Sciligo A, Thies C, Tscharntke T, Venturini E, Veromann E, Vollhardt IMG, Wäckers F, Ward K, Westbury DB, Wilby A, Woltz M, Wratten S, Sutter L (2020) The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol Lett 23(10):1488–1498. https://doi.org/10.1111/ele.13576
Signorell, A. et mult. al. (2022) DescTools: tools for descriptive statistics. R package version 0.99.46. https://cran.r-project.org/package=DescTools
Arnholt AT, Evans B (2021) BSDA: Basic Statistics and Data Analysis. R package version 1.2.1. https://CRAN.R-project.org/package=BSDA
Basche AD, Miguez FE, Kaspar TC, Castellano MJ (2014) Do Cover Crops Increase or Decrease Nitrous Oxide Emissions? A Meta-Analysis. J Soil Water Conserv 69(6):471–482. https://doi.org/10.2489/jswc.69.6.471
Bellassen V, Angers D, Kowalczewski T, Olesen A (2022) Soil Carbon Is the Blind Spot of European National GHG Inventories. Nat Clim Chang 12(4):324–331. https://doi.org/10.1038/s41558-022-01321-9
Biffi S, Chapman PJ, Grayson RP, Ziv G (2022) Soil Carbon Sequestration Potential of Planting Hedgerows in Agricultural Landscapes. J Environ Manage 307:114484. https://doi.org/10.1016/j.jenvman.2022.114484
Blanco-Canqui H, Shaver TM, Lindquist JL, Shapiro CA, Elmore RW, Francis CA, Hergert GW (2015) Cover Crops and Ecosystem Services: Insights from Studies in Temperate Soils. Agron J 107(6):2449–2474. https://doi.org/10.2134/agronj15.0086
Buhk C, Oppermann R, Schanowski A, Bleil R, Lüdemann J, Maus C (2018) Flower Strip Networks Offer Promising Long Term Effects on Pollinator Species Richness in Intensively Cultivated Agricultural Areas. BMC Ecol 18(1):55. https://doi.org/10.1186/s12898-018-0210-z
Carreck NL, Williams IH (2002) Food for Insect Pollinators on Farmland: Insect Visits to Flowers of Annual Seed Mixtures. J Insect Conserv 6(1):13–23. https://doi.org/10.1023/A:1015764925536
Coleman K, Jenkinson DS (1996) ‘RothC-26.3 - A Model for the Turnover of Carbon in Soil’. In: Powlson DS, Smith P, Smith JU (eds) Evaluation of Soil Organic Matter Models, NATO ASI Series. Berlin, Springer: Heidelberg, pp. 237–46
Cong W, Dupont YL, Søegaard K, Eriksen J (2020) Optimizing Yield and Flower Resources for Pollinators in Intensively Managed Multi-Species Grasslands. Agric Ecosyst Environ 302:107062. https://doi.org/10.1016/j.agee.2020.107062
Dechow R, Franko U, Kätterer T, Kolbe H (2019) Evaluation of the RothC Model as a Prognostic Tool for the Prediction of SOC Trends in Response to Management Practices on Arable Land. Geoderma 337:463–478. https://doi.org/10.1016/j.geoderma.2018.10.001
Drexler S, Gensior A, Don A (2021) Carbon Sequestration in Hedgerow Biomass and Soil in the Temperate Climate Zone. Reg Environ Change 21(3):74. https://doi.org/10.1007/s10113-021-01798-8
European Commission (2022) ‘Carbon Farming’. Retrieved https://ec.europa.eu/clima/eu-action/forests-and-agriculture/sustainable-carbon-cycles/carbon-farming_en. Accessed 21 July 2022
Fan Ji, Yan L, Zhang P, Zhang Ge (2015) Effects of Grass Contour Hedgerow Systems on Controlling Soil Erosion in Red Soil Hilly Areas, Southeast China. Int J Sedim Res 30(2):107–116. https://doi.org/10.1016/j.ijsrc.2015.03.001
Federal Environmental Agency (2021) Struktur der Flächennutzung. https://www.umweltbundesamt.de/daten/flaeche-boden-land-oekosysteme/flaeche/struktur-der-flaechennutzung. Accessed 21 July 2022
Feltham H, Park K, Minderman J, Goulson D (2015) Experimental Evidence That Wildflower Strips Increase Pollinator Visits to Crops. Ecol Evol 5(16):3523–3530. https://doi.org/10.1002/ece3.1444
Foutain (2022) Wildflower interventions on beneficial insects in fruit crops: A review. Insects. https://doi.org/10.3390/insects13030304
Franko U, Kolbe H, Thiel E, Ließ E (2011) Multi-Site Validation of a Soil Organic Matter Model for Arable Fields Based on Generally Available Input Data. Geoderma 166(1):119–134. https://doi.org/10.1016/j.geoderma.2011.07.019
Freibauer A, Rounsevell MDA, Smith P, Verhagen J (2004) Carbon Sequestration in the Agricultural Soils of Europe. Geoderma 122(1):1–23. https://doi.org/10.1016/j.geoderma.2004.01.021
Fritz A (2019) Artenschutz: Über 200.000 Kilometer Blühstreifen in Deutschland. AgrarHeute. https://www.agrarheute.com/pflanze/artenschutz-ueber-200000-kilometer-bluehstreifen-deutschland-553936. Accessed 21 July 2022
Garibaldi, Lucas A., Ingolf Steffan-Dewenter, Rachael Winfree, Marcelo A. Aizen, Riccardo Bommarco, Saul A. Cunningham, Claire Kremen, Luísa G. Carvalheiro, Lawrence D. Harder, Ohad Afik, Ignasi Bartomeus, Faye Benjamin, Virginie Boreux, Daniel Cariveau, Natacha P. Chacoff, Jan H. Dudenhöffer, Breno M. Freitas, Jaboury Ghazoul, Sarah Greenleaf, Juliana Hipólito, Andrea Holzschuh, Brad Howlett, Rufus Isaacs, Steven K. Javorek, Christina M. Kennedy, Kristin M. Krewenka, Smitha Krishnan, Yael Mandelik, Margaret M. Mayfield, Iris Motzke, Theodore Munyuli, Brian A. Nault, Mark Otieno, Jessica Petersen, Gideon Pisanty, Simon G. Potts, Romina Rader, Taylor H. Ricketts, Maj Rundlöf, Colleen L. Seymour, Christof Schüepp, Hajnalka Szentgyörgyi, Hisatomo Taki, Teja Tscharntke, Carlos H. Vergara, Blandina F. Viana, Thomas C. Wanger, Catrin Westphal, Neal Williams, and Alexandra M. Klein. 2013. ‘Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance’. Science (New York, N.Y.) 339(6127):1608–11. https://doi.org/10.1126/science.1230200
Gentsch N, Boy J, Batalla JDK, Heuermann D, von Wirén N, Schweneker D, Feuerstein U, Groß J, Bauer B, Reinhold-Hurek B, Hurek T, Céspedes FC, Guggenberger G (2020) Catch Crop Diversity Increases Rhizosphere Carbon Input and Soil Microbial Biomass. Biol Fertil Soils 56(7):943–957. https://doi.org/10.1007/s00374-020-01475-8
Gill RA, Jackson RB (2000) Global Patterns of Root Turnover for Terrestrial Ecosystems. New Phytol 147(1):13–31. https://doi.org/10.1046/j.1469-8137.2000.00681.x
Gontijo LM (2019) Engineering Natural Enemy Shelters to Enhance Conservation Biological Control in Field Crops. Biol Control 130:155–163. https://doi.org/10.1016/j.biocontrol.2018.10.014
Haaland C, Naisbit RE, Bersier L-F (2011) Sown Wildflower Strips for Insect Conservation: A Review. Insect Conserv Divers 4(1):60–80. https://doi.org/10.1111/j.1752-4598.2010.00098.x
Haberl H, Heinz Erb K, Krausmann F, Gaube V, Bondeau A, Plutzar C, Gingrich S, Lucht W, Fischer-Kowalski M (2007) Quantifying and Mapping the Human Appropriation of Net Primary Production in Earth’s Terrestrial Ecosystems. Proc Natl Acad Sci 104(31):12942–12947. https://doi.org/10.1073/pnas.0704243104
Haddaway NR, Brown C, Eales J, Eggers S, Josefsson J, Kronvang B, Randall NP, Uusi-Kämppä J (2018) The Multifunctional Roles of Vegetated Strips around and within Agricultural Fields. Environ Evid 7(1):14. https://doi.org/10.1186/s13750-018-0126-2
Harwood RWJ, Wratten SD, Nowakowski M, Marshall EPJ (1994) ‘Wild Flower Strips and Winter/Summer Populations of Beneficial Invertebrates on Farmland’. In: Bulletin OILB SROP (France). OILB
Hu T, Taghizadeh-Toosi A, Olesen JE, Jensen ML, Sørensen P, Christensen BT (2019) Converting Temperate Long-Term Arable Land into Semi-Natural Grassland: Decadal-Scale Changes in Topsoil C, N, 13C and 15N Contents. Eur J Soil Sci 70(2):350–360. https://doi.org/10.1111/ejss.12738
Jacobs Anna, Heidecke Claudia, Jumshudzade Zaur (2020) ‘Soil Organic Carbon Certificates - Potential and Limitations for Private and Public Climate Action’. Landbauforschung J Sustain Org Agric Syst 70(2020)2):31–35 https://doi.org/10.3220/LBF1605778405000
Janzen H Henry, van Groenigen Kees Jan, Powlson David S, Schwinghamer Timothy, van Groenigen Jan Willem (2022) ‘Photosynthetic Limits on Carbon Sequestration in Croplands’. Geoderma 416:115810. https://doi.org/10.1016/j.geoderma.2022.115810
Johnston AE, Poulton PR, Coleman K (2009) Soil Organic Matter: Its Importance in Sustainable Agriculture and Carbon Dioxide Fluxes. Adv Agron 101:1–57. https://doi.org/10.1016/S0065-2113(08)00801-8
Kätterer T, Bolinder MA, Andrén O, Kirchmann H, Menichetti L (2011) Roots Contribute More to Refractory Soil Organic Matter than Above-Ground Crop Residues, as Revealed by a Long-Term Field Experiment. Agric Ecosyst Environ 141(1):184–192. https://doi.org/10.1016/j.agee.2011.02.029
Kay S, Rega C, Moreno G, den Herder M, Palma JHN, Borek R, Crous-Duran J, Freese D, Giannitsopoulos M, Graves A, Jäger M, Lamersdorf N, Memedemin D, Mosquera-Losada R, Pantera A, Paracchini ML, Paris P, Roces-Díaz JV, Rolo V, Rosati A, Sandor M, Smith Jo, Szerencsits E, Varga A, Viaud V, Wawer R, Burgess PJ, Herzog F (2019) Agroforestry Creates Carbon Sinks Whilst Enhancing the Environment in Agricultural Landscapes in Europe. Land Use Policy 83:581–593. https://doi.org/10.1016/j.landusepol.2019.02.025
Keel SG, Leifeld J, Mayer J, Taghizadeh-Toosi A, Olesen JE (2017) Large Uncertainty in Soil Carbon Modelling Related to Method of Calculation of Plant Carbon Input in Agricultural Systems. Eur J Soil Sci 68(6):953–963. https://doi.org/10.1111/ejss.12454
Klimm Fabian (2020) ‘The Value of Flower Strips in Agricultural Landscapes - Effects on Pollination, Herbivory and Decomposition’
Lugato E, Bampa F, Panagos P, Montanarella L, Jones A (2014) Potential Carbon Sequestration of European Arable Soils Estimated by Modelling a Comprehensive Set of Management Practices. Glob Change Biol 20(11):3557–3567. https://doi.org/10.1111/gcb.12551
Marshall EJP, Moonen AC (2002) Field Margins in Northern Europe: Their Functions and Interactions with Agriculture. Agric Ecosyst Environ 89(1):5–21. https://doi.org/10.1016/S0167-8809(01)00315-2
Meyfroidt P, Börner J, Garrett R, Gardner T, Godar J, Kis-Katos K, Soares-Filho BS, Wunder S (2020) Focus on Leakage and Spillovers: Informing Land-Use Governance in a Tele-Coupled World. Environ Res Lett 15(9):090202. https://doi.org/10.1088/1748-9326/ab7397
Montgomery Ian, Caruso Tancredi, Reid Neil (2020) ‘Hedgerows as Ecosystems: Service Delivery, Management and Restoration’. Ann Rev Ecol Evol Syst. https://doi.org/10.1146/annurev-ecolsys-012120-100346
Nicholls CI, Altieri MA (2013) Plant Biodiversity Enhances Bees and Other Insect Pollinators in Agroecosystems. A Review. Agron Sustain Dev 33(2):257–274. https://doi.org/10.1007/s13593-012-0092-y
Pausch J, Kuzyakov Y (2018) Carbon Input by Roots into the Soil: Quantification of Rhizodeposition from Root to Ecosystem Scale. Glob Change Biol 24(1):1–12. https://doi.org/10.1111/gcb.13850
Pellerin, S, Laure B, Camille L, Raphaël M, Schiavo M, Angers D, Augusto L, Balesdent J, Basile-Doelsch I, Bellassen V, Cardinael R, Cécillon L, Ceschia E, Chenu C, Constantin J, Daroussin J, Delacote P, Delame N, Gastal F, Gilbert D, Graux A, Guenet B, Houot S, Klumpp K, Letort E, Litrico I, Martin M, Menasseri-Aubry S, Meziere D, Morvan T, Mosnier C, Roger-Estrade J, Saint-André L, Sierra J, Therond O, Viaud V, Grateau R, Le Perchec S, Savini I, Rechauchère O (2020) Stocker Du Carbone Dans Les Sols Français, Quel Potentiel Au Regard de l’objectif 4 Pour 1000 et à Quel Coût? Agence de l’Environnement et de la Maîtrise de l’Energie. https://doi.org/10.15454/1.5433098269609653E12
Pfiffner L, Cahenzli F, Steinemann B, Jamar L, Bjørn MC, Porcel M, Tasin M, Telfser J, Kelderer M, Lisek J, Sigsgaard L (2019) Design, Implementation and Management of Perennial Flower Strips to Promote Functional Agrobiodiversity in Organic Apple Orchards: A Pan-European Study. Agric Ecosyst Environ 278:61–71. https://doi.org/10.1016/j.agee.2019.03.005
Pielou EC (1966) The Measurement of Diversity in Different Types of Biological Collections. J Theor Biol 13:131–144. https://doi.org/10.1016/0022-5193(66)90013-0
Piqueray J, Gilliaux V, Decruyenaere V, Cornelis J-T, Uyttenbroeck R, Mahy G (2019) Management of Grassland-like Wildflower Strips Sown on Nutrient-Rich Arable Soils: The Role of Grass Density and Mowing Regime. Environ Manage 63(5):647–657. https://doi.org/10.1007/s00267-019-01153-y
Poeplau C, Don A (2013) Sensitivity of Soil Organic Carbon Stocks and Fractions to Different Land-Use Changes across Europe. Geoderma 192:189–201. https://doi.org/10.1016/j.geoderma.2012.08.003
Poeplau C, Don A, Schneider F (2021) Roots Are Key to Increasing the Mean Residence Time of Organic Carbon Entering Temperate Agricultural Soils. Glob Change Biol 27(19):4921–4934. https://doi.org/10.1111/gcb.15787
Poeplau C, Jacobs A, Don A, Vos C, Schneider F, Wittnebel M, Tiemeyer B, Heidkamp A, Prietz R, Flessa H (2020) Stocks of Organic Carbon in German Agricultural Soils—Key Results of the First Comprehensive Inventory. J Plant Nutr Soil Sci 183(6):665–681. https://doi.org/10.1002/jpln.202000113
Poeplau C, Zopf D, Greiner B, Geerts R, Korvaar H, Thumm U, Don A, Heidkamp A, Flessa H (2018) Why Does Mineral Fertilization Increase Soil Carbon Stocks in Temperate Grasslands? Agric Ecosyst Environ 265:144–155. https://doi.org/10.1016/j.agee.2018.06.003
Prout JM, Shepherd KD, McGrath SP, Kirk GJD, Haefele SM (2021) What Is a Good Level of Soil Organic Matter? An Index Based on Organic Carbon to Clay Ratio. Eur J Soil Sci 72(6):2493–2503. https://doi.org/10.1111/ejss.13012
Pywell RF, Heard MS, Woodcock BA, Hinsley S, Ridding L, Nowakowski M, Bullock JM (2015) Wildlife-Friendly Farming Increases Crop Yield: Evidence for Ecological Intensification. Proc R Soc B Biol Sci 282(1816):20151740. https://doi.org/10.1098/rspb.2015.1740
R Core Team (2020) ‘R: A Language and Environment for Statistical Computing’
Ram J, Singh JS, Singh SP (1989) Plant Biomass, Species Diversity and Net Primary Production in a Central Himalayan High Altitude Grassland. J Ecol 77(2):456–468. https://doi.org/10.2307/2260762
Riggers C, Poeplau C, Don A, Bamminger C, Höper H, Dechow R (2019) Multi-Model Ensemble Improved the Prediction of Trends in Soil Organic Carbon Stocks in German Croplands. Geoderma 345:17–30. https://doi.org/10.1016/j.geoderma.2019.03.014
Sanderman J, Hengl T, Fiske GJ (2017) Soil Carbon Debt of 12,000 Years of Human Land Use. Proc Natl Acad Sci 114(36):9575–9580. https://doi.org/10.1073/pnas.1706103114
Seitz D, Fischer LM, Dechow R, Wiesmeier M, Don A (2022) The Potential of Cover Crops to Increase Soil Organic Carbon Storage in German Croplands. Plant Soil. https://doi.org/10.1007/s11104-022-05438-w
Shannon C, Weaver W (1964) The mathematical theory of communication. The University of Illinois Press, Urbana, p 131
Shcherbak I, Millar N, Philip Robertson G (2014) Global Metaanalysis of the Nonlinear Response of Soil Nitrous Oxide (N2O) Emissions to Fertilizer Nitrogen. Proc Natl Acad Sci 111(25):9199–9204. https://doi.org/10.1073/pnas.1322434111
Sierra C, Mueller M, Trumbore S (2012) Models of soil organic matter decomposition: the SoilR package. R package version 1.0. https://doi.org/10.5194/gmd-5-1045-2012
Smith P (2004) How Long before a Change in Soil Organic Carbon Can Be Detected? Glob Change Biol 10(11):1878–1883. https://doi.org/10.1111/j.1365-2486.2004.00854.x
Sparling GP, Cheshire MV, Mundie CM (1982) Effect of Barley Plants on the Decomposition of 14C-Labelled Soil Organic Matter. J Soil Sci 33(1):89–100. https://doi.org/10.1111/j.1365-2389.1982.tb01750.x
Sutter L, Jeanneret P, Bartual AM, Bocci G, Albrecht M (2017) Enhancing Plant Diversity in Agricultural Landscapes Promotes Both Rare Bees and Dominant Crop-Pollinating Bees through Complementary Increase in Key Floral Resources. J Appl Ecol 54(6):1856–1864. https://doi.org/10.1111/1365-2664.12907
Thiel B, Smukler SM, Krzic M, Gergel S, Terpsma C (2015) Using Hedgerow Biodiversity to Enhance the Carbon Storage of Farmland in the Fraser River Delta of British Columbia. J Soil Water Conserv 70(4):247–256. https://doi.org/10.2489/jswc.70.4.247
Thomas CFG, Marshall EJP (1999) Arthropod Abundance and Diversity in Differently Vegetated Margins of Arable Fields. Agric Ecosyst Environ 72(2):131–144. https://doi.org/10.1016/S0167-8809(98)00169-8
Tschumi M, Albrecht M, Bärtschi C, Collatz J, Entling MH, Jacot K (2016) Perennial, Species-Rich Wildflower Strips Enhance Pest Control and Crop Yield. Agric Ecosyst Environ 220:97–103. https://doi.org/10.1016/j.agee.2016.01.001
Tschumi M, Albrecht M, Entling MH, Jacot K (2015) High Effectiveness of Tailored Flower Strips in Reducing Pests and Crop Plant Damage. Proc R Soc B Biol Sci 282(1814):20151369. https://doi.org/10.1098/rspb.2015.1369
Wang H, Liu H, Cao G, Ma Z, Li Y, Zhang F, Zhao X, Zhao X, Jiang L, Sanders NJ, Classen AT, He J-S (2020) Alpine Grassland Plants Grow Earlier and Faster but Biomass Remains Unchanged over 35 Years of Climate Change. Ecol Lett 23(4):701–710. https://doi.org/10.1111/ele.13474
Wendt JW, Hauser S (2013) An Equivalent Soil Mass Procedure for Monitoring Soil Organic Carbon in Multiple Soil Layers. Eur J Soil Sci 64(1):58–65. https://doi.org/10.1111/ejss.12002
Westphal C, Vidal S, Horgan FG, Gurr GM, Escalada M, Van Chien Ho, Tscharntke T, Heong KL, Settele J (2015) Promoting Multiple Ecosystem Services with Flower Strips and Participatory Approaches in Rice Production Landscapes. Basic Appl Ecol 16(8):681–689. https://doi.org/10.1016/j.baae.2015.10.004
Acknowledgements
This work was conducted in the framework of the German Agricultural Soil Inventory funded by the Federal ministry of food and agriculture. We express our thanks to Roland Prietz, Frank Hegewald and Nicole Altwein for great support and are grateful for rapid sample processing by the Thünen Laboratory of Soil Monitoring.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
Christopher Poeplau, Henrike Heinemann and Gesa Schulz contributed to the conception and design of the study, as well as the material preparation and data collection. Data analysis was performed by Laura Sofie Harbo and Gesa Schulz. Modelling was performed by Laura Sofie Harbo and Rene Dechow. The first draft of the manuscript was mainly written by Laura Sofie Harbo, Gesa Schulz and Christopher Poeplau, and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
This work was conducted in the framework of the German Agricultural Soil Inventory, funded by the Federal Ministery of Food and Agriculture.
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Responsible Editor: Tida Ge.
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Harbo, L.S., Schulz, G., Heinemann, H. et al. Flower strips as a carbon sequestration measure in temperate croplands. Plant Soil 482, 647–663 (2023). https://doi.org/10.1007/s11104-022-05718-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-022-05718-5