
Abstract
Background
Combinations of grasses and nitrogen-fixing legumes are ubiquitous in most natural and derived pastoral grasslands. This was not formerly the case in New Zealand’s unique indigenous grasslands that are now frequently impacted by exotic pasture grasses and legumes. Understanding the co-existence of native and exotic plants is the broad focus of this research.
Aims
Spillover of nitrogen (N) from clovers to grasses in diverse pasture is well known. We question whether grasses provide reciprocal nutritional benefits to legumes. Does the mutual exploitation of soil biogeochemistry by legumes and grasses help to explain their coexistence and, if so, does this have implications for biodiversity in NZ’s novel native grassland communities?
Methods
Combinations of grasses and legumes, including a native tussock grass, were grown in a nutrient-poor (low P, S, Ca, Mg, Mn, and B) high country soil in a pot experiment, quantifying the foliar acquisition of nutrients from soil. Field data were obtained by sampling foliage of clover in single- and mixed-species patches in a more fertile lowland pasture.
Results
Benefits of legume and grass growing together were reflected in enhanced productivity and higher uptake of a range of key nutrients. This was most evident but not restricted to a combination of two exotic species: cocksfoot and white clover. In the nutrient-poor soil, legumes grew better in combination with different species of introduced grasses. Uptake of key elements from soil to plants significantly differed with combinations of legumes and grasses compared to individual species. Elevated concentrations of P, K and S were recorded in clover when growing with grasses, although Ca uptake was lower. Expected reciprocal reduction of clover N or enhanced grass N were not recorded. Mass balance data (total extraction of key nutrients from the soil pool) showed that combination of grasses and legumes exploit soil nutrients (particularly P, Zn, Mn and Mo) more effectively than single species alone. In grasses, only tissue concentrations of K, S and Zn significantly increased when growing with legumes, but native tussock grass procured less nutrients when growing with the exotic legumes. Field sampling of clover from the more fertile lowland soil showed significantly higher foliar concentrations of K, Mn, Cu and B, but less Ca.
Conclusions
The findings are indicative of a mutualistic relationship: legumes derive nutritional benefits from growing with grasses. Native tussock grass contained less N when growing with the exotic legumes, suggesting less compatibility and a lack of adaptation to coexistence.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Coexistence and maintenance of diversity in plant communities are explained as a result of different species exploiting resources that are spatially and temporally variable between and within habitats (Burrows, 1990; Høgh-Jensen and Schjoerring 2001; Homulle et al. 2021). Competition for resources is much more extensively studied than the mutual benefits that may be derived for different plant species growing together (Craine and Dybzinski 2013; Trinder et al. 2021). However, complementarity between species can be highly significant; agricultural intercropping of two or more species is one obvious and demonstrable example (Li et al. 2014; Gliessman 2015). This has been explained in terms of the abilities of each species to exploit different niches in the above-ground environment and root zone (Zhang et al. 2015). In terms of plant nutrition, the best-known example of complementarity is symbiotic nitrogen fixation in legumes that provides incidental spillover of nitrogen to soil, for example from clovers to grasses in mixed-species grasslands (Fornara and Tilman 2008; Pirhofer-Walzl 2012). A greater total herbage yield is obtained by the two functional types growing together rather than individually (Schipanski and Drinkwater 2012; Sturludóttir et al. 2014); this also balances palatability and improves nutritional benefits to stock. In this example, and in view of widespread and naturally occurring grass-legume associations, we question whether this relationship and its association with soil fertility operates only in one direction, or if grasses provide other elements to clovers as reciprocal nutrition. Our hypothesis is that spillover of N from clovers is an intentional strategy to procure limiting nutrients from neighbouring grasses. This is based on an assumption that there would be no selective advantage to legumes of fixing N in excess of their own requirements. Conversely, there would be a clear advantage to be able to acquire limiting soil nutrients without direct metabolic cost (Lynch et al. 2021).
This topic has contemporary interest in New Zealand where unique indigenous grasslands existed largely without herbaceous legumes and entirely without grazing mammals, thus providing a near unique location to study this particular type of plant functional complementarity. Pastoral farming systems first arrived with European settlers in the 19th Century, and clovers were introduced less than 100 years ago (Caradus 1996). Indigenous tussock grasslands currently account for about 13% of land cover, with a further 34% of land converted to sheep, beef and dairy pasture that is largely exotic (StatsNZ 2021). Native grassland plant communities that are associated particularly with higher altitude (High Country) habitats now co-exist with grazing mammals and invasive plants species, and they are frequently impacted by nutrient top-dressing and over-sowing of exotic pasture grasses and legumes. This means they now exist as novel native grassland communities (idem Wilsey et al. 2018). Pasture containing exotic fodder species is far more responsive to enhanced fertility. Exotic legumes (including Trifolium spp. and Lotus spp.) have become widely naturalised, including in high country native tussock grasslands, potentially threatening more than one-third of the country’s native plant species, of which 93% are endemic in the High Country (Mark and Galloway 2012). Herbaceous legumes are virtually absent from native flora and this family is restricted to only a relatively small number of species: 34 species that are mostly trees and shrubs, compared to at least 113 species of fully naturalized exotic legumes (Wardle 1991; Allen and Lee 2006; Dollery et al, 2019).
Graminaceous species are known to release phytosiderophores, which promote absorption of Fe and Zn (Erenoglu et al. 2000; Banakar et al. 2017). There are only a few examples where this has been shown to benefit adjacent species; for example, both Fe and Zn acquisition are increased in peanuts when they are grown with maize (Inal et al. 2007; Zuo and Zhang 2008; Xiong et al. 2013) and in guava growing with grasses (Kamal et al. 2000). However, there appear to be few insights into whether legumes similarly receive nutritional benefits from grasses in grassland pastures.
The aim of the present study was simply to investigate whether native and exotic grasses provide nutritional benefits to invasive legumes, rather than this relationship being a one-way provision of nitrogen to grasses. This is investigated using glasshouse experiments and field sampling.
Materials and methods
Plants and soils
The legumes selected for this study were white clover (Trifolium repens L.) and birdsfoot trefoil (Lotus pedunculatus Cav.), both of which have been widely seeded in New Zealand and elsewhere. White clover is the main legume of temperate pastures and meadows (Rochon et al. 2004), planted in New Zealand since the late 1920s alongside several other species of annual and perennial clovers (Rattray 2005). Lotus is also commonly sown on hill country in New Zealand, particularly at wetter, more acid, or more infertile sites (Armstrong 1974). Two common exotic pasture species of grasses and one native tussock grass were selected: perennial ryegrass (Lolium perenne), cocksfoot (Dactylis glomerata) and silver tussock (Poa cita). Ryegrass is the most productive, palatable and digestible grass species under fertile conditions and is also the most widely planted species in New Zealand but is less suited to drier locations and the high country. Cocksfoot is more hardy and drought-tolerant (Sanada et al. 2010). Poa cita is a common tussock grass species particularly suited to exposed, windy locations and dry soil (Hunter and Scott 1997), and is widespread in natural grasslands and less developed grazed pasture. Ryegrass and cocksfoot were grown from seed and Poa tussock seedlings were collected from the Lincoln University campus where the species has been widely planted.
A previously-identified nutritionally-poor soil, vegetated largely by more hardy shrubs and grasses with no known history of fertilization, was collected (1–20 cm depth) from a site near Lake Lyndon in Canterbury (altitude 611 m) described by Gutierrez-Gines et al. (2019). The soil was thoroughly mixed, then air-dried and sieved (2 mm). Samples were analysed using standard methods by Analytical Services, Soils and Physical Sciences Department, Lincoln University (Table 1), showing a range of key determinants (pH, calcium, sulphate-sulphur, soluble phosphorus, copper, magnesium, manganese) were less than optimum for plant growth. Available phosphorus, calcium and boron were extremely deficient.
In a smaller sampling exercise, white clover was also sampled from a planted lowland pasture on a more fertile soil on the university campus (Table 1). This was an attempt to investigate whether differences in foliar nutrient concentrations of clover differ if the plants are established in single-species patches of clover, compared to when it is growing in assemblages with other species.
Pot experiment
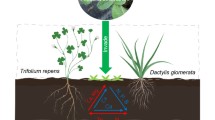
A pot experiment was located in a glasshouse at the Lincoln University Nursery. All legumes and grasses were initially propagated and established in seed trays, then transplanted, singly or in combination with each other, into (15 cm diameter, 20 cm height, 3.5L) plastic pots (Fig. 1). Eleven experimental treatments consisted of three grasses and two legumes growing either singly (5 species) or in six paired combinations (Clover & Ryegrass; Clover & Cocksfoot; Clover & Tussock; Lotus & Ryegrass; Lotus & Cocksfoot; Lotus & Tussock). Each treatment had 5 replicates (11 × 5 = 55 pots), arranged in a randomized single block design on a glasshouse bench with natural daylight. Pots were watered each day to maintain field capacity. Mean glasshouse temperatures for the duration of the experiment were 18.2 °C (range 14.2 – 32.1 °C). After four months’ growth, above-ground plant biomass was harvested, sorted into separate species, then dried (65 °C, 48 h), weighed and finely ground, prior to microwave digested using 5 M nitric acid and chemical analysis using ICP-OES following standard methodology.

Legumes and grasses were grown as either single or mixed species
Field sampling
White clover foliage was sampled from the lowland pasture on the university campus which was a fenced enclosure containing ryegrass, cocksfoot, white clover and few other species apart from plantain (Plantago lanceolata L.). The site had been planted about 5 years previously on a Wakanui soil type consisting of silt/sandy alluviums that are widespread in the Canterbury lowlands on South Island (Molloy 1998). The paddock had no recent fertilizer application or grazing. Clover foliage was sampled where it was growing in patches alone, or else in patches with cocksfoot, ryegrass or multiple assemblages of species. Each sample was excised at the ground surface and consisted of approximately 20 leaves and petioles, without attached stolons. Soil contamination of the samples was carefully avoided. Bulked samples each consisting of 2 separate samples were replicated 5 times. Samples were analysed as described above.
Statistical analysis
Data not normally distributed were log-transformed before analysis. Differences between means were determined using one-way ANOVA, with a post-hoc Fisher LSD test. All analyses were conducted using Minitab 19.
Results
Pot experiment
Clover grew better in combination with cocksfoot, and Lotus grew better in combination with ryegrass (Fig. 2). None of the grasses had significantly different yields with or without legumes; yields of grasses were in the range of 1.36 – 1.96 g pot −1 (c.f. 4.0 – 4.9 g pot −1 for legumes). There were significant differences in nutrient concentrations in the foliage of the two functional types (legumes, grasses) according to whether they were growing singly or in combination. In the legumes (Table 2), this was mostly reflected in higher concentrations of P, K and S, and lower concentrations of Ca, when growing with at least one of the three grasses. Manganese concentrations in foliage were also lower in the legumes when they were growing with cocksfoot. Nitrogen concentrations in legume foliage were not significantly different, whether or not the legumes were growing with grasses.

Yield (g. dry wt. pot −1) of each species growing alone or in combination in the pot experiment after 4 months growth (Open bars show species growing alone. Shaded bars show clover and lotus yield when growing together with ryegrass (grey), cocksfoot (dark grey) or tussock (black)). Statistical differences (*, P < 0.05) are shown for growth of clover and Lotus grown alone or in combination
When grasses were growing with or without legumes (Table 3), there was no evidence that N concentrations increased in grass foliage, although P concentrations were frequently lower (Table 3). Otherwise, elemental concentrations in the foliage of the three grasses showed few consistent significant differences when growing with legumes. In ryegrass, only K and S concentrations were significantly higher when this species was growing with one of the legumes, but five of the 13 elements were in lower concentrations. In cocksfoot, only Ni and Zn were elevated when it was growing with clover. In tussock, six elements (N, P, K, Ca, B, Zn) were in lower foliage concentrations in grasses when growing together with one of the legumes.
The data were also analysed to show total uptake of each element into grasses or legumes, calculated as biomass x element concentration after 4 months’ growth (Fig. 3). As well as having higher tissue concentrations of P, significantly more P was taken up by clover foliage when it was growing with cockfoot or tussock, and seven other elements were taken up in larger amounts when it was growing with cocksfoot. Total uptake of both P and K into Lotus was higher when this legume grew with ryegrass. When growing with clover, K and S uptake into ryegrass was higher, and Zn and Ni uptake were higher in cocksfoot. In tussock, P, B and Zn uptake into cocksfoot and tussock were lower when they were growing with legumes.

Total foliar content of selected nutrients in legumes and grasses, growing either alone or with each other (companion species are shown in the keys in brackets)
An estimate of the total acquisition of nutrients from the soil pool contained within the pots was calculated by adding together data for all of the foliage contained in each pot. Total offtake of nutrients often was not always simply explained by higher biomass yields or higher tissue concentrations, but by a combination of both (Fig. 4). When clover grew with cocksfoot, a 30% larger harvest of biomass corresponded to nearly 50% greater extraction of P from soil, and substantially more Mn, Zn and Mo.

Differences in the total harvest of vegetation and nutrients from pots when clover (A) or lotus (B) were growing with companion species. Symbols descriptions: squares = P; triangles = Mn; circles = Zn; diamonds = Mo. Open symbols are the legume with ryegrass; shaded symbols are the legume with native tussock; black symbols are the legume with cocksfoot. Percentage change is the difference to when the legumes grew alone
Field sampling
When clover grew with any of the companion plant species in a field situation, K and Mn were consistently in higher concentration in its foliage (Table 4). In some cases, Ca, B and Cu were also higher, but foliar concentrations of P and Mo were lower in clover when growing with cocksfoot.
Discussion
Legume-grass assemblages are usually more productive than either of these plant functional types in monoculture (Sleugh 2000; Sturludóttir et al. 2014). This is an example of transgressive overyielding in which functional differences between species lead to a strong complementarity (Schmid 2008; Hooper and Dukes 2004). It is generally understood that productivity is enhanced by better spatial and temporal capture of light in the mixed-species canopy, and of soil resources below-ground (Gliessman 2015; Homulle et al. 2021). In particular, nitrogen fixation in legumes, spillover of this nutrient to soil and its exploitation by adjacent species has provided an explanation for overyielding in grassland pastures (Høgh-Jensen and Schjoerring 2001; Gylfadóttir et al. 2007; Scott et al. 2018). Little attention appears to have been given to the possibility that grasses may also improve the nutritional status of legumes through modification of soil biogeochemistry, but the findings of the present study suggest this is the case.
Clover and Lotus had higher yields when growing with grasses than when growing alone. No more or less N was measured in foliage of any of the species apart from the native tussock grass that actually acquired less nitrogen when growing with legumes. Otherwise, there were no significant differences in the total foliar content of N. However, as was also indicated by the routine soil analysis, N did not appear to be an important nutrient limiting plant growth in this soil. The experimental conditions provided adequate soil moisture for growth which is likely to have favoured the growth of legumes over grasses, and there may have been little investment in root nodules and reliance on N from this source.
At least six elements (P, S, Ca, Mg, Mn, and B) were deficient in the pot experiment soil. There was evidence from the present study that all these elements, and also Mo, were extracted in higher concentrations, quantities or total amounts from the total soil pool by a combination of legumes and grasses, compared to either plant type alone. The best example was of clover growing with cocksfoot; eight elements were taken up in larger amounts into clover foliage. Field sampling of a more fertile soil similarly showed significantly higher uptake of K, Mn, Cu and B, which suggests compatibility is a real phenomenon in the field situation in grasslands.
Phosphorus, K and S concentrations in the foliage of legumes were higher in the presence of grasses. Foliar concentrations of K and S were also higher in ryegrass when it was growing with legumes. Otherwise, key elements were in lower foliar concentrations in grasses when they were growing with clovers. This was most evident in the native tussock where N, P, K, Ca, B and Zn were in lower foliage concentrations when growing with legumes. Grasses consistently extracted lesser amounts of the total soil pool of nutrients when growing alone. The native tussock grass did not benefit nutritionally from the presence of legumes, but higher P concentrations were found in tussock foliage and, as a companion plant, clover had a higher concentration of P when it was growing with tussock. It appears that tussock grass has some ability to mobilise P in its rhizosphere.
Possible mechanistic explanations for some of these findings are provided in the scientific literature. Phosphorus is a particularly critical element due to its low solubility and mobility in soil (Scott and Condron 2003; Saleem et al. 2020); P availability in the rhizosphere is increased by secretion of organic exudates and also by the release of protons, the latter particularly in acid soil (Cu et al. 2005). Evidence of partner plants being complementary to one another in the context of exploiting this element has been found previously (Homulle et al. 2021; Lambers et al. 2021). Potassium was not deficient in the experimental soil, but it is known that legumes have a higher K absorption efficiency than grasses (Wang et al. 2014), which would suggest legumes have an advantage in competition for K when growing with grasses. Root exudates containing glutamic acid, tyrosine, and leucine increase K uptake from the soil. In the pot experiment of the present study, K concentrations in Lotus and grass foliage were both higher when they were growing together rather than alone. In the field sampling, clover had higher foliar K concentrations when it was growing with grasses. This may be due to K mobility being increased in the grass rhizosphere soil, but then shared with the legumes.
Of course there are also functional interactions between physico-chemical variables. The interaction of S with other elements through its modification of rhizosphere conditions generally exceed its own direct nutritional value, but legumes require more S than grasses and microbial activity associated with grass rhizospheres accelerate the oxidation and mineralization of organic S into S2− or SO42− (Wainwright 1984). When soil pH is lower, soil redox potential decreases and solubility of Fe and Mn increases. Grasses also respond to Fe, Zn, Mn and Cu deficiency with enhanced siderophore release from roots (Marschner and Römheld 1994; Erenoglu, et al. 2000). Uptake of Zn, Mn and Cu is poorly controlled in plants (Lambers et al. 2021). In the present study, clover had higher foliar concentrations of these elements when it grew with ryegrass and tussock. Clover-cocksfoot and Lotus-ryegrass assemblages procured more Mn in the present study. Also, in the field sampling of the present study, when clover grew with any of the grass species or multi-species assemblages, foliage concentrations of Mn were higher.
Phytosiderophores may have played a key role in higher Cu and B concentrations in clover foliage when growing with cocksfoot, but mechanistic explanations are likely to be more complex. Understanding Zn deficiency provides a good example (Hafeez et al. 2013). Each essential nutrient has specific physiological and biochemical roles; Zn influences the nitrogen content of legumes (Buerkert et al. 1990; Bolanos et al. 1994) and rhizobia can mobilise Zn in soil to some extent (Chen 2003). The results of the present study indicate that cocksfoot mobilized Zn and its uptake when growing by itself. Clover may then have consumed mobile Zn, limiting availability of this element to cocksfoot, in turn stimulating cocksfoot to synthesize and secrete mugineic acids. Molybdenum is known to be an important trigger for photosynthesis and N metabolism (Imran et al. 2021), and this element was better exploited from the soil pool by mixtures of legumes and grasses in the pot experiment of the present study. Similarly, Mg is known to improve N-use efficiency (Tian et al. 2021); foliar concentrations of Mg were lower in grasses in the presence of legumes.
It is recognised that it is difficult to identify the specific plant traits that explain species complementarity and transgressive overyielding (Hooper and Dukes 2004; von Felten and Schmid 2008), and clearly nutrient mobility in soil and acquisition by plants is particularly complex. The present study lacks any investigation or analysis of the root systems in the pot experiment or field site, but it is known that there is a substantial rhizophere priming effect on nutrient cycling (Lu 2020). Variations between plant species in root exudation pattern and efficiency of acquiring nutrients has long been discussed (Bardgett et al. 1999), and combinations of clovers and grasses have been shown to increase microbial enzyme activity in soil and release of nutrients from organic matter (Sekaran et al. 2020). Findings in the present study relating to each of the elements that were analysed are hardly definitive, but the body of evidence presented clearly illustrates benefits to legumes through partnership with grasses. Productivity is increased and nutrient acquisition from the soil is enhanced by neighbouring grasses that have a complimentary role. Increases of both productivity and foliar concentrations showed a greater total offtake of P, Zn, Mo and Mn from the soil nutrient pool.
In a broader practical context, increasing the legume component of pasture has been a goal of high-country farming in New Zealand for several decades by attempting to introduce a wide range of exotic clovers and other herbaceous legumes to increase vegetation yields and stock productivity. The findings of the present study indicate that cocksfoot and other grasses, including native tussock, play a mutually beneficial role that previously has been largely overlooked. Native tussock grass contained lesser amounts of N and five other nutrients when it was growing with the exotic legumes, suggesting less compatibility, a lack of adaptation to coexistence and perhaps to competition for soil N. Exotic herbaceous legumes appear to offer little benefit to tussock grasses, but sustaining residual assemblages of indigenous species within the productive farming landscape is hugely important to biodiversity conservation in New Zealand. This study suggests it would be worthwhile to elucidate functionality of plant diversity in a wider range of native plants in terms of biodiversity protection, soil biogeochemistry and efficient exploitation of nutrient resources.
Conclusions
Legume nutrition is improved by neighbouring grasses, and coexistence of these functional groups provides more efficient soil biogeochemistry. This manuscript appears to be a first report that spillover of N from clovers to grasses is an adaptation and strategy to procure quid pro quo other limiting nutrients from neighbouring grasses. In a nutrient-poor soil, productivity increased and the mobility of key elements in plants and soils significantly differed with combinations of legumes and grasses, compared to either functional group alone. Foliar concentrations differed and mass balance results demonstrated that mixed species assemblages improved the efficiency of exploitation of the soil pool of nutrients. White clover and Lotus grew better in combination with different species of introduced grasses. The results provide a demonstration of transgressive overyielding. Sampling of clover in a more fertile lowland pastoral grassland provided results that suggested mutual compatibility between legumes and other species is a more widespread and frequent phenomenon. Mechanistic explanations for these responses were not sought in this experimental work, but numerous potential explanations exist in the scientific literature; it is likely that modification of Ca and S and subsequently pH in the rhizosphere, which is further modified by different root structures and root exudates play key roles. These findings have obvious significance towards understanding the value of plant diversity in pastoral farming, but they also advance knowledge of native and exotic plant co-existence that is highly relevant to conservation and management of New Zealand’s native biodiversity. The indigenous tussock grass contained less N and no other nutritional benefits when growing with exotic legumes, but did improve the overall offtake of nutrients. This suggests less compatibility, a lack of adaptation to coexistence and a limited ability to compete for soil N. Further research is justified.
Data availability
Not applicable.
Code availability
Not applicable.
References
Allen RB, Lee WG (2006) Biological invasions in New Zealand. Springer Science & Business Media.
Armstrong C (1974) Grasslands Maku tetraploid lotus (Lotus pedunculatus Cav.). N Z J Exp Agric 2:333–336
Banakar R, Alvarez Fernandez A, Díaz-Benito P, Abadia J, Capell T, Christou P (2017) Phytosiderophores determine thresholds for iron and zinc accumulation in biofortified rice endosperm while inhibiting the accumulation of cadmium. J Exp Bot 68:4983–4995
Bardgett R, Mawdsley J, Edwards S, Hobbs P, Rodwell J, Davies WJ (1999) Plant species and nitrogen effects on soil biological properties of temperate upland grasslands. Funct Ecol 13:650–660
Bolanos L, Esteban E, de Lorenzo C, Fernandez-Pascual M, de Felipe MR, Garate A, Bonilla I (1994) Essentiality of boron for symbiotic dinitrogen fixation in pea (Pisum sativum) rhizobium nodules. Plant Physiol 104:85–90
Buerkert A, Cassman K, De la Piedra R, Munns D (1990) Soil acidity and liming effects on stand, nodulation, and yield of common bean. Agron J 82:749–754
Burrows CJ (1990) Processes of vegetation change. Unwin Hyman, Boston
Caradus J (1996) Overview and vision for white clover. In: Woodfield DR (ed) White clover: New Zealand’s competitive edge. New Zealand Grassland Association: Palmerston North, pp. 1–6.
Chen W, Huang Q, Guo X (2003) Effects of rhizobia on morphological distribution of Cu, Zn and Cd in soil. J App Ecol 14:1278–1282
Craine JM, Dybzinski R (2013) Mechanisms of plant competition for nutrients, water and light. Funct Ecol 27:833–840
Cu ST, Hutson J, Schuller KA (2005) Mixed culture of wheat (Triticum aestivum L.) with white lupin (Lupinus albus L.) improves the growth and phosphorus nutrition of the wheat. Plant Soil 272:143–151
Dollery R, Li S, Dickinson NM (2019) Nutrient-enriched soils and native N-fixing plants in New Zealand. J Plant Nutr Soil Sci 182:104–110
Erenoglu B, Eker S, Cakmak I, Derici R, Römheld V (2000) Effect of iron and zinc deficiency on release of phytosiderophores in barley cultivars differing in zinc efficiency. J Plant Nutr 23:1645–1656
Fornara D, Tilman D (2008) Plant functional composition influences rates of soil carbon and nitrogen accumulation. J Ecol 96:314–322
Gliessman S (2015) Agroecology: the ecology of sustainable food systems, 3rd edn. Taylor & Francis, Boca Raton
Gutiérrez-Ginés MJ, Madejón E, Lehto NJ, McLenaghen RD, Horswell J, Dickinson N, Robinson BH (2019) Response of a pioneering species (Leptospermum scoparium JR Forst. & G. Forst.) to heterogeneity in a low-fertility soil. Front Plant Sci 10:1–12
Gylfadóttir T, Helgadóttir Á, Høgh-Jensen H (2007) Consequences of including adapted white clover in northern European grassland: transfer and deposition of nitrogen. Plant Soil 297:93–104
Hafeez B, Khanif Y, Saleem M (2013) Role of zinc in plant nutrition-a review. Journal of Experimental Agriculture International: 374–391.
Høgh-Jensen H, Schjoerring JK (2001) Rhizodeposition of nitrogen by red clover, white clover and ryegrass leys. Soil Biol Biochem 33:439–448
Homulle Z, George T, Karley A (2021) Root traits with team benefits: understanding belowground interactions in intercropping systems. Plant and Soil: 1–26. [https://doi.org/10.1007/s11104-021-05165-8]
Hooper DU, Dukes JS (2004) Overyielding among plant functional groups in a long-term experiment. Ecol Lett 7:95–105
Hunter GG, Scott D (1997) Changes in tussock grasslands, South Island high country, 1973–93. Department of Conservation, Wellington.
Imran M, Hussain S, Rana MS, Saleem MH, Rasul F, Ali KH, Potcho MP, Pan S, Duan M, Tang X (2021) Molybdenum improves 2-acetyl-1-pyrroline, grain quality traits and yield attributes in fragrant rice through efficient nitrogen assimilation under cadmium toxicity. Ecotoxicol Environ Safe 211:111911
Inal A, Gunes A, Zhang F, Cakmak I (2007) Peanut/maize intercropping induced changes in rhizosphere and nutrient concentrations in shoots. Plant Physiol Biochem 45:350–356
Kamal K, Hagagg L, Awad F (2000) Improved Fe and Zn acquisition by guava seedlings grown in calcareous soils intercropped with graminaceous species. J Plant Nutr 23:2071–2080
Lambers H, Wright IJ, Pereira CG, Bellingham PJ, Bentley LP, Boonman A, Cernusak LA, Foulds W, Gleason SM, Gray EF (2021) Leaf manganese concentrations as a tool to assess belowground plant functioning in phosphorus-impoverished environments. Plant Soil 461:43–61
Li L, Tilman D, Lambers H, Zhang FS (2014) Plant diversity and overyielding: insights from belowground facilitation of intercropping in agriculture. New Phytol 203:63–69
Lu JY, Yang JF, Keitel C, Yin LM, Wang P, Cheng WX, Dijkstra FA (2020) Rhizosphere priming effects of Lolium perenne and Trifolium repens depend on phosphorus fertilization and biological nitrogen fixation. Soil Biol Biochem 150:108005
Lynch JP, Strock CF, Schneider HM, Sidhu JS, Ajmera I, Galindo-Castañeda T, Klein SP, Hanlon MT (2021) Root anatomy and soil resource capture. Plant Soil 466:21–63
Mark AF, Galloway DJ (2012) Above the treeline: a nature guide to alpine New Zealand. Craig Potton Publishing, Nelson
Marschner H, Römheld V (1994) Strategies of plants for acquisition of iron. Plant Soil 165:261–274
Pirhofer-Walzl K, Rasmussen J, Høgh-Jensen H, Eriksen J, Søegaard K, Rasmussen J (2012) Nitrogen transfer from forage legumes to nine neighbouring plants in a multi-species grassland. Plant Soil 350:71–84
Rattray PV, By C, SFF SFF (2005) Clover management, research, development and extension in the New Zealand pastoral industries: A review of R&D literature and R&D opportunities, extension initiatives and extension opportunities. Sustainable Farming Fund, Ministry for Primary Industries, NZ Government.
Rochon J, Doyle C, Greef J, Hopkins A, Molle G, Sitzia M, Scholefield D, Smith C (2004) Grazing legumes in Europe: a review of their status, management, benefits, research needs and future prospects. Grass Forage Sci 59:197–214
Saleem M, Pervaiz ZH, Contreras J, Lindenberger JH, Hupp BM, Chen D, Zhang Q, Wang C, Iqbal J, Twigg P (2020) Cover crop diversity improves multiple soil properties via altering root architectural traits. Rhizosphere 16:100248
Sanada Y, Gras M, Van Santen E (2010) Cocksfoot. In: Boller BU, Posselt K, Veronesi DF (eds) Fodder crops and amenity grasses: Handbook of plant breeding. Springer, New York, pp 317–328
Schipanski ME, Drinkwater LE (2012) Nitrogen fixation in annual and perennial legume-grass mixtures across a fertility gradient. Plant Soil 357:147–159
Schmid B, Hector A, Saha P, Loreau M (2008) Biodiversity effects and transgressive overyielding. Journal of Plant Ecology 1:95–102
Scott JT, Condron LM (2003) Dynamics and availability of phosphorus in the rhizosphere of a temperate silvopastoral system. Biol Fertil Soils 39:65–73
Scott GD, Baddeley JA, Watson CA, Robinson D (2018) Demographic quantification of carbon and nitrogen dynamics associated with root turnover in white clover. Plant, Cell Environ 41:2045–2056
Sekaran U, Loya JR, Abagandura GO, Subramanian S, Owens V, Kumar S (2020) Intercropping of kura clover (Trifolium ambiguum M Bieb) with prairie cordgrass (Spartina pectinata link) enhanced soil biochemical activities and microbial community structure. App Soil Ecol 147:103427
Sleugh B, Moore KJ, George JR, Brummer EC (2000) Binary legume–grass mixtures improve forage yield, quality, and seasonal distribution. Agron J 92:24–29
StatsNZ (2021) Agricultural and horticultural land use. StatsNZ Tatauranga Aotearoa. https://www.stats.govt.nz/indicators/agricultural-and-horticultural-land-use. Accessed 24 Jan 2022
Sturludóttir E, Brophy C, Belanger G, Gustavsson AM, Jørgensen M, Lunnan T, Helgadóttir Á (2014) Benefits of mixing grasses and legumes for herbage yield and nutritive value in N orthern E urope and C anada. Grass Forage Sci 69:229–240
Tian S, Zhang Y, Yang C, Tie S, Nan J (2021) Nitrogen-doped carbon nanosheet coated multilayer graphite as stabilized anode material of potassium-ion batteries with high performances. Electrochimica Acta 380:138254
Trinder CJ, Brooker RW, Davidson H, Robinson D (2021) Directly quantifying multiple interacting influences on plant competition. Plant, Cell Environ 44:1268–1277
von Felten S, Schmid B (2008) Complementarity among species in horizontal versus vertical rooting space. Journal of Plant Ecology 1:33–41
Wainwright M (1984) Sulfur oxidation in soils. Adv Agron 37:349–396
Wang Z, Lu J, Yang H, Zhang X, Luo C, Zhao Y (2014) Resorption of nitrogen, phosphorus and potassium from leaves of lucerne stands of different ages. Plant Soil 383:301–312
Wardle P (1991) Vegetation of New Zealand. Cambridge Universioty Press, Cambridge
Wilsey BJ, Martin LM, Kaul AD (2018) Phenology differences between native and novel exotic-dominated grasslands rival the effects of climate change. J Appl Ecol 55:863–873
Xiong H, Kakei Y, Kobayashi T, Guo X, Nakazono M, Takahashi H, Nakanishi H, Shen H, Zhang F, Nishizawa NK (2013) Molecular evidence for phytosiderophore-induced improvement of iron nutrition of peanut intercropped with maize in calcareous soil. Plant, Cell Environ 36:1888–1902
Zhang WP, Liu GC, Sun JH, Zhang LZ, Weiner J, Li L (2015) Growth trajectories and interspecific competitive dynamics in wheat/maize and barley/maize intercropping. Plant Soil 397:227–238
Zuo Y, Zhang F (2008) Effect of peanut mixed cropping with gramineous species on micronutrient concentrations and iron chlorosis of peanut plants grown in a calcareous soil. Plant Soil 306:23–36
Acknowledgements
The research team are grateful to the Miss E.L. Hellaby Indigenous Grasslands Research Trust for grant funding to support Zhang Wei for his PhD stipend and operational funds. Special thanks also to Dasom Jeon, Brent Richards and Roger Cresswell for practical support.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions This study was supported by Lincoln University and the Miss E.L. Hellaby Indigenous Grasslands Research Trust.
Author information
Authors and Affiliations
Contributions
All authors contributed to the planning and execution of the project, and to manuscript preparation. Zhang Wei carried out the practical work and data analysis as part of his PhD programme supervised by Nicholas Dickinson, Thomas Maxwell and Brett Robinson. Nicholas Dickinson is responsible for the final manuscript draft.
Corresponding author
Ethics declarations
Conflict of interest
There are no conflicts of interest.
Additional information
Responsible Editor: Peter Christie.
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wei, Z., Maxwell, T.M.R., Robinson, B. et al. Legume nutrition is improved by neighbouring grasses. Plant Soil 475, 443–455 (2022). https://doi.org/10.1007/s11104-022-05379-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-022-05379-4