
Abstract
Background and aims
The reasons for partial recovery after a soil drought are not fully understood and have not been studied so far. This study investigated the physiological and biochemical responses of triticale cultivars with differential recovery ability after soil water deficit.
Methods
Activity of the photosynthetic apparatus under soil drought followed by rehydration was estimated. Plant antioxidant potential was determined based on the measurement of catalase and peroxidase activity. The levels of hydrogen peroxide and superoxide radical were assessed.
Results
Under rehydration, the not fully-recovered cultivar experienced further significant increase in the content of H2O2 and inhibited activity of the photosynthetic apparatus, as compared to the drought period. On 42nd day of the rehydration, the not fully-recovered cultivar showed also a reduced photosynthetic activity in the flag leaves, which resulted in a significant decrease in its grain yield. The first week of a rapid rehydration involved a decrease in total peroxidase and catalase activities. The increased content of H2O2 was compensated only when leaf water content was gradually restored in the first week of the rehydration and no further decrease in the activity of the photosynthetic apparatus was noticed.
Conclusions
A destructive effect of the rapid rehydration was manifested in an intensification of the physiological processes associated with reactive oxygen species (ROS) overproduction. An important cause of hydrogen peroxide overproduction seems to be the electron leakage due to overloading of the electron transport chain (ETC) in the PSI and PSII.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Soil drought is one of disadvantageous environmental factors significantly affecting the productivity of cereal crops (Cannell et al. 1984; Shah and Paulsen 2003; Hura et al. 2007a). The extent of grain yield loss depends on the intensity of the soil drought, i.e., whether the drought is mild or acute, its duration, frequency, plant growth stage during which adverse soil moisture conditions occur, and the efficiency of plant rehydration after the soil drought ceases (Souza et al. 2004; Barnabás et al. 2008).
Rehydration, i.e., favorable soil moisture conditions following a drought (Beckett et al. 2012), is a period when plant physiological activity can return to the optimum, or even rise above the normal level, as in the case of the photosynthetic apparatus (Hura et al. 2009a, b, c). However, rapid plant rehydration may also cause adverse effects, such as an oxidative burst of hydrogen peroxide. This is often observed in mosses and lichens following a desiccation period, and the main reasons for reactive oxygen species generation seem to be extracellular and intracellular peroxidases, high light intensity or specifically localized the plasma membrane NADPH and NADH (Minibayeva and Beckett 2001; Mayaba et al. 2002; Weissman et al. 2005). Other studies showed that full recovery of plants when restoring favorable soil moisture conditions also depended on the mechanisms related to hydraulic failure and carbon starvation (McDowell et al. 2008; Sala et al. 2010; O’Grady et al. 2013; Sevanto et al. 2014).
Full recovery of crops after a drought means restoring normal cellular metabolism, manifested by the yield corresponding to optimal conditions of plant growth. In the case of winter triticale, soil drought at the heading or anthesis stage causes the greatest yield loss (Saini and Westgate 2000; Yang and Zhang 2006). Restoring favorable soil moisture conditions does not result in full recovery of the crops, as the plants are at the final growth stages. Contrary to that, full recovery of winter triticale is very often observed following the soil drought occurring at the tillering stage (Hura et al. 2006; 2009a, b, c). However, for some cultivars, even a long rehydration period after a drought experienced at the tillering stage is still insufficient and is manifested by unexpectedly large grain yield reduction.
The reasons for partial recovery after a soil drought are not fully understood and have not been studied so far. Majority of papers on rehydration emphasize its ‘beneficial’ effects on plant growth following a drought cessation (Srivalli et al. 2003; Hura et al. 2006, 2009a; Lovisolo et al. 2008; Beckett et al. 2012; Furlan et al. 2014). There are few publications addressing the adverse effects of the rehydration period on plant regeneration following the soil drought. Therefore, the aim of the experiment presented in this paper was to determine the physiological and biochemical responses of triticale cultivars with differential recovery ability after soil water deficit experienced at the tillering stage. To evaluate whether rapid rehydration causing intensification of metabolic processes could be the only source of hydrogen peroxide burst, we hypothesized that permanent and adverse changes in the activity of the photosynthetic apparatus are initiated by rapid rehydration following the drought, regardless light availability. This research hypothesis was verified in experimental conditions allowing for assessing the effects of light and different rehydration rates at which optimum soil water content was restored. Our study focused on the level of hydrogen peroxide, which on the one hand is considered to be a biological indicator of stress intensity (Bartosz 1997; Blokhina et al. 2002; Hura et al. 2014), and on the other hand is a secondary messenger in the signal transduction pathways leading to gene expression in response to adverse environmental factors (Orozco-Cárdenas et al. 2001; Maksymiec 2007).
Materials and methods
Plant material
As presented in Table 1S, the experiment included one not fully-recovered cultivar ‘Kazo’ and one fully-recovered cultivar ‘Hortenso’ of winter triticale (x Triticosecale Wittmack). These cultivars are not significantly different in terms of physiological and biochemical responses to drought stress. The data presented in the table are the results of pot experiments conducted under controlled greenhouse conditions in the years 2008–2010. The seeds were obtained from ‘DANKO’ Plant Breeders Ltd., Choryń, Poland. The part of the experiment concerning plant growth was carried out in air-conditioned chambers and greenhouses of the Department of Plant Physiology, Faculty of Agriculture and Economics at the University of Agriculture in Kraków, Poland.
Plant growth conditions
The seeds of ‘Kazo’ and ‘Hortenso’ cultivars were sown into plastic pots 3.7 L in volume. Before sowing, the pots were tared to equal weight and filled with a homogeneous mixture of soil and sand (1:3; v/v). The seeds were sown with a surplus, i.e., 9 seeds per pot, and after the emergence, the number of plants per pot was limited to 7.
The experiment covered an entire growth cycle of winter triticale from germination to ripening. Therefore, the seedlings at the stage of 2 leaves were subjected to vernalization in cold chambers, lasting for 7 weeks at +3 °C (±1 °C), and PPFD (photosynthetic photon flux density) of 150 μmol m−2 s−1, photoperiod 10 h. After the vernalization, the plants at the 4-leaf stage were transferred into air-conditioned greenhouse chambers. Air temperature in the greenhouse was 28/18 °C (±2 °C) day/night, and relative air humidity was about 40 %. The plants were additionally illuminated to ensure that the PPFD was about 250 μmol m−2 s−1 (provided by high pressure sodium lamps, 400 W; Philips SON-T AGRO, Brussels, Belgium), at the level of the top leaf. Light intensity at the leaf level was measured with a QSPAR Quantum Sensor (Hansatech Instruments LTD, Kings Lynn, England). The plants were irrigated once a week with full-strength Hoagland’s nutrient solution (Hoagland 1948).
Drought conditions
The treatment of low water availability was initiated when the plants of both cultivars developed three leaves on the main shoot and two lateral shoots with 3 and 2 leaves (8 leaves in total). Water content in the pots was gradually lowered to 30 % (about -5.46 MPa) by not watering the plants for 7 days, and these conditions were maintained for the next 2 weeks. Water content in the control pots was maintained at 75 % (about -0.31 MPa). Water content in the pots for each treatment was set based on the maximum water holding capacity of soil and sand mixture. Soil moisture level in the pots was monitored daily between 8 and 10 a.m., using the gravimetric method and taking into account the weight of the plants. An increase in the fresh weight of the plants was determined at the harvest to adjust the correct level of soil moisture.
Rehydration
By the end of the soil drought period both cultivars developed a 5-leaf main shoot and one 3-leaf lateral shoot (8 leaves in total). Control plants of both cultivars developed a 5-leaf main shoot and two lateral shoots with 4 and 3 leaves (12 leaves in total).
The soil drought at the tillering stage was followed by three different schemes of rehydration:
-
Rehydration 1 (R1). Immediate restoration of soil water content up to 75 % for PPFD of 250 μmol m−2 s−1.
-
Rehydration 2 (R2). Gradual restoration of soil water content up to 75 % for PPFD of 250 μmol m−2 s−1. The soil water content was maintained at 40 % for the first 2 days, then at 50 % for the next 2 days, at 60 % for 1 day and at 75 % for the last 2 days.
-
Rehydration 3 (R3). Immediate restoration of soil water content up to 75 % for PPFD of 25 μmol m−2 s−1.
R2 and R3 treatments were applied only in the case of not fully-recovered ‘Kazo’ cultivar.
Measurements
The analyses were performed 14 days after the pot soil water content reached 30 %. Additionally, 1, 3 and 7 days after the rehydration, the activity of peroxidase and catalase was evaluated, and hydrogen peroxide and water content in the leaves were assessed. Chlorophyll fluorescence, gas exchange and emission intensity of blue and red fluorescence were measured 7 days after the rehydration. The analyses involved the second (from the top) fully developed leaf. Furthermore, on 42nd day of the rehydration, the measurements were repeated for the flag leaves at the anthesis stage in both cultivars. Total catalase and peroxidase activity was recalculated for protein content according to Bradford (1976).
Soil water content (SWC)
The soil water content was estimated by measuring the fresh weight of the soil without plant residues and then the dry weight after 24 h of drying in an oven at 90 °C. Soil samples (100 g) were taken from three pots within treatments, every other day for 2 weeks in the drought variant and every day for 7 days since the end of the drought period in the rehydration variant. SWC analysis in the control samples was run in parallel for the last 7 days of the drought period and 7 days of the rehydration every other day. When the plants were in the generative phase, SWC was analyzed on 36th, 38th, 40th and 42nd day of the rehydration (data not shown).
Midday relative water content (RWC) and predawn leaf water potential (Ψw)
Relative water content (RWC) of the leaves was calculated as per the following equation: RWC(%) = (fw ‐ dw)/(tw ‐ dw) × 100. The leaves were weighed (fw - fresh weight), and then soaked in freshly deionized water for 12 h in darkness at 5 °C and their turgid weight (tw) was determined. The samples were then oven-dried at 80 °C for 72 h and weighed (dw - dry weight) (Barrs and Weatherley 1962; Turner 1981).
Measurements of leaf water potential were taken with a psychrometer HR 33 T (WESCOR, Inc., Logan, Utah, USA), equipped with leaf sample chambers C-52 (WESCOR, Inc., Logan, Utah, USA). The leaf discs (collected from the central part of a leaf blade), 5 mm in diameter, were placed in C-52 chambers and left for 60 min. The measurements were taken in the dew point mode.
Pre-dawn samples were collected at 6:00 to 7:00 a.m., while midday samples were collected at 12:00 to 1:00 p.m.
Chlorophyll fluorescence measurements
Chlorophyll fluorescence analysis during the drought and rehydration at the tillering stage was carried out using a fluorometer Handy PEA (Hansatech Ltd. Kings Lynn, UK). Data were scored after 25 min of leaf adaptation to darkness. Fv/Fm (quantum yield of PSII) was calculated according to van Kooten and Snel (1990) as (Fm - Fo)/Fm, where Fo and Fm represent the minimal and maximal chlorophyll fluorescence, respectively. The excitation irradiance was 3000 μmol (quanta) m−2 s−1 (peak at 650 nm). Fluorescence was recorded during irradiation between 10 μs and 1 s. During the initial 2 ms, the data were scored every 10 μs with a 12 bit resolution. The following parameters were calculated per excited leaf cross-section (CSm): PI (overall performance index of PSII photochemistry), DIo/CSm (energy amount dissipated from PSII), RC/CSm (number of active reaction centers), ETo/CSm (amount of energy used for the electron transport) and TRo/CSm (amount of excitation energy trapped in PSII reaction centers), ψRo (probability, at time 0, that a trapped exciton moves an electron into the electron transport chain beyond QA −), δRo (the efficiency with which an electron can move from the reduced intersystem of electron acceptors to the PSI end electron acceptors), φRo (the quantum yield of electron transport from QA − to the PSI end electron acceptors). Calculations for the parameters were based on the theory of energy flow in PSII using the JIP test (Strasser and Tsimilli-Michael 2001; Tsimilli-Michael and Strasser 2008; Strasser et al. 2010).
Chlorophyll fluorescence measurements of the flag leaves were performed with a fluorometer FMS 2 (Hansatech Instruments, Kings Lynn, UK). The leaves were adapted to darkness for 25 min. Fv/Fm (quantum yield of PSII) was calculated according to van Kooten and Snel (1990). Then, the leaves were continuously illuminated with white actinic light (400 μmol m−2 s−1). After 7 min, the fluorescence in steady state (Fs) was recorded and a second saturating pulse at 8000 μmol m−2 s−1 was imposed to determine the maximal fluorescence in the light-adapted state (Fm’). The minimal fluorescence in the light-adapted state (Fo’) was determined by brief application (3 s) of low-intensity (6 μmol m−2 s−1) far-red light (730 nm). Photochemical quenching coefficient qP = (Fm ’- Fs)/(Fm ’ - Fo ’), efficiency of excitation transfer to open PSII centers Fv ’/Fm ’ = (Fm ’ - Fo ’)/Fm ’, PSII quantum efficiency ФPSII = (Fm ’- Fs)/Fm ’ and the electron transport rate ETR = PAR × 0.5 × ФPSII × 0.84 were calculated as in Genty et al. (1989). Non-photochemical quenching qN = 1- (Fm ’ - Fo ’)/(Fm - Fo) was calculated according to van Kooten and Snel (1990).
Fluorescence emission spectra of red (690 nm) and far-red fluorescence (740 nm) were measured using a spectrofluorometer (Perkin-Elmer LS 50B, Norwalk, USA), according to Hura et al. (2007a). Fluorescence intensity was recorded between 650 and 800 nm. The leaves were excited at 450 nm. The spectral slit width was set at 10 nm (excitation and emission).
Fluorescence images of the leaves were made using a PSI Fluorcam 700MF chlorophyll fluorescence imaging system (PSI, Brno, Czech Republic), as described by Nedbal et al. (2000). Fluorcam v. 3.5 software was used to control the imaging system and to process the images.
Leaf photosynthetic activity
Leaf gas exchange measurements were performed at ambient temperature (25 ± 2 °C) and ambient relative humidity (40 ± 5 %) in the greenhouse conditions, on fully expanded leaves, using an infrared gas analyzer (CID; CI 301 CO2; Inc. Vancouver, Washington, USA), operated within an open system with a leaf chamber. The net photosynthesis (PN), transpiration rate (E) and stomatal conductance (gs) were determined between 11:00 a.m. and 1:00 p.m. under air flow 200 mL min−1, the saturating PPFD of 1100 μmol (photons) m−2 s−1 (Hura et al. 2006, 2007b) provided by LED light module (CI-301LA, CID, Camas, WA, USA), and at air CO2 concentration of 380 ppm, provided by CO2 control unit (CI-301 AD, CID, Camas, WA, USA).
Pigment content
Chlorophyll and carotenoids content were analyzed spectrophotometrically (Ultrospec II, Biochrom, Cambridge, England), after extraction in 95 % ethanol. The concentration of chlorophyll and carotenoids was then calculated according to Lichtenthaler and Wellburn (1983).
Histochemical identification of superoxide radical
Dye-based identification of superoxide radical in the leaves was done according to Doke and Ohashi (1988). The leaves were infiltrated in darkness for 15 min under the pressure of 0.8 MPa with a solution containing 0.5 % (w/v) nitroblue tetrazolium (NBT), 10 mM potassium phosphate buffer pH 7.0, and 0.005 % (w/v) Triton X-100, and then exposed to light for 15 min. The leaves were rinsed several times with hot ethanol (96 %) to remove chlorophyll from the plant tissues. The colored spots (dark blue) of superoxide radical accumulation in the leaves were scanned (CanoScan, Lide 110). Staining was carried out on the seventh day of the rehydration.
Hydrogen peroxide content
Histochemical detection of hydrogen peroxide was performed according to Thordal-Christensen et al. (1997). The leaves were infiltrated in darkness for 15 min under the pressure of 0.8 MPa in 3,3’-diaminobenzidine (DAB) solution. DAB aqueous solution was prepared in the ratio 1 mg DAB: 1 mL H2O, and pH was adjusted to 3.8 with 0.1 M HCl. To remove chlorophyll, the leaves were rinsed with hot (60 °C) ethanol (95 %). The colored spots, indicating H2O2 accumulation in the leaves, were scanned.
Hydrogen peroxide content was analyzed according to Ishikawa et al. (1993). The leaves were homogenized in 1.4 mL of the extraction buffer containing 50 mM potassium phosphate buffer pH 7.5, 5 % trichloroacetic acid, 1 mM EDTA, and 1 % w/v polyvinylpyrrolidone. The reaction mixture consisted of 2.5 mL of 1.25 mM homovanillic acid, 2.5 μL of horseradish peroxidase (1380 U mg−1) and 20 μL of the buffered extract. H2O2 content was analyzed with a spectrofluorometer (Perkin-Elmer LS 50B, Norwalk, CT, USA). The reaction mixtures were excited at 315 nm and fluorescence emission was detected at 400–450 nm. The excitation and emission slit width of the monochromators was adjusted to 10 nm.
Total catalase (CAT) activity
Catalase activity was assessed according to Aebi (1984). Plant material (150–200 mg of fresh weight) was homogenized in the extraction buffer (50 mM potassium phosphate buffer pH 7.5, 1 mM EDTA) at 4 °C. Plant extract (100 μL) was added to the reaction mixture consisting of 0.3 mL of 30 % H2O2 in 50 mM potassium phosphate buffer pH 7.8 and 1 mM EDTA. Absorbance was analyzed at 240 nm. The enzyme activity was recalculated, assuming that absorbance drop of about 0.0145 corresponds to the degradation of 1 μmol of H2O2 within 1 min per g of protein.
Total peroxidase activity (POX)
The leaves (150–200 mg of fresh weight) were homogenized in the extraction buffer (50 mM potassium phosphate buffer pH 7.0, 1 mM EDTA) at 4 °C. The reaction mixture contained 2 mL of 50 mM potassium phosphate buffer (pH 7.8), 1 mM EDTA, 12 μL of p-phenylenediamine, 12 μL of 30 % H2O2 and 15 μL of the plant extract. Absorbance was measured at 460 nm. According to Bergmeyer (1965), increasing absorbance correlates with an accumulation of oxidized products of p-phenyldiamine in H2O2 presence. One unit of POX activity was defined as an increase in absorbance (0.01 ΔA460 = 1 EU) per 1 min per g of protein.
Excitation spectra of blue fluorescence
Blue fluorescence excitation spectra were recorded using Perkin-Elmer LS 50B spectrofluorometer, according to Hura et al. (2006). Leaf samples were excited at 250–400 nm and blue fluorescence emission was observed at 440 nm. The excitation and emission monochromator slit width was adjusted to 10 nm.
Statistical analysis
Statistical analysis was carried out using Statistica v. 9.0 (Statsoft Inc.). Analysis of variance was used to determine the main effects of treatments on the physiological parameters within each studied cultivar. Duncan’s multiple range test at 0.05 probability level was chosen to determine the significance of differences among the treatment means. Correlations between measured parameters were tested at a probability of P < 0.05.
Results
Soil water content and plant water relations during drought and rehydration
Soil water content (SWC) under optimal plant growth conditions (Fig. 1a) and during R1 and R3 rehydration (Fig. 1c) ranged between 70 and 75 %, and in the drought period it amounted to 30–35 % (Fig. 1b). For the R2 variant, SWC ranges were 40–45 %, 50–55 %, 60–65 % and 70–75 % during the rehydration period (Fig. 1c).

Soil water content (SWC) in the pots for control (a;  ), drought (b; □), and rehydration (c): R1 (○), R2 (
), drought (b; □), and rehydration (c): R1 (○), R2 ( ), R3 (△)
), R3 (△)
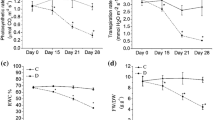
Following 3 weeks of drought, the plants of both triticale cultivars either growing in optimum water soil conditions or exposed to the rehydration showed similar leaf water content (Fig. 2). After 3 weeks of drought, the leaf relative water content (RWC) in ‘Hortenso’ and ‘Kazo’ cultivars reached 65 % (RWC) (Fig. 2a), and leaf water potential was about -2.2 MPa (Fig. 2b). Gradual restoration of soil water content in R2 variant was reflected in the gradual increase in leaf water content in the not fully-recovered cv. ‘Kazo’. After 24 h of R1 and R3 rehydration, leaf water content of both cultivars was the same as in the control plants (about 95 % RWC and −0.90 MPa).

Relative water content (RWC, [%]) (a) and leaf water potential (b, [MPa]) for fully-recovered ‘Hortenso’ cultivar and not fully-recovered ‘Kazo’ cultivar. The analyses were performed 1, 3 and 7 days after rehydration. Control-(○), drought-(●), rehydration: R1-(□), R2-( ), R3-(■). Means ± SE (n = 10 for RWC; n = 7 for leaf water potential)
), R3-(■). Means ± SE (n = 10 for RWC; n = 7 for leaf water potential)
Reactive oxygen species and antioxidant enzyme activity
The plants exposed to R1 and R3 treatments showed high content of superoxide radical after 7 days of the rehydration as compared to R2 treatment (Fig. 3).

Histochemical detection of superoxide radical with the use of nitroblue tetrazolium (NBT) performed after 7 days of rehydration for fully-recovered ‘Hortenso’ cultivar and not fully-recovered ‘Kazo’ cultivar (C -control plants, R1/R2/R3-rehydration). Black bars = 1 cm
The leaves of fully-recovered cv. ‘Hortenso’ showed a decrease in H2O2 content down to the control level during R1 rehydration (Fig. 4a and d). Low content of hydrogen peroxide in this cultivar was accompanied by high activity of catalase (Fig. 4b) and peroxidase (Fig. 4c). In the not fully-recovered cv. ‘Kazo’, increased activity of catalase and peroxidase and low H2O2 content were observed only during R2 treatment.

The content of hydrogen peroxide (a, [μmol H2O2 g−1 DW]), activity of catalase (CAT, [μmol H2O2 min−1 mg−1 proteins]) (b) and total activity of peroxidase (POX, [units−1 min−1 mg−1 proteins]) (c) for fully-recovered ‘Hortenso’ cultivar and not fully-recovered ‘Kazo’ cultivar. The analyses were performed 1, 3 and 7 days after rehydration. Control-(○), drought-(●), rehydration: R1-(□), R2-( ), R3-(■). Means ± SE (n = 10 for H2O2; n = 7 for CAT and POX). d Histochemical detection of H2O2 with the use of 3,3’-diaminobenzidine (DAB), performed after 7 days of rehydration for fully-recovered ‘Hortenso’ cultivar and not fully-recovered ‘Kazo’ cultivar (C-control plants, D-drought, R1/R2/R3-rehydration). Black bars = 1 cm
), R3-(■). Means ± SE (n = 10 for H2O2; n = 7 for CAT and POX). d Histochemical detection of H2O2 with the use of 3,3’-diaminobenzidine (DAB), performed after 7 days of rehydration for fully-recovered ‘Hortenso’ cultivar and not fully-recovered ‘Kazo’ cultivar (C-control plants, D-drought, R1/R2/R3-rehydration). Black bars = 1 cm
Intensity of blue fluorescence (F440) excitation
In the fully-recovered cv. ‘Hortenso‘, a significant increase in the intensity of blue fluorescence excitation was noticed on the last day of drought and after 7 days of R1 treatment (Fig. 5). In the not fully-recovered ‘Kazo’ plants, large and statistically significant increase in the intensity of blue fluorescence excitation in relation to the control and drought variant was observed only for R2 treatment.

Intensity of blue fluorescence (F440) excitation for fully-recovered ‘Hortenso’ cultivar and not fully-recovered ‘Kazo’ cultivar. The analyses were performed after 3 weeks of drought and 7 days of rehydration. Control -(C, ○), drought-(D, ●), rehydration (R): R1-(□), R2-( ), R3-(■). Duncan’s test at 0.05 probability level was performed in order to determine the significance of differences between treatment means. Means indicated with the same letters within cultivar were not significantly different. Means ± SE (n = 10)
), R3-(■). Duncan’s test at 0.05 probability level was performed in order to determine the significance of differences between treatment means. Means indicated with the same letters within cultivar were not significantly different. Means ± SE (n = 10)
Activity of the photosynthetic apparatus in the vegetative growth phase
Leaf photosynthetic activity was fully restored after 7 days of rehydration in ‘Hortenso’ cultivar exposed to R1 treatment and in ‘Kazo’ plants subjected to R2 scheme (Fig. 6). After 7 days of R1 rehydration, the net photosynthesis rate (Pn) (Fig. 6a), transpiration (E) (Fig. 6b) and stomatal conductance (gs) (Fig. 6c) in the not fully-recovered cultivar were significantly lower than those recorded in the drought-exposed plants. After 7 days of R3 rehydration, gas exchange intensity was at the same level as in the drought-exposed plants.

Net photosynthetic activity (Pn, [μmol (CO2) m−2 s−1]) (a), transpiration activity (E, [mmol (H2O) m−2 s−1]) (b) and stomatal conductance (gs, [mmol (CO2) m−2 s−1]) (c) for fully-recovered ‘Hortenso’ cultivar and not fully-recovered ‘Kazo’ cultivar. The analyses were performed after 3 weeks of drought and 7 days of rehydration. Control -(C,○), drought-(D, ●), rehydration (R): R1-(□), R2-( ), R3-(■). Duncan’s test at 0.05 probability level was performed in order to determine the significance of differences between treatment means. Means indicated with the same letters within cultivar were not significantly different. Means ± SE (n = 10)
), R3-(■). Duncan’s test at 0.05 probability level was performed in order to determine the significance of differences between treatment means. Means indicated with the same letters within cultivar were not significantly different. Means ± SE (n = 10)
After 7 days of R2 rehydration, fluorescence parameters in the leaves of not fully-recovered cultivar were the same as in the control plants (Table 1). Further decline in the activity of the photosynthetic apparatus, as compared with drought-exposed plants, was observed during R1 rehydration. It was manifested by significantly lower (PI, ETo/CSm, RC/CSm, ψRo, δRo, φRo) or higher (DIo/CSm) chlorophyll fluorescence parameters. In the fully-recovered ‘Hortenso’ cultivar, chlorophyll fluorescence parameters after 7 days of R1 treatment were equal to the control (Table 1).
Statistically significant correlations were observed between chlorophyll fluorescence parameters and hydrogen peroxide level (Fig. 7). High amounts of both the energy used for electron transport (ETo/CSm) (Fig. 7a) and the excitation energy trapped in PSII reaction centers (TRo/CSm) (Fig. 7b) correlated with increased content of H2O2. Additionally, higher content of H2O2 corresponded to a reduced probability that a trapped exciton moves an electron into the electron transport chain beyond QA − (ψRo) (Fig. 7c), the efficiency with which an electron can move from the reduced intersystem of electron acceptors to the PSI end electron acceptors (δRo) (Fig. 7d), and the quantum yield of electron transport from QA − to the PSI end electron acceptors (φRo) (Fig. 7e).

Correlations between hydrogen peroxide level and amount of energy used for the electron transport (ETo/CSm) (a), amount of excitation energy trapped in PSII reaction centers (TRo/CSm) (b), probability that a trapped exciton moves an electron into the electron transport chain beyond QA − (ψRo) (c), the efficiency with which an electron can move from the reduced intersystem of electron acceptors to the PSI end electron acceptors (δRo) (d) and the quantum yield of electron transport from QA − to the PSI end electron acceptors (φRo) (e). Control (C) for R1, R2 and R3 treatments - white circles; drought (D, data collected on the last day of drought treatment) - black circles; rehydration (data recorded on the seventh day of rehydration): R1 - white squares, R2 - gray squares, R3 - black squares. The lines represent linear adjustment at a probability level p < 0.05
An analysis of red (F690) and far-red (F740) fluorescence emission spectra revealed that in the not fully-recovered plants exposed to 7 days of R2 rehydration low intensity of red fluorescence emission (equal to control) (Fig. 8a) was accompanied by high intensity of far-red fluorescence emission (Fig. 8b and c). The not fully-recovered plants exposed to R1 treatment responded with a significant increase in the intensity of red fluorescence emission, while F740 fluorescence was not significantly different from either the control or drought-exposed plants. Intensity of F740 emission during R3 rehydration was similar to that observed for control and drought-exposed plants. In contrast, the intensity of F690 emission was significantly higher than that of the control but did not differ significantly from that observed in drought-exposed plants.

a Exemplary emission spectra of red fluorescence and far-red fluorescence performed for the control plants (C) after 3 weeks of drought (D) and 7 days of rehydration (R) for fully-recovered ‘Hortenso’ cultivar and not fully-recovered ‘Kazo’ cultivar. C, D, R1 - solid line, R2 - dashed line, R3 - dotted line. b–c Intensity of red fluorescence (F690) and far-red fluorescence (F740) emission for fully-recovered ‘Hortenso’ cultivar and not fully-recovered ‘Kazo’ cultivar. The analyses were performed after 3 weeks of drought and 7 days of rehydration. Control -(C, ○), drought-(D, ●), rehydration (R): R1-(□), R2-( ), R3-(■). Duncan’s test at 0.05 probability level was performed in order to determine the significance of differences between treatment means. Means indicated with the same letters within cultivar were not significantly different. Means ± SE (n = 10). d Fluorescence imaging of leaf fragments for fully-recovered ‘Hortenso’ cultivar and not fully-recovered ‘Kazo’ cultivar. C-control, D-drought, R1/R2/R3-rehydration. White bars = 1 cm
), R3-(■). Duncan’s test at 0.05 probability level was performed in order to determine the significance of differences between treatment means. Means indicated with the same letters within cultivar were not significantly different. Means ± SE (n = 10). d Fluorescence imaging of leaf fragments for fully-recovered ‘Hortenso’ cultivar and not fully-recovered ‘Kazo’ cultivar. C-control, D-drought, R1/R2/R3-rehydration. White bars = 1 cm
Fluorescence imaging of leaf segments, in which gas exchange and chlorophyll fluorescence were analyzed, indicated increased intensity of red fluorescence emission in ‘Hortenso’ and ‘Kazo’ cultivars following 3 weeks of drought (Fig. 8d). Higher intensity of red fluorescence emission in the drought-exposed plants was observed for R1 rehydration. In R2 variant, red fluorescence emission was similar to the control, while after 7 days of R3 treatment it was similar to that observed for drought-exposed plants.
Three weeks of soil drought resulted in a decrease of chlorophyll and carotenoid content in both cultivars (Table 2). After 7 days of R1 rehydration in the fully-recovered cv. ‘Hortenso’ and after 7 days of R1, R2 or R3 rehydration in the not fully-recovered cv. ‘Kazo’ the content of chlorophyll and carotenoids did not differ significantly from the controls.
Activity of the photosynthetic apparatus in flag leaf and grain yield analysis
The analyses carried out after 42 days of rehydration showed no significant differences in flag leaf water content, pigment content, hydrogen peroxide level, and the activity of antioxidant enzymes (Table 3). The leaves of not fully-recovered cv. ‘Kazo’, rehydrated using R1 and R3 schemes, still showed low photosynthetic activity (Fig. 9a), accompanied by lack of differences concerning transpiration (Fig. 9b) and stomatal conductance (Fig. 9c) when compared to the control plants. However, Pn values in the not fully-recovered cultivar were significantly higher for R3 than for R1 (Fig. 9a). It was also observed that R1 and R3 rehydration triggered permanent reduction in the activity of flag leaf photosynthetic apparatus. In comparison with the control, R1 treatment resulted in a significant drop in the maximum photochemical efficiency of PSII (Fv/Fm) (Fig. 10a), a decrease in effective excitation energy capture by PSII centers and trapping of light quanta by PSII antennae (Fv’/Fm’) (Fig. 10b), a reduction in the PSII quantum efficiency (ΦPSII) (Fig. 10c), less effective transfer of excitation energy towards further photochemical reactions (qP) (Fig. 10d), an increase in energy dissipation as heat (qN) (Fig. 10e), and lower electron transport rate (ETR) (Fig. 10f).

Net photosynthetic activity (Pn, [μmol (CO2) m−2 s−1]) (a), transpiration activity (E, [mmol (H2O) m−2 s−1]) (b) and stomatal conductance (gs, [mmol (CO2) m−2 s−1]) (c) for fully-recovered ‘Hortenso’ cultivar (white bars) and not fully-recovered ‘Kazo’ cultivar (black bars). The analyses were performed for flag leaves on the 42nd day of rehydration. C-control, D-drought, R1/R2/R3-rehydration. Duncan’s test at 0.05 probability level was performed in order to determine the significance of differences between treatment means. Means indicated with the same letters within the investigated parameters were not significantly different. Means ± SE (n = 10)

The maximum potential PSII efficiency (Fv/Fm) (a), the efficiency of excitation transfer to open PSII centers (Fv ’/Fm ’) (b), PSII quantum efficiency (ФPSII) (c), photochemical quenching coefficient (qP) (d), non-photochemical quenching coefficient (qN) (e) and the electron transport rate (ETR, [μmol electrons m−2 s−1]) (f) for fully-recovered ‘Hortenso’ cultivar (white bars) and not fully-recovered ‘Kazo’ cultivar (black bars). The analyses were performed for flag leaves on the 42nd day of rehydration. C-control, D-drought, R1/R2/R3-rehydration. Duncan’s test at 0.05 probability level was performed in order to determine the significance of differences between treatment means. Means indicated with the same letters within the investigated parameters were not significantly different. Means ± SE (n = 10). g–h Changes in the emission intensity of red fluorescence (F690) and far-red fluorescence (F740) depending on the number of point excitation events (from 1 to 10, 1-min interval) of the same fragment of a flag leaf with light of 450 nm in not fully-recovered ‘Kazo’ cultivar on the 42nd day of rehydration. There were ten excitation events for 10 leaves in R1, R2 and R3 treatments. Each excitation event was followed by reading the intensity of red and far-red fluorescence emission. R1-white squares, R2-gray squares, R3-black squares. Means ± SE (n = 10)
Flag leaves of the not fully-recovered cv. ‘Kazo’ exposed to R2 rehydration and subjected to an increasing number of point excitations (from 1 to 10) with the light of 450 nm, emitted less intense red fluorescence (Fig. 10g) but more intense far-red fluorescence (Fig. 10h). During R1 treatment of the same cultivar, higher number of excitation events corresponded to more intense F690 emission, while the intensity of F740 emission remained unchanged. In contrast, R3 rehydration resulted in a slight increase in the intensity of both types of fluorescence with growing number of excitation events.
No significant differences in grain yield were found for individual rehydration treatments in cv. ‘Hortenso’ (Fig. 11). In not fully-recovered ‘Kazo’ plants, grain yield was the same as in control following R2 treatment but it was significantly lower than in control for the other two treatments.

Analysis of grain yield for fully-recovered ‘Hortenso’ cultivar (white bars) and not fully-recovered ‘Kazo’ cultivar (black bars). C-control, D-drought, R1/R2/R3-rehydration. Duncan’s test at 0.05 probability level was performed in order to determine the significance of differences between treatment means. Means indicated with the same letters within cultivar were not significantly different. Means ± SE (n = 18)
Discussion
So far, numerous papers have been published on different mechanisms of plant acclimation to soil drought and on their role in the selection of cultivars or genotypes resistant to soil water deficit (Flexas et al. 1999, 2006; Escalona et al. 2000; Flexas and Medrano 2002; Hura et al. 2007a, 2009a, b, c, 2011, 2012; Grzesiak et al. 2010; Rollins et al. 2013; Nolf et al. 2014; Borrell et al. 2014). However, very few studies investigated the problem of adverse functional changes brought about by the rehydration stage (Oliver et al. 1998; Hura et al. 2009a). In mosses and lichens, characterized by high tolerance to desiccation, rehydration stage is accompanied by hydrogen peroxide burst that may be caused by increased activity of extracellular peroxidases (Mayaba et al. 2002) and light (Proctor and Smirnoff 2000). It is well-known that an increased level of hydrogen peroxide in the plant cells is, on the one hand, a biochemical indicator of stress that can damage the plant cell structural elements and disturb its functioning (Mittler 2002), and on the other hand, it is an important signal transduction factor controlling the expression of genes involved in adaptive mechanisms activated by environmental stressors (Neill et al. 2002; Apel and Hirt 2004).
In our study, the rapid rehydration (R1) that, similarly to drought, triggered additional accumulation of H2O2 in the leaves of not fully-recovered triticale cultivar may initiate may initiate permanent and adverse changes in plants metabolism (Fig. 4a and d). However, it should be pointed out that the increase in H2O2 content could also correspond to the cumulative and lag response to soil drought.
The photosynthetic apparatus is an important source of ROS overproduction under environmental stresses (Asada 2006). Rapid increase in H2O2 content observed for R1 treatment could be due to low efficiency of excess excitation energy dissipation through the photosynthetic apparatus. In the not fully-recovered cultivar, R1 treatment resulted in increased amount of energy used for electron transport (ETo/CSm) and higher amount of excitation energy trapped in PSII reaction centers (TRo/CSm). Other effects of R1 treatment included low probability that a trapped exciton moves an electron into the electron transport chain beyond QA − (ψRo), low efficiency with which an electron can move from the reduced intersystem electron acceptors to the PSI end electron acceptors (δRo), and reduced quantum yield of electron transport from QA − to the PSI end electron acceptors (φRo) (Table 1). Reduction in chlorophyll fluorescence parameters (ψRo, δRo, φRo) during R1 rehydration may indicate electron transfer from plastoquinone QA to oxygen molecules. The above mentioned statements were supported by statistically significant correlations between chlorophyll fluorescence parameters (TRo/CSm, ETo/CSm, ψRo, δRo, φRo) and the content of hydrogen peroxide (Fig. 7). Takahashi et al. (1988) showed that in PSII, in the places of plastoquinone QA and QB activity, electron leak may occur, accompanied by a reduction of molecular oxygen to superoxide radical, further dismutated to H2O2. Another possible source of ROS generation during R1 could be PSI, where overloading of the electron transport chain (ETC) could cause a redirection of some electrons from ferredoxin to O2 and reducing it via Mehler reaction to superoxide radical, which was then spontaneously dismutated to H2O2 by Cu/Zn-SOD (Polle 1996; Edreva 2005). However, Wiese et al. (1998) showed that oxygen reduction in the Mehler reaction was insufficient to protect photosystem I and II against photoinactivation. An important mechanism, preventing overloading of PSII with excitation energy during R2 treatment, was its effective transfer to PSI and then dissipation in the form of far-red fluorescence (Fig. 8a–c) (Baker 1991; Huner et al. 1998; Agati et al. 2000).
High level of hydrogen peroxide observed during R1 and R3 was accompanied by low activity of catalase (Fig. 4b) and peroxidases (Fig. 4c) as compared to gradual rehydration of leaf cells during R2 treatment. The fact that hydrogen peroxide content under stress depends on the antioxidant status of a plant cell, associated e.g., with the activity of superoxide radical dismutase and H2O2 scavenging enzymes, has been confirmed by numerous authors (Fath et al. 2001; Neill et al. 2002; Srivalli et al. 2003). An increase in blue fluorescence excitation intensity (Fig. 5) may indicate another possible mode of H2O2 neutralization during R2 treatment. This type of fluorescence usually indicates the presence of phenolic compounds, mainly ferulic acid, associated with the cell wall carbohydrates (Lichtenthaler and Schweiger 1998). Schopfer (1996) proved that the incorporation of phenols in the cell wall is mediated by peroxidases and hydrogen peroxide as an oxidizing agent. The observed increase in the intensity of blue fluorescence excitation in R2 treatment may indicate one of the mechanisms for reducing the content of hydrogen peroxide in the process of phenolics incorporation in the cell wall. A similar increase in the intensity of emission or excitation of blue fluorescence from the plant leaves was observed under other environmental stresses that may be accompanied by a secondary oxidative stress (Lang et al. 1996; Schweiger et al. 1996).
High level of hydrogen peroxide was maintained also during R3 treatment at PPFD of 25 μmol m−2 s−1 (Fig. 4a and d). This may suggest an intensification of light-independent metabolic processes (e.g., mitochondrial electron transport chain, fatty acid β-oxidation, Mn-SOD activity in peroxisomes, NADH oxidase activity) that may also result in hydrogen peroxide formation (Møller 2001; del Río et al. 2006). It should be mentioned, however, that in PSII, even under low-light conditions, the reactive oxygen species might be generated as by-products of photochemical reactions (Gill and Tuteja 2010).
The analyses performed in flag leaves revealed permanent changes in the photosynthetic activity in the not fully-recovered cv. ‘Kazo’ (Figs. 9 and 10). Changes in the parameters of chlorophyll fluorescence and photosynthetic activity, observed 7 days after finishing the drought period during R1 rehydration, were probably due to damage to the photosynthetic apparatus caused by ROS, including hydrogen peroxide (Miller et al. 2010). Moreover, hydrogen peroxide is involved in abscisic acid-induced stomatal closure (Zhang et al. 2001; Bright et al. 2006; Miao et al. 2006), which may be another reason for reduced photosynthetic activity after 7 days of R1 or R3 treatment.
Low photosynthetic activity of flag leaves at the generative development stage during R1 and R3 treatment is an example of a negative permanent effect of rapid rehydration. High level of hydrogen peroxide that acts as a secondary cellular messenger could induce the expression of genes controlling photosynthetic activity during R1 and R3 treatment (Foyer et al. 1997; Mittler 2002; Mittler et al. 2004). Low photosynthetic activity observed during R1 and R3 rehydration was not caused by disturbances in the stomatal conductance (Fig. 9c), changes in chlorophyll content or leaf water content (Table 3). It was also not due to differences in hydrogen peroxide content or activity of antioxidant enzymes (Table 3). Measurements of the flag leaves chlorophyll fluorescence during R1 and R3 treatment (Fig. 10) revealed that the reduced rate of photosynthesis could be caused by disturbances in the photosynthetic apparatus activity (Bolhar-Nordenkampf and Öquist 1993; Maxwell and Johnson 2000; Baker and Rosenqvist 2004). Permanent and unfavorable changes in the photosynthetic activity of the flag leaves in not fully-recovered cv. ‘Kazo’ triggered by R1 and R3 treatment involved also a drop in the efficiency of excitation energy transfer between PSII and PS I (Fig. 10g and h) (Huner et al. 1998; Tikkanen and Aro 2012).
Hydrogen peroxide is the most stable and long-distance ROS signaling molecule that regulates stress responses and developmental processes in plants (Vranová et al. 2002). Vandenabeele et al. (2002) showed that hydrogen peroxide could provoke transcriptional responses that mimicked both biotic and abiotic stresses. Therefore, sustained increases in H2O2 content under rapid rehydration could also induce genes expression involved in physiological adjustments, e.g., the photosynthetic apparatus activity. It has been known since the 1970s that an increase in hydrogen peroxide content affects the activity of the Calvin cycle enzymes (Kaiser 1979). Tanou et al. (2009) reported that plants exposed to H2O2 activated specific proteome reprogramming to prevent accumulation of proteins, e.g., the key enzymes of the Calvin cycle (Rubisco activase, Rubisco large subunit, fructose 1,6-bisphosphate aldolase, phosphoglycerate kinase, glyceraldehyde-3-phosphate dehydrogenase, phosphoribulokinase, transketolase and carbonic anhydrase). Hence, it may be speculated that the impact of rapid rehydration after drought at the tillering stage on low photosynthesis during the reproductive phase is connected with H2O2-induced gene expression related to persistent limitation in CO2 assimilation.
Our results indicate that rapid rehydration can initiate adverse changes in the activity of the photosynthetic apparatus. Therefore, to show a comprehensive process of plant acclimation to drought, this rehydration stage should be taken into account in the experiments aimed at a selection of cultivars resistant to soil drought and during the search for the genome regions related to the resistance to soil water deficit. Further studies are currently being conducted to elucidate the molecular mechanisms of the photosynthetic apparatus acclimation in the rehydration conditions.
Conclusions
This study has proved that rapid rehydration, occurring after soil drought at the tillering stage, caused negative changes manifested by lowered photosynthetic activity of the flag leaves and, consequently, a significant drop in triticale grain weight. We suggest that, despite rehydration of triticale leaves, further accumulation of hydrogen peroxide during the rehydration at the tillering stage was a biochemical factor determining full or partial recovery of triticale. A destructive effect of the rapid rehydration was manifested in an intensification of the physiological processes associated with reactive oxygen species (ROS) production. An important cause of hydrogen peroxide overproduction could be the electron leakage due to overloading of the electron transport chain (ETC) in PSI and PSII.
References
Aebi H (1984) Catalase in vitro. In: Willam BJ (ed) Methods in enzymology. Academic, New York, pp 121–126
Agati G, Cerovic ZG, Moya I (2000) The effect of decreasing temperature up to chilling values on the in vivo F685/F735 chlorophyll fluorescence ratio in Phaseolus vulgaris and Pisum sativum: the role of the photosystem I contribution to the 735 nm fluorescence band. Photochem Photobiol 72:75–84
Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 55:373–39
Asada K (2006) Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol 141:391–396
Baker NR (1991) A possible role for photosystem II in environmental perturbations of photosynthesis. Physiol Plant 81:563–570
Baker NR, Rosenqvist E (2004) Applications of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. J Exp Bot 55:1607–1621
Barnabás B, Jäger K, Fehér A (2008) The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ 31:11–38
Barrs HD, Weatherley PE (1962) A re-examination of the relative turgidity techniques for estimating water deficits in leaves. Aust J Biol Sci 15:413–428
Bartosz G (1997) Oxidative stress in plants. Acta Physiol Plant 19:47–64
Beckett M, Loreto F, Velikova V, Brunetti C, Di Ferdinando M, Tattini M, Calfapietra C, Farrant JM (2012) Photosynthetic limitations and volatile and non-volatile isoprenoids in the poikilochlorophyllous resurrection plant Xerophyta humilis during dehydration and rehydration. Plant Cell Environ 35:2061–2074
Bergmeyer HU (1965) Methods of enzymatic analysis. Verlag Chemie G.m.b.H., Weinheim/Bergstr. Academic Press, New York and London
Blokhina O, Virolainen E, Fagerstedt KV (2002) Antioxidants, oxidative damage and oxygen deprivation stress: a review. Ann Bot 91:179–194
Bolhar-Nordenkampf HR, Öquist G (1993) Chlorophyll fluorescence as a tool in photosynthesis research. In: Hall DO, Scurlock JMO, Bolhàr-Nordenkampf HR, Leegood RC, Long SP (eds) Photosynthesis and production in a changing environment: a field and laboratory manual. Chapman & Hall, London, pp 193–206
Borrell AK, Mullet JE, George-Jaeggli B, van Oosterom EJ, Hammer GL, Klein PE, Jordan DR (2014) Drought adaptation of stay-green sorghum is associated with canopy development, leaf anatomy, root growth, and water uptake. J Exp Bot 65:6251–6263
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Bright J, Desikan R, Hancock JT, Weir IS, Neill SJ (2006) ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J 45:113–122
Cannell RQ, Belford RK, Gales K, Thomson RJ, Webster CP (1984) Effects of waterlogging and drought on winter wheat and winter barley grown on a clay and a sandy loam soil. Plant Soil 80:53–66
del Río LA, Sandalio LM, Corpas FJ, Palma JM, Barroso JB (2006) Reactive oxygen species and reactive nitrogen species in peroxisomes. production, scavenging, and role in cell signalling. Plant Physiol 141:330–335
Doke N, Ohashi Y (1988) Involvement of and O2 •– generating system in the induction of necrotic lesions on tobacco leaves infected with tobacco mosaic virus. Physiol Mol Plant Pathol 32:165–175
Edreva A (2005) Generation and scavenging of reactive oxygen species in chloroplasts: a submolecular approach. Agric Ecosyst Environ 106:119–133
Escalona JM, Flexas J, Medrano H (2000) Stomatal and non-stomatal limitations of photosynthesis under water stress in field-grown grapevines. Aust J Plant Physiol 27:87–87
Fath A, Bethke PC, Jones RL (2001) Enzymes that scavenge reactive oxygen species are down-regulated prior to gibberellic acid-induced programmed cell death in barley aleurone. Plant Physiol 126:156–166
Flexas J, Medrano H (2002) Energy dissipation in C3 plants under drought. Funct Plant Biol 29:1209–1215
Flexas J, Escalona JM, Medrano H (1999) Water stress induces different levels of photosynthesis and electron transport rate regulation in grapevines. Plant Cell Environ 22:39–48
Flexas J, Ribas-Carbó M, Bota J, Galmés J, Henkle M, Martínez-Cañellas S, Medrano H (2006) Decreased Rubisco activity during water stress is not induced by decreased relative water content but related to conditions of low stomatal conductance and chloroplast CO2 concentration. New Phytol 172:73–82
Foyer CH, Lopez-Delgado H, Dat JF, Scott IM (1997) Hydrogen peroxide- and glutathione-associated mechanisms of acclimatory stress tolerance and signalling. Physiol Plant 100:241–254
Furlan AL, Bianucci E, Tordable MC, Castro S, Dietz KJ (2014) Antioxidant enzyme activities and gene expression patterns in peanut nodules during a drought and rehydration cycle. Funct Plant Biol 41:704–713
Genty B, Briantais JM, Adams WW (1989) The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta 990:87–92
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Grzesiak MT, Filek W, Hura T, Kocurek M, Pilarski J (2010) Leaf optical properties during and after drought stress in triticale and maize genotypes differing in drought tolerance. Acta Physiol Plant 32:433–442
Hoagland DR (1948) Lectures on the inorganic nutrition of plants. Chronica Botanica Co, Waltham
Huner NPA, Öquist G, Sarhan F (1998) Energy balance and acclimation to light and cold. Trends Plant Sci 3:224–230
Hura T, Grzesiak S, Hura K, Grzesiak MT, Rzepka A (2006) Differences in the physiological state between triticale and maize plants during drought stress and followed rehydration expressed by the leaf gas exchange and spectrofluorimetric methods. Acta Physiol Plant 5:433–443
Hura T, Grzesiak S, Hura K, Thiemt E, Tokarz K, Wedzony M (2007a) Physiological and biochemical tools useful in drought-tolerance detection in genotypes of winter triticale: accumulation of ferulic acid correlates with drought tolerance. Ann Bot 100:767–775
Hura T, Hura K, Grzesiak MT, Rzepka A (2007b) Effect of long-term drought stress on leaf gas exchange and fluorescence parameters in C3 and C4 plants. Acta Physiol Plant 29:103–113
Hura T, Hura K, Grzesiak S (2009a) Possible contribution of cell wall-bound ferulic acid in drought resistance and recovery in triticale seedlings. J Plant Physiol 166:1720–1733
Hura T, Hura K, Grzesiak S (2009b) Physiological and biochemical parameters for identifications of QTLs controlling the winter triticale drought tolerance at the seedling stage. Plant Physiol Biochem 47:210–214
Hura T, Hura K, Grzesiak S (2009c) Leaf dehydration induces different content of phenolics and ferulic acid in drought-resistant and -sensitive genotypes of spring triticale. Z Naturforsch 64c:85–95
Hura T, Hura K, Grzesiak M (2011) Soil drought applied during the vegetative growth of triticale modifies physiological and biochemical adaptation to drought during the generative development. J Agron Crop Sci 197:113–123
Hura T, Hura K, Dziurka K, Ostrowska A, Bączek-Kwinta R, Grzesiak MT (2012) An increase in the content of cell wall-bound phenolics correlates with the productivity of triticale under soil drought. J Plant Physiol 169:1728–1736
Hura K, Hura T, Dziurka K, Dziurka M (2014) Biochemical defense mechanisms induced in winter oilseed rape seedlings with different susceptibility to infection with Leptosphaeria maculans. Physiol Mol Plant Pathol 87:42–50
Ishikawa T, Takeda T, Shigeoka S, Hirayama O, Mitsunaga T (1993) Hydrogen peroxide generation in organelles of Euglena gracilis. Phytochemistry 33:1297–1299
Kaiser WM (1979) Reversible inhibition of the Calvin cycle and activation of oxidative pentose phosphate cycle in isolated intact chloroplasts by hydrogen peroxide. Planta 145:377–382
Lang M, Lichtenthaler HK, Sowinska M, Heisel F, Miehé JA (1996) Fluorescence imaging of water and temperature stress in plant leaves. J Plant Physiol 148:613–621
Lichtenthaler HK, Schweiger J (1998) Cell wall bound ferulic acid, the major substance of the blue-green fluorescence emission of plants. J Plant Physiol 152:272–282
Lichtenthaler HK, Wellburn RR (1983) Determination of total carotenoids and chlorophylls a and b of extracts in different solvents. Biochem Soc Trans 603:591–592
Lovisolo C, Perrone I, Hartung W, Schubert A (2008) An abscisic acid-related reduced transpiration promotes gradual embolism repair when grapevines are rehydrated after drought. New Phytol 180:642–651
Maksymiec W (2007) Signaling responses in plants to heavy metal stress. Acta Physiol Plant 29:177–187
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence - a practical guide. J Exp Bot 51:659–668
Mayaba N, Minibayeva F, Beckett RP (2002) An oxidative burst of hydrogen peroxide during rehydration following desiccation in the moss Atrichum androgynum. New Phytol 155:275–283
McDowell N, Pockman WT, Allen CD, Breshears DD, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams DG, Yepez EA (2008) Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytol 178:719–739
Miao Y, Lv D, Wang P, Wang XC, Chen J, Miao C, Song CP (2006) An Arabidopsis glutathione peroxidase functions as both a redox transducer and a scavenger in abscisic acid and drought stress responses. Plant Cell 18:2749–2766
Miller G, Suzuki N, Ciftci-Yilmaz S, Mittler R (2010) Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ
Minibayeva F, Beckett RP (2001) High rates of extracellular superoxide production in bryophytes and lichens, and an oxidative burst in response to rehydration following desiccation. New Phytol 152:333–343
Mittler R (2002) Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 7:405–410
Mittler R, Vanderauwera S, Gollery M, van Breusegem F (2004) Reactive oxygen gene network of plants. Trends Plant Sci 9:490–498
Møller IM (2001) Plant mitochondria and oxidative stress: electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annu Rev Plant Physiol Plant Mol Biol 52:561–591
Nedbal L, Soukupová J, Kaftan D, Whitmarsh J, Trtílek M (2000) Kinetic imaging of chlorophyll fluorescence using modulated light. Photosynth Res 66:3–12
Neill S, Desikan R, Hancock J (2002) Hydrogen peroxide signalling. Curr Opin Plant Biol 5:388–395
Nolf M, Pagitz K, Mayr S (2014) Physiological acclimation to drought stress in Solidago canadensis. Physiol Plant 150:529–539
O’Grady AP, Mitchell PJM, Pinkard EA, Tissue DT (2013) Thirsty roots and hungry leaves: unravelling the roles of carbon and water dynamics in tree mortality. New Phytol 200:294–297
Oliver MJ, O’Mahony P, Wood AJ (1998) ‘To dryness and beyond’ – preparation for the dried state and rehydration in vegetative desiccation-tolerant plants. Plant Growth Regul 24:193–201
Orozco-Cárdenas ML, Narváez-Vásquez J, Ryan CA (2001) Hydrogen peroxide acts as a second messenger for the induction of defense genes in tomato plants in response to wounding, systemin, and methyl jasmonate. Plant Cell 13:179–191
Polle A (1996) Mehler reaction: friend or foe in photosynthesis? Bot Acta 109:84–89
Proctor MCF, Smirnoff N (2000) Rapid recovery of photosystems on rewetting desiccation-tolerant mosses: chlorophyll fluorescence and inhibitor experiments. J Exp Bot 51:1695–1704
Rollins JA, Habte E, Templer SE, Colby T, Schmidt J, von Korff M (2013) Leaf proteome alterations in the context of physiological and morphological responses to drought and heat stress in barley (Hordeum vulgare L.). J Exp Bot 64:3201–3212
Saini HS, Westgate ME (2000) Reproductive development in grain crops during drought. In: Spartes DL (ed) Advances in agronomy. Academic, San Diego, pp 59–96
Sala A, Piper F, Hoch G (2010) Physiological mechanisms of drought-induced tree mortality are far from being resolved. New Phytol 186:274–281
Schopfer P (1996) Hydrogen peroxide-mediated cell-wall stiffening in vitro in maize coleoptiles. Planta 199:43–49
Schweiger J, Lang M, Lichtenthaler HK (1996) Differences in fluorescence excitation spectra of leaves between stressed and non-stressed plants. J Plant Physiol 148:536–547
Sevanto S, McDowell NG, Dickman LT, Pangle R, Pockman WT (2014) How do trees die? A test of the hydraulic failure and carbon starvation hypotheses. Plant Cell Environ 37:153–161
Shah NH, Paulsen GM (2003) Interaction of drought and high temperature on photosynthesis and grain-filling of wheat. Plant Soil 257:219–226
Souza RP, Machado EC, Silva JAB, Lagôa AMMA, Silveira JAG (2004) Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environ Exp Bot 51:45–56
Srivalli B, Sharma G, Khanna-Chopra R (2003) Antioxidative defense system in an upland rice cultivar subjected to increasing intensity of water stress followed by recovery. Physiol Plant 119:503–512
Strasser RJ, Tsimilli-Michael M (2001) Stress in plants, from daily rhythm to global changes, detected and quantified by the JIP–Test. Chim Nouv 75:3321–3326
Strasser RJ, Tsimilli-Michael M, Qiang S, Goltsev V (2010) Simultaneous in vivo recording of prompt and delayed fluorescence and 820 nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim Biophys Acta 1797:1313–1326
Takahashi M, Shiraishi T, Asada K (1988) Superoxide production in aprotic interior of chloroplast thylakoids. Arch Biochem Biophys 267:714–722
Tanou G, Job C, Rajjou L, Arc E, Belghazi M, Diamantidis G, Molassiotis A, Job D (2009) Proteomics reveals the overlapping roles of hydrogen peroxide and nitric oxide in the acclimation of citrus plants to salinity. Plant J 60:795–804
Thordal-Christensen H, Zhang Z, Wei Y, Collinge DB (1997) Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J 11:1187–1194
Tikkanen M, Aro EM (2012) Thylakoid protein phosphorylation in dynamic regulation of photosystem II in higher plants. Biochim Biophys Acta 1817:232–238
Tsimilli-Michael M, Strasser RJ (2008) In vivo assessment of plant’s vitality: applications in detecting and evaluating the impact of mycorrhization on host plants. In: Varma A (ed) Mycorrhiza, state of the art, genetics, and molecular biology, eco-function, biotechnology, eco-physiology, structure, and systematics, 3rd edn. Springer, Dordrecht, pp 679–703
Turner NC (1981) Techniques and experimental approaches for the measurement of plant water status. Plant Soil 58:339–366
van Kooten O, Snel FH (1990) The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth Res 25:147–150
Vandenabeele S, Van Der Kelen K, Dat J, Gadjev I, Boonefaes T, Morsa S, Rottiers P, Slooten L, Van Montagu M, Zabeau M, Inzé D, Van Breusegem F (2002) A comprehensive analysis of hydrogen peroxide-induced gene expression in tobacco. Proc Natl Acad Sci U S A 100:16113–16118
Vranová E, Inzé D, Van Breusegem F (2002) Signal transduction during oxidative stress. J Exp Bot 53:1227–1236
Weissman L, Garty J, Hochman A (2005) Rehydration of the lichen Ramalina lacera results in production of reactive oxygen species and nitric oxide and a decrease in antioxidants. Appl Environ Microbiol 71:2121–2129
Wiese C, Shi LB, Heber U (1998) Oxygen reduction in the Mehler reaction is insufficient to protect photosystems I and II of leaves against photoinactivation. Physiol Plant 102:437–446
Yang JC, Zhang JH (2006) Grain filling of cereals under soil drying. New Phytol 169:223–236
Zhang X, Zhang L, Dong F, Gao J, Galbraith DW, Song CP (2001) Hydrogen peroxide is involved in abscisic acid-induced stomatal closure in Vicia faba. Plant Physiol 126:1438–1448
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Rafael S. Oliveira,.
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 14 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Hura, T., Hura, K., Ostrowska, A. et al. Rapid plant rehydration initiates permanent and adverse changes in the photosynthetic apparatus of triticale. Plant Soil 397, 127–145 (2015). https://doi.org/10.1007/s11104-015-2607-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-015-2607-1