
Abstract
Context
Montados are dynamic agroforestry systems of southern Portugal, with high economic and ecological values. Changes in land use and cover have important implications for landscape-level biodiversity and its conservation.
Objectives
Our objectives were to evaluate the biodiversity values and trends in a montado system in the Alentejo, Portugal so as to inform landscape level conservation approaches. In doing so, we aimed to develop a replicable and robust approach drawing together field observation, expert opinion, and remote sensing to produce predictions relevant to land management planning.
Methods
Field sampling and subsequent analysis of data on the birds, butterflies and plants in eight distinct land covers allowed the identification of two principal habitat groupings of importance: ‘montado mosaic’ and ‘shrubland’. Morphological spatial pattern analysis was performed on Landsat-derived GIS habitat layers for 1984 and 2009, generating maps and statistics for change in the different landscape functional classes. In addition, we demonstrated how the modelling of ecotones between open and closed biomes can identify the preferred hunting grounds of the threatened Iberian lynx and black vulture, flagship species whose conservation provides benefits to the area’s wider biodiversity values.
Results
Total and core area of montado mosaics and shrubland increased over the 25 year period, whilst the amount of habitat connectivity declined in the case of shrubland. Considerable local variation in these trends highlighted targetable areas for conservation action (e.g. through agri-environment spending).
Conclusions
A rapid and robust approach was demonstrated, with potentially wider utility for biodiversity assessment and planning.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The wood pastures of southern Portugal (we use the Portuguese term montados hereafter) are an important agroforestry system and landscape type that is characteristic of the western Mediterranean region as a whole (Blondel and Aronson 1999). Sometimes also referred to as oak savannahs, they comprise a low density of holm oak (Quercus ilex) or cork oak (Q. suber) trees (30–60 per hectare) and understorey of grassland and shrub species (Díaz et al. 1997; Aronson et al. 2009). As well as being economically important, they are of high value for biodiversity, representing ecotonal habitats supporting species that specialise in forests and open habitats (Tellería 2001; Díaz 2008; Bugalho et al. 2009; Godinho and Rabaça 2010; Bugalho et al. 2011a, c). Farmland and forest species benefit from areas with different tree densities and patches of Mediterranean shrubs interspersed with pastureland (Godinho and Rabaça 2010). The conservation importance of montados is recognised by their status as protected Annex I ‘Habitats of Community importance’ under the European Community Habitats Directive (Council Directive 92/43/EEC on the Conservation of wild habitats and of wild fauna and flora), and by the protected species that they harbour. The patch-mosaic of open and closed land typical of a montado provides habitat for endangered Iberian lynx (Lynx pardinus) and black vulture (Aegypius monachus) (Diaz and Campos 1997). Forests, dense woodlands and shrubs serve as shelter for the lynx, while open landscapes supply their prey, commonly rabbits. Transition zones (shrubland–pastureland ecotones) are considered particularly important for prey and predator alike (Fernández et al. 2003; Calvete et al. 2004; Curado and Lourenço 2011).
Conservation strategies for these landscapes depend on an understanding of landscape patterns and how they are generated (Alados et al. 2004). In the Mediterranean region, significant changes to the characteristically heterogeneous landscapes result from agricultural abandonment or intensification, urban development and other socio-economic processes (Atauri and de Lucio 2001; Plieninger 2006; Koniak et al. 2011). Such changes in land use and landscape structure have profound effects on biodiversity; the challenge is to identify and apply metrics that provide information at the landscape level on biodiversity change (Gude et al. 2007; Walz and Syrbe 2013). These metrics can be based on different approaches, including direct measurements of the presence of forest species (Ochoa-Quintero et al. 2015), key/focal species (Watts et al. 2010) or areas of habitat (White et al. 1997). The spatial configuration and connectivity of habitat, as they affect species dispersion and movement or minimum areas of territory, is often considered (Saura et al. 2011) within island biogeographical theory of species–area relationships, upon which predictions can be made based on quantifying the changing area of patches. An important conceptual development in this respect is that of ‘countryside biogeography’ (Mendenhall et al. 2014), which demonstrates that farmland is not a neutral matrix within which forest (or other habitat) fragments sit, but itself supports novel species assemblages. This challenges the notion, found in many studies of land use–biodiversity relationships, of a binary habitat/non-habitat landscape. Through habitat complementation, mosaics of forest fragments and shrubland, for example, can support species that are not present in either one of those habitats alone (Brotons et al. 2005).
In this study we evaluate the biodiversity value and trends in a montado system in the south-east Alentejo, Portugal, to inform landscape level conservation approaches to its fauna and flora. The approach we adopt recognises the importance, especially in a typical Mediterranean landscape, of the mosaic of different vegetation structures and land covers and the extent to which they offer distinct habitats for species (Moreira and Russo 2007). We characterise species assemblages of eight principal land covers of montado mosaics, looking for similarities and differences in species composition and thereby reclassifying the landscape into areas of habitat important for different species groups (Tomaselli et al. 2013). We are then able to intersect this analysis with land cover change information (Allen et al. 2018) to model potential impact on overall biodiversity using Morphological Spatial Pattern Analysis (MSPA) to identify core habitat patches and their connectivity, and the modelling of ecotones. We concentrated on birds (wintering and breeding), butterflies and vascular plants as biodiversity indicators, being three taxonomic groups that are relatively well-studied and conspicuous in the field, and with value for indicating a range of habitat conditions suitable for species of other taxonomic groups—see for example Mandelik et al. (2010) and Herrando et al. (2016). Our aim was to develop a rapid, replicable and robust approach drawing together field observation, expert opinion, and remote sensing techniques to answer the question: how does biodiversity change spatially and temporally in this landscape, and what are the implications for land management planning and conservation policy?
Materials and methods
Study area
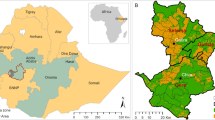
Our study was conducted in an area of 1400 km2 in the vicinity of Moura, Barrancos and Chança in the Alentejo region, Portugal (Fig. 1). This area is the location of an EU LIFE-Nature project ‘Enhancing Habitat for the Iberian lynx and black vulture in the Southeast of Portugal’ (LIFE08 NAT/P/000227), led by the Liga para a Protecção da Natureza (LPN). Eight distinct land cover types within this region were targeted in our study (Table 1). For each type, three replicate sites were selected such that they were representative of the range of conditions/states observed for that habitat in the region. Each site consisted of a habitat patch of at least 50 ha in size, and for each habitat type, the replicates were dispersed as much as possible across the region. Access arrangements with specific landowners meant that some sites of different habitats fell in clusters.

Study area, situated in the Baixo Alentejo region of Portugal and adjoining the Spanish border
Field surveys
Transects
The bird, butterfly and plant surveys were performed along line transects. The approximate route of the transect at each replicate site was defined in advance and adapted during the course of the first counts. The exact route taken by the observer was recorded with GPS waymarks, to ensure the correct distance was travelled. Each transect was of 1 km in length, and avoided habitat edges (usually 150 m from the recognised habitat boundary) and, as far as possible, major tracks or other linear features. The use of narrow tracks was unavoidable for some habitats (e.g. matagal shrubland).
Bird surveys
The bird surveys were undertaken twice to cover wintering birds (January/February) and springtime breeding populations (May). We used the standard line transect methodology, as described by Bibby et al. (2000). The surveys were all undertaken within 3 h of sunrise, in dry weather conditions and good visibility. Whilst the distance of the transect was fixed at 1 km, the duration of recording varied depending on the density of the vegetation: a steady pace of 2 km/h for open habitats and 1 km/h for closed habitats (Gibbons and Gregory 2006). For each of ten sections along the transect, we counted and recorded all birds seen or heard in the distance bands 0–25, 25–100 and > 100 m on either side of the transect route. Double-counting was avoided as best as possible. Birds flying overhead were noted separately.
Butterfly surveys
The butterfly surveys were undertaken in May 2014. An earlier study in the Algarve indicated that this was the optimal month for recording the majority of species (Gardiner, 1994, unpublished). The butterfly recording method was based on that used for the British Butterfly Monitoring Scheme described by Pollard and Yates (1993). The butterfly surveys were organised to occur on the same days as the breeding bird surveys. They were undertaken within the time period 10:00–15:00 h, in sunny weather (or > 20 °C air temperature if cloudy). Wind speed was no higher than force 5 on the Beaufort scale. The observer walked at a steady pace recording all butterflies seen within 2.5 m either side of the route and up to 5 m ahead. No attempt was made to count butterflies flying higher than the recorder. A sweep net was carried by the observer and used to catch and identify individuals when needed. If the recorder was unsure of an identification, the butterfly was recorded as the most common species (according to the expert opinion of the recorder at the time) of possible options (Pollard and Yates 1993).
Vascular plant survey
The vascular plant survey was undertaken 2–10 April 2014, which previous experience showed to be optimal for identifying spring flowering plants. We employed the same survey sites and transects as the fauna surveys. Data collection was from four quadrats of 4 × 4 m size situated at intervals of 200–250 m, with the first starting at least 100 m in from the end of the transect. The distances between quadrats were paced, and their positions recorded by GPS.
Within each quadrat, the presence of vascular plants was noted and cover/abundance was estimated by two observers according to the cover-abundance scale of Braun-Blanquet (1932). Specimens were collected of species not identifiable in the field, and were later identified with the help of flora available in the Herbarium library of the University of Cambridge. Grasses, sedges and rushes, as well as clovers (Trifolium spp.), were often not yet flowering at the time of survey, and were therefore not differentiated to species level. After completion of the quadrat, an extended 10 × 10 m quadrat, encompassing the original one in its corner, was surveyed for woody species, to capture the presence of any additional species that by their stature and dispersion would be less likely to be present in the smaller quadrat size.
Definition of habitats
We were most interested in a comparison of the species composition of the different land covers, and for this purpose two methods were employed. Firstly, we used nonmetric multidimensional scaling (NMDS) ordination (Minchin 1987) on the species data to map the relative similarities and differences of site species composition, as indicated by clustering and dispersion in the environmental space of the plotted ordination diagram. Land covers were grouped by visual identification of these clusters where present. Secondly, having identified these groupings, the species of bird, butterfly or plant responsible for differentiating groupings (i.e. being characteristic of them) were searched for using the methods of tabular comparison. A full explanation of this approach, adapted from the field of phytosociology, is detailed by Mueller-Dombois and Ellenberg (1974).
The species abundance scores used in NMDS varied between groups. For most species of birds, an estimate of bird density (Gibbons and Gregory 2006) was not possible because there were too few observations to calculate a detectability function. Instead, for each species a relative abundance score was calculated, being the total number of individuals seen in the three counting bands as a proportion of all records. Birds flying overhead were not included. Similar relative abundance scores for butterflies at each site were derived from the total number of individuals per species recorded along the transect within the counting region. For plants, the ordinal scale cover-abundance values were converted to percentage cover based on Braun-Blanquet’s (1964) conversion used by Maarel (1979). They were then averaged across the four quadrats for each transect.
For each of the identified habitats, species richness (mean and maximum values), species diversity according to the Shannon index (Magurran 2004), and number of habitat specialists (species occurring in no more than two of the land covers) were calculated.
Change in habitat area and connectivity
Landscapes are characterised by mosaics of habitat patches and associated land uses, in which biodiversity and ecological processes are contained and connected by linkages or zones of dispersion. The spatial configuration of these mosaics contributes to the integrity of the landscape for maintaining healthy ecosystems and biodiversity features (Walz 2011). We used Morphological Spatial Pattern Analysis (MSPA; Vogt et al., 2007) to perform a connectivity analysis of ecosystems of interest in our study area. MSPA has been successfully utilised in several conservation applications to assess the spatial dynamic and function of habitats: for example, habitat connectivity and suitability index modelling (Elbakidze et al. 2011); least cost path analysis and scenario modelling of land use changes (Höbinger et al. 2011); and, habitat fragmentation and loss (Wickham et al. 2010; Chuvieco et al. 2013). Using simple mathematical operators and a binary classification of pixels designated as either foreground or background (habitat of interest vs. no habitat respectively), MSPA segments pixels into seven mutually exclusive landscape categories (Table 2).
We selected two habitat combinations for connectivity modelling (see results 3.3 below). For each, we used the following user-defined operations within MSPA: a 2-pixel (c. 60 m) edge-width and an eight cell connectivity search window. To reduce the effect of fragmentation within large habitat patches and to remove small isolated habitat patches, the habitat layers were generalised with a 3 × 3 smoothing window. The MSPA output produces maps that show, for 1984 and 2009, the extent of the seven landscape connectivity classes.
Ecotone modelling
Many species require ecotones between open and closed habitats to provide their full resource and dispersal requirements. Open and mixed areas are preferred by rabbits for grazing (Lombardi et al. 2007) and therefore by top predators, such as the Iberian lynx, for hunting, but for example the lynx will only access hunting grounds from a closed habitat due to their need for shrubby cover concealment (Rodríguez and Delibes 2002; Curado and Lourenço 2011). Similarly, black vulture and the Spanish imperial eagle (Aquila adalberti) require an ecotone mosaic to access their prey in open areas, but nest in closed areas (Lombardi et al. 2007). Within MSPA, and using the 2009 land cover classification, we identified two open-closed ecotones: (1) at the interface of open and closed habitats (montado/olive groves vs shrubby montado and shrubland); and (2) at the interface of closed habitats and pasture (shrubby montado and shrubland vs. pasture/fallow). We defined the width of the ecotone as 90 m, in line with previous studies in similar Mediterranean habitats (Santos and Tellería 1992). As with the connectivity modelling, the open/closed habitat layers were generalised with a 3 × 3 smoothing window. All open habitat, closed habitat, and pasture classes that did not comprise the ecotone interface were retained. The output creates a map showing pasture, open habitat, closed habitat, and open-closed ecotones.
Results
Species’ usage of different land covers
Fifty-six wintering bird species were recorded in our survey. Fifteen of the species were found in just one or two of the land cover types due to habitat specialism or rarity of observation, whilst 10 species were of conservation concern (EU Birds Directive Annex 1; Table S1 in Supplementary Material). Species richness varied significantly between habitats, with the different montado habitats generally being the richest, and the pinewoods the poorest (Table 3). Bird densities also varied substantially, both within and between land covers. The montados and olive groves held the highest numbers of birds, though the significance of differences between land covers needs to be treated cautiously due to varying levels of detectability and disturbance.
We recorded 100 breeding bird species in the survey plots including 13 of conservation concern (Table S2 in Supplementary Material). Together with an additional 13 bird species observed flying over the survey transects, the consolidated bird list for both wintering and breeding birds numbered 118. Four species registered in the winter were not observed in the springtime survey. Twenty-six breeding bird species were observed in just one or two habitats, whilst 13 species were ubiquitous across all land covers (occurring in two-thirds or more of the survey sites). As for the wintering birds, montado land covers were the most species rich.
We recorded 41 butterfly taxa in the survey; all but two were identified to the species level. Some were ubiquitous (five species were recorded in all 24 sites); three were rarely recorded (three sites or fewer). Fourteen butterfly species were frequently present in the species-rich sites, but absent (or almost so) from the species-poor ones (Table S3 in Supplementary Material).
In our study, 225 vascular plant species were recorded. This is a conservative estimate of species richness, as graminoids and clovers (Trifolium spp.) were not differentiated in the study, and some additional species would become apparent in other seasons. The average species richness of discrete land covers varied between 23 (pines) and 47 (olives) (Table 3). With an average number of species per site of 46/47, olives and high tree density montado were significantly richer in species than other land covers.
Ordination of species data and definition of principal habitats
Ordination of the wintering bird data revealed considerable overlap of species composition in high tree density montado, shrubby montado, esteval, matagal and olive (Fig. 2a). Open fields had a distinctly different fauna, as did the pinewoods. The species composition of low tree density montado fell somewhere in between the main grouping (i.e. of more woody land covers) and open fields. The differentiated table of wintering bird occurrence revealed a grouping of eight species characteristic of the woody habitats, and five species shared by the open fields (Table S4 in Supplementary Material). The species complements of both habitat types were well represented in the low tree density montado, helping to explain the relatively high species richness there. Three species were restricted to the pinewoods.

NMDS ordination of a wintering bird, b breeding bird, c butterfly and d vascular plant species. The clusters are based on land cover categories (except in the case of butterflies, for which species rich and poor sites are distinguished) and the numbers within the clusters identify the sampling sites
Ordination of the breeding bird relative abundance data revealed a strong degree of clustering of the montado, olive and matagal sites, contrasting with the community composition of the open fields and esteval on the one hand, and pinewoods on the other (Fig. 2b). The differentiated table of species occurrence reveals four main groups of bird species responsible for this pattern (Table S5 in Supplementary Material):
-
(a)
Species only occurring in the montado/olive/matagal (23 in total). In addition to these, two species had high constancy, albeit in low numbers, in the three montado land covers: short-toed treecreeper (Certhya brachydactyla) and mistle thrush (Turdus viscivorus).
-
(b)
Species occurring in these land covers as well as pinewood (9);
-
(c)
Species occurring in montado/olive/matagal as well as open and/or esteval, but not pinewood (10)
-
(d)
Species only occurring in pinewood (2)
One of the matagal sites (6 in Fig. 2b) had a distinct species assemblage not falling in the above scheme. This site had a stronger signal of old field plant species in its vegetation composition, as well as a suite of heathland shrubs not present or common in the other matagal sites (Osyris quadripartita, Phillyrea angustifolia, Stauracanthus boivinii). In general, the different avifaunas were based more on which of the rich assembly of montado birds were present in other land cover types, than the presence of any differentiating species within the latter. In particular, the lack of species characteristic of the open field and esteval sites was notable. Tetrax tetrax (little bustard) was observed in one of the open field sites, and was characteristic of this habitat in the winter. Two species (crested tit, Parus cristatus, and coal tit, Parus ater) appeared to be specialists of pine forest.
Ordination of the butterfly data reveals two main groups of sites (Fig. 2c): 11 sites of high species richness (range 21–40, average 31), encompassing all montado (low density, high density, shrubby) sites and two of the matagal sites; and 8 sites of low species richness (range 14–21; average 16), encompassing the third matagal site, the esteval and olive sites, and two of the pine sites. Species richness here was in the range 14–21 (average 16). In addition, four sites (three open field sites and one pine), represented outliers in the ordination diagram, with little affinity to each other and a particularly poor complement of species (range 7–13).
In terms of the flora, the land covers were generally more different from each other than was the case with the fauna, although there were some overlaps (Fig. 2d). Matagal had the most distinct vegetation structure as well as species composition, with a large number of shrubs and other perennials requiring undisturbed conditions and therefore absent from other areas. The esteval sites shared a similar species composition to the shrubby montados according to the ordination analysis, and some of their defining species were also often shared with matagal (Table S6 in Supplementary Material). Another distinct grouping of land covers consisted of the high and low tree density montados and open fields; habitats in which pastureland represented a significant component. Their differential species (Table S7 in Supplementary Material) were often shared with olives, though the latter had their own distinct suite of annuals associated with frequently disturbed ground.
On the basis of the above results, and knowledge gained from the LIFE project on the requirements of Iberian lynx and black vultures, we chose the following two nested groupings of habitats to infer changes in biodiversity quality of the study region for 1984–2009:
-
‘Montado mosaic’: olive groves and montado, shrubby montado (understorey > 1.5 m tall) and shrubland (matagal and esteval). These collectively represent the richest areas for birds and butterflies, and also provide the land-use complementarity important for top predators that seek shelter in denser forested habitats whilst feeding in more open areas (Gonçalves et al. 2011), such as the Iberian lynx and black vulture (Carrete and Donazár 2005).
-
‘Shrubland’: Shrubby montado (understorey > 1.5 m tall) and shrubland (matagal and esteval). This subset of shrubby habitats emphasises areas with a distinct woody flora, a specialised community of birds benefiting from multi-layered woody habitat, e.g. tits (Parus spp.), chaffinches (Fringilla coelebs), leaf warblers (Phylloscopus spp.) and other forest birds (Díaz 2009; Pereira et al. 2012), and potential shelter for the aforementioned top predators.
Change in principal habitat area and connectivity
Allen et al. (2018) report on the change of land covers between 1984 and 2009. Areas of trees with herbaceous vegetation (montado and olive groves) increased in area by approximately 33% and shrubland by approximately 30%. By contrast areas of pasture and trees/shrubs (mostly shrubby montado) declined by approximately 28 and 44% respectively. Simplistically, the increase in extent of montado, olive groves and shrubland is of potential benefit for the biodiversity associated with these habitats, including the lynx and vulture. However, the spatial arrangement of these habitats and their changing configuration, as revealed by MSPA, is of critical importance. When considering the montado mosaic (montado and olive groves, shrubby montado and shrubland) (Table 4, Fig. 3a), total area increased between 1984 and 2009 by about 19% (from 47,148 to 56,043 ha) and core habitat area by over 6500 ha. Connectivity also improved with an increase in bridge corridor habitats, while edge areas declined. These results suggest an improvement in the regularity of patch shapes with more bridges linking core habitat. Areas of loop also increased, highlighting potential connectivity between core habitat patches which is not yet being realised. For shrubland (Table 5, Fig. 3b), MSPA results show that while total area decreased between 1984 and 2009 by about 17% (from 8345 to 6951 ha), core habitat area increased from 28 to 35% of total which represents an overall improvement to patch shape. However bridge habitat, and therefore connectivity, decreased. It is unclear whether this has occurred through habitat loss increasing fragmentation or through regeneration of habitat around core patches creating larger contiguous patches. Area of edge also increased, which has implications for areas of ecotonal habitat.

a: Functional classes for montado mosaic habitat (montado, shrubby montado, olive groves and shrubland) for 1984 and 2009. b: Functional classes for shrubland habitat (shrubby montado, matagal and esteval) for 1984 and 2009
Ecotone modelling
An example close-up from the modelled ecotone map for 2009 is shown in Fig. 4. Over the whole study area, closed habitat amounts to 9675 ha while open habitat (which includes open agricultural land) comprises about 45,550 ha. The ecotone between these two habitats is 9265 ha, or 14% of the total area. This represents potentially sensitive and important habitat for predator resource requirements, though this needs confirmation through field observation.

Ecotone mapping for open biome and closed biome interface for the year 2009, showing an area north of Barrancos at the western edge of the study area (compare Fig. 1)
Discussion
Biodiversity value of the montado landscape
This study has confirmed the importance of the montado mosaic landscapes for biodiversity (Blondel et al. 2010). Our field surveys revealed 10 wintering bird species of conservation concern (EU Birds Directive Annex 1; Table S1 in Supplementary Material). Two of these are classified as Near Threatened in the IUCN Red List: Dartford warbler (Sylvia undata) and little bustard (Tetrax tetrax). Sylvia undata, and another Annex 1 species, woodlark (Lullula arborea), were common across all the studied land covers. Montado landscape mosaics are noted for offering important food resources for over-wintering birds. Most of the estimated 60–70,000 overwintering common cranes (Grus grus) in Iberia are to be found in these areas between November and February (Diaz and Campos 1997). Such areas also support 6–7 M wood pigeon (Columba palumbus), as well as many passerines including species favouring shrubland for refuge and food (e.g. Sylvia species) and those associated with the tree layer (e.g. the insectivorous great tit (Parus major) and blue tit (P. caeruleus)) which feed on arthropods among the foliage of oak trees (Diaz and Campos 1997; Onofre 2007).
Of the 100 species of breeding bird recorded in the survey plots, 26 species were of conservation concern (EU Birds Directive Annex 1, Table S2 in Supplementary Material), three of which are also classified as Near Threatened in the IUCN Red List: red kite (Milvus milvus), Dartford warbler (Sylvia undata) and little bustard (Tetrax tetrax). The six raptor species observed flying over the survey transects benefit from the mosaic nature of the montado landscape (Onofre 2007); all but one of these (honey buzzard; Pernis apivorus) are Annex 1 Birds Directive protected species, and black vulture (Aegypius monachus) is assessed as Near Threatened, with decreasing population trend, in the IUCN Red List. Black vulture, like black stork (Ciconia nigra), breed in remote and dense Mediterranean forests, but utilise neighbouring montado landscapes as their main feeding areas (Diaz and Campos 1997).
Montados were the richest in avifauna of the range of land covers we surveyed in this mosaic landscape. Such habitats are considered to have the highest breeding bird richness of any habitat in Iberia (Telleria et al. 2001). This is explained by the ecotonic nature of these habitats, supporting both farmland species and forest species benefiting from a range of tree densities (Godinho and Rabaça 2010). Furthermore, they offer good hunting areas for raptors due to prey abundance and accessibility (low tree cover and herbaceous field layer) (Onofre 2007).
While montados are not noted for their butterfly fauna (Slancarova et al. 2015), nine of the butterfly species recorded in our study were endemic to SW Europe and NW Africa. Analysing their documented habitat preferences, many of this group are known from lightly-wooded or scrubby, flower-rich, dry, hot areas. Their food plants include various species of Brassicaceae (crucifers), Fabaceae (legumes) and Poaceae (grasses), in two cases the Mediterranean arborescent shrub Rhamnus alaternus, and in another two cases Quercus ilex or other oaks. In a comparison of butterfly habitats in central Spain, the highest diversities were found in the open oak woodlands rather than forests and other habitats (Viejo 1989), and our results are consistent with this pattern. Shrubland removal as a forest fuel management practice has been shown to be beneficial for butterflies by returning areas to an early successional state dominated by a herbaceous layer, although the value of undisturbed patches benefiting some specialist species improves diversity at the landscape scale (Verdasca et al. 2012).
Montado systems in Iberia are characterised by botanically rich herbaceous plant assemblages (Díaz-Villa et al. 2003; Bugalho et al. 2011b), more so than open pastures (Moreno et al. 2016). More than 135 species of herbaceous plant and grass were recorded in a 0.1 ha plot in a previous study, being one of the highest known plant diversities in the world at this scale (Díaz-Villa et al. 2003). Management practices that encompass areas with and without grazing increase habitat heterogeneity and herbaceous plant (as well as invertebrate) diversity (Bugalho et al. 2011c). The richness values we recorded for the montado land covers (averaging 33 for low tree density and 46 for high tree density plots) can be compared with a study of Spanish montados (dehesas) in which an average of 27 species per 4 m2 was recorded in open pastureland (Marañón 1986). In this latter study, the equivalent value was 16.5 under the canopy of the trees, and 31.9 at the canopy edge. In our survey, we did not sample under the oaks, but did observe that whilst they seemingly had a reduced number of species, there were shade and disturbance tolerant plants there which would increase the overall (gamma) diversity of these systems.
In summary, our field results indicated that, with the possible exception of esteval shrubland, all land covers contribute significantly to landscape-level biodiversity. Montados were the most biodiverse and should be a priority for conservation. Shrubland encroachment into montado and succession to matagal is unlikely to harm the butterfly fauna as long as open, grassy and herb-rich areas remain in the landscape. A balance between shrubby and open montados would seem to be most beneficial for birds, whilst open field sites are also important for overwintering steppe grassland specialists. Traditional management of olive groves produces a high plant diversity in our study area, but these sites are unremarkable for the fauna. Pinewoods are species poor, but small extents should be maintained for the few specialist birds that inhabit them. Expansion of areas of undisturbed habitat (such as the ecological corridors created through the LIFE project) will increase the area of rare assemblages of Mediterranean shrubs and perennials.
Changes in biodiversity value of our study montado landscape 1984–2009
We charted significant changes in the areas and functional classes of two habitat groupings—montado mosaics and shrublands—between 1984 and 2009. There was an overall increase in area of montado mosaic of 19%, and decrease of shrubland of 17%. Allen et al. (2018) describe the land cover trends behind these changes, and their drivers. The amount of core habitat increased in both habitat groupings, and this indicates an improvement of conditions for species sensitive to patch size. In the case of montado mosaics, bridge habitat also increased, suggesting improved connectivity between the patches. Notwithstanding changes in the intensity of land use, one can surmise that for the complement of species associated with montado mosaics, habitat conditions will have improved over the study period. The amount of bridge habitat for shrublands decreased, however, and the loss of patch connectivity may have to some degree cancelled any advantage of having greater areas of core habitat. Within these trends for the study landscape as a whole, inspection of the maps (Fig. 3) shows how they are not uniform across the area, but rather, often concentrated in hotspots of dynamic change. The increase in montado mosaics between 1984 and 2009, for example, is largely driven by new areas in the southern landscape of Chança (Fig. 3a). The increase in core shrubland habitat, in part owes itself to the abandonment and subsequent shrub encroachment of hill-top olive groves east of Moura (Fig. 3b; Allen et al. 2018).
The capturing of these trends provides information on how the availability of important habitat has changed, and also the functioning and ecological integrity of the landscape as a whole. The interface between open and closed habitat, as indicated by the ecotone map generated for 2009 (Fig. 4), is a resource requirement shared by many key species in Mediterranean ecosystems. Whilst we did not model change in these ecotones, this would be possible to monitor in the long term using the methods that we demonstrate, and would provide a wider picture of the true impact of land use and development in the landscape. Similarly, our remote sensing-derived products can deliver indicators of overall landscape heterogeneity, which can be more important to some groups of animals than the presence of certain land covers (Atauri and de Lucio 2001).
Implications for conservation policy
The study area is a Natura 2000 Special Protected Area (PTZPE0045) and Site of Community Importance (PTCON0053) and this comes with both land management restrictions and opportunities. Agri-environment scheme funding and forest certification (Bugalho et al. 2011b; Dias et al. 2012) are two possible routes for supporting management interventions that enhance biodiversity on a farm property. Our work aids this process by combining knowledge of species habitat preferences with maps of core/fragmented habitat. It enables better targeting of habitat conservation and restoration measures, and allows farmers and landowners to make decisions about habitat management, for example, providing information to inspire farmers to prioritise field-layer plants in open montado for butterflies and floral diversity, or patches of shrubland for their shrub and tree values, or open fields for over-wintering birds, such as cranes and bustards. In this way landscapes can be made for biodiversity alongside maximising economic returns, for example through the creation of biodiverse permanent pastures (Teixeira et al. 2015).
Creating landscapes that are better for biodiversity is a challenge that needs to be tackled at three levels: landscape design, policy making, and public engagement. For landscape design, our results show core areas of different key habitats, such as shrublands and where these integrate with montados and olive groves to provide the best areas for diverse assemblages of birds, butterflies and plants. Other maps show where ecotones exist between open and closed biomes, the best hunting areas for top predators, such as Iberian lynx and black vulture. The current status of core habitat and ecotones across the region reveals not only where they exist and need conserving, but also where they are noticeably absent or need restoring. In general, maintaining a dynamic (a soft and varied) interface between open and closed habitat areas supports the conditions favoured by key species in many Mediterranean ecosystems. The practical challenge for policy-makers is to design policies that stimulate management approaches to maintaining a mosaic of land cover types. Our maps of land cover changes provide an historical context for making today’s decisions.
We have revealed locations where abandonment of farmed land has been beneficial in increasing core shrubland habitat. The benefits to biodiversity can be further enhanced by ecological corridors to improve their connectivity. Policies to promote corridors include compensation for loss of revenue per hectare of habitat created. Under the LIFE project agreements have been made with 13 olive growers to create 56 ha of corridor, about 8.5 km in length, which connects patches of shrubland in the Adiҫa-Ficalho hills near Moura. In exchange for compensation, with payments negotiated until 2020, farmers have agreed not to clear scrub or disturb the development of herbaceous and shrubby vegetation, not to use chemical products on soils or vegetation, and not to proceed with the harvest of olives or undertake other interventions that might negatively affect natural regeneration of the vegetation. In addition, grazing cattle have been excluded on 27 ha and 48 bird feeders put in place over 18 ha, with the purpose of promoting seed dispersal. It is a good example of cooperation between farmers and conservationists and reconciliation of agricultural and conservation needs, thereby addressing the need for public engagement with building landscapes for conservation.
Conclusions
Our study measured three important emergent landscape properties (Bennett and Radford 2007): the total amount of suitable habitat, landscape composition, and landscape configuration (e.g., access to open hunting grounds for predators sheltering in tall shrubland patches). By using the tools of remote sensing and landscape modelling, we created maps by which it is possible to identify and prioritise areas for conservation management, for example management of shrubland succession or establishing pastures and watering holes for rabbits.
This study has attempted to capture the diversity to be found in three taxonomic groups in a landscape dominated by montado, and to characterise differences between the species assemblages to be found in different landscape elements. We were testing the premise that these elements, with their different biophysical characteristics, would offer different habitat conditions and this would be reflected in the species complement associated with them. Ultimately, we were interested in being able to predict the biodiversity impacts of landscape change, in relation to the relative availability and connectedness of the different habitats.
We have demonstrated an approach that is rapid but robust at making assessments at the landscape level, recognising that the generation of any landscape metric of biodiversity implies a set of generalisations and simplifications. Here we have focussed on a small range of the total taxonomic biodiversity, and our investigation of habitat associations was not able to cover the full variation of soils, tree ages and management. Small-scale features such as stone walls and streams have neither been considered, whilst they can be important micro-habitats (Moreira and Russo 2007) or barriers to connectivity. Different species and species assemblages view landscape connectivity according to their own resource requirements and dispersal constraints. In the current study connectivity was defined according to habitat types, not species requirements. Future applications may look at the dispersal ecology of species and their area and resource requirements to model connectivity across the landscape. This would help to inform habitat corridors which serve to provide connectivity for a suite of species and assemblages.
The mapping of connectivity and ecotones in our assessment is an important decision support tool for identifying critical areas in the landscape which provide vital resources for key species and support ecological processes. Moreover, they may help to understand threats to the integrity of the ecosystem or identify potential conflicts in the landscape between biodiversity and development.
References
Alados CL, Pueyo Y, Barrantes O, Escós J, Giner L, Robles AB (2004) Variations in landscape patterns and vegetation cover between 1957 and 1994 in a semiarid mediterranean ecosystem. Landscape Ecol 19(5):545–561. https://doi.org/10.1023/B:LAND.0000036149.96664.9a
Allen HD, Simonson WD, Parham E, Santos E, Hotham P (2018) Satellite remote sensing of land cover change in a mixed agro-silvo-pastoral landscape in the Alentejo, Portugal. Int J Remote Sens. https://doi.org/10.1080/01431161.2018.1440095
Aronson J, Pereira JS, Pausas JG (2009) Introduction. In: Aronson J, Pereira JS (eds) Cork oak woodlands on the edge. Island Press, Washington D.C., pp 1–6
Atauri JA, de Lucio JV (2001) The role of landscape structure in species richness distribution of birds, amphibians, reptiles and lepidopterans in mediterranean landscapes. Landscape Ecol 16:147–159. https://doi.org/10.1023/A:1011115921050
Bennett AF, Radford JQ (2007) Emergent properties of land mosaics: implications for land management and biodiversity conservation. Managing and designing landscapes in conservation—moving from perspectives to principles. Blackwell, London, pp 201–214
Bibby CJ, Burgess ND, Hill DA, Mustoe SH (2000) Bird census techniques. Academic Press, London
Blondel J, Aronson J (1999) Biology and wildlife of the mediterranean region. Oxford University Press, Oxford
Blondel J, Aronson J, Bodiou J-Y (2010) The mediterranean region: biological diversity in space and time. Oxford University Press, Oxford
Braun-Blanquet, J. (1932) Plant Sociology. (trans: Fuller GD, Conrad HS). McGraw-Hill, New York
Brotons L, Herrando S, Martin J-l (2005) Bird assemblages in forest fragments within mediterranean mosaics created by wild fires. Landscape Ecol 19(6):663–675. https://doi.org/10.1007/s10980-005-0165-2
Bugalho MN, Caldeira MC, Pereira JS, Aronson J, Pausas JG (2011a) Mediterranean cork oak savannas require human use to sustain biodiversity and ecosystem services. Front Ecol Environ 9(5):278–286. https://doi.org/10.1890/100084
Bugalho MN, Lecomte X, Gonçalves M, Caldeira MC, Branco M (2011b) Establishing grazing and grazing-excluded patches increases plant and invertebrate diversity in a mediterranean oak woodland. For Ecol Manag 261(11):2133–2139. https://doi.org/10.1016/j.foreco.2011.03.009
Bugalho MN, Plieninger T, Aronson J, Ellatifi M, Gomes Crespo D (2009) Open woodlands: a diversity of uses (and overuses). In: Aronson J, Pereira JA, Pausas JG (eds) Cork oak woodlands on the edge: ecology, adaptive management and restoration. Island Press, Washington DC, pp 33–45
Bugalho MN, Caldeira MC, Pereira JS, Aronson J, Pausas JG (2011c) Mediterranean cork oak savannas require human use to sustain biodiversity and ecosystem services. Front Ecol Environ 9:278–286. https://doi.org/10.1890/100084
Calvete C, Estrada R, Angulo E, Cabezas-Ruiz S (2004) Habitat factors related to wild rabbit conservation in an agricultural landscape. Landscape Ecol 19:531–542. https://doi.org/10.1023/B:LAND.0000036139.04466.06
Carrete M, Donazár J (2005) Application of central-place foraging theory shows the importance of mediterranean dehesas for the conservation of the cinereous vulture. Biol Conserv 126(4):582–590. https://doi.org/10.1016/j.biocon.2005.06.031
Chuvieco E, Martínez S, Román MV, Hantson S, Lucrecia Pettinari M (2013) Integration of ecological and socio-economic factors to assess global vulnerability to wildfire. Glob Ecol Biogeogr 23(2):245–258. https://doi.org/10.1111/geb.12095
Curado N, Lourenço P (2011) Cartografia E Identificação de Áreas Prioritárias Para O Lince-Ibérico E Para O Abutre-Preto. LPN, Lisbon
Dias FS, Bugalho MN, Orestes Cerdeira J, João Martins M (2012) Is forest certification targeting areas of high biodiversity in cork oak savannas? Biodivers Conserv 22(1):93–112. https://doi.org/10.1007/s10531-012-0401-4
Díaz-Villa MD, Marañón T, Arroyo J, Garrido B (2003) Soil seed bank and floristic diversity in a forest-grassland mosaic in Southern Spain. J Veg Sci 14(5):701–709. https://doi.org/10.1111/j.1654-1103.2003.tb02202.x
Díaz M (2008) La Biodiversité de La Dehesa. In: Mosquera MR, Fernández JL, Rigueiro A (eds) Systèmes Agroforestiers 895 Comme Technique Pour Le Gestion Durable Du Territoire—Programme Azahar. AECI, Madrid, pp 183–201
Díaz M (2009) Animal communities. In: Aronson J, Pereira JS, Pausas JG (eds) Cork oak woodlands on the edge. Island Press, Washington DC, pp 205–206
Diaz M, Campos P (1997) The Spanish Dehesas: a diversity of land uses and wildlife. Farming and birds in Europe: the common agricultural policy and its implications for bird conservation. Academic Press, London, pp 178–209
Díaz M, Campos P, Pulido FJ (1997) The Spanish Dehesas: A Diversity in Land-Use and Wildlife. In: Pain D, Pienkowski M (eds) Farming and birds in Europe. Academic Press, London, UK, pp 178–209
Elbakidze M, Angelstam P, Andersson K, Nordberg M, Pautov Y (2011) How does forest certification contribute to boreal biodiversity conservation? Standards and outcomes in Sweden and NW Russia. For Ecol Manag 262(11):1983–1995. https://doi.org/10.1016/j.foreco.2011.08.040
Fernández N, Delibes M, Palomares F, Mladenoff DJ (2003) Identifying breeding habitat for the iberian lynx: inferences from a fine-scale spatial analysis. Ecol Appl 13(5):1310–1324. https://doi.org/10.1890/02-5081
Gibbons DW, Gregory RD (2006) Birds. In: Sutherland WJ (ed) Ecological census techniques, 2nd edn. Cambridge University Press, Cambridge, pp 308–500
Godinho C, Rabaça JE (2010) Birds like it corky: the influence of habitat features and management of ‘montados’ in breeding bird communities. Agrofor Syst 82(August):183–195. https://doi.org/10.1007/s10457-010-9345-4
Gude PH, Hansen AJ, Jones DA (2007) Biodiversity consequences of alternative future land use scenarios in greater yellowstone. Ecol Appl 17(4):1004–1018
Herrando S, Brotons L, Anton M, Paramo F, Villero D, Titeux N, Quesada J, Stefanescu C (2016) Assessing impacts of land abandonment on mediterranean biodiversity using indicators based on bird and butterfly monitoring data. Environ Conserv 43(1):69–78. https://doi.org/10.1017/S0376892915000260
Höbinger T, Schindler S, Seaman BS, Wrbka T, Weissenhofer A (2011) Impact of oil palm plantations on the structure of the agroforestry mosaic of La Gamba, Southern Costa Rica: potential implications for biodiversity. Agrofor Syst 85(3):367–381. https://doi.org/10.1007/s10457-011-9425-0
Koniak G, Noy-Meir I, Perevolotsky A (2011) Modelling dynamics of ecosystem services basket in mediterranean landscapes: a tool for rational management. Landscape Ecol 26(1):109–124. https://doi.org/10.1007/s10980-010-9540-8
Lombardi L, Fernández N, Moreno S (2007) Habitat use and spatial behaviour in the european rabbit in three mediterranean environments. Basic Appl Ecol 8(5):453–463. https://doi.org/10.1016/j.baae.2006.09.004
Magurran AE (2004) Measuring biological diversity. Blackwell, Oxford
Mandelik Yael, Roll Uri, Fleischer Aliza (2010) Cost-efficiency of biodiversity indicators for mediterranean ecosystems and the effects of socio-economic factors. J Appl Ecol 47(6):1179–1188. https://doi.org/10.1111/j.1365-2664.2010.01864.x
Marañón T (1986) Plant species richness and canopy effect in the savanna-like ‘dehesa’ of S.W. Spain. Ecol Mediter 12:131–141
Mendenhall CD, Karp DS, Meyer CFJ, Hadly EA, Daily GC (2014) Predicting biodiversity change and averting collapse in agricultural landscapes. Nature 509(7499):213–217. https://doi.org/10.1038/nature13139
Minchin PR (1987) An evaluation of the relative robustness of techniques for ecological ordination. Vegetatio 69:89–107
Moreira F, Russo D (2007) Modelling the impact of agricultural abandonment and wildfires on vertebrate diversity in mediterranean europe. Landscape Ecol 22(10):1461–1476. https://doi.org/10.1007/s10980-007-9125-3
Moreno G, Gonzalez-Bornay G, Pulido F, Lopez-Diaz ML, Bertomeu M, Juárez E, Diaz M (2016) Exploring the causes of high biodiversity of iberian dehesas: the importance of wood pastures and marginal habitats. Agrofor Syst 90(1):87–105. https://doi.org/10.1007/s10457-015-9817-7
Mueller-Dombois D, Ellenberg H (1974) Aims and methods of vegetation ecology. Wiley, London
Ochoa-Quintero JM, Gardner TA, Rosa I, de Barros Ferraz SF, Sutherland WJ (2015) Thresholds of species loss in amazonian deforestation frontier landscapes. Conserv Biol 29(2):440–451. https://doi.org/10.1111/cobi.12446
Onofre N (2007) A Fauna Dos Montados de Azinho. In: Silva JS (ed) Os Montados—Muito Para Além Das Árvores. Publico/Fundação Luso-Americano para o Desenvolvimento, Lisbon, pp 131–159
Pereira P, Godinho C, Roque I, Marques A, Branco M, Rabaça JE (2012) Time to rethink the management intensity in a mediterranean oak woodland: the response of insectivorous birds and leaf-chewing defoliators as key groups in the forest ecosystem. Ann For Sci 71(August):25–32. https://doi.org/10.1007/s13595-012-0227-y
Plieninger T (2006) Habitat loss, fragmentation, and alteration – quantifying the impact of land-use changes on a spanish dehesa landscape by use of aerial photography and GIS. Landscape Ecol 21(1):91–105. https://doi.org/10.1007/s10980-005-8294-1
Pollard E, Yates TJ (1993) Monitoring butterflies for ecology and conservation. Chapman & Hall, London
Rodríguez A, Delibes M (2002) Internal structure and patterns of contraction in the geographic range of the Iberian Lynx. Ecography 25:314–328. https://doi.org/10.1034/j.1600-0587.2002.250308.x
Santos T, Tellería J (1992) Edge effects on nest predation in mediterranean fragmented forests. Biol Conserv 60(1):1–5. https://doi.org/10.1016/0006-3207(92)90792-L
Saura S, Vogt P, Velázquez J, Hernando A, Tejera R (2011) Key structural forest connectors can be identified by combining landscape spatial pattern and network analyses. For Ecol Manag 262(2):150–160. https://doi.org/10.1016/j.foreco.2011.03.017
Slancarova J, Garcia-Pereira P, Fric ZF, Romo H, Garcia-Barros E (2015) Butterflies in Portuguese Montados: relationships between climate, land use and life-history traits. J Insect Conserv 19(5):823–836. https://doi.org/10.1007/s10841-015-9801-6
Teixeira RFM, Proença V, Crespo D, Valada T, Domingos T (2015) A conceptual framework for the analysis of engineered biodiverse pastures. Ecol Eng 77:85–97. https://doi.org/10.1016/j.ecoleng.2015.01.002
Tellería JL (2001) Passerine bird communities of Iberian Dehesas: a review. Anim Biodivers Conserv 24(2):67–78
Tomaselli V, Dimopoulos P, Marangi C, Kallimanis AS, Adamo M, Tarantino C, Panitsa M, Terzi M, Veronico G, Lovergine F, Nagendra H (2013) Translating land cover/land use classifications to habitat taxonomies for landscape monitoring: a mediterranean assessment. Landscape Ecol 28(5):905–930. https://doi.org/10.1007/s10980-013-9863-3
van der Maarel E (1979) Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 39(2):97–114
Verdasca MJ, Leitão AS, Santana J, Porto M, Dias S, Beja P (2012) Forest fuel management as a conservation tool for early successional species under agricultural abandonment: the case of mediterranean butterflies. Biol Conserv 146(1):14–23
Viejo JL (1989) The Importance of woodlands in the conservation of butterflies (Lep.: Papilionoidea and Hesperioidea) in the centre of the iberian peninsula. Biol Conserv 48:101–114
Vogt P, Riitters KH, Iwanowski M, Estreguil C, Kozak J, Soille P (2007) Mapping landscape corridors. Ecol Indic 7(2):481–488. https://doi.org/10.1016/j.ecolind.2006.11.001
Walz U (2011) Landscape structure, landscape metrics and biodiversity. Living Rev Landsc Res 5:1–35. https://doi.org/10.12942/lrlr-2011-3
Walz U, Syrbe RU (2013) Linking landscape structure and biodiversity. Ecol Indic 31(8):1–5. https://doi.org/10.1016/j.ecolind.2013.01.032
Watts K, Eycott AE, Handley P, Ray D, Humphrey JW, Quine CP (2010) Targeting and evaluating biodiversity conservation action within fragmented landscapes: an approach based on generic focal species and least-cost networks. Landscape Ecol 25(9):1305–1318. https://doi.org/10.1007/s10980-010-9507-9
White D, Minotti PG, Barczak MJ, Sifneos JC, Freemark KE, Santelmann MV, Steinitz CF, Ross KA, Preston EM (1997) Assessing risks to biodiversity from future landscape change. Conserv Biol 11(2):349–360. https://doi.org/10.1046/j.1523-1739.1997.95458.x
Wickham JD, Riitters KH, Wade TG, Vogt P (2010) A national assessment of green infrastructure and change for the conterminous united states using morphological image processing. Landsc Urban Plan 94(3–4):186–195. https://doi.org/10.1016/j.landurbplan.2009.10.003
Acknowledgements
The authors would like to thank land-owners and farmers of the study region for allowing and facilitating access to the field sampling sites, Nuno Curado (formerly of LPN) for his logistical help, and Mário Carmo for the field recording of birds and butterflies. We are grateful to the two anonymous reviewers whose comments helped improve the manuscript. The project was supported by a Cambridge Conservation Initiative Collaborative Fund grant (CCI 05-13-008) sponsored by Arcadia.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Simonson, W.D., Allen, H.D., Parham, E. et al. Modelling biodiversity trends in the montado (wood pasture) landscapes of the Alentejo, Portugal. Landscape Ecol 33, 811–827 (2018). https://doi.org/10.1007/s10980-018-0627-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-018-0627-y