
For in that profession there is no such thing as coincidence. (John le Carré)
Similar content being viewed by others

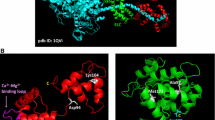

Introduction—domain structure and binding partners
Myosin binding protein C (also known as C-protein: MyBP-C; cardiac isoform abbreviated: cMyBP-C) was discovered in 1973 (Offer et al. 1973) and is located on 7–9 stripes of 43 nm spacing in each half of the A-band (cross bridge bearing zone, C-region) of the sarcomere in skeletal and cardiac muscles (Fig. 1). To date three different MYBPC proteins are known, each encoded by different genes (MYBPC 1–3), where slow (MYBPC1, 1141 aa) and fast (MYBPC2 1141 aa) twitch muscle isoforms are restricted to skeletal muscle tissues and whereas MYBPC3 (1273 aa) is expressed exclusively in cardiac myocytes (Furst et al. 1992; Gautel et al. 1995; Weber et al. 1993). All MYBPC molecules share common architectural features: they are composed of seven immunoglobulin domains (Ig) and three fibronectin type III domains (FnIII) called C1–10 where a 105 amino acid myosin binding protein motif is localized between C1 and C2. In addition, a proline and alanine rich (PA) domain is localized near the aminoterminus (Fig. 2). Domains C9–C10 interact with titin and C10 interacts with light meromyosin (LMM) (Freiburg and Gautel 1996). The actin and S2 (head region) myosin heavy chain binding regions are located at the aminoterminus (Fig. 2). Skeletal muscle MYBPC also interacts with nebulin, but this interaction is not well studied (Jin and Wang 1991) (for a review (Kontrogianni-Konstantopoulos et al. 2009)). Also the MYBPC1-variant 1 interacts with the giant protein obscurin (Ackermann et al. 2009). Another novel interaction between the cardiac specific C0 domain and the regulatory light chain (RLC) has been analyzed by Ratti et al. (2011).

Electron micrograph of rat cardiac muscle labelled with a MYBPC antibody. It shows that MYBPC in cardiac muscle occurs over 9 stripes. The scale bar is 500 nm (Luther et al. 2008) (with Dr Pradeep Luther’s kind permission)

Depicts the structure of MYBPC3 as well as highlights important structural domains and indicates MYBPC3 interacting proteins. Please note: the number of phosphorylation sites localized in the MYBPC motif is species dependent. Ig immunoglobulin, P/A proline/alanine, Fn fibronectin)
In addition, domains in the mid-region (C5–C8) have been hypothesized to interact with one another, forming a “trimeric collar” that constrains the thick filament (Moolman-Smook et al. 2002). Also, a slightly different model of MYBPC organisation within the sarcomere has been proposed by Squire et al. (2003). These authors due to structural considerations and because domains C7–C10 are also reported to interact with titin, which is unlikely to run other than axially along the myosin filament, propose a model whereby the carboxyterminal end of MYBPC is aligned axially, which contrasts the collar alignment structure (Fig. 3).

Summarizes the major differences between the MYBPC models proposed by (Moolman-Smook et al. 2002) (a) and (Squire et al. 2003) (b). Please note in a MYBPC forms a “trimeric collar” that constrains the thick filament (Moolman-Smook et al. 2002). Top: schematic diagram showing a single MYBPC protein interacting with myosin (“top view”), bottom: side view. Whereas in b the carboxyterminal end of MYBPC is aligned axially (Squire et al. 2003). Top view: a thick filament is represented along with six surrounding actin filaments (in striated muscle), the broken circle represents the C0 domain in the MYBPC3 isoform. Bottom: side view, domains C7–C10 running axially along the myosin filament backbone. A few myosin heads are drawn as “transparent” ghosts to indicate their position (grey circles: MYBPC3 with green indicating the P/A rich domain; yellow/brown: myosin, dark grey: actin; figures are from: (Squire et al. 2003) with kind permission from the authors and the publisher (Journal of Molecular Biology))
However three major differences exist between the cardiac and skeletal muscle isoforms: MYBPC3 consists of an additional immunoglobulin domain (C0) at its amino-terminus, the domain C5 contains a proline rich 25 residue insertion and several phosphorylation sites are species dependent localized in the myosin binding motif (Gautel et al. 1995; Mohamed et al. 1998).
Implication in human disease
MYBPC3 mutations were first reported in 1995, which was an important discovery (Bonne et al. 1995; Watkins et al. 1995a). Indeed HCM is a frequent disease, affecting 1:500 individuals (Maron et al. 2006) and depending on the population analyzed, MYBPC3 mutations are found in up to 40–50% of the genotyped HCM patients (Richard et al. 2003). To date about 200 different MYBPC3 mutations have been reported (Marston 2011; Schlossarek et al. 2011). In general, MYBPC3 mutations are associated with a slightly lower penetrance, later onset of disease and with milder forms of disease progression in comparison to other HCM causing mutations located for example in the beta myosin heavy chain (MYH7) gene (Niimura et al. 1998; Watkins et al. 1995b). However this general statement may not necessarily be true for all MYBPC3 mutations and indeed some MYBPC3 mutations are associated with a poor prognosis. For example—a recently discovered deletion in intron 32 in MYBPC3 (nt 21348-21372, accession no. U91629), leading to a pre-terminal stop codon, has been found at a frequency of about 4% in India and South East Asia, with significant impact on human heart failure (Dhandapany et al. 2009; Waldmuller et al. 2003). While this mutation has been initially described in HCM patients, it is important to note that this mutation is also associated with other types of cardiomyopathy such as DCM. Other MYBPC3 mutations primarily found in DCM patients have also been reported, for example the Asn948Thr missense mutation (Daehmlow et al. 2002).
In addition the MYBPC3 Arg502Trp mutation with a frequency of about 2.4% is the most common HCM-causing mutation among individuals of European descent in the USA (Saltzman et al. 2010). However other mutations may be prevalent in different European populations, such as in the Netherlands, where three different founder mutations are present: (i) the c.2373_2374insG MYBPC3 which is present in the great majority of HCM patients (up to 25%) and where (ii) the c.2864_2865delCT and (iii) the c.2827C > T mutations occur in about 5% of HCM patients each (Alders et al. 2003; Christiaans et al. 2010).
Beside the important mutations in MYBPC3, MYBPC1mutations are a cause of distal arthrogryposis type 1 (DA1), a disease characterized by congenital contractures of the hands and feet (Gurnett et al. 2010).
Although by now MYBPC3 mutations are well known causes of HCM and DCM including associated heart failure, the underlying molecular mechanisms are not well defined, some of which will be discussed in the next chapters.
Animal models
To gain more insight into the underlying molecular mechanisms, Mybpc3 has been deleted in genetically altered mouse models by two independent groups. Loss of this protein is not associated with any embryonic lethality and MYBPC3 also is not essential for sarcomere formation but its absence results in profound eccentric hypertrophy in the homozygous animals (Carrier et al. 2004; Harris et al. 2002). Hemodynamic analysis in the mice generated by the Carrier group, where exons 1 and 2 have been deleted, revealed the presence of normal contractility but severe diastolic defects. In addition heterozyogous animals develop septal hypertrophy, a hallmark of HCM (Carrier et al. 2004). However these animals were engineered so that they harbour a complete ablation of the gene and therefore are useful to identify basic mechanisms, but the overwhelming majority of human mutation carriers express mutant mRNAs and probably proteins, which makes it difficult to relate these data directly to the situation in patients.
Therefore, in addition to the pure knockout models, wildtype Mybpc3 and different Mybpc3 mutants have been overexpressed in various models such as an amino-terminal truncated MYBPC3, which mimics a certain type of human mutations which lead to the loss of the carboxyterminal domains including the titin and myosin binding sites. Overexpression of the mutant, but not the wildtype protein, caused major features of HCM including hypertrophy and an increase in calcium sensitivity (Yang et al. 1998).
Whereas above mentioned transgenes were well expressed at the mRNA and protein levels, overexpression of a Mybpc3 mutant lacking only the myosin binding domain resulted in the expression of only very modest levels of mutant protein (i.e. about 5%) which led to a mild hypertrophy and heart failure phenotype (Yang et al. 1999).
Two additional knock in mouse models have been engineered to carry mutations found in patients and which affect titin and myosin binding. Interestingly animals homozygous for these mutations develop a DCM like phenotype with depressed contractility and hypertrophy (McConnell et al. 1999). Another knock in mouse model was generated such that the mutant protein did not contain the aminoterminal myosin binding domain. The mutant protein was readily integrated into the sarcomeres of heterozygous and homozygous animals, was PKA phosphorylatable and major structural defects could not be detected. However this mutation was associated with a significant increase in calcium sensitivity (Witt et al. 2001). An additional mouse knock in model, based on a G > A transition located on the last nucleotide of exon 6 and which was found in a patient with HCM, has been generated by Lucie Carrier’s group. Interestingly this mutation gives rise to three different mRNAs: (i) missense mutation (ii) nonsense due to exon skipping, frameshift and premature stop codon and (iii) deletion/insertion as nonsense but with additional partial retention of a downstream intron which restores the reading frame and which leads to an almost full length protein. Homozygous animals develop hypertrophy, interstitial fibrosis and decreased myocardial function whereas heterozygous animals do not have any obvious phenotype (Vignier et al. 2009).
Additional genetically altered mouse models have been generated to study MYBPC3 phosphorylation, which will be discussed in the next chapter.
Phosphorylation
Human MYBPC3 contains at least four phosphorylation sites which are localized inside the myosin binding motif (serines 275, 284, 304, with an additional phosphorylation site not unequivocally identified) (Copeland et al. 2010)), the mouse myosin binding motif contains three to four sites (serines 273, 282, 302, (305)) and five sites have been identified in the canine genome as well as three sites in the rat genome (Jia et al. 2010; Yuan et al. 2006, 2008). These phosphorylation sites are targets of protein kinase A (PKA; serines 273, 282, 302) (Gautel et al. 1995), protein kinase C (PKC; serines 273, 302) (Mohamed et al. 1998), calmodulin dependent kinase II (CamKII; serine 302) (Gautel et al. 1995), protein kinase D (PKD; serine 302) (Bardswell et al. 2010) and ribosomal S6 kinase (RSK; serine 282) (Cuello et al. 2011).
MYBPC3 phosphorylation also has been studied in various transgenic mouse models, for example lines have been established where the phosphorylation sites (Ser273, Ser282, and Ser302), along with two adjacent sites that could be potentially phosphorylated (Thr272, Thr281), were converted to alanines. While overexpression of wildtype MYBPC3 was able to rescue the MYBPC3 null phenotype, the non-phosphorylatable MYBPC3 was not (Sadayappan et al. 2005). Also a similar approach has been used to analyze MYBPC3 phosphorylation by generating animals whereby only the three known phosphorylation sites were converted into alanines (t3SA). Again these animals showed, when crossed into the Mybpc3 null background, in addition to hypertrophy systolic and diastolic defects, further supporting the notion that PKA phosphorylation of MYBPC3 is important for myocardial function (Tong et al. 2008). Another type of transgenic animals, where the phosphorylation motif “LAGAGRRTS” was completely deleted, where also generated. These animals showed an increase in contractility and relaxation of about 22 and 25%, respectively. An increase in the phosphorylation of the remaining MYBPC3 and other proteins such as troponin I and phospholamban was also observed (Yang et al. 2001).
The reverse experiment, i.e. overexpression of a phosphomimetic MYBPC3 in a Mybpc3 null background, has also been performed, which resulted in subtle changes in sarcomeric ultrastructure characterized by increased distances between the thick filaments which may indicate that phosphomimetic MYBPC3 affects thick–thin filament relationship. The phosphomimetic MYBPC3 also prevented interaction with myosin heavy chain in vitro as analyzed by yeast two hybrid and pull down assays. Moreover these transgenic animals were also resistant to ischemia reperfusion injury. This effect is difficult to explain, but it may well be that phosphomimetic MYBPC3 prevents an increase of thick filament packing density which is associated with reduced calcium activated force generation. An alternative explanation could be that phosphorylated MYBPC3 is protected from calpain (see also later) mediated proteolysis (Sadayappan et al. 2006, 2009). In this context it is probably important to mention, that loss of MYBPC3 phosphorylation also has been observed in failing human hearts and strategies to increase its phosphorylation may have cardio-protective effects (Copeland et al. 2010; Sadayappan et al. 2006).
Above mentioned studies have been intensified by generating mouse models overexpressing various single or double phosphomimetic or non phosphorylatable serines in the Mybpc3 knockout background (i.e. the cMyBP-CSAS(t/t), cMyBP-CADA(t/t), cMyBP-CDAD(t/t); on position S-273, S-282 and S-302, respectively). One major conclusion of these studies is that S-282 has a unique regulatory role in that its phosphorylation is critical for the subsequent phosphorylation of S-302, but that all residues play a role in regulating the contractile response to β-agonist stimulation (Sadayappan et al. 2011).
Although there is now compelling evidence supporting the notion that MYBPC3 phosphorylation modulates contractility by controlling the proximity of the myosin heads to actin, however the precise molecular mechanism remains unclear (reviewed in (Barefield and Sadayappan 2010; Schlossarek et al. 2011)). In particular when using in vitro systems it was shown that MYBPC3 phosphorylation can abolish the ability of MYBPC3 to interact with the S2 region of the myosin heavy chain but may enhance MYBPC3 interactions with the thin filament. Vice versa—dephosphorylation results in strong binding of MYBPC3 to the myosin head, probably preventing its force generating strong interaction with actin (Kulikovskaya et al. 2003a; Sadayappan et al. 2006). However recent data also indicate that MYBPC3 may act synergistically with the myosin RLC to enhance cross-bridge formation by altering the interaction of the myosin head with actin. It may well be that this interaction depends on phosphorylation of either MYBPC3 or RLC and may well be able to provide another mechanism how phosphorylation of sarcomeric proteins may affect protein/protein interactions and kinetics of force development (Colson et al. 2010). Although it is well known that the distance between thick and thin filaments is a major determinant of calcium sensitivity and that loss of MYBPC3 is associated with increased calcium sensitivity, the precise molecular mechanism of how MYBPC3 phosphorylation affects this system is unknown. Part of this problem is that PKA phosphorylation decreases calcium sensitivity via troponin I phosphorylation which leads to a decrease in tropinin C calcium sensitivity. However MYBPC3 phosphorylation, which decreases calcium sensitivity, has no effect on interfilament spacing (Colson et al. 2010) (for more details on calcium sensitivity see also next chapter). Future studies, based on various transgenic animals, will certainly help to answer this question.
MYBPC3 mutations and molecular mechanisms leading to myocardial dysfunction
In general two different molecular mechanisms may account for the observed pathologic effects of any mutation: (i) poison peptide (functional integration of a mutant protein) (ii) haploinsufficiency (loss of one allele).
However, any mutation may also cause disturbances in the degradation of such a product, including its mRNA. For example, the mRNA might be degraded via nonsense mediated mRNA decay or the protein may be degraded via the ubiquitin proteasome system (UPS) with implications for protein degradation in general, including autophagy.
Most of the more than 600 known HCM causing mutations located in other sarcomeric protein genes are point mutations or single amino acid deletions (Van Driest et al. 2004), hence the poison peptide mechanism seems to be the likely mechanism for these mutations. In contrast, the most frequent type of MYBPC3 mutation affects splice acceptor and donor sites or are deletions or insertions predicted to result in truncated MYBPC3 proteins (Van Driest et al. 2004).
However, multiple studies were unable to detect or document the presence of the predicted truncated MYBPC3 molecules. For example van Dijk and colleagues analyzed the c.2373dupG and the c.2864_2865delCT mutations and found a significant decrease in mutant MYBPC3 mRNA. Also they detected a significant loss of MYBPC3 protein but were unable to detect the truncated proteins (van Dijk et al. 2009). Marston and colleagues studied MYBPC3 protein expression in two patients carrying the Glu258Lys and the Arg502Trp missense mutations in addition to seven other mutations which are predicted to lead to premature terminations and truncated proteins. However no truncated protein was detectable and in all samples studied significant lower amounts of MYBPC3 protein was detected (Marston et al. 2009). Another complex mutation where a T to A transition occurred on position 2604 together with a deletion of C at 2605 was predicted to lead to a truncated protein (1-868 + 13 nonsense amino acids with a premature stop codon in C7) was also studied in great detail, but no truncated MYBPC3 protein could be detected (Jacques et al. 2008). In summary, several different studies failed to detect truncated MYBPC3 proteins in MYBPC3 mutation carriers, which makes the poison peptide mechanism unlikely to occur in the studied mutations/individuals. Also these studies documented loss of MYBPC3 protein in mutations carriers, with only 67–75% of the normal protein present, which indicates haploinsufficiency may contribute to the phenotype.
MYBPC3 plays a modulatory role in the regulation of actin–myosin interaction and by binding to both proteins it may directly affect calcium sensitivity. By employing skinned myocytes from patients with HCM mutations, a variety of studies showed that MYBPC3 mutations are associated with higher calcium sensitivity. However, these data may also result from low levels of troponin I (TnI) phosphorylation present in the myectomy tissue rather than the mutation itself (Hoskins et al. 2010; Jacques et al. 2008; van Dijk et al. 2009). Nevertheless an increase in calcium sensitivity in myocardial samples obtained from patients affected by MYBPC3 mutations is likely because removal of MYBPC3 from myocardial samples under experimental conditions is associated with an increase in calcium sensitivity (Hofmann et al. 1991a, b; Kulikovskaya et al. 2003b) and an increase in calcium sensitivity has also been shown in Mybpc3 deficient myocardium (Rybakova et al. 2011). Calcium sensitivity depends on the rate of calcium binding to or calcium release from troponin C and kinetic studies suggest that for HCM causing mutations the rate of calcium binding to TnC is increased, but it remains to be elucidated whether this is a general phenomenon or whether other determinants of calcium sensitivity such as the TnC–calcium dissociation rate may also be affected and whether this holds true for MYBPC3 mutations (Dong et al. 2008).
Therefore, MYBPC3 mutations may primarily increase calcium sensitivity (gain of function) which may cause faster cross bridge cycling (hypercontractility), but which may also cause incomplete relaxation, a cause of diastolic dysfunction frequently observed in HCM. This concept also is supported by hemodynamic data obtained from Mybpc3 deficient animals, where no systolic but severe diastolic dysfunction has been observed (Carrier et al. 2004).
It might well be that MYBPC3 mutations are primarily linked to an increase in calcium sensitivity, but it is difficult to link this event directly to the secondary effects such as myocardial hypertrophy including myofibrillar disarray, fatal arrhythmias and sudden cardiac death (Fig. 4). One hypothesis proposes energy deficiency due to reduced thermal efficiency in HCM or another hypothesis states that troponin C functions as an intracellular calcium buffer or calcium store (Bers 2008). An increase in calcium sensitivity could affect calcium transients, which indeed have been found to be increased in Myh6 Arg403Gln knock in animals (Gao et al. 1999). An increase in calcium transients may also have effects on calcium dependent enzymes such as calcineurin, calmodulin dependent kinase, and protein kinase C all of which have been shown to be important for the initiation of myocardial hypertrophy (Kubis et al. 2003).

Summary of possible pathologic events caused by a primary increase in calcium sensitivity, which has been shown to be present in various Mybpc3 transgenic animal models as well as in patients affected by mutations in the MYBPC3 gene. CamKII calmodulin dependent kinase II, PKC protein kinase C
The hypothesis that sarcomeric HCM mutations lead to inefficient ATP utilization was proposed by the Watkins group and is an attractive model, because it is able to explain many different features found in HCM (Ashrafian et al. 2003). According to this theory a lower ATP/ADP + P ratio may contribute to energy depletion in critical compartments, which may affect calcium re-uptake and lead to an increase in calcium concentrations and thus activation of calcium dependent enzymes such as calcineurin. Another consequence is the activation of adenosine monophosphate kinase (AMPK), which is also able to induce hypertrophy and yet another consequence is the increase in mitochondrial demands which may lead to an increase in reactive oxygen species, mitochondrial abnormalities and apoptosis—all of which have been observed in various patients affected by HCM (Ishikawa et al. 2005; Kavantzas et al. 2000; Unno et al. 2009).
As pointed out above most MYBPC3 mutations lead to haploinsufficiency but some missense mutations might escape targeted removal and might well be expressed in the myocardium. This might particularly be true for the reported aminoterminal mutations G5R, R35W and T59A in the human genome (Sarcomere Protein Gene Mutation Data Base) as well as for the A31P mutation observed in Maine Coon cats (Meurs et al. 2005), all of which are linked to HCM. Ratti and co-workers (Ratti et al. 2011) studied above mentioned missense mutations. While G5R had only mild effects, A31P could not be expressed as a soluble protein and T59A was not chosen as in many other species a threonine residue is substituted by an alanine and no extreme effects were expected, the R35W seemed to be important for binding to RLC.
MYBPC3 mutations by modulating calcium sensitivity may also have effects, either directly or indirectly, on mechanosensation and mechanotransduction. For example, changes in calcium transients caused either via an increase in calcium sensitivity or via inefficient ATP usage, may have effects on calcium dependent enzymes which may in turn affect mechanosensory effects. For example, calcineurin mediated NFAT and GATA4 activation, protein kinase C mediated effects on MYBPC3 itself or calmodulin kinase II effects on phospholamban and or ryanodine receptor phosphorylation may contribute to changes in gene expression and myocardial remodelling. Myocardial hypertrophy and remodelling itself may lead to changes in bio-mechanical properties of the heart, which feeds back to changes in intracellular signalling (Knöll et al. 2002, 2010). The sarcomeric Z-disc is a central nodal point for signalling (Knöll et al. 2011a) and changes in Z-disc structure or composition may well affect its function. In this context, the E3 ubiquitin ligase atrogin 1 has been found to colocalize at the sarcomeric Z-disc with a truncated MYBPC3 which may have consequences for Z-disc mediated signalling (Mearini et al. 2010) (see also later). However it remains unclear whether above mentioned changes in intracellular signalling, if not alone able to cause effects, will sensitize the myocardium at predisposed locations to maladaptation. This may include the possibility of increased shear stress causing septal hypertrophy. It also remains to be seen whether Z-disc mediated apoptotic events play a role in MYBPC3 related HCM (Knöll et al. 2011c).
It remains also unclear, how MYBPC3 mutations are linked to fibrosis as well to intracellular and cellular myofibrillar disarray. It might well be that fibrosis, which is probably an early event in HCM, causes myofibrillar disarray by displacing existing cardiac myocytes, but it remains unclear how intracellular disarray develops. In this context, myofibrillar disarray contributes certainly to inefficient energy use (i.e. cardiac myocytes are not aligned properly). A recent study on HCM myocardial biopsies, including those from patients affected by MYBPC3 mutations, supports the view that fibrosis is an early event and is present even before the onset of hypertrophy (Ho et al. 2010). Inhibition of fibrosis, either via anti TGFβ antibodies or the angiotensin II type 1 receptor antagonist losartan prevented onset of hypertrophy, non myocyte proliferation and fibrosis in genotype positive but hypertrophy negative (pre-hypertrophic) animals (Teekakirikul et al. 2010). These data point as well to the importance of other cell types than cardiac myocytes which are inherent in the myocardium and which contribute significantly to the phenotype (Knöll et al. 2011b).
A novel link between an increase in myofibrillar calcium sensitivity and arrhythmias has been uncovered recently. An increase in calcium sensitivity led to a change in the shape of the ventricular action potentials in mice which results in shorter effective refractory periods, greater beat to beat variability of action potential durations and increased dispersion of ventricular conduction velocities at higher heart rates—all of which may predispose to the occurrance of arrhythmias. These effects were greatest in HCM causing troponin T mutant animals and most importantly, these effects were reproduced when the calcium sensitizer EMD57033 was used and they were reversible when blebbistatin, a calcium de-sensitizer, was employed. Another important finding of this study is that the degree of myocardial hypertrophy does not correlate with the arrhythmia risk (Baudenbacher et al. 2008). Other mechanisms may involve above mentioned energy depletion with consecutive effects on energy dependent ion exchanges (Ashrafian et al. 2003), loss of cardiac myocytes via apoptosis, necrosis or autophagy which may lead to electric isolation of single cardiac myocytes and fibrosis (Teekakirikul et al. 2010), all of which have been shown to contribute to arrhythmogenic events.
Another important approach has been used by Lucie Carrier’s group, when they analyzed the degradation process of MYBPC3. This group found that the E3 ubiquitin ligase atrogin 1 interacts with both, mutant (a 32 kDa MYBPC3 truncated protein, based on a human mutation and which comprises only of domains C0 and parts of C1 termed M7t-cMyBP-C) and wildtype MYBPC3, but only the mutant MYBPC3 was targeted for degradation. This makes atrogin 1 also a novel MYBPC3 interacting protein, but whether the interaction of atrogin 1 and full length wildtype MYBPC3 is physiologically relevant remains to be seen. Interestingly atrogin 1 and M7t-cMyBP-C co-localize at the sarcomeric Z-disc, which points to the importance of this structure in protein degradation processes (Mearini et al. 2010) (reviewed in: (Knöll et al. 2011a)). Their data also indicated that muscle ring finger protein 1 (MuRF1) regulates MYBPC3 mRNA and protein via a posttranscriptional mechanism (Mearini et al. 2010). Also, calpain 1, a ubiquitously expressed calcium dependent protease and which localizes to the sarcomeric Z-disc and I-band via interactions with titin, interacts with MYBPC3 as well and might contribute to MYBPC3 turnover (Jiang et al. 2002; Kontrogianni-Konstantopoulos et al. 2009). In addition, the lysosomal inhibitor bafilomycin A1 caused an increase in truncated MYBPC3 proteins, which points to the possible involvement of autophagy in MYBPC3 related degradation processes.
As briefly mentioned above, domains C5–C8 have been hypothesized to interact with one another, forming a “trimeric collar” that constrains the thick filament. Interestingly, mutations R654H and N755K, both are HCM causing and located in C5, decreased the affinity to C8 about 2-fold and by at least 10-fold, respectively (Moolman-Smook et al. 2002). Disturbances of intra- MYBPC3 interactions by mutations are a completely novel approach to explain the complex phenotype observed in genetically altered animal models as well as in human patients.
Implications for therapy
MYBPC3 mutations like the majority of all other cardiomyopathy causing mutations are associated with incomplete, age and gender dependent penetrance. Powerful epigenetic and environmental factors are thought to have a major impact on the phenotype, which also implies the possibility of effective therapeutical interventions.
If increased calcium sensitivity is the primary event which leads to HCM in MYBPC3 mutation carriers then calcium desensitizers might be able to influence positively disease progression, which has been shown in animal models (Baudenbacher et al. 2008) (see also above). However DCM is associated with decreased calcium sensitivity and precise calibration of calcium sensitivity is certainly important and needs to be monitored properly, if applied in therapy. Another although still very experimental approach is to use short oligonucleotides to intervene in mRNA processing such that “exon skipping” events can be used therapeutically to prevent the synthesis of pathologic mRNAs or to use siRNAs/miRNAs to suppress pathologic mRNAs—an approach potentially important for MYBPC3 mutations. Calcium antagonists such as diltiazem have been successfully used to suppress HCM in the R403Q alpha MHC transgenic mouse (Semsarian et al. 2002) as well as in a mouse model carrying the TnT-I79 N mutation (Westermann et al. 2006), which led to an ongoing human trial (NCT00319982). However it will be important to find out, whether MYBPC3 mutation positive patients will respond appropriately to this type of treatment.
Also overexpression of SERCA2a in a Glu180Gly tropomyosin (Pena et al. 2010) as well as parvalbumin overexpression in a Glu180Gln tropomyosin animal model rescued the phenotype in both of these lines, which points to the possibility of manipulating the calcium metabolism in order to treat the disease (Coutu et al. 2004). However while phospholamban ablation in muscle LIM protein deficient (Minamisawa et al. 1999) and in Glu180Gly tropomysin transgenic animals (Alves et al. 2010; Gaffin et al. 2011) is favourable, ablation of the very same gene in Mybpc3 transgenic animals is not (Song et al. 2003). It remains to be determined whether these differences are due to the well known differences between human and mouse physiology or whether these differences reflect differences in the underlying molecular mechanisms.
Another possibility might be to intervene at the level of energy metabolism via perhexiline which shifts substrate utilization away from free fatty acids to carbohydrates by inhibiting the enzyme carnitine palmitoyl transferase. Perhexiline improved cardiac energetics, normalized exercise diastolic dysfunction and increased exercise capacity in patients (Abozguia et al. 2010; Lele et al. 1995).
It remains also to be seen whether moderate physical activity in MYBPC3 mutation carriers is favourable, as suggested by animal experimentation (Konhilas et al. 2006), and anti-apoptotic strategies may prove helpful as well. Indeed anti-apoptotic gene expression has been observed when animals were subjected to limited physical exercise (Konhilas et al. 2006). Also calcineurin inhibition may have favourable effects as suggested by animal experimentation (Sussman et al. 1998), but application of available calcineurin inhibitors such as cyclosporine A and tacrolimus, at doses needed to affect calcineurin activity in the heart, is associated with significant side effects such as hypertension, nephro-toxicity and diabetes, which precludes their application in human individuals.
Another way to inhibit hypertrophy is to use statins (3-hydroxy-3-methyglutarylcoenzyme A reductase inhibitors), which act via reducing membrane bound Ras and thus reducing pERK44/42 and ERK1/2 activation which reduced HCM related symptoms in a R403Q beta MHC transgenic rabbit model (Patel et al. 2001; Senthil et al. 2005). However human pilot studies based on these animal experiments remained disappointing (Bauersachs et al. 2007; Nagueh et al. 2010). Therefore it remains to be determined whether this approach is not applicable in general or whether a genotype–phenotype specific analysis will identify patient subgroups more susceptible to this type of treatment.
Summary
Although MYBPC3 has been the subject of intense research for almost over four decades, our understanding of its function in physiology and patho-physiology is still limited. However, MYBPC3 mutations are a major cause of human cardiomyopathy and associated heart failure, with some disease causing mutations having frequencies of up to 4% in various populations. Most MYBPC3 mutations lead to haploinsufficiency and to a primary increase in calcium sensitivity. This may explain major features observed in patients such as the hypercontractile phenotype, changes in calcium transients, myocardial hypertrophy, the defect in relaxation and effects on remodelling including the development of myofibrillar disarray as well as the link to live threatening arrhythmias (Fig. 4).
Future research will focus on the identification of pathways leading from a single MYBPC3 mutation to the complex disease phenotype in patients. These pathways should be defined for every single “private” mutation. Although challenging and certainly difficult to achieve, this approach will undoubtedly result in the development and design of patient specific, novel therapies. Inducible pluripotent stem cells (iPS), gene therapy approaches, whole genome sequencing and determination of the explicit genotypes, as well as medicinal chemistry will have to be combined to increase our knowledge in this field.
Abbreviations
- DCM:
-
Dilated cardiomyopathy
- HCM:
-
Hypertrophic cardiomyopathy
- LMM:
-
Light meromyosin
- MYBPC:
-
Myosin binding protein C (in italics: gene)
- MYBPC1:
-
Slow twitch isoform (in italics: gene)
- MYBPC2:
-
Fast twitch isoform (in italics: gene)
- MYBPC3:
-
Cardiac isoform (in italics: gene)
- cMyBP-C:
-
Cardiac myosin binding protein C
- PKA:
-
Protein kinase A
- RLC:
-
Regulatory light chain
- TnC, I, T:
-
Troponin C, I, T
References
Abozguia K, Elliott P, McKenna W, Phan TT, Nallur-Shivu G, Ahmed I, Maher AR, Kaur K, Taylor J, Henning A, Ashrafian H, Watkins H, Frenneaux M (2010) Metabolic modulator perhexiline corrects energy deficiency and improves exercise capacity in symptomatic hypertrophic cardiomyopathy. Circulation 122(16):1562–1569. doi:CIRCULATIONAHA.109.934059[pii]10.1161/CIRCULATIONAHA.109.934059
Ackermann MA, Hu LY, Bowman AL, Bloch RJ, Kontrogianni-Konstantopoulos A (2009) Obscurin interacts with a novel isoform of MyBP-C slow at the periphery of the sarcomeric M-band and regulates thick filament assembly. Mol Biol Cell 20(12):2963–2978. doi:E08-12-1251[pii]10.1091/mbc.E08-12-1251
Alders M, Jongbloed R, Deelen W, van den Wijngaard A, Doevendans P, Ten Cate F, Regitz-Zagrosek V, Vosberg HP, van Langen I, Wilde A, Dooijes D, Mannens M (2003) The 2373insG mutation in the MYBPC3 gene is a founder mutation, which accounts for nearly one-fourth of the HCM cases in the Netherlands. Eur Heart J 24(20):1848–1853. doi:S0195668X03004664[pii]
Alves ML, Gaffin RD, Wolska BM (2010) Rescue of familial cardiomyopathies by modifications at the level of sarcomere and Ca2+ fluxes. J Mol Cell Cardiol 48(5):834–842. doi:S0022-2828(10)00004-0[pii]10.1016/j.yjmcc.2010.01.003
Ashrafian H, Redwood C, Blair E, Watkins H (2003) Hypertrophic cardiomyopathy:a paradigm for myocardial energy depletion. Trends Genet 19(5):263–268. doi:S0168952503000817[pii]
Bardswell SC, Cuello F, Rowland AJ, Sadayappan S, Robbins J, Gautel M, Walker JW, Kentish JC, Avkiran M (2010) Distinct sarcomeric substrates are responsible for protein kinase D-mediated regulation of cardiac myofilament Ca2+ sensitivity and cross-bridge cycling. J Biol Chem 285(8):5674–5682. doi:M109.066456[pii]10.1074/jbc.M109.066456
Barefield D, Sadayappan S (2010) Phosphorylation and function of cardiac myosin binding protein-C in health and disease. J Mol Cell Cardiol 48(5):866–875. doi:S0022-2828(09)00489-1[pii]10.1016/j.yjmcc.2009.11.014
Baudenbacher F, Schober T, Pinto JR, Sidorov VY, Hilliard F, Solaro RJ, Potter JD, Knollmann BC (2008) Myofilament Ca2+ sensitization causes susceptibility to cardiac arrhythmia in mice. J Clin Invest 118(12):3893–3903. doi:36642[pii]10.1172/JCI36642
Bauersachs J, Stork S, Kung M, Waller C, Fidler F, Hoyer C, Frantz S, Weidemann F, Ertl G, Angermann CE (2007) HMG CoA reductase inhibition and left ventricular mass in hypertrophic cardiomyopathy: a randomized placebo-controlled pilot study. Eur J Clin Invest 37(11):852–859. doi:ECI1877[pii]10.1111/j.1365-2362.2007.01877.x
Bers DM (2008) Calcium cycling and signaling in cardiac myocytes. Annu Rev Physiol 70:23–49. doi:10.1146/annurev.physiol.70.113006.100455
Bonne G, Carrier L, Bercovici J, Cruaud C, Richard P, Hainque B, Gautel M, Labeit S, James M, Beckmann J, Weissenbach J, Vosberg HP, Fiszman M, Komajda M, Schwartz K (1995) Cardiac myosin binding protein-C gene splice acceptor site mutation is associated with familial hypertrophic cardiomyopathy. Nat Genet 11(4):438–440
Carrier L, Knoll R, Vignier N, Keller DI, Bausero P, Prudhon B, Isnard R, Ambroisine ML, Fiszman M, Ross J Jr, Schwartz K, Chien KR (2004) Asymmetric septal hypertrophy in heterozygous cMyBP-C null mice. Cardiovasc Res 63(2):293–304
Christiaans I, Nannenberg EA, Dooijes D, Jongbloed RJ, Michels M, Postema PG, Majoor-Krakauer D, van den Wijngaard A, Mannens MM, van Tintelen JP, van Langen IM, Wilde AA (2010) Founder mutations in hypertrophic cardiomyopathy patients in the Netherlands. Neth Heart J 18(5):248–254
Colson BA, Locher MR, Bekyarova T, Patel JR, Fitzsimons DP, Irving TC, Moss RL (2010) Differential roles of regulatory light chain and myosin binding protein-C phosphorylations in the modulation of cardiac force development. J Physiol 588(Pt 6):981–993. doi:jphysiol.2009.183897[pii]10.1113/jphysiol.2009.183897
Copeland O, Sadayappan S, Messer AE, Steinen GJ, van der Velden J, Marston SB (2010) Analysis of cardiac myosin binding protein-C phosphorylation in human heart muscle. J Mol Cell Cardiol 49(6):1003–1011. doi:S0022-2828(10)00338-X[pii]10.1016/j.yjmcc.2010.09.007
Coutu P, Bennett CN, Favre EG, Day SM, Metzger JM (2004) Parvalbumin corrects slowed relaxation in adult cardiac myocytes expressing hypertrophic cardiomyopathy-linked alpha-tropomyosin mutations. Circ Res 94(9):1235–1241. doi:10.1161/01.RES.0000126923.46786.FD01.RES.0000126923.46786.FD[pii]
Cuello F, Bardswell SC, Haworth RS, Ehler E, Sadayappan S, Kentish JC, Avkiran M (2011) Novel role for p90 ribosomal S6 kinase in the regulation of cardiac myofilament phosphorylation. J Biol Chem 286(7):5300–5310. doi:M110.202713[pii]10.1074/jbc.M110.202713
Daehmlow S, Erdmann J, Knueppel T, Gille C, Froemmel C, Hummel M, Hetzer R, Regitz-Zagrosek V (2002) Novel mutations in sarcomeric protein genes in dilated cardiomyopathy. Biochem Biophys Res Commun 298(1):116–120
Dhandapany PS, Sadayappan S, Xue Y, Powell GT, Rani DS, Nallari P, Rai TS, Khullar M, Soares P, Bahl A, Tharkan JM, Vaideeswar P, Rathinavel A, Narasimhan C, Ayapati DR, Ayub Q, Mehdi SQ, Oppenheimer S, Richards MB, Price AL, Patterson N, Reich D, Singh L, Tyler-Smith C, Thangaraj K (2009) A common MYBPC3 (cardiac myosin binding protein C) variant associated with cardiomyopathies in South Asia. Nat Genet 41(2):187–191. doi:ng.309[pii]10.1038/ng.309
Dong WJ, Xing J, Ouyang Y, An J, Cheung HC (2008) Structural kinetics of cardiac troponin C mutants linked to familial hypertrophic and dilated cardiomyopathy in troponin complexes. J Biol Chem 283(6):3424–3432. doi:M703822200[pii]10.1074/jbc.M703822200
Freiburg A, Gautel M (1996) A molecular map of the interactions between titin and myosin-binding protein C. Implications for sarcomeric assembly in familial hypertrophic cardiomyopathy. Eur J Biochem 235(1–2):317–323
Furst DO, Vinkemeier U, Weber K (1992) Mammalian skeletal muscle C-protein: purification from bovine muscle, binding to titin and the characterization of a full-length human cDNA. J Cell Sci 102(Pt 4):769–778
Gaffin RD, Pena JR, Alves MS, Dias FA, Chowdhury SA, Heinrich LS, Goldspink PH, Kranias EG, Wieczorek DF, Wolska BM (2011) Long-term rescue of a familial hypertrophic cardiomyopathy caused by a mutation in the thin filament protein, tropomyosin, via modulation of a calcium cycling protein. J Mol Cell Cardiol 51(5):812–820. doi:S0022-2828(11)00314-2[pii]10.1016/j.yjmcc.2011.07.026
Gao WD, Perez NG, Seidman CE, Seidman JG, Marban E (1999) Altered cardiac excitation-contraction coupling in mutant mice with familial hypertrophic cardiomyopathy. J Clin Invest 103(5):661–666
Gautel M, Zuffardi O, Freiburg A, Labeit S (1995) Phosphorylation switches specific for the cardiac isoform of myosin binding protein-C: a modulator of cardiac contraction? EMBO J 14(9):1952–1960
Gurnett CA, Desruisseau DM, McCall K, Choi R, Meyer ZI, Talerico M, Miller SE, Ju JS, Pestronk A, Connolly AM, Druley TE, Weihl CC, Dobbs MB (2010) Myosin binding protein C1: a novel gene for autosomal dominant distal arthrogryposis type 1. Hum Mol Genet 19(7):1165–1173. doi:ddp587[pii]10.1093/hmg/ddp587
Harris SP, Bartley CR, Hacker TA, McDonald KS, Douglas PS, Greaser ML, Powers PA, Moss RL (2002) Hypertrophic cardiomyopathy in cardiac myosin binding protein-C knockout mice. Circ Res 90(5):594–601
Ho CY, Lopez B, Coelho-Filho OR, Lakdawala NK, Cirino AL, Jarolim P, Kwong R, Gonzalez A, Colan SD, Seidman JG, Diez J, Seidman CE (2010) Myocardial fibrosis as an early manifestation of hypertrophic cardiomyopathy. N Engl J Med 363(6):552–563. doi:10.1056/NEJMoa1002659
Hofmann PA, Greaser ML, Moss RL (1991a) C-protein limits shortening velocity of rabbit skeletal muscle fibres at low levels of Ca2+ activation. J Physiol 439:701–715
Hofmann PA, Hartzell HC, Moss RL (1991b) Alterations in Ca2+ sensitive tension due to partial extraction of C-protein from rat skinned cardiac myocytes and rabbit skeletal muscle fibers. J Gen Physiol 97(6):1141–1163
Hoskins AC, Jacques A, Bardswell SC, McKenna WJ, Tsang V, dos Remedios CG, Ehler E, Adams K, Jalilzadeh S, Avkiran M, Watkins H, Redwood C, Marston SB, Kentish JC (2010) Normal passive viscoelasticity but abnormal myofibrillar force generation in human hypertrophic cardiomyopathy. J Mol Cell Cardiol 49(5):737–745. doi:S0022-2828(10)00243-9[pii]10.1016/j.yjmcc.2010.06.006
Ishikawa K, Kimura S, Kobayashi A, Sato T, Matsumoto H, Ujiie Y, Nakazato K, Mitsugi M, Maruyama Y (2005) Increased reactive oxygen species and anti-oxidative response in mitochondrial cardiomyopathy. Circ J 69(5):617–620. doi:JST.JSTAGE/circj/69.617[pii]
Jacques A, Hoskins AC, Kentish JC, Marston SB (2008) From genotype to phenotype: a longitudinal study of a patient with hypertrophic cardiomyopathy due to a mutation in the MYBPC3 gene. J Muscle Res Cell Motil 29(6–8):239–246. doi:10.1007/s10974-009-9174-0
Jia W, Shaffer JF, Harris SP, Leary JA (2010) Identification of novel protein kinase A phosphorylation sites in the M-domain of human and murine cardiac myosin binding protein-C using mass spectrometry analysis. J Proteome Res 9(4):1843–1853. doi:10.1021/pr901006h
Jiang LQ, Wen SJ, Wang HY, Chen LY (2002) Screening the proteins that interact with calpain in a human heart cDNA library using a yeast two-hybrid system. Hypertens Res 25(4):647–652
Jin JP, Wang K (1991) Cloning, expression, and protein interaction of human nebulin fragments composed of varying numbers of sequence modules. J Biol Chem 266(31):21215–21223
Kavantzas NG, Lazaris AC, Agapitos EV, Nanas J, Davaris PS (2000) Histological assessment of apoptotic cell death in cardiomyopathies. Pathology 32(3):176–180
Knöll R, Hoshijima M, Hoffman HM, Person V, Lorenzen-Schmidt I, Bang ML, Hayashi T, Shiga N, Yasukawa H, Schaper W, McKenna W, Yokoyama M, Schork NJ, Omens JH, McCulloch AD, Kimura A, Gregorio CC, Poller W, Schaper J, Schultheiss HP, Chien KR (2002) The cardiac mechanical stretch sensor machinery involves a Z disc complex that is defective in a subset of human dilated cardiomyopathy. Cell 111(7):943–955
Knöll R, Kostin S, Klede S, Savvatis K, Klinge L, Stehle I, Gunkel S, Kotter S, Babicz K, Sohns M, Miocic S, Didie M, Knoll G, Zimmermann WH, Thelen P, Bickeboller H, Maier LS, Schaper W, Schaper J, Kraft T, Tschope C, Linke WA, Chien KR (2010) A common MLP (muscle LIM protein) variant is associated with cardiomyopathy. Circ Res 106(4):695–704. doi:CIRCRESAHA.109.206243[pii]10.1161/CIRCRESAHA.109.206243
Knöll R, Buyandelger B, Lab M (2011a) The sarcomeric z-disc and z-discopathies. J Biomed Biotechnol 2011:569628. doi:10.1155/2011/569628
Knöll R, Iaccarino G, Tarone G, Hilfiker-Kleiner D, Bauersachs J, Leite-Moreira AF, Sugden PH, Balligand JL (2011b) Towards a re-definition of ‘cardiac hypertrophy’ through a rational characterization of left ventricular phenotypes: a position paper of the Working Group ‘Myocardial Function’ of the ESC. Eur J Heart Fail 13(8):811–819. doi:hfr071[pii]10.1093/eurjhf/hfr071
Knöll R, Linke WA, Zou P, Miocic S, Kostin S, Buyandelger B, Ku CH, Neef S, Bug M, Schafer K, Knoll G, Felkin LE, Wessels J, Toischer K, Hagn F, Kessler H, Didie M, Quentin T, Maier LS, Teucher N, Unsold B, Schmidt A, Birks EJ, Gunkel S, Lang P, Granzier H, Zimmermann WH, Field LJ, Faulkner G, Dobbelstein M, Barton PJ, Sattler M, Wilmanns M, Chien KR (2011c) Telethonin deficiency is associated with maladaptation to biomechanical stress in the mammalian heart. Circ Res. doi:10.1161/CIRCRESAHA.111.245787
Konhilas JP, Watson PA, Maass A, Boucek DM, Horn T, Stauffer BL, Luckey SW, Rosenberg P, Leinwand LA (2006) Exercise can prevent and reverse the severity of hypertrophic cardiomyopathy. Circ Res 98(4):540–548. doi:01.RES.0000205766.97556.00[pii]10.1161/01.RES.0000205766.97556.00
Kontrogianni-Konstantopoulos A, Ackermann MA, Bowman AL, Yap SV, Bloch RJ (2009) Muscle giants: molecular scaffolds in sarcomerogenesis. Physiol Rev 89(4):1217–1267. doi:89/4/1217[pii]10.1152/physrev.00017.2009
Kubis HP, Hanke N, Scheibe RJ, Meissner JD, Gros G (2003) Ca2+ transients activate calcineurin/NFATc1 and initiate fast-to-slow transformation in a primary skeletal muscle culture. Am J Physiol Cell Physiol 285(1):C56–C63. doi:10.1152/ajpcell.00377.2002
Kulikovskaya I, McClellan G, Flavigny J, Carrier L, Winegrad S (2003a) Effect of MyBP-C binding to actin on contractility in heart muscle. J Gen Physiol 122(6):761–774. doi:10.1085/jgp.200308941
Kulikovskaya I, McClellan G, Levine R, Winegrad S (2003b) Effect of extraction of myosin binding protein C on contractility of rat heart. Am J Physiol 285(2):H857–H865. doi:10.1152/ajpheart.00841.2002
Lele SS, Thomson HL, Seo H, Belenkie I, McKenna WJ, Frenneaux MP (1995) Exercise capacity in hypertrophic cardiomyopathy. Role of stroke volume limitation, heart rate, and diastolic filling characteristics. Circulation 92(10):2886–2894
Luther PK, Bennett PM, Knupp C, Craig R, Padron R, Harris SP, Patel J, Moss RL (2008) Understanding the organisation and role of myosin binding protein C in normal striated muscle by comparison with MyBP-C knockout cardiac muscle. J Mol Biol 384(1):60–72. doi:S0022-2836(08)01135-2[pii]10.1016/j.jmb.2008.09.013
Maron BJ, Towbin JA, Thiene G, Antzelevitch C, Corrado D, Arnett D, Moss AJ, Seidman CE, Young JB (2006) Contemporary definitions and classification of the cardiomyopathies: an American Heart Association Scientific Statement from the Council on Clinical Cardiology, Heart Failure and Transplantation Committee; Quality of Care and Outcomes Research and Functional Genomics and Translational Biology Interdisciplinary Working Groups; and Council on Epidemiology and Prevention. Circulation 113(14):1807–1816
Marston SB (2011) How do mutations in contractile proteins cause the primary familial cardiomyopathies? J Cardiovasc Transl Res 4(3):245–255. doi:10.1007/s12265-011-9266-2
Marston S, Copeland O, Jacques A, Livesey K, Tsang V, McKenna WJ, Jalilzadeh S, Carballo S, Redwood C, Watkins H (2009) Evidence from human myectomy samples that MYBPC3 mutations cause hypertrophic cardiomyopathy through haploinsufficiency. Circ Res 105(3):219–222. doi:CIRCRESAHA.109.202440[pii]10.1161/CIRCRESAHA.109.202440
McConnell BK, Jones KA, Fatkin D, Arroyo LH, Lee RT, Aristizabal O, Turnbull DH, Georgakopoulos D, Kass D, Bond M, Niimura H, Schoen FJ, Conner D, Fischman DA, Seidman CE, Seidman JG (1999) Dilated cardiomyopathy in homozygous myosin-binding protein-C mutant mice. J Clin Invest 104(12):1771
Mearini G, Gedicke C, Schlossarek S, Witt CC, Kramer E, Cao P, Gomes MD, Lecker SH, Labeit S, Willis MS, Eschenhagen T, Carrier L (2010) Atrogin-1 and MuRF1 regulate cardiac MyBP-C levels via different mechanisms. Cardiovasc Res 85(2):357–366. doi:cvp348[pii]10.1093/cvr/cvp348
Meurs KM, Sanchez X, David RM, Bowles NE, Towbin JA, Reiser PJ, Kittleson JA, Munro MJ, Dryburgh K, Macdonald KA, Kittleson MD (2005) A cardiac myosin binding protein C mutation in the Maine Coon cat with familial hypertrophic cardiomyopathy. Hum Mol Genet 14(23):3587–3593. doi:ddi386[pii]10.1093/hmg/ddi386
Minamisawa S, Hoshijima M, Chu G, Ward CA, Frank K, Gu Y, Martone ME, Wang Y, Ross J Jr, Kranias EG, Giles WR, Chien KR (1999) Chronic phospholamban-sarcoplasmic reticulum calcium ATPase interaction is the critical calcium cycling defect in dilated cardiomyopathy. Cell 99(3):313–322
Mohamed AS, Dignam JD, Schlender KK (1998) Cardiac myosin-binding protein C (MyBP-C): identification of protein kinase A and protein kinase C phosphorylation sites. Arch Biochem Biophys 358(2):313–319. doi:S0003-9861(98)90857-8[pii]10.1006/abbi.1998.0857
Moolman-Smook J, Flashman E, de Lange W, Li Z, Corfield V, Redwood C, Watkins H (2002) Identification of novel interactions between domains of Myosin binding protein-C that are modulated by hypertrophic cardiomyopathy missense mutations. Circ Res 91(8):704–711
Nagueh SF, Lombardi R, Tan Y, Wang J, Willerson JT, Marian AJ (2010) Atorvastatin and cardiac hypertrophy and function in hypertrophic cardiomyopathy: a pilot study. Eur J Clin Invest 40(11):976–983. doi:ECI2349[pii]10.1111/j.1365-2362.2010.02349.x
Niimura H, Bachinski LL, Sangwatanaroj S, Watkins H, Chudley AE, McKenna W, Kristinsson A, Roberts R, Sole M, Maron BJ, Seidman JG, Seidman CE (1998) Mutations in the gene for cardiac myosin-binding protein C and late-onset familial hypertrophic cardiomyopathy. N Engl J Med 338(18):1248–1257. doi:10.1056/NEJM199804303381802
Offer G, Moos C, Starr R (1973) A new protein of the thick filaments of vertebrate skeletal myofibrils. Extractions, purification and characterization. J Mol Biol 74(4):653–676
Patel R, Nagueh SF, Tsybouleva N, Abdellatif M, Lutucuta S, Kopelen HA, Quinones MA, Zoghbi WA, Entman ML, Roberts R, Marian AJ (2001) Simvastatin induces regression of cardiac hypertrophy and fibrosis and improves cardiac function in a transgenic rabbit model of human hypertrophic cardiomyopathy. Circulation 104(3):317–324
Pena JR, Szkudlarek AC, Warren CM, Heinrich LS, Gaffin RD, Jagatheesan G, del Monte F, Hajjar RJ, Goldspink PH, Solaro RJ, Wieczorek DF, Wolska BM (2010) Neonatal gene transfer of Serca2a delays onset of hypertrophic remodeling and improves function in familial hypertrophic cardiomyopathy. J Mol Cell Cardiol 49(6):993–1002. doi:S0022-2828(10)00341-X[pii]10.1016/j.yjmcc.2010.09.010
Ratti J, Rostkova E, Gautel M, Pfuhl M (2011) Structure and interactions of myosin-binding protein C domain C0: cardiac-specific regulation of myosin at its neck? J Biol Chem 286(14):12650–12658. doi:M110.156646[pii]10.1074/jbc.M110.156646
Richard P, Charron P, Carrier L, Ledeuil C, Cheav T, Pichereau C, Benaiche A, Isnard R, Dubourg O, Burban M, Gueffet JP, Millaire A, Desnos M, Schwartz K, Hainque B, Komajda M (2003) Hypertrophic cardiomyopathy: distribution of disease genes, spectrum of mutations, and implications for a molecular diagnosis strategy. Circulation 107(17):2227–2232
Rybakova IN, Greaser ML, Moss RL (2011) Myosin binding protein C interaction with actin: characterization and mapping of the binding site. J Biol Chem 286(3):2008–2016. doi:M110.170605[pii]10.1074/jbc.M110.170605
Sadayappan S, Gulick J, Osinska H, Martin LA, Hahn HS, Dorn GW II, Klevitsky R, Seidman CE, Seidman JG, Robbins J (2005) Cardiac myosin-binding protein-C phosphorylation and cardiac function. Circ Res 97(11):1156–1163. doi:01.RES.0000190605.79013.4d[pii]10.1161/01.RES.0000190605.79013.4d
Sadayappan S, Osinska H, Klevitsky R, Lorenz JN, Sargent M, Molkentin JD, Seidman CE, Seidman JG, Robbins J (2006) Cardiac myosin binding protein C phosphorylation is cardioprotective. Proc Natl Acad Sci USA 103(45):16918–16923. doi:0607069103[pii]10.1073/pnas.0607069103
Sadayappan S, Gulick J, Klevitsky R, Lorenz JN, Sargent M, Molkentin JD, Robbins J (2009) Cardiac myosin binding protein-C phosphorylation in a {beta}-myosin heavy chain background. Circulation 119(9):1253–1262. doi:CIRCULATIONAHA.108.798983[pii]10.1161/CIRCULATIONAHA.108.798983
Sadayappan S, Gulick J, Osinska H, Barefield D, Cuello F, Avkiran M, Lasko VM, Lorenz JN, Maillet M, Martin JL, Brown JH, Bers DM, Molkentin JD, James J, Robbins J (2011) A critical function for Ser-282 in cardiac Myosin binding protein-C phosphorylation and cardiac function. Circ Res 109(2):141–150. doi:CIRCRESAHA.111.242560[pii]10.1161/CIRCRESAHA.111.242560
Saltzman AJ, Mancini-DiNardo D, Li C, Chung WK, Ho CY, Hurst S, Wynn J, Care M, Hamilton RM, Seidman GW, Gorham J, McDonough B, Sparks E, Seidman JG, Seidman CE, Rehm HL (2010) Short communication: the cardiac myosin binding protein C Arg502Trp mutation: a common cause of hypertrophic cardiomyopathy. Circ Res 106(9):1549–1552. doi:CIRCRESAHA.109.216291[pii]10.1161/CIRCRESAHA.109.216291
Schlossarek S, Mearini G, Carrier L (2011) Cardiac myosin-binding protein C in hypertrophic cardiomyopathy: mechanisms and therapeutic opportunities. J Mol Cell Cardiol 50(4):613–620. doi:S0022-2828(11)00054-X[pii]10.1016/j.yjmcc.2011.01.014
Semsarian C, Ahmad I, Giewat M, Georgakopoulos D, Schmitt JP, McConnell BK, Reiken S, Mende U, Marks AR, Kass DA, Seidman CE, Seidman JG (2002) The L-type calcium channel inhibitor diltiazem prevents cardiomyopathy in a mouse model. J Clin Invest 109(8):1013–1020
Senthil V, Chen SN, Tsybouleva N, Halder T, Nagueh SF, Willerson JT, Roberts R, Marian AJ (2005) Prevention of cardiac hypertrophy by atorvastatin in a transgenic rabbit model of human hypertrophic cardiomyopathy. Circ Res 97(3):285–292. doi:01.RES.0000177090.07296.ac[pii]10.1161/01.RES.0000177090.07296.ac
Song Q, Schmidt AG, Hahn HS, Carr AN, Frank B, Pater L, Gerst M, Young K, Hoit BD, McConnell BK, Haghighi K, Seidman CE, Seidman JG, Dorn GW II, Kranias EG (2003) Rescue of cardiomyocyte dysfunction by phospholamban ablation does not prevent ventricular failure in genetic hypertrophy. J Clin Invest 111(6):859–867
Squire JM, Luther PK, Knupp C (2003) Structural evidence for the interaction of C-protein (MyBP-C) with actin and sequence identification of a possible actin-binding domain. J Mol Biol 331(3):713–724. doi:S0022283603007812[pii]
Sussman MA, Lim HW, Gude N, Taigen T, Olson EN, Robbins J, Colbert MC, Gualberto A, Wieczorek DF, Molkentin JD (1998) Prevention of cardiac hypertrophy in mice by calcineurin inhibition. Science 281(5383):1690–1693
Teekakirikul P, Eminaga S, Toka O, Alcalai R, Wang L, Wakimoto H, Nayor M, Konno T, Gorham JM, Wolf CM, Kim JB, Schmitt JP, Molkentin JD, Norris RA, Tager AM, Hoffman SR, Markwald RR, Seidman CE, Seidman JG (2010) Cardiac fibrosis in mice with hypertrophic cardiomyopathy is mediated by non-myocyte proliferation and requires Tgf-beta. J Clin Invest 120(10):3520–3529. doi:42028[pii]10.1172/JCI42028
Tong CW, Stelzer JE, Greaser ML, Powers PA, Moss RL (2008) Acceleration of crossbridge kinetics by protein kinase A phosphorylation of cardiac myosin binding protein C modulates cardiac function. Circ Res 103(9):974–982. doi:CIRCRESAHA.108.177683[pii]10.1161/CIRCRESAHA.108.177683
Unno K, Isobe S, Izawa H, Cheng XW, Kobayashi M, Hirashiki A, Yamada T, Harada K, Ohshima S, Noda A, Nagata K, Kato K, Yokota M, Murohara T (2009) Relation of functional and morphological changes in mitochondria to myocardial contractile and relaxation reserves in asymptomatic to mildly symptomatic patients with hypertrophic cardiomyopathy. Eur Heart J 30(15):1853–1862. doi:ehp184[pii]10.1093/eurheartj/ehp184
van Dijk SJ, Dooijes D, dos Remedios C, Michels M, Lamers JM, Winegrad S, Schlossarek S, Carrier L, ten Cate FJ, Stienen GJ, van der Velden J (2009) Cardiac myosin-binding protein C mutations and hypertrophic cardiomyopathy: haploinsufficiency, deranged phosphorylation, and cardiomyocyte dysfunction. Circulation 119(11):1473–1483. doi:CIRCULATIONAHA.108.838672[pii]10.1161/CIRCULATIONAHA.108.838672
Van Driest SL, Vasile VC, Ommen SR, Will ML, Tajik AJ, Gersh BJ, Ackerman MJ (2004) Myosin binding protein C mutations and compound heterozygosity in hypertrophic cardiomyopathy. J Am Coll Cardiol 44(9):1903–1910. doi:S0735-1097(04)01614-6[pii]10.1016/j.jacc.2004.07.045
Vignier N, Schlossarek S, Fraysse B, Mearini G, Kramer E, Pointu H, Mougenot N, Guiard J, Reimer R, Hohenberg H, Schwartz K, Vernet M, Eschenhagen T, Carrier L (2009) Nonsense-mediated mRNA decay and ubiquitin-proteasome system regulate cardiac myosin-binding protein C mutant levels in cardiomyopathic mice. Circ Res 105(3):239–248. doi:CIRCRESAHA.109.201251[pii]10.1161/CIRCRESAHA.109.201251
Waldmuller S, Sakthivel S, Saadi AV, Selignow C, Rakesh PG, Golubenko M, Joseph PK, Padmakumar R, Richard P, Schwartz K, Tharakan JM, Rajamanickam C, Vosberg HP (2003) Novel deletions in MYH7 and MYBPC3 identified in Indian families with familial hypertrophic cardiomyopathy. J Mol Cell Cardiol 35(6):623–636. doi:S0022282803000506[pii]
Watkins H, Conner D, Thierfelder L, Jarcho JA, MacRae C, McKenna WJ, Maron BJ, Seidman JG, Seidman CE (1995a) Mutations in the cardiac myosin binding protein-C gene on chromosome 11 cause familial hypertrophic cardiomyopathy. Nat Genet 11(4):434–437
Watkins H, McKenna WJ, Thierfelder L, Suk HJ, Anan R, O’Donoghue A, Spirito P, Matsumori A, Moravec CS, Seidman JG et al (1995b) Mutations in the genes for cardiac troponin T and alpha-tropomyosin in hypertrophic cardiomyopathy. N Engl J Med 332(16):1058–1064. doi:10.1056/NEJM199504203321603
Weber FE, Vaughan KT, Reinach FC, Fischman DA (1993) Complete sequence of human fast-type and slow-type muscle myosin-binding-protein C (MyBP-C). Differential expression, conserved domain structure and chromosome assignment. Eur J Biochem 216(2):661–669
Westermann D, Knollmann BC, Steendijk P, Rutschow S, Riad A, Pauschinger M, Potter JD, Schultheiss HP, Tschope C (2006) Diltiazem treatment prevents diastolic heart failure in mice with familial hypertrophic cardiomyopathy. Eur J Heart Fail 8(2):115–121. doi:S1388-9842(05)00213-8[pii]10.1016/j.ejheart.2005.07.012
Witt CC, Gerull B, Davies MJ, Centner T, Linke WA, Thierfelder L (2001) Hypercontractile properties of cardiac muscle fibers in a knock-in mouse model of cardiac myosin-binding protein-C. J Biol Chem 276(7):5353–5359
Yang Q, Sanbe A, Osinska H, Hewett TE, Klevitsky R, Robbins J (1998) A mouse model of myosin binding protein C human familial hypertrophic cardiomyopathy. J Clin Invest 102(7):1292–1300. doi:10.1172/JCI3880
Yang Q, Sanbe A, Osinska H, Hewett TE, Klevitsky R, Robbins J (1999) In vivo modeling of myosin binding protein C familial hypertrophic cardiomyopathy. Circ Res 85(9):841–847
Yang Q, Hewett TE, Klevitsky R, Sanbe A, Wang X, Robbins J (2001) PKA-dependent phosphorylation of cardiac myosin binding protein C in transgenic mice. Cardiovasc Res 51(1):80–88. doi:S0008636301002735[pii]
Yuan C, Guo Y, Ravi R, Przyklenk K, Shilkofski N, Diez R, Cole RN, Murphy AM (2006) Myosin binding protein C is differentially phosphorylated upon myocardial stunning in canine and rat hearts—evidence for novel phosphorylation sites. Proteomics 6(14):4176–4186
Yuan C, Sheng Q, Tang H, Li Y, Zeng R, Solaro RJ (2008) Quantitative comparison of sarcomeric phosphoproteomes of neonatal and adult rat hearts. Am J Physiol 295(2):H647–H656. doi:10.1152/ajpheart.00357.2008
Acknowledgments
Prof. Max Lab and Dr. Pradeep Luther are gratefully acknowledged for helpful discussions. Dr. Pradeep Luther is also acknowledged for providing the MYBPC3 related electron microscopy (Fig. 1). Dr. R. Knöll is supported by DFG Kn 448/9-1, DFG Kn 448 10-1, Fritz Thyssen Stiftung, British Heart Foundation (PG11/34/28793) and FP7-PEOPLE-2011-IRSES, Proposal No 291834—Acronym: SarcoSi.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Knöll, R. Myosin binding protein C: implications for signal-transduction. J Muscle Res Cell Motil 33, 31–42 (2012). https://doi.org/10.1007/s10974-011-9281-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10974-011-9281-6