
Abstract
The Late Jurassic docodontan Haldanodon exspectatus from the Guimarota coal mine in Portugal provides important information on the evolution of the dental replacement pattern in mammaliaforms. Haldanodon shows diphyodont replacement of antemolars and non-replacement of molars. Lower incisors are replaced in alternating order with early replacement of i2 and i4, and late replacement of i1 and i3. Upper and lower premolars were replaced sequentially from front to back. In the maxilla and mandible, four deciduous premolar positions are present (dP1-4, dp1-4), but only three permanent premolars (P1/p1, P3/p3, P4/p4) erupt with loss of the dP2/dp2 position. The anterior deciduous premolars (dP1-2/dp1-2) are small and peg-like, dP3-4/dp3-4 are much larger and molariform. The lower canine and dp4 are the last lower teeth to be replaced. The permanent lower premolars are premolariform and consist mainly of large main cusp a. The ultimate permanent lower premolar (p4) erupts at the same time as m4. After replacement of the antemolars, one or two more molars (m5-6) are added at the posterior end of the tooth row of the mandible. Growth of the ramus occurs at the anterior and posterior ends as evident from the much larger permanent canine and addition of m5-6. In the maxilla, all permanent premolars differ morphologically clearly from the molars. The replacing P1 is small and consists mainly of labial cusps A, B, and C. The P3 and P4 are increasingly larger and show a lingual extension with cusps X and Y which is shorter than in the molars. The adult dental formula of Haldanodon is 6I/4i, 1C/1c, 3P/3p, 5 M/5-6m, the deciduous dental formula is ?dI/4di, 1dC/1dc, 4dP/4dp. The tooth eruption sequence in the mandible is m1 → p1 + m2 → i2 + i4 → ?i1 + ?i3 → p3 + m3 → c + p4 + m4 → m5 → m6. Our results corroborate the earlier suggestions that “Peraiocynodon inexpectatus” is based on deciduous lower premolars (dp1-4) of Docodon victor, and that “Peraiocynodon major“ and Cyrtlatherium canei are based on deciduous lower premolars.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Docodontans are Jurassic-Cretaceous non-mammalian mammaliaforms that play an important role for the understanding of early mammalian evolution. They show a remarkable ecomorphological diversity both in dietary and locomotory adaptation (e.g., Krusat 1991; Martin 2005; Ji et al. 2006; Luo et al. 2015; Meng et al. 2015; Panciroli et al. 2022). Docodontans were the first mammaliaforms to evolve a complex molar dentition with a crushing and grinding function, similar to that of molars of tribosphenidans (Jenkins 1969; Gingerich 1973; Butler 1988; Pfretzschner et al. 2005; Brinkkötter 2018; Martin et al. 2020). Their dietary adaptations range from insectivory (e.g., Docodon, Haldanodon), faunivory-carnivory (e.g., Itatodon), piscivory (e.g., Castorocauda) to omnivory-herbivory (e.g., Simpsonodon, Dsungarodon) (Jenkins 1969; Gingerich 1973; Krusat 1980; Kermack et al. 1987; Lopatin and Averianov 2005; Pfretzschner et al. 2005; Ji et al. 2006; Luo and Martin 2007; Schultz et al. 2019; Martin et al. 2020). Deciduous teeth of docodontans are poorly known and were first recognized by Butler (1939) who suggested that the four teeth preserved in the type mandible of “Peraiocynodon inexpectatus” Simpson, 1928 from the Berriasian of the British Purbeck are deciduous premolars (dp1-4), a view that since has been corroborated by subsequent workers (Krusat 1980; Sigogneau-Russell 2003; Averianov 2004). Sigogneau-Russell (2001) demonstrated the docodontan nature of Cyrtlatherium canei Freeman, 1979 from the Bathonian of Kirtlington and suggested that the two isolated lower teeth which are known for this taxon are deciduous premolars. Schultz et al. (2019) re-studied dentaries and maxillae of Docodon victor from the Tithonian of the Morrison Formation by µCT and found three fragmentary mandibles to bear deciduous canines and premolars.
A large growth series of juvenile mandibles and maxillae of the docodontan Haldanodon exspectatus Kühne and Krusat, 1972 has been recovered from the Upper Jurassic lignite of the Guimarota coal mine in Portugal, during more than ten years of field work. After its discovery in 1959 and several short initial field campaigns in 1960–1963, a more extensive year-round field program from 1973 to 1982 yielded a wealth of vertebrate fossils, among which are more than 800 gnathic remains with teeth of mammaliaforms and mammals including 152 mandibular and 67 maxillary remains of Haldanodon exspectatus (see Krebs 2000 for historic overview and Martin and Nowotny 2000). Among the Haldanodon specimens, there are nine mandibles and four maxillae of juvenile individuals of various ontogenetic stages that provide detailed insight into mandibular growth and mode of tooth replacement. Apart from Krusat (1980), who has studied two fragmentary juvenile mandibles (MG 6800 [VJ 1004 − 155] and MG 6801 [VJ 1005 − 155]) from the early field campaigns, only short reports on the juvenile Haldanodon exspectatus specimens have been published so far (Martin and Nowotny 2000; Nowotny et al. 2001; Martin et al. 2010). Here we provide a detailed description and analysis of the juvenile maxillaries and mandibles of Haldanodon exspectatus from the Guimarota coal mine based on µCT analysis and their implications for the evolution of dental replacement and mandibular growth in early mammals. Additionally, previous interpretations on deciduous teeth of docodontans (“Peraiocynodon” and Cyrtlatherium) are discussed in the light of the new data presented here.
Materials and methods
Most mandibles and maxillae were collected between 1973 and 1982. Fossiliferous coaly marl was mined underground exclusively for paleontological purposes, brought to the surface and subsequently split by local workers for the detection of fossils (Krebs 2000). Fossils were marked, hermetically sealed in plastic bags to prevent dehydration and brought to the laboratory at Free University Berlin for preparation. Some specimens derive from the field campaigns of the early 1960s and were collected from coal heaps of the then still active coal mine. Specimens were hardened with cyanoacrylate after being completely freed from the coal matrix (Drescher 2000). The specimens are currently housed in the Institute for Geosciences, Section Paleontology at the University of Bonn, Germany for scientific study. After completion of study, the Guimarota specimens will be transferred to the Geological Museum of the National Laboratory of Energy and Geology of Portugal (MG/LNEG) in Lisbon, Portugal for permanent repository under the MG specimen numbers provided (MG is the acronym used for the Geological Museum [Museu Geológico]). Formerly on various occasions the acronym “SGP” (for Serviços Geológicos de Portugal) was used (e.g., in Lillegraven and Krusat 1991), but this institution was merged into the LNEG in 2003. The Guimarota collection specimen numbers VJ (Vertebrados Jurassicos) and Gui Mam (Guimarota Mammalia) have been widely used in previous publications since 1980 and will be retained with the permanent MG numbers after transferral of the specimens to Lisbon. We include the Guimarota specimen number in brackets, following the MG specimen number.
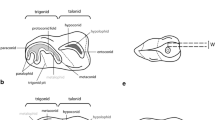
For molar cusp terminology, we follow the alphabetical scheme as outlined by Butler (1997) and Luo and Martin (2007) (Fig. 1). Upper case letters refer to upper teeth and lower case letters to lower teeth: I/i, incisor; C/c, canine; P/p, premolar; M/m, molar; deciduous teeth are denoted by the prefix d (e.g., dP2).

Modified from Luo and Martin (2007: figs. 3 and 4). Cusp “df” stands for “docodont cusp f”, which is not homologous to cusp f of morganucodontans, kuehneotheriids, crown therians, and their near relatives (Luo and Martin 2007: table 1). The lower molar is mirrored. Not to scale
Cusp terminology of upper (a) and lower (b) molars of Haldanodon used in this study.
As “deciduous” we consider teeth that are replaced by successors (or that are shed without replacement) and that differentiate earlier than their successors during ontogeny (Luckett 1993). The developing tooth stages are defined as follows in our study. The “germ” stage is a tiny mineralized structure (in an initial stage of mineralization), detectable by µCT. The “cap” stage is a tooth crown in an advanced stage of mineralization that has not yet developed the full set of crown structures (all cusps and crests). The “crown” stage is a fully developed and mineralized unerupted tooth crown which is ready to erupt but has the roots not yet formed.
The maxillae and mandibles were scanned with varying resolution ranging between 3.5 and 11.0 μm (see Table 1) using the 180 x-ray tube of the v|tome|x s µCT device (GE Sensing & Inspection Technologies GmbH phoenix|x-ray) housed in the Institute of Geosciences, Section Paleontology, University of Bonn, Germany. Scan settings varied between the specimens from 82 to 90 kV and 140–180 µA, and exposure time of 500 or 1000 ms per capture. The µCT device produces isotropic voxels (i.e., same pixel size in each dimension) with a single image size of 2024 × 2024 pixels. Avizo 8 (Thermo Fisher Scientific) was used for segmentation and reconstruction of maxillae, dentaries, and dentitions. Institutional abbreviations: MG, Geological Museum (Museu Geológico) of the National Laboratory of Energy and Geology of Portugal, Lisbon; NHMUK (formerly BMNH), Natural History Museum, London; YPM VPPU, Princeton University Collection, Yale Peabody Museum, New Haven, CT.
Results
Description of maxillae and upper teeth
MG 6834 [Gui Mam 44/81]
The right maxilla fragment is broken anterior to dC and represents the ontogenetically youngest individual examined (Fig. 2). In the part of the maxilla containing dP1-2 the maxilla is laterally constricted, and bears a number of small foramina and pits above dP1 and dC.

Haldanodon exspectatus (MG 6834 [Gui Mam 44/81]). Right maxilla in occlusal (a), lateral (b), and medial (c-d) views; tooth row without maxilla (d). The deciduous canine is highlighted in blue, and deciduous premolars are highlighted in red. 3D models based on µCT-scan data. Scale bar equals 1 mm
The double-rooted deciduous canine has a laterally compressed acute triangular main cusp with a slightly recurved tip which bears a small apical wear facet. On the lingual side of the crown base a weak lingual cingulum is present which is interrupted in the middle. At the crown base there is a small distal accessory cuspule which sits on a heel, and a tiny mesial cuspule. The roots of the dC diverge and are of equal size. The dP1 and dP2 are small and are separated from each other by a small gap.
The dP1 is single-rooted and peg-like, with a triangular, labio-lingually slightly compressed crown tip. It bears a small apical wear facet. The dP2 is slightly larger (mesio-distally longer) and has a broad-triangular, labio-lingually compressed main cusp with a distal wear facet. It possesses a single root which is labio-lingually compressed with a vertical groove on the labial side suggesting an incomplete separation. The dP3 is molarized but less so than the dP4, and 25% smaller. The lingual portion is less developed than in dP4 and cusp Y (Fig. 1a) is very small. The dp3 is moderately worn on the main cusps. Above dP3 an empty crypt for P3 was detected in the maxilla by µCT. The dP4 is more strongly molarized with well-developed lingual portion and distinct cusp Y besides cusp X; it is slightly worn at the apices of the main cusps. Both dp3 and dp4 have one lingual and two labial roots. Posteriorly to dP4 the maxilla is broken and missing.
MG 6742 [Gui Mam 91/76]
The right maxilla fragment is broken anterior to dC and posterior to dP4 (Fig. 3).

Haldanodon exspectatus (MG 6742 [Gui Mam 91/76]). Right maxilla in occlusal (a), lateral (b), and medial views (c). The deciduous canine is highlighted in blue, and the deciduous premolars are highlighted in red. 3D models based on µCT-scan data. Scale bar equals 1 mm
Of the dC, the crown base, distal root and part of the mesial root are preserved. The distal root is diverging posteriorly, and the preserved origin of the mesial root indicates that it was diverging anteriorly. Immediately behind dC a single collapsed alveolus of single-rooted dP1 has been detected. The dP2 has a triangular main cusp with an apical wear facet and a well-developed mesial accessory cuspule at the base. There are two roots of equal size which are spread. Of the molariform dP3 only the labial portion with moderately worn cusps A, B, and C is preserved. The labial crown fragment of dP4 is somewhat displaced and unworn. The maxillary bone is crushed, but a large maxillary foramen is evident dorsal to dP2 and a smaller one dorsal to dP3. Dorsal to the broken canine, about ten small foramina are detectable.
MG 6757 [Gui Mam 128/76]
The right maxilla fragment (Fig. 4) preserves the dC which is somewhat lifted out of its alveoli by the succeeding C and two premolar alveoli. The dC is double-rooted with the mesial root slightly stronger than the distal one. The roots diverge. The triangular crown is labio-lingually compressed and corresponds in size and shape to the dC seen in MG 6834 [Gui Mam 44/81]. It has an acute triangular and slightly recurved main cusp with small apical wear facet and a tiny accessory cuspule on the distal side of the crown base; the accessory cuspule is connected by a faint crest to the main cusp. Lingual of the accessory cuspule, there is a ledge-like heel. A second faint crest connects the main cusp with the lingual end of the heel. The triangular area between these two crests, oriented in disto-lingual direction, is slightly concave. From the mesial side of the crown base runs a tubercular cingulum in disto-cervical direction that fades after two thirds of the crown length. The crown of dC has about half the size of the permanent C and is somewhat more obtuse angled. The acute angled crown of the unerupted permanent C is situated between the roots (mesial root partially resorbed) of dC and has shifted the dC out of its original position. It has double the size of the crown of the dC and is more acute; roots are not detectable because they are not mineralized yet. The exposed part corresponds in size and shape to the C of adult specimens such as MG 6732 [Gui Mam 60/76] (Luo and Martin 2007: fig. 5D-F). Associated with MG 6757 [Gui Mam 128/76], but preserved on a separate fragment of the maxillary without direct contact, are two fully erupted molars with slight wear on the main cusps, probably M1 and M2.

Haldanodon exspectatus (MG 6757 [Gui Mam 128/76]). Fragment of right maxilla in occlusal (a), lateral (b-c), and medial (d) views. The unerupted crown of the upper canine is highlighted in green, and in c, the maxilla is transparent. 3D models based on µCT-scan data. Scale bar equals 1 mm

Haldanodon exspectatus (MG 6731 [Gui Mam 12/74]). Left maxilla in occlusal (a), lateral (b-c), and medial (d) views; in c, the maxilla is transparent, with cap of C, unerupted crowns of P1 and P3, and germ of P4 highlighted in green; e. tooth row without maxilla in lingual view. The roots of the deciduous canine are highlighted in blue, the deciduous premolars in red, and the molars in yellow. 3D models based on µCT-scan data. Scale bar equals 1 mm
MG 6731 [Gui Mam 12/74]
Anteriorly, the left maxilla (Fig. 5) is broken but in front of the roots of dC an alveolus for a single-rooted incisor (I6) is preserved. The crown of dC is broken off. The large roots of dC diverge and deep in the maxilla in between the roots of dC, the enamel cap of the germ of the permanent canine was detected by µCT.
At the position of the first premolar an alveolus with part of the single root of dP1 is visible. Above the root fragment of dP1, the crown of P1 was detected. A small peg-like, unicuspid dP2 is in place. It has a labio-lingually compressed root with vertical grooves on both the lingual and labial sides indicating incomplete separation; apically the roots are fully separated (Fig. 5e). No trace of a developing P2 has been detected inside the maxilla. The main cusp of dP2 bears an apical wear facet that extends onto the lingual side. The dP3 is molarized, with three roots and exhibits strong apical wear on the main cusps. Immediately below dP3, the crown of P3 – ready to erupt – was detected by µCT. The dp4 is in place and is moderately worn. It is molarized but 25% smaller than M1. The germ of P4 in its crypt was detected above dP4 inside the maxilla. The M1 is fully erupted and exhibits slight apical wear. The unworn M2 is almost completely erupted. Behind M2 the maxilla is broken off.
Immediately behind the dC, the maxilla is laterally constricted; the constriction is deepest at the dP2 position and becomes shallower at the level of dP3. There are four large maxillary foramina, two anterodorsal to dC and one dorsal to dP4 and of dP3. In the part of the maxilla containing dP2-dC, the maxillary bone is perforated by about ten tiny foramina.
MG 6729 [Gui Mam 116/77]
The right maxilla (Fig. 6) is from an adult individual. It holds the root of I6, roots and parts of the crown base of C, P4, and M1-5. This specimen is remarkable because P1 and P3 are completely reduced or lost and their alveoli are plugged by bone, leaving a gap between the distal canine root and P4. Anterolingual to the mesial canine root a pit for the lower canine is present in the maxillary bone (Fig. 6a). The P4 is almost unworn, M1 is moderately worn (lingual portion with cusps X and Y damaged), M2-4 exhibit an increasing wear gradient, and M5 exhibits moderate wear (distolabial portion broken).

Haldanodon exspectatus (MG 6729 [Gui Mam 116/77]). Right maxilla in occlusal (a), lateral (b-c), and medial (d) views; in c, the maxillary bone is transparent, showing roots of teeth. The I6 is broken and only its root is preserved. The root of i6 is highlighted in green, the canine in blue, the P4 in orange, and the molars in yellow. 3D models based on µCT-scan data. Scale bar equals 1 mm
Description of mandibles and lower teeth
MG 6827 [Gui Mam 36/80]
The right mandible (Fig. 7) is ontogenetically the youngest specimen available and its erupted dentition is comprised only by deciduous teeth. The di1 appears fully erupted and is unworn. It projects about 45° in anterior direction. It has a laterally slightly compressed root with an oval cross section. Its crown is spatulate with a pointed tip. A faint bulge separates the occlusal surface of di1 in two shallow concave lingual and labial portions. The interpretation of this tooth being the di1 is based on comparison with the i1 of adult mandible MG 6831 [Gui Mam 34/74] which is much larger (root diameter i1 of MG 6831 [Gui Mam 34/74] = 0.93 mm; di1 of MG 6827 [Gui Mam 36/80] = 0.41 mm) and has a labio-lingually strongly compressed root. Incisors di2 and di3 are lost due to breakage. Anterior to the dc sits the tiny di4, in its original position and separated by a small intact bony bar from it. It is inserted almost vertically in the mandible, only slightly inclined 10° in anterior direction. The root is rounded with a diameter of about 25% of that of i1. The crown is spatulate with a convex mesial side and slightly concave distal side (oriented linguo-distally); at the tip, the di4 bears an apical wear facet. Below the di4, the cap of the i4 was detected which has more than twice the diameter of the crown of the di4.

Haldanodon exspectatus (MG 6827 [Gui Mam 36/80]). Right mandible in occlusal (a), lateral (b-c), and medial (d) views; in c, the mandible is transparent, with caps of i4 and p1 and germ of p3 highlighted in green. 3D models based on µCT-scan data. Scale bar equals 1 mm
The dp1 is broken, and mesioventral to its preserved distal root the cap of p1 is visible. The caps of i4 and p3 are hidden in the mandible below their deciduous predecessors but have been detected by µCT analysis (Fig. 7c). The early ontogenetic age of MG 6827 [Gui Mam 36/80] is also evident from the cancellous bone surface of the mandible.
The dc is large with a mesio-distally wide triangular main cusp which bears a small distally oriented apical wear facet. The main cusp has a convex labial flank and a shallow concave lingual flank that is divided into a mesial and a distal portion by a ridge that is running down from its tip. Mesially and distally there are small cuspules near the crown base of which the distal one is slightly more prominent. The distal cusp sits on a heel and is slightly shifted labially; the heel is concave on the lingual and labial sides. The mesial and distal cuspules are connected by a shelf-like lingual cingulid. The labial flank is smooth without a cingulid, but the labial side of the heel bears some small enamel knots. The deciduous canine has two roots of which the ventral portion of the mesial root is broken off. The distal root is complete and stronger than the mesial one as evident from the preserved dorsal portion of the latter. According to the orientation of the distal root and the dorsal part of the mesial one, the roots were spread as is typical for deciduous teeth.
Of the dp1 only the ventral part of the distal root is preserved, the mesial root and the crown are broken off. Deep in the jaw the developing triangular crown of the unerupted p1 is visible. The crown of p1 has a convex labial side, a distally recurved tip and a small mesial cuspule at the base.
The dp2 is very small and single-rooted. Its crown consists of a triangular main cusp with a mesial and a distal small accessory cusp of equal size. The main cusp is conical and labio-lingually compressed; at the tip, it bears a small rounded apical wear facet. The labial flank is smooth, whereas on the lingual side a sharp cingulid connects the mesial and distal cuspules.
The dp3 is much larger and has 2.5 times the length of the dp2. The dp3 possesses two roots that diverge and on the labial side a wide interradical notch is evident. Its mesio-distally elongated crown with a large main cusp (= cusp a) and well-developed distolingual cusp (= cusp c) appears semi-molariform (for cusp terminology see Fig. 1). The conical cusp a is labio-lingually compressed and bears a well-developed apical wear facet that continues onto the distal crest. Cusp c has also a large apical wear facet. Mesiolabial cusp b and distolabial cusp d are of equal size and smaller than cusp c; apically they bear small wear facets. On the lingual side the crown bears a tubercular cingulid, whereas the labial side is smooth. Cusps g, df, and e are not developed. (Fig. 7b, d). The µCT images revealed a small mineralized germ of p3 between the roots of dp3 (Fig. 7c).
The dp4 is larger and has 1.5 times the length of dp3. It is molariform in shape, with main cusp a, large cusp c and well-developed labial cusps b and df. Lingual cuspules e and f are small. Cusp g is not developed. A strong crest connects the tip of cusp a to cusp c, and a much weaker crest is running from cusp a in disto-labial direction. Cusp a bears a well-developed apical wear facet that continues onto the connecting crest to cusp c. In disto-labial direction, cusp c is connected by a strong crest with cusp d; this crest is worn. A much weaker unworn crest connects cusp a with cusp b. Cusps b and d bear small apical wear facets. Tiny cuspule e is integrated into the lingual cingulid and is connected to cusp b by a faint s-shaped crest. The lingual cingulid is interrupted by cusp c; distal to cusp c it reappears under inclusion of cupule df and wraps around the distal crown base, connecting to cusp d. The interradical notch is wide, and the two roots are spread as is evident from the µCT image.
Posterior to dp4 there are two coal-filled alveoli of equal size for m1 which apparently was already erupted but has fallen out post mortem (Fig. 7d). In lingual aspect, below the origin of the coronoid process, the mandible reveals a collapsed large crypt for m2. The µCT analysis did not reveal a germ of m2 within the collapsed crypt suggesting that a mineralized structure was not yet developed.
Below and posterior to the collapsed crypt of m2, the wide postdentary trough is visible with insertion scars of the postdentary bones. A well-developed shelf with medial ridge is preserved above the postdentary trough. From the postdentary trough, the Meckel´s sulcus is extending towards the symphysis. Due to damage, the anterior end of Meckel´s sulcus is not discernible.
MG 6828 [Gui Mam 33/80]
The left mandible (Fig. 8) represents a slightly older individual than MG 6827 [Gui Mam 36/80] but the coronoid process is broken.

Haldanodon exspectatus (MG 6828 [Gui Mam 33/80]). Left mandible in occlusal (a), lateral (b-c), and medial (d-e) views; in c and e, the mandible is transparent, with caps of i2 and i4 highlighted in green, and large alveolus for di1. 3D models based on µCT-scan data. Scale bar equals 1 mm
The di1 is lost, but its large alveolus is preserved and indicates that it was procumbent (inclination 45°). The di2-4 are vertically oriented respectively slightly inclined distally (di4) and are unnaturally crowded which is due to distortion and post-mortem compression. The largest deciduous incisor is di3, second-largest di2 and the smallest is di4. All incisors are single-rooted and their crowns resemble morphologically those of MG 6827 [Gui Mam 36/80] and bear small apical wear facets. The µCT analysis revealed mineralized enamel caps of both i2 and i4 beneath their deciduous predecessors (Fig. 8c).
The dc corresponds in shape and size to that of MG 6827 [Gui Mam 36/80]. It bears a small, round, and distally oriented wear facet on the tip of the main cusp. Both roots are preserved and the distal root is stronger than the mesial one and recurved in posterior direction (Fig. 8d, e).
There are four premolar positions present. The crowns of both dp1 and dp2 are broken and of dp3 only the distal portion is preserved; dp4 is complete. The crown base of dp1 is preserved with the mesial root and the cervical portion of the distal root. A lingual portion of the dp2 root as well as the alveolus are preserved; as in MG 6827 [Gui Mam 36/80], dp2 has only a thin single root, whereas the other deciduous premolars are double-rooted. A crown fragment of the dp3 with its distal root and the mesial alveolus are present. The largest deciduous premolar is dp4 which resembles that of MG 6827 [Gui Mam 36/80] but is slightly more worn, suggesting that MG 6828 [Gui Mam 33/80] represents a slightly older individual.
The m1 is unworn except for tiny apical wear facets on cusps a, c, and b which indicates that it had just erupted. The crown closely resembles those of adult Haldanodon mandibles and is 25% larger than that of dp4. Besides its larger size, m1 differs from dp4 by presence of small cusps g and d. The small size of cusp g is typical for anterior (m1 and m2) molars in Haldanodon; it is largest in m3 but still only 50% of cusp c (e.g., MG 6832 [Gui Mam 6/82]). Immediately behind m1 and below the origin of the coronoid process, the crypt of m2 (open due to breakage) is visible.
The surface of the mandible exhibits a cancellous bone structure. A wide postdentary trough with insertions scars for the postdentary bones and a well-developed Meckel´s sulcus are visible on the medial side of the mandible. There are three mental foramina present, below di2/di3, di4, and dc/dp1.
MG 6829 [Gui Mam 73/77]
The left mandible fragment (Fig. 9) is broken at the anterior end and the distal root of dc is present. The first preserved tooth is unworn p1 which apparently was freshly erupted. It has a large, slightly recurved main cusp and a well-developed, shelf-like cingulid on the lingual side. The semimolariform dp3 and molariform dp4 are strongly worn (Fig. 9a) and have diverging roots (distal root of dp4 is broken). Below dp3, deep in the mandible, the mineralized cap of p3 (turned 90° in posterior direction) was detected by µCT. A high-oval resorption crypt (no mineralized germ of p4 detectable) is present below dp4 (Fig. 9b, c). Immediately posterior to dp4 sits the fully erupted m1, which exhibits only small apical wear facets on the main cusps. Posterior to m1, the alveoli for m2 are preserved. They are filled with coaly matrix which indicates that m2 was erupted and lost post-mortem before fossilization.

Haldanodon exspectatus (MG 6829 [Gui Mam 73/77]). Fragment of left mandible in occlusal (a), lateral (b-c), and medial views (d); in c. the mandible is transparent, with tilted cap of p3 highlighted in green. 3D models based on µCT-scan data. Scale bar equals 1 mm
MG 6794 [Gui Mam 115/74]
The lingual side of the left mandible (Fig. 10) is partially broken and exposes the alveoli and roots of the teeth. The first preserved tooth is dp3. Beneath the roots of dp3, slightly shifted lingually, the crown of the p3 is preserved. Mesial to dp3 are two alveoli, one of which is filled with coal matrix. The anterior-most alveolus is lingually open due to breakage and is empty. These two alveoli are interpreted as alveolus for the single-rooted dp2 and distal alveolus for the double-rooted dp1. Anterior to the distal alveolus for dp1 there is a large crypt deep in the mandible (open due to breakage of the lingual side) which probably housed the germ of p1. Below dp4, deep in the mandible a crypt with the germ of p4 was detected by µCT. Both preserved deciduous premolars are strongly worn.

Haldanodon exspectatus (MG 6794 [Gui Mam 115/74]). Left mandible in occlusal (a), lateral (b-c), and medial views (d); in c, the mandible is transparent, with unerupted crown of p3 and germ of p4 highlighted in green. 3D models based on µCT-scan data. Scale bar equals 1 mm
The m1 is about 20% smaller than m2 and exhibits moderate wear on cusps a and c as well as small apical wear facets on cusps b and g. The m2 is fully erupted and exhibits slight apical wear on the main cusps. The large empty crypt posterior to m2 suggests that m3 was in the process of eruption or close to it. The posterior portion of the mandible is missing.
MG 6786 [Gui Mam 28/77]
The crown of the unerupted p3 is visible because the lateral side of the left mandible (Fig. 11) is broken. The roots are not yet fully formed and it had not yet penetrated the mandible surface. The dp4 is still in place and well worn. Due to breakage of the lateral side of the mandible, its diverging roots are exposed. The ventral portion of the distal root of dp4 appears partially resorbed by the putative germ of p4; although the germ of p4 is lost, its resorption cavity is clearly detectable in the µCT image (Fig. 11c).

Haldanodon exspectatus (MG 6786 [Gui Mam 28/77]). Left mandible in occlusal (a), lateral (b-c), and medial (d) views; in c, the mandible is transparent, with unerupted crown of p3 highlighted in green, and resorption pit of p4 germ. 3D models based on µCT-scan data. Scale bar equals 1 mm
Both m1 and m2 are fully erupted and have tiny apical wear facets (slightly larger on m1). Posterior to m3 the lingual border of the mandible is broken and gives view to a large crypt for an unerupted m3 below the origin of the coronoid process. Posterior to the large crypt there is a tiny crypt for the germ of m4. The bone surface of the mandible is somewhat cancellous on the lateral side and the posterior part of the lingual side.
MG 6830 [Gui Mam 2/79]
The right mandible (Fig. 12) is broken into two pieces which have no direct contact surface. In the anterior fragment, i2 is the mesial-most preserved tooth with an unworn crown and cervical part of the root in place. The i2 is almost fully erupted and is vertically implanted. The i3 has a fully formed crown but was not yet erupted; the tip of its crown is exposed because the mandibular bone surface is damaged (Fig. 12b, d). The di3 is broken off. Of i4 only a small coal filled alveolus is present; it was lost post mortem. The dc is in place and resembles in shape and size the dc of MG 6827 [Gui Mam 36/80] and MG 6828 [Gui Mam 33/80]. Below dc the large enamel cap of the permanent canine was detected by µCT (Fig. 12c). It differs from the deciduous canine by its acute triangular shape and much larger size. Between dc and p1 there is a small gap of 0.4 mm length.

Haldanodon exspectatus (MG 6830) [Gui Mam 2/79]. Right mandible, preserved in two parts, in occlusal (a), lateral (b), and medial (c-d) views; in c, the mandible is transparent, with cap of c and unerupted crown of i3 highlighted in green. 3D models based on µCT-scan data. Scale bar equals 1 mm
The p1 is fully erupted and bears a tiny wear facet on the tip of the main cusp. In terms of morphology and size, it resembles p1 of the adult specimens MG 6804 [VJ 1001 − 155] (holotype), MG 6833 [Gui Mam 95/75], and MG 6796 [Gui Mam 122/76] (Luo and Martin 2007: figs. 7 and 8). On the lingual side of the crown-base it bears a shelf-like tubercular cingulid. The premolariform p3 follows immediately behind p1. The p3 is about 25% larger than the p1 and morphologically similar including the large, slightly recurved main cusp and a mesial and a distal accessory cuspule. Like p1, it bears a shelf-like tubercular cingulid on the lingual side of the crown base. The p3 is not yet fully erupted, with the lingual cingulid sitting on the rim of the mandibular bone (Fig. 12d). The molariform dp4 is still in place and is strongly worn on cusps a and c with the latter almost completely worn off. The distal root of dp4 is broken. Distal to dp4, the anterior fragment of the mandible is broken.
In the posterior fragment, the crown and mesial root of m1 are missing due to breakage. Based on the position of the preserved distal root, we believe that m1 was fully erupted. Posterior to the distal root of m1, a large crypt (open due to breakage) for the m2 is present. Behind the crypt of m2, below the origin of the coronoid process, a very small crypt for the germ of m3 is visible (Fig. 12d). Both crypts sit closely to the lingual side of the mandible and apparently were covered only by thin layers of bone which are now missing. A lingual position of the molar crypts has also been observed in the other mandibles examined.
The mandibular body exhibits a cancellous bone surface of the preserved part of the ventral side, in the wide postdentary trough and immediately above the medial ridge of the postdentary trough. The surface of the preserved part of the coronoid process is smooth. The Meckel´s sulcus is well-developed but broken off anterior to dp4. There are three mental foramina present on the labial side of the maxilla, below p1 (large), the anterior root of dc (large), and the posterior root of dc (small) (Fig. 12a).
MG 6782 [Gui Mam 42/74]
The right mandible (Fig. 13) is anteriorly broken, the first tooth preserved is p4. Anterior to p4, both alveoli of p3 (distal alveolus containing ventral portion of root) and the distal root of p1 are preserved. The p4 is in the process of eruption; the upper half of the main cusp is already emerged above the mandibular border.

Haldanodon exspectatus (MG 6782 [Gui Mam 42/74]). Right mandible in occlusal (a), lateral (b), and medial (c-d) views; in c, the mandible is transparent, with erupting p4 and unerupted crown of m4 highlighted in green. 3D models based on µCT-scan data. Scale bar equals 1 mm
The mesial two thirds of the crown of m1 are broken. Judging by the preserved mesial root and the distal portion of the crown base, it was fully erupted. The crown of the fully erupted m2 is intact; cusps a, b, c, and g bear small apical wear facets. The m3 is broken and lost. Judging by the alveoli with remaining root fragments it was fully erupted or in an advanced eruption stage. Behind m3, on the lingual side of the mandible the opened crypt with the fully developed crown of m4 (ready to erupt) is visible. Behind and slightly above the crypt of m4, a depression is present at the origin of the coronoid process (Fig. 13d) which is the crypt for the germ of m5.
The bony surface of the mandible is only slightly cancellous; the medial side of the coronoid process is smooth. The postdentary trough is wide and exhibits scars for the attachment of the splenial and other accessory bones. Meckel´s sulcus is well-developed and extends anteriorly up to the symphyseal region where it fades.
MG 6801 [VJ 1005 − 155]
The right mandible is preserved in two parts which are partially transferred to an artificial plastic matrix and which were virtually reassembled after µCT scanning (Fig. 14). The crown of p1 (split between both fragments), the dp3, germ of p3, dp4, m1, and the crown of m2 (split between both fragments) are preserved. Between crowns of p1 and dp3 is a small space for the tiny single-rooted dp2 which is not preserved. The crown of p1 is just piercing the mandibular surface. The dp3 is still in place and its crown is corroded. Below dp3, somewhat shifted towards the anterior root, the germ of p3 was detected. Below well-worn dp4 no mineralized germ of p4 was found. The m1 is fully erupted and almost unworn. The m2 is in the process of eruption and its main cusp a has just emerged above the bony surface of the mandible.

Haldanodon exspectatus (MG 6801 [VJ 1005 − 155]). Right mandible in occlusal (a), lateral (b-c), and medial (d) views; in c, the mandible is transparent, with unerupted crowns of p1 and m2 and germ of p3 highlighted in green. 3D models based on µCT-scan data. Scale bar equals 1 mm
Krusat (1980: Fig. 23) interpreted the teeth preserved in MG 6801 [VJ 1005 − 155] as p1, dp2, dp3, m1, and m2 which is here emended based on µCT analysis as outlined above.
MG 6800 [VJ 1004 − 155]
The right mandible (Fig. 15) is preserved in two parts of which the tooth-bearing anterior portion is embedded in an artificial matrix. Both fragments have been virtually repositioned. The anterior-most preserved tooth is the fragmentary dc; below the distal crown portion and root of dc is the cap of the canine (Fig. 15b, d). The p1 was lost post mortem but its two alveoli are present. The crown of p3 is unerupted and just about to pierce the dentary surface. A moderately worn dp4 is still in place without a germ of p4 below. Both m1 and m2 are fully erupted; m1 is slightly worn and m2 is almost unworn. The m3 has fallen out post-mortally and is represented by its two alveoli. The large crypt of m4 is lingually broken and empty, and its shape demonstrates that both roots were already developed. On the slope of the ascending ramus sits a small crypt with a tiny enamel cap of m5. The mandible has a well-developed postdentary trough. The anterior extension of Meckel´s sulcus is unclear because the anterior mandibular body is somewhat distorted and partially preserved as a plastic cast of the natural mould, therefore we are unsure of its original position.

Haldanodon exspectatus (MG 6800 [VJ 1004 − 155]). Right mandible in occlusal (a), lateral (b), and medial (c-d) views; in d, the mandible is transparent, with unerupted cap of c, unerupted crown of p3 and germ of m5 in crypt highlighted in green. 3D models based on µCT-scan data. Scale bar equals 1 mm
Krusat (1980: Fig. 24) interpreted the teeth preserved in MG 6800 [VJ 1004 − 155] as root of p1, p2, dp3, m1, and m2 which is here emended based on the μCT analysis.
MG 6830 [Gui Mam 2/79]
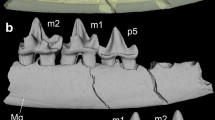
The adult left mandible fragment (Fig. 16) is the only specimen from the entire Guimarota sample that preserves the complete tooth row with i1-4, p1, p3, p4, and m1-6 in place. The i1 (tip broken) is labio-lingually strongly compressed and is procumbent (inclination 20°). The more posterior incisors are increasingly more steeply inclined (i2: 45°; i3: 65°; i4: 85°) and the size gradient from largest to smallest is i1 > i3 > i2 > i4. All incisors are single-rooted, and the roots of i2-4 are slightly compressed labio-lingually; their crowns bear small apical wear facets. The double-rooted canine is the tallest tooth and bears a large apical wear facet but is otherwise unworn. Premolars p1, p3, and p4 are all double-rooted and moderately worn, and increase in size from front to back (p1 < p3 < p4). The molars are double-rooted (m1-5) and strongly worn (especially m3 and m4). In m4 and m5 the roots are fused on the labial side and in m6 the roots are fully fused with vertical grooves on the lingual and labial side. The m6 is the smallest molar with a simplified crown morphology, a typical character of the ultimate molar which has the tendency to become vestigial.

Haldanodon exspectatus (MG 6831 [Gui Mam 34/74)). Left mandible in occlusal (a), lateral (b), and medial (d) views; c. complete permanent tooth row in labial view. The i1 is weakly inclined and procumbent (tip broken), i2-4 are increasingly more steeply inserted. The molars are strongly worn (except for small m6). The incisors are highlighted in green, the canine in blue, the premolars in orange, and the molars in yellow. 3D models based on µCT-scan data. Scale bar equals 5 mm
Sequence of tooth replacement in the maxilla
Deciduous upper incisors are not preserved in any of the studied specimens (Fig. 17; Table 2). In MG 6834 [Gui Mam 44/81] dC and dP1-4 are still in place when M1 is fully erupted. In MG 6732 [Gui Mam 12/74], with fully erupted M1-2 and nearly fully erupted M3, dC is still in place and deep in the maxilla a small enamel cap of the permanent C is present. The dP1-4 are still in place, with crowns of both P1 and P3 just about to break through the mandibular surface. Above dP4 a crypt with tiny mineralized germ of P4 is present. None of the maxillae preserved a germ or crypt of a permanent successor above dP2 which suggests that the second deciduous premolar position (dP2) is lost in the permanent dentition, as for the mandible. The dP2 can have a single root (MG 6834 [Gui Mam 44/81]), incompletely separated roots (MG 6731 [Gui Mam 12/74]), or two roots (MG 6742 [Gui Mam 91/76]), whereas the slightly smaller dP1 is single-rooted in all studied specimens. In MG 6757 [Gui Mam 128/76] the crown of the permanent C has lifted the dC slightly out of its alveoli and is ready to erupt, when at least M1 and M2 are in place. Because M1 and M2 (both with slight apical wear) are preserved on a separate maxilla fragment without direct contact to the piece containing dC and C, possibly one or two more molars were erupted or were in the process of eruption in MG 6757 [Gui Mam 128/76].

Haldanodon exspectatus. Studied juvenile maxillae from the Guimarota coal mine in ontogenetic order from top to bottom. Shaded pencil drawings in medial view. The dashed lines indicate the position of the deciduous canine. Scale bar equals 2 mm
Sequence of tooth replacement in the mandible
Haldanodon exspectatus has four lower permanent incisors (Martin and Nowotny 2000) (Fig. 15; Table 3). In most of the juvenile mandibles the anterior portion of the jaw with the incisors in place is damaged, but in MG 6827 [Gui Mam 36/80] and MG 6828 [Gui Mam 33/80] it is largely intact. In MG 6828 [Gui Mam 33/80], di2, di3, and di4 are present (anterior-most incisor position damaged). In MG 6827 [Gui Mam 36/80], di1 and di4 are still in place, both i2 and i3 and their roots/alveoli are lost due to breakage. The deciduous incisors can be distinguished from the permanent incisors by their much smaller size (44% of the di1 root diameter). The combination of information from juvenile mandibles (MG 6827 [Gui Mam 36/80], MG 6828 [Gui Mam 33/80], and MG 6830 [Gui Mam 6830]) suggests an alternating incisor replacement, with early replacement of the i2 and i4 positions and late replacement of the i1 and i3 positions. The space between di1 and di4 in MG 6827 [Gui Mam 36/80] appears wide enough to have accommodated di2 and di3.
The dc is replaced relatively late in ontogeny. In MG 6830 [Gui Mam 2/79], a mineralized enamel cap (without roots) of the permanent canine is below dc; dp1-3 are already shed and p1 and p3 are erupted, but dp4 is still in place and lacks a germ of p4 below. This suggests that the deciduous canine is replaced after dp3 and simultaneously with or shortly before dp4.
Haldanodon has three permanent lower premolar positions but four deciduous premolars, as is evident from MG 6827 [Gui Mam 36/80] and MG 6828 [Gui Mam 33/80] (Figs. 7 and 8). The dp2 is single-rooted and the smallest deciduous premolar; all other deciduous lower premolars are double-rooted. In MG 6827 [Gui Mam 36/80] the cap of p1 and germ of p3 were detected below both dp1 and dp3, but no successor is present below dp2. Both dp3 (semimolariform) and dp4 (molariform) resemble the molars in shape, but are smaller and lack cusp g. The dp3 has half the size of m1 and lacks both cuspules df and e; dp4 is more molarized with tiny cuspules df and e (for cusp terminology, see Fig. 1), and is 25% smaller than m1. Both dp3 and dp4 are replaced by premolariform permanent successors. The suppression of the second premolar position in the permanent dentition suggests that the lower premolar formula in Haldanodon is p1, p3, p4. The premolars are replaced sequentially from front to back. In the ontogenetically youngest mandible MG 6827 [Gui Mam 36/80] all four deciduous premolars are present. The dp1 is broken but its distal root still sits in its alveolus with a cap of p1 visible deep in the mandible. Below dp3 is a tiny germ of p3, clearly in a much earlier stage than the cap of p1. Below dp4 no trace of a mineralized germ is seen in the µCT scan. In the mandible MG 6830 [Gui Mam 2/79], p1 is fully erupted, p3 is not yet fully erupted and unworn, whereas a strongly worn dp4 is still in place. In MG 6794 [Gui Mam 115/74], both dp3 and dp4 are in place with the crown of p3 and crypt with germ of p4 inside the mandible; m1-2 are fully erupted and m3 (lost) was still in crypt. In MG 6782 [Gui Mam 42/74], p4 is erupting, with the upper third of the main cusp above the mandibular border; m1-2 are erupted, m3 (broken) was in the process of eruption or fully erupted, and fully developed crown of m4 is seen in its crypt at the base of the ascending ramus. This suggests that the p4 erupts shortly before m4.
The m1 erupts before the deciduous premolars are replaced as is evident from MG 6828 [Gui Mam 33/80]. The m2 is ready to erupt at the time when p1 is present and p3 is in a late eruption stage (MG 6830 [Gui Mam 2/79]). The m3 is in the process of eruption and m4 is fully developed but has not yet started to erupt when p4 is in the process of eruption. Antemolar replacement is completed when four molars are in place (Table 3). Successively, one or two (m5-6) molar positions are added during later ontogeny. The mandible has determinate growth, reaching its final (adult) length of about 30 mm when five molars are erupted. The molars erupt at the origin (slope) of the ascending ramus which successively is shifting backwards, which has also been observed in Docodon victor (Schultz et al. 2019). In MG 6782 [Gui Mam 42/74], the still shallow crypt of m5 is evident on the medial side of the mandible, posterodorsal to m4 which is still sitting in its large crypt, ready to erupt. The erupting molars and the molar germs are forming close to the lingual surface of the mandible and are covered by a thin bony layer only. Small (vestigial) m6 erupts somewhat shifted lingually immediately at the base of the ascending ramus, filling the space posteriorly of m5. Based on combined information from the mandibles described above, the tooth eruption sequence is reconstructed as m1 → p1 + m2 → i2 + i4 → ?i1 + ?i3 → p3 + m3 → c + p4 + m4 → m5 → m6.
Ontogenetic changes of the mandible
During ontogeny, the Haldanodon mandible experiences considerable allometric changes. There is an elongation of the horizontal ramus and a backwards shift of the coronoid process during ontogeny (Fig. 18). These are weaker than in Docodon, however, because Haldanodon possesses only six lower molars, whereas Docodon has eight (Simpson 1929; Schultz et al. 2019). An elongation occurs also at the anterior end of the dentary, because the replacing permanent canine is about 50% larger than its deciduous predecessor. The anterior-most incisor (di1) is 45° inclined anteriorly (procumbent) in the ontogenetically youngest specimen MG 6827 [Gui Mam 36/80]. The di4 of the same specimen is implanted much steeper, at 80°. The vertical to distally inclined orientation of di2, di3, and di4 in the slightly older individual MG 6828 [Gui Mam 33/80] is caused by post-mortal compression. A similar phenomenon of diagenetic compression, resulting in unnatural steep inclination of the incisors, occurred in the adult mandible MG 6769 [Gui Mam 35/75] (Martin 2018: Fig. 6.12B). Mandible MG 6831 [Gui Mam 34/74] shows the undistorted orientation of the incisors with a procumbent i1 and increasingly more steeply inclined i2-4 (Fig. 16).

Haldanodon exspectatus. Mandibular growth and dental replacement pattern in the studied mandibles from the Guimarota coal mine. Specimens MG 6828, 6829, 6794, and 6786 mirrored for easier comparison. Shaded pencil drawings of medial aspect. Dashed lines indicate the position of the dp4/p4. Scale bar equals 2 mm
In the juvenile specimens, the Meckel´s sulcus extends on the medial side of the mandible from the postdentary trough to the symphysis in anterior direction (e.g., MG 6827 [Gui Mam 36/80] and MG 6828 [Gui Mam 33/80]). In adult specimens, it fades before reaching the symphysis (Martin 2018: fig. 6.12) which is related to the anterior elongation of the mandible caused by the larger permanent canine and the larger permanent incisors. The same observation was made by Schultz et al. (2019) on the mandible of Docodon.
The bone structure of most juvenile specimens is cancellous with longitudinally oriented canaliculi which are most expressed on the lateral side of the posterior end of the mandible, the anterior medial side, and within the postdentary trough. The postdentary trough is relatively larger in the juveniles than in the adults. There are three mental foramina present which show some variation in position, largely independent from ontogenetic age. In the youngest specimen (MG 6828 [Gui Mam 33/80]), the mental foramina are below di2, di4, and between dc and dp1 positions. In the somewhat older specimen MG 6830 [Gui Mam 2/79], the mental foramina are below dc, immediately behind dc, and below p1. In two adult specimens, the positions of the mental foramina vary. They lie below i2, c, and between c and p1 in MG 6769 [Gui Mam 35/75], and below i4, anterior root of c, and p1 in MG 6777 [Gui Mam 81/79] (Martin and Nowotny 2000: fig. 14.4).
Discussion
Mode of tooth replacement
The analysis of the mandibles and maxillae available for study corroborated that Haldanodon exspectatus has a diphyodont replacement of the antemolar dentition, a derived mammalian character. This is in accordance with the observation by Schultz et al. (2019) who suggested a diphyodont premolar replacement for Docodon victor. Multiple replacements of the upper canine as suggested earlier for Haldanodon (Martin and Nowotny 2000; Nowotny et al. 2001; Martin et al. 2010) were not endorsed. Replacement of molars does not occur in Haldanodon, or in Docodon victor Schultz et al. (2019). Haldanodon, Docodon and the other docodontans so far known differ from Sinoconodon where one replacement of the posterior “molars” (M3-5) did occur, and the anterior molars (M1-2) were lost during ontogeny (Zhang et al. 1998; Luo et al. 2004: fig. 2). Sinoconodon shows a single replacement of premolars (Zhang et al. 1998, Luo et al. 2004), as seen in Haldanodon and Docodon, which is a derived feature of crown mammals. Similar to Sinoconodon, the lower incisors (information on uppers is lacking) of Haldanodon are replaced in alternating mode. The upper and lower canines of Haldanodon show a single replacement, whereas for Sinoconodon multiple replacements for the upper canine have been reported (Crompton and Luo 1993; Zhang et al. 1998; Luo et al. 2004). Sinoconodon shows a loss of anterior postcanines (premolars and anterior molars) during ontogeny (Zhang et al. 1998). The two deciduous premolar positions are replaced once and then are lost, as are the two anterior molar positions (M1-2/m1-2) which results in an increasing larger diastema and a posterior shift of the functional tooth row in the jaw by successive addition of newly erupting molariforms; this is plesiomorphic for mammals and is seen also in mammaliamorph synapsids (Zhang et al. 1998). A loss of anterior postcanines occurs, less strongly expressed, in other non-mammalian mammaliaforms such as Morganucodon, Hadrocodium, and possibly Kuehneotherium (Kermack et al. 1968; Parrington 1971; Mills 1971, 1984; Luo et al. 2001, 2004). Both the non-replacement and loss of the P2/p2 position in Haldanodon (“P2” diastema; Butler 1939; Rougier et al. 2015) correspond to these observations. Simpson (1929: p. 89) observed a lack of the p2 in the mandible of Ennacodon crassus” (synonymized with Docodon victor by Schultz et al. 2019) and attributed this feature to its young ontogenetic age. According to our observations, it is more likely that this is due to the fact that p2 is lost, as evident from Haldanodon. In the adult Haldanodon maxilla MG 6729 [Gui Mam 116/77] premolar reduction is even more advanced as P1 and P3 are lost and their alveoli are plugged by bone, whereas P4 is in place (Fig. 16). In Sinoconodon, small deciduous postcanines are replaced by larger and more molariform successors, which resembles the pattern seen in diademodontids (Luo et al. 2004). Haldanodon differs from that pattern by showing molarized dP3-4 and dp3-4 which are replaced by premolariform permanent premolars with morphologically simpler crowns. In the maxilla, the small and peglike dP1 is replaced by a somewhat larger premolariform successor whereas the dP2 position is lost. Premolar replacement in the mandible corresponds to this pattern: dp1-2 are small and peglike, and dp1 is replaced by a somewhat larger premolariform successor, whereas the dp2 position is lost. This corresponds to the pattern seen in Sinoconodon but differs from that of crown mammals where the replacing premolars generally are morphologically less complex than their predecessors. An exception among crown mammals is Gobiconodon where the successor tooth is similar to its deciduous predecessor in the complexity of molariform shape (Luo et al. 2004).
In contrast to Sinoconodon (Crompton and Sun 1985), the complex molars of Haldanodon (and of other docodontans) show a precise occlusal fit with well-defined and precisely fitting wear facets (Jenkins 1969; Gingerich 1973; Brinkkötter 2018; Schultz et al. 2019; Martin et al. 2020). This precise occlusal fit and mastication coincide with the non-replacement of molars in Haldanodon (and other docodontans). After replacement of the incisors, canine, and premolars which was completed around the time when m4 was erupting (Table 3), one or two supplementary (m5-6) molars were added to the tooth row, coinciding with an elongation of the posterior portion of the mandible. The number of lower molars in Haldanodon appears reduced compared to Docodon victor, where up to eight molar positions are present (Simpson 1929; Schultz et al. 2019). The non-replacement of molars and their limited number (5–6, with m6 small and often vestigial) suggest that Haldanodon had a determinate growth of its skull, although the lack of epiphyses in the humeri and femora indicate an extremely extended or even lifelong growth of the postcranium (Martin 2005). Luo et al. (2004) suggested for Sinoconodon an indeterminate growth of the skull, together with continual tooth replacement as in non-mammalian cynodonts (Crompton 1963; Hopson 1971; Osborn and Crompton 1973; Osborn 1974).
A molariform (“molar”) replacement has been suggested for morganucodontans (Luo et al. 2004), but there is no direct evidence so far. In Megazostrodon the replacement of m2 has been postulated based on less wear in this tooth as compared to m1 and m3 (Gow 1986), but no successor has been detected. It is generally accepted that there was no replacement of molars in Morganucodon watsoni (Mills 1971; Crompton 1972; Parrington 1973; Crompton and Parker 1978; Clemens and Lillegraven 1986; Luo 1994). According to Kielan-Jaworowska et al. (2004: p. 152), Morganucodon had developed a diphyodont replacement pattern in all of its antemolars, but molars were not replaced. The mode of premolar replacement is sequential from front to back, similar to Sinoconodon (Luo et al. 2004). Haldanodon and Docodon correspond to this replacement pattern. According to Luo et al. (2004), there is little evidence for replacement of ultimate molars from the growth series of complete mandibles of Morganucodon oehleri (Yang 1982; Crompton and Luo 1993) and Dinnetherium (Jenkins et al. 1983; Crompton and Luo 1993). According to Z.-X. Luo (personal communication February 7, 2022) the statement that dental replacement occurred in the anterior molars of Morganucodon (Luo et al. 2004: p. 165) is no longer considered to be valid. An X-ray of two Morganucodon oehleri skulls provided no evidence for replacement successors to the ultimate and penultimate functional molars (Luo et al. 2004). The non-replacement of molars as seen in Haldanodon and Docodon was apparently established in morganucodontans. O´Meara and Asher (2016) found evidence for determinate growth in a large sample of Morganucodon mandibles, supporting the hypothesis that determinate growth enabled diphyodont antemolar replacement and non-replacement of molars with precise occlusion (Crompton and Jenkins 1973; Crompton 1995).
Position of the canine
Haldanodon possesses six upper incisors, of which I5 and I6 are implanted in the maxilla (Lillegraven and Krusat 1991; Martin and Nowotny 2000). In therian mammals, according to the criterion of Lage [position] (Remane 1952) the first tooth implanted in the maxilla is considered to be the canine. However, the occurrence of maxillary incisors is not uncommon in mammaliaforms and stem therians (Kermack et al. 1981: p. 11; Lillegraven and Krusat 1991; Martin 1997, 1999) and probably represents the plesiomorphic mammalian condition. A maxillary incisor (I4) is also found in the docodontan Borealestes cuillinensis (Panciroli et al. 2021: fig. 15). As a general rule, the lower canine occludes anterior to the upper canine in mammals, and therefore the occlusal relationships can be used for determining the respective tooth category. Haldanodon maxillae exhibit a large pit mesiolingual of the upper caniniform tooth (Fig. 6a) for accommodation of the occluding lower caniniform tooth (a feature also seen in B. cuillinensis; Panciroli et al. 2021). This is evidence that the upper and lower caniniform teeth of Haldanodon are indeed canines, although the upper is not the anterior-most tooth in the maxilla. The lower deciduous premolars are sequentially shed and replaced (dp2 is not replaced) from front to back (dp1 → dp2 → dp3 → dp4, and the deciduous lower canine is replaced about at the same time as dp4. This is evident from MG 6830 [Gui Mam 2/79] where dc is still in place when p1 and p3 are already replaced and dp4 is still present. The large cap of the permanent canine below dc of MG 6830 [Gui Mam 2/79] suggests that deciduous canine replacement occurred simultaneously with or shortly before replacement of dp4. The late replacement of the lower canine which does not follow the sequential replacement of the lower premolars supports the interpretation that it belongs to a different tooth category than the premolars. Because of the occlusal relationship of upper and lower caniniform tooth, we consider the upper caniniform tooth as the canine, even if it is not the anterior-most tooth implanted in the maxilla. A late replacement of the deciduous canine is also observed in Docodon. Schultz et al. (2019) reported the presence of deciduous canines in juvenile mandibles YPM VPPU 10649 and YPM VPPU 13735. According to our observations, the deciduous premolars are already replaced by their permanent successors in these mandibles, contra Schultz et al. (2019) who interpreted the third and fourth premolar position as still having the deciduous premolars in place. The interpretation by Schultz et al. (2019) was based on the shape of the roots, but no germs of permanent premolars had been detected below the teeth in question. The crown morphology of the supposed dp4 in YPM VPPU 13735 is premolariform and clearly differs from molariform dp4 of YPM VPPU 23748. The supposed dp3 of YPM VPPU 10649 is also premolariform, and both teeth closely resemble p3 and p4 of the adult mandibles YPM VPPU 11826 and YPM VPPU 11823 (Schultz et al. 2019: fig. 7), and are interpreted as permanent premolars here.
“Peraiocynodon” and Cyrtlatherium
“Peraiocynodon inexpectatus” was established by Simpson (1928) based on a left ramus fragment with four teeth (NHMUK PV OR48248, formerly BMNH 48248) which is the only known specimen for this taxon. Simpson (1928) interpreted the teeth as m1-4 and germ of m5. Later Butler (1939) reinterpreted the teeth in the mandibular fragment as deciduous premolars (dp1-4). When Butler (1939: p. 332) compared the dp4 of “Peraiocynodon” (“m4” of Simpson 1928) with the m1 of Docodon, he noted clear differences: “…the paraconid [= cusp b] is lower and close to the anterior cusp; the metaconid [= cusp c] and the adjacent cusps are less distinct; the outline is narrower relatively to its breadth; the protoconid [= cusp a] is placed more nearly in the centre of the crown… The other teeth are progressively simpler, and the most anterior one bears the protoconid [= cusp a] and a marginal ledge. It therefore seems very probable that they are the milk molars of a form related to Docodon” (additions in [ ] by the present authors). The characters listed by Butler (1939) are exactly the same that we have observed in dp4 of Haldanodon. Sigogneau-Russell (2003) described isolated lower docodontan molariform teeth from the Bathonian of Kirtlington that she attributed to a new species of “Peraiocynodon” (“P. major”). The teeth assigned to “Peraiocynodon major” are not only much larger (170%) than those of “Peraiocynodon inexpectatus”, but also differ strongly by morphology. Furthermore, “Peraiocynodon major” is of Middle Jurassic (Bathonian) age, whereas “Peraiocynodon inexpectatus” comes from the Lower Cretaceous (Purbeckian), and is approximately 25 million years younger. When describing the two molariform teeth from Kirtlington, Sigogneau-Russell (2003: p. 361) noted: “The two teeth [holotype NHMUK PV M46693 and specimen M46679] here referred to as Peraiocynodon major sp. nov. (Fig. 3d) have a very unusual configuration in that they possess no mesio-lingual cusp; in that the mesio-labial cusp does not send a crest lingually (= antero-basal crest)…” The absence of a mesiolabial cusp (= cusp g) is a character seen in dp3 and dp4 of Haldanodon, which suggests that the teeth attributed to “Peraiocynodon major” could be docodontan posterior deciduous premolars (dp3, dp4). In any case, by morphology and stratigraphic age difference, it is highly improbable that the Kirtlington teeth (attributed to “Peraiocynodon major” by Sigogneau-Russell 2003) belong to the same genus as the holotype mandible of “Peraiocynodon inexpectatus”. The perceived superficial similarities between “Pereiocynodon inexpectatus” and “Peraiocynodon maior” may be explained by them both being deciduous premolars. Sigogneau-Russell (2003) described eight additional lower teeth and tooth fragments from the British Purbeck and attributed them to “Peraiocynodon inexpectatus”; six of them exhibit possible deciduous premolar characters such as more elongated trigonid, lower cusp b, and absence of cusp g. Two of the eight teeth (fragmentary) are larger than the others. According to Sigogneau-Russell (2003: p. 370) “…it seems unlikely that we would have here six deciduous teeth [the smaller teeth] for two permanent molars [the larger fragmentary teeth] of one single taxon. Given that argument … I would interpret the six teeth as permanent molars of Peraiocynodon and the two fragments [of the larger teeth] as the remains of a Docodon species…” (additions in [ ] by the present authors). Averianov (2004) confirmed Butler´s (1939) interpretation of the “Peraiocynodon inexpectatus” holotype dentition as dp1-4. He suggested that the absence of cusp g in the teeth of the two species of “Peraiocynodon” (and in Cyrtlatherium) indicates that these are milk teeth. According to Averianov (2004), Sigogneau-Russell´s (2003) argument against interpreting six out of the eight lower teeth as deciduous premolars does not apply, because the sample size is too small, and in some environments and localities deciduous teeth may be quite abundant (see Cifelli 1999). Schultz et al. (2019: p. 31) observed major differences in the deciduous teeth (premolars) of “Peraiocynodon inexpectatus” and Docodon based on µCT study. However, the dp4 of mandible YPM VPPU 23748 (Schultz et al. 2019: fig. 7a) is very similar to dp4 of the “Peraiocynodon inexpectatus” holotype mandible NHMUK PV OR48248. YPM VPPU 23748 is the only specimen studied by Schultz et al. (2019) where an unerupted germ of a premolar (p3) has been detected. The dp4 of YPM VPPU 23748 exhibits the same characteristics as dp4 of Haldanodon and “Peraiocynodon inexpectatus” (e.g., absence of cusp g) and is without doubt a deciduous premolar. The teeth in two other anterior mandibular fragments (YPM VPPU 10649 and YPM VPPU 13735), which were interpreted as dp3 and dp4 by Schultz et al. (2019), differ in crown morphology from the dp4 of YPM VPPU 23748. Their assignment as deciduous premolars is based on the shape of the roots, but no germs of permanent p3 and p4 have been detected (Schultz et al. 2019: Fig. 7). The crowns of the premolars in question of YPM VPPU 10649 and YPM VPPU 13735 are partly broken; the crowns of the preserved teeth (p3 of YPM VPPU 10649 and p4 of YPM VPPU 13735) differ clearly from the dp3 and dp4 of Haldanodon (and from dp4 of YPM VPPU 23748) which are all molarized. We consider these teeth as permanent premolars, and the observed differences to “Peraiocynodon” account for their permanent premolar nature. Based on our observations of the deciduous premolars of Haldanodon, we concur with Averianov (2004) that “Peraiocynodon inexpectatus” is an objective junior synonym of Docodon, based on milk teeth.
Cyrtlatherium canei had been erected based on two right lower molars by Freeman (1979) who considered it as a “symmetrodontan” kuehneotheriid. Sigogneau-Russell (2001) recognized the docodontan nature of Cyrtlatherium and suggested that this taxon is based on two milk teeth. Averianov (2004) corroborated the deciduous nature of the Cyrtlatherium premolars. He suggested that they are milk teeth of Simpsonodon oxfordensis, which would make Simpsonodon oxfordensis Kermack et al. 1987 a junior synonym of Cyrtlatherium canei. This suggestion is probable, but currently cannot be proven with certainty on the limited material base. Even if Simpsonodon will turn out to be a junior synonym of Cyrtlatherium, it appears reasonable to maintain Simpsonodon, because this name is widely introduced into the literature and Cyrtlatherium is rarely used, and a formal case needs to be placed at the International Commission of Zoological Nomenclature if the synonymy is corroborated. Based on our observations of the deciduous dentition of Haldanodon exspectatus, we confirm that “Peraiocynodon inexpectatus” and Cyrtlatherium canei are based on milk teeth as suggested by Butler (1939) and Averianov (2004) for the former and by Sigogneau-Russell (2001) and Averianov (2004) for the latter.
Conclusion
Haldanodon exspectatus exhibits the diphyodont replacement of the antemolar dentition as seen in crown mammals. The non-replacement of molars is probably linked to the complex molar crown pattern with precise occlusion, because the chewing apparatus would have lost its functionality with replacing molars. The mandible exhibits determinate growth which is attained when m5 is erupted; subsequently vestigial m6 fills the space between m5 and base of the ascending ramus. The sequential replacement of premolars from front to back is a derived mammalian character, the alternating replacement of the incisors is plesiomorphic and seen on the non-mammalian synapsid stem lineage (Parrington 1936; Osborn and Crompton 1973). The upper canine is not the anterior-most tooth implanted in the maxilla (small peg-like maxillary incisors i5 and i6 are present), but occlusal relationship with the lower caniniform demonstrates its canine nature. During ontogeny, the Haldanodon mandible experiences considerable changes in shape with growth in the anterior region for accommodation of the larger permanent incisors and canine and additional growth at the origin of the ascending ramus for accommodation of m5-6. The adult dental formula of Haldanodon is 6I/4i, 1C/1c, 3P/3p, 5 M/5-6m, and that of the deciduous dentition ?dI/4di, 1dC/1dc, 4dP/4dp. Upper and lower second premolar positions are present in the deciduous dentition, but are not replaced after shedding (lost in the permanent dentition). The lower permanent tooth eruption sequence of Haldanodon is m1 → p1 + m2 → i2 + i4 → ?i1 + ?i3 → p3 + m3 → c + p4 + m4 → m5 → m6. The series of juvenile Haldanodon mandibles corroborates the earlier suggestions (Butler 1939; Averianov 2004) that “Peraiocynodon” and Cyrtlatherium are likely to be based on docodontan milk dentitions.
Data availability
Specimens are deposited in the cited collections and institutions. Images of segmentation of all specimens scanned by µCT for this study are presented in the text figures. The scan data and 3D meshes are available from the corresponding author upon reasonable request.
Change history
04 September 2023
A Correction to this paper has been published: https://doi.org/10.1007/s10914-023-09684-2
References
Averianov AO (2004) Interpretation of the Early Cretaceous mammal Peraiocynodon (Docodonta) and taxonomy of some British Mesozoic docodonts. Russ J Theriol 3:1–4
Brinkkötter JJ (2018) Molar dentition of the docodontan Haldanodon (Mammaliaformes) as functional analog to tribosphenic teeth. Dissertation, Universität Bonn. https://hss.ulb.uni-bonn.de/2019/5487/5487.htm
Butler PM (1939) The teeth of the Jurassic mammals. Proc Zool Soc Lond 109:329–356
Butler PM (1988) Docodont molars as tribosphenic analogues (Mammalia, Jurassic). In: Russell DE, Santoro J-P, Sigogneau-Russell D (eds) Teeth Revisited: Proceedings of the VIIth International Symposium on Dental Morphology. Mém Mus nat Hist nat Paris C 53:329–340
Butler PM (1997) An alternative hypothesis on the origin of docodont molar teeth. J Vertebr Paleont 17:435–439
Cifelli RL (1999) Therian teeth of unusual design from the medial Cretaceous (Albian-Cenomanian) Cedar Mountain Formation, Utah. J Mamm Evol 6:247–270
Clemens WA, Lillegraven JA (1986) New Late Cretaceous, North American advanced therian mammals that fit neither the marsupial nor eutherian molds. Contrib Geol Univ Wyo Spec Pap 3:55–85
Crompton AW (1963) Tooth replacement in the cynodont Thrianxodon liorhinus Seeley. Ann South Afr Mus 46:279–521
Crompton AW (1972) Postcanine occlusion in cynodonts and tritylodonts. Bull Brit Mus (Nat Hist) Geol 21:30–71
Crompton AW (1995) Masticatory function in nonmammalian cynodonts and early mammals. In Thomason JJ (ed) Functional Morphology in Vertebrate Paleontology. Cambridge University Press, Cambridge, pp 55–75
Crompton AW, Jenkins FA (1973) Mammals from reptiles: a review of mammalian origins. Annu Rev Earth Pl Sci 1:131–155.
Crompton AW, Luo Z-X (1993) The relationships of the Liassic mammals Sinoconodon, Morganucodon oehleri and Dinnetherium. In: Szalay FS, Novacek MJ, McKenna MC (eds) Mammal Phylogeny. Vol.1. Mesozoic Differentiation, Multituberculates, Monotremes, Early Therians, and Marsupials. Springer-Verlag, New York, pp 30–44
Crompton AW, Parker P (1978) Evolution of the mammalian masticatory apparatus. Am Sci 66:192–201
Crompton AW, Sun AL (1985) Cranial structure and relationships of the Liassic mammal Sinoconodon. Zool J Linn Soc 85:99–119
Drescher E (2000) Preparation of vertebrate fossils from the Guimarota mine. In: Martin T, Krebs B (eds) Guimarota – A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, Munich, pp 137–142
Freeman EF (1979) A Middle Jurassic mammal bed from Oxfordshire. Palaeontology 22:135–166
Gingerich PD (1973) Molar occlusion and function in the Jurassic mammal Docodon. J Mammal 254:1008–1013
Gow CE (1986) A new skull of Megazostrodon (Mammalia: Triconodonta) from the Elliot Formation (Lower Jurassic) of southern Africa. Palaeontol Afr 26:13–23
Hopson JA (1971) Postcanine replacement in the gomphodont cynodont Diademodon. In Kermack DM, Kermack KA (eds) Early Mammals. Academic Press, London, pp 1–21
Jenkins FA Jr (1969) Occlusion in Docodon (Mammalia, Docodonta). Postilla 139:1–24
Jenkins FA Jr, Crompton AW, Downs WR (1983) Mesozoic mammals from Arizona: new evidence on mammalian evolution. Science 222:1233–1235
Ji Q, Luo Z-X, Yuan C-X, Tabrum AR (2006) A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals. Science 311:1123–1127
Kermack DM, Kermack KA, Mussett F (1968) The Welsh pantothere Kuehneotherium praecursoris. Zool J Linn Soc 47:407–423
Kermack KA, Mussett F, Rigney HW (1981) The skull of Morganucodon. Zool J Linn Soc 71:1–158
Kermack KA, Lee AJ, Lees PM, Mussett F (1987) A new docodont from the Forest Marble. Zool J Linn Soc 89:1–39
Kielan-Jaworowska Z, Cifelli RL, Luo Z-X (2004) Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. Columbia University Press, New York
Krebs B (2000) The excavations in the Guimarota mine. In: Martin T, Krebs B (eds) Guimarota ̶ A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, Munich, pp 9–20
Krusat G (1980) Contribuçăo para o conhecimento da fauna do Kimeridgiano da mina de lignito Guimarota (Leiria, Portugal). IV Parte. Haldanodon exspectatus Kühne & Krusat 1972 (Mammalia, Docodonta). Mems Serv Geol Portugal 27:1–79
Krusat G (1991) Functional morphology of Haldanodon exspectatus (Mammalia, Docodonta) from the Upper Jurassic of Portugal. In: Kielan-Jaworowska Z, Heintz N, Nakrem HA (eds) Fifth Symposium on Mesozoic Terrestrial Ecosystems and Biota. Contrib Paleont Mus Univ Oslo 363:37–38
Kühne W, Krusat G (1972) Legalisierung des Taxon Haldanodon (Mammalia, Docodonta). N Jb Geol Paläont Mh 1972:300–302
Lillegraven JA, Krusat G (1991) Cranio-mandibular anatomy of Haldanodon exspectatus (Docodonta; Mammalia) from the Late Jurassic of Portugal and its implications to the evolution of mammalian characters. Contrib Geol Univ Wyo 28:39–138
Lopatin AV, Averianov AO (2005) A new docodont (Docodonta, Mammalia) from the Middle Jurassic of Siberia. Doklady Biol Sci 405:434–436
Luckett WP (1993) An ontogenetic assessment of dental homologies in therian mammals. In: Szalay FS, Novacek MJ, McKenna MC (eds) Mammal Phylogeny. Mesozoic Differentiation, Multituberculates, Monotremes, Early Therians, and Marsupials. Springer-Verlag, Berlin, New York, Heidelberg, pp182–204.
Luo Z-X (1994) Sister-group relationship of mammals and transformations of diagnostic mammalian characters. In: Fraser NC, Sues HD (eds) In the Shadow of the Dinosaurs: Early Mesozoic Tetrapods. Cambridge University Press, Cambridge, pp 98–128
Luo Z.-X., Martin T (2007) Analysis of molar structure and phylogeny of docodontan genera. Bull Carnegie Mus Nat Hist 39:27–47
Luo Z-X, Crompton AW, Sun A-L (2001) A new mammaliaform from the Early Jurassic of China and evolution of mammalian characteristics. Science 292:1535–1540
Luo Z-X, Kielan-Jaworowska Z, Cifelli RL (2004) Evolution of dental replacement in mammals. Bull Carnegie Mus Nat Hist 36:159–175
Luo Z-X, Meng Q-J, Ji Q, Liu D, Zhang Y-G, Neander AI (2015) Evolutionary development in basal mammaliaforms as revealed by a docodontan. Science 347:760–764
Martin T (1997) Tooth replacement in Late Jurassic Dryolestidae (Eupantotheria, Mammalia). J Mammal Evol 4:1–18
Martin T (1999) Dryolestidae (Eupantotheria, Mammalia) aus dem Oberen Jura von Portugal. Abh senckenberg naturforsch Ges 550:1–119
Martin T (2005) Postcranial anatomy of Haldanodon exspectatus (Mammalia, Docodonta) from the Late Jurassic (Kimmeridgian) of Portugal and its bearing for mammalian evolution. Zool J Linn Soc 145:219–248. https://doi.org/10.1111/j.1096-3642.2005.00187.x
Martin T (2018) Mesozoic mammals - early mammalian diversity and ecomorphological adaptations. In Zachos FE, Asher RJ (eds) Handbook of Zoology, Mammalia. Mammalian Evolution, Diversity and Systematics. De Gruyter, pp 199–299
Martin T, Nowotny M (2000) The docodont Haldanodon from the Guimarota Mine. In: Martin T, Krebs B (eds) Guimarota – A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, Munich, pp 91–96
Martin T, Nowotny M, Fischer M (2010) New data on tooth replacement in the Late Jurassic docodont mammal Haldanodon exspectatus. SVP Program and Abstracts Book 2010:130A
Martin T, Jäger KRK, Plogschties T, Schwermann AH, Brinkkötter JJ, Schultz JA (2020) Molar diversity and functional adaptations in Mesozoic mammals. In: Martin T, Koenigswald Wv (eds) Mammalian Teeth – Form and Function. Verlag Dr. Friedrich Pfeil, Munich, pp 187–214
Meng Q-J, Ji Q, Zhang Y-G, Liu D, Grossnickle DM, Luo Z-X (2015) An arboreal docodont from the Jurassic and mammaliaform ecological diversification. Science 347:764–768
Mills JRE (1971) The dentition of Morganucodon. In: Kermack DM, Kermack KA (eds) Early Mammals. Zool J Linn Soc 50(Supp 1):29–63
Mills JRE (1984) The molar dentition of a Welsh pantothere. Zool J Linn Soc 82:189–205
Nowotny M, Martin T, Fischer M (2001) Dental anatomy and tooth replacement of Haldanodon exspectatus (Docodonta, Mammalia) from the Upper Jurassic of Portugal. J Morphol 248:268.
O´Meara RN, Asher RJ (2016) The evolution of growth patterns in mammalian versus nonmammalian cynodonts. Paleobiology 42:439–464.
Osborn JW (1974) On the tooth succession in Diademodon. Evolution 28:141–157
Osborn JW, Crompton AW (1973) The evolution of mammalian from reptilian dentitions. Breviora 399:1–18
Panciroli E, Benson RB, Fernandez V, Butler RJ, Fraser, NC, Luo, Z-X, Walsh S (2021) New species of mammaliaform and the cranium of Borealestes (Mammaliformes: Docodonta) from the Middle Jurassic of the British Isles. Zool J Linn Soc 192:1323–1362.
Panciroli E, Benson RBJ, Vernandez V, Humpage M, Martín-Serra A, Walsh S, Luo Z-X, Fraser NC (2022) Postcrania of Borealestes (Mammaliaformes, Docodonta) and the emergence of ecomorphological diversity in early mammals. Palaeontology 65(1):e12577. https://doi.org/10.1111/pala.12577
Parrington FR (1936) On the tooth-replacement in theriodont reptiles. Phil Trans Roy Soc Lond B 226:121–142
Parrington FR (1971) On the Upper Triassic mammals. Phil Trans Roy Soc Lond B 261:231–272
Parrington FR (1973) The dentitions of the earliest mammals. Zool J Linn Soc 52:85–95
Pfretzschner H-U, Martin T, Maisch M, Matzke A, Sun G (2005) A new docodont from the Late Jurassic of the Junggar Basin of northwest China. Acta Palaeontol Pol 50:799–808
Remane A (1952) Die Grundlagen des natürlichen Systems, der vergleichenden Anatomie und der Phylogenetik. Geest & Portig KG, Leipzig
Rougier GW, Sheth AS, Carpenter K, Appella-Guiscafre L, Davis BM (2015) A new species of Docodon (Mammaliaformes: Docodonta) from the Upper Jurassic Morrison Formation and a reassessment of selected craniodental characters in basal mammaliaforms. J Mammal Evol 22:1–16 https://doi.org/10.1007/s10914-014-9263-8
Schultz JA, Bhullar B-AS, Luo ZX (2019) Re-examination of the Jurassic mammaliaform Docodon victor by computed tomography and occlusal functional analysis. J Mammal Evol 26:9–38.
Sigogneau-Russell D (2001) Docodont nature of Cyrtlatherium, an upper Bathonian mammal from England. Acta Palaeontol Pol 46:427–430
Sigogneau-Russell D (2003) Docodonts from the British Mesozoic. Acta Palaeontol Pol 48:357–374
Simpson GG (1928) A Catalogue of the Mesozoic Mammalia in the Geological Department of the British Museum. Trustees of the British Museum, London
Simpson GG (1929) American Mesozoic Mammalia. Mem Peabody Mus Yale Univ 3:1–235
Yang Z-J [Young CC] (1982) Selected Works of Yang Zhongjian (Young Chung-Chien). (Edited by the Editorial Committee for Selected Works of Yang Zhongjian, Chaired by MC Chow [M-C Zhou]) Science Press, Beijing
Zhang F-K, Crompton AW, Luo Z-X, Schaff CR (1998) Pattern of dental replacement of Sinoconodon and its implications for evolution of mammals. Vertebr PalAsiatica 36:197–217 (Chinese and English)
Acknowledgements
Field work in Portugal under the auspices of Bernard Krebs and Siegfried Henkel (both Freie Universität Berlin) was extensively supported by the former Serviços Geológicos de Portugal (Lisbon). Special thanks are due to the then directors of this institution, António de Castelo Branco, Fernando Moitinho de Almeida, and Miguel M. Ramalho for their long-term goodwill. Ellen Eggert (née Drescher; Freie Universität Berlin) masterfully prepared the Haldanodon jaws. Angelika Helfricht (Senckenberg Forschungsinstitut und Naturmuseum, Frankfurt/M.) provided the drawings of Figs. 17 and 18, and Anna Kizner (Universität Bonn) and Cathrin Pfaff (now Universität Wien) assisted with the 3D reconstructions. Reviewers Elsa Panciroli (University of Oxford Museum of Natural History) and Helder Gomes Rodrigues (Géosciences, Université II Montpellier) as well as editor Robert Asher (University of Cambridge) provided valuable comments that greatly improved our manuscript. The excavations in the Guimarota mine (1973–1982) were funded by the Freie Universität Berlin and the DFG (German Research Foundation). The Universität Bonn granted a sabbatical leave to TM.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
T.M. wrote the main manuscript text and J.A.S. performed the scanning and segmentation of specimens and prepared the figures. Both authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this article was revised: The article included some left/right and numbering errors in the text and three formatting issues in the Results section. Also, the specimen numbers in Table 1 should be prefixed by “MG” rather than “MGL”.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Martin, T., Schultz, J.A. Deciduous dentition, tooth replacement, and mandibular growth in the Late Jurassic docodontan Haldanodon exspectatus (Mammaliaformes). J Mammal Evol 30, 507–531 (2023). https://doi.org/10.1007/s10914-023-09668-2
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10914-023-09668-2