
Abstract
In two independent experiments, we compared: (1) water depth selection (and accompanying temperature selection) by male and female Daphnia magna under different kinds of environmental stress, including the presence of filamentous cyanobacteria, the risk of predation from fish, and the presence of toxic compounds; and (2) sex-dependent production of heat shock proteins (HSP60, 70, and 90) in response to a sudden change in temperature. Male D. magna selected deep water strata, which offer a relatively stable environment, and thereby avoided the threat of predation and the presence of toxic compounds in surface waters. Correlated with this behavior, males reduce their molecular defenses against stress, such as the production of heat shock proteins (HSPs), and do not maintain the physiological machinery that triggers an increase in HSP levels in response to stress. In contrast, female D. magna actively select habitats that offer optimal conditions for growth and production of offspring. Consequently, females are exposed to variable environmental conditions that may be associated with increased stress. To permit survival in these different habitats, D. magna females require molecular mechanisms to protect their cells from rapid changes in stress levels. Thus, they maintain high constitutive levels of the heat shock proteins from HSP 60, 70, and 90 families, and they have the potential to further enhance the production of the majority of these proteins under stress conditions. The results of this study indicate that the separate habitats selected by male and female D. magna result in different patterns of HSP production, leading us to hypothesize that that male and female Daphnia magna adopt different strategies to maximize the fitness of the species.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In many species, differences between males and females are not restricted to the morphology of sexual structures involved in mating or fertilization (Andersson, 1994). Individuals of each sex possess unique adaptations to maximize fitness. Under favorable environmental conditions, most water fleas, Daphnia spp. (Cladocera, Daphniidae) reproduce parthenogenetically. When conditions become adverse, Daphnia females produce resting eggs and deposit them into a special carapace structure called the ephippium. For successful hatching, resting eggs require fertilization before storage in the ephippium (Hobæk and Larsson, 1990; Pijanowska and Stolpe, 1996; Shurin and Dodson, 1997; Gyllström and Hansson, 2004).
The optimal strategy of female Daphnia involves a trade-off between the necessity to assimilate enough energy to grow and produce offspring, and the avoidance of environmental risks (e.g., predation or toxicity). The impact of majority of stress factors is associated with surface water. Harmful chemicals that reach the lake in summer remain in the epilimnion (surface, warm water stratum). Even when these stresses reach the hypolimnion (deep, cold, and dark water stratum), their influence on organisms is limited due to low temperature (Cairns et al., 1975). Risk from predaceous fish that rely on vision to locate their prey, strongly depends on light and, as a consequence, is constrained to the upper strata. Because of the high temperatures and the abundance of food, epilimnetic conditions allow rapid growth (Wetzel, 2001). The necessity of avoiding the most dangerous locations at the surface and, at the same time, maximizing growth and reproduction has promoted the evolution of vertical migration behavior in Daphnia (e.g., Dawidowicz and Loose, 1992). Unlike females, males do not need to attain a large size or gather as much energy for reproduction. Their fitness depends on the number of females they fertilize during their lifetime. Therefore, their main goal is continued survival. We hypothesize that male Daphnia magna Straus should remain deeper in the water column than females, where they experience lower temperatures and food concentrations, but avoid predation by fish and adverse factors like cyanobacteria or toxins that are more prevalent in surface waters. Such male behavior has been described in Daphnia galeata by Spaak and Boersma (2001), who explained the inter-sexual differences in migratory behavior as a strategy to avoid inbreeding.
One of the main consequences of vertical migration in stratified lakes is exposure to rapid changes in temperature. For example, D. magna migrating twice a day – from the epilimnion to the hypolimnion and back – can experience a temperature gradient of 10-15°C. The reaction to thermal shock at the cellular level involves the expression of heat shock (or stress) proteins (HSPs) that protect cell structures against thermal stress (Jolly and Morimoto, 2000).
A change in the level of HSPs is one of the most widely recognized molecular responses to stress in pro- and eukaryotic organisms (Lindquist and Craig, 1988; Mager and de Kruijff, 1995; Jolly and Morimoto, 2000). HSPs were first discovered in Drosophila busckii following exposure to high temperature (Ritossa, 1962). They are divided into several groups (families) of different molecular weights (kDa): e.g., HSP100, HSP90, HSP70, HSP60, HSP40, and small proteins. HSP60s control modifications of newly synthesized proteins, repair damaged ones, and prevent peptides from accumulating (Jolly and Morimoto, 2000; Pockley, 2003). HSP70s are the most important molecular chaperones permanently active in the cell. In cooperation with other proteins, they are responsible for cellular homeostasis under non-stress conditions. Moreover, they play a vital role in stress tolerance and survival under adverse conditions. HSP70s also act to reduce accumulation of peptide aggregates, but if this does happen they can dissolve aggregates. HSP70s also promote the correct folding of newly synthesized proteins (Mayer and Bukau, 2005). HSP90s play a major role in stress tolerance. They act to remove proteins with incorrect structure, and by mediating proper folding under stress conditions, they prevent the accumulation of peptide aggregates. They also regulate the activity of other proteins (e.g., kinases) and stabilize the cytoskeleton (Csermely et al., 1998).
The induction of HSPs in Daphnia sp. was first described as a reaction to arsenic stress (Chen et al., 1999). Few studies have examined changes in HSP levels in response to biotic factors such as predation risk (Pijanowska and Kloc, 2004; Pauwels et al., 2005) and parasite infection (Pauwels et al., 2007). We recently have observed enhanced production of HSP60, HSP70, and HSP90 in heat-shocked D. magna (Mikulski et al., 2009). If the amplitude of migration of D. magna females is indeed broader than that of males, there are likely to be sex-related differences in the molecular response of HSPs to changes in temperature experienced by these animals.
The primary aim of this study was to examine whether depth (temperature) selection by D. magna in response to different kind of stress [the presence of cyanobacteria, fish, and polychlorinated biphenyls (PCBs) and their various combinations] differs between males and females. A second aim was to determine whether male and female Daphnia differ in the constitutive level of stress proteins and their production in response to changing temperature.
Methods and Materials
Two experiments were conducted on genetically identical individuals of a D. magna clone originating from Binnensee, a shallow coastal lake in northwestern Germany. To avoid any maternal effect, D. magna were cultured in 0.8 l of medium in glass jars (20 animals per jar) at 20°C under a summer photoperiod (16:8 h L:D) for several generations before the experiment. The experimental medium was lake water that had been aerated for a few weeks and filtered through a 0.2 μm capsule filter. Animals were fed green algae, Scenedesmus obliquus, at the non-limiting concentration of 1 mg C/l (Lampert, 1987), and the experimental medium was changed daily. Neonates from the second clutch were used for further breeding.
In the first experiment, differences in depth selection between D. magna males and females exposed to different kinds of stress were recorded. The stresses tested were the presence of filamentous cyanobacteria, the threat of fish predation, and the presence of toxic compounds, all of which are more severe in surface waters (reviewed by Mengel and Clarck, 1988). Moreover, all of these factors are known to influence depth selection by cladocerans (Bernatowicz and Pijanowska, 2010). In the second experiment, the levels of HSPs were quantified in male and female D. magna exposed to thermal stress as a surrogate for depth selection.
Depth Selection of Males and Females
A single cohort of D. magna neonates from the pre-experimental culture was transferred to several 0.8 l glass jars (20 animals of unknown sex per jar) and cultured at 20°C under a summer photoperiod (16:8 h L:D) for 6 d after birth. The animals were fed green algae, Scenedesmus obliquus, at 1 mg C/l, and the growth medium was changed daily.
On the seventh day, after all animals had reached maturity, some were selected for further experiments and transferred (3 females or 5 males per tube) to flow-through tubes (60 cm long, 1 cm diam) placed in a water bath, with surface waters heated to 22°C and bottom waters cooled to 9°C, and illuminated from above. This system, known as a “plankton organ,” is widely used for studies on the diel vertical migration behavior of Daphnia (see Dawidowicz and Loose, 1992). The temperature gradient (Fig. 1) was stable throughout the experiment in all tubes. The specific stress factors then were applied, in four replicates (tubes) each, to both females and males: (1) the toxic cyanobacterium, Cylindrospermopsis raciborskii (Cy), from a laboratory culture added to the standard medium at a concentration of 1 mg C/l; (2) fish-conditioned (Fi) medium prepared by using water in which crucian carp, Carassius carassius, had been held for 24 h (1 fish per 10 l water); (3) toxic polychlorinated biphenyls PCB 52; and (4) PCB 153 added at a concentration of 10 μg/l. In the control treatment, standard medium containing only the Scenedesmus obliquus food was used. In addition to these single stressor treatments, 7 different combinations of these stress factors also were applied (see Fig. 2). The flow-through system ensured even distribution of food and chemicals in all tubes.

The thermal profile in the experimental "plankton organ" tubes used to assay responses of Daphnia magna to various stress factors

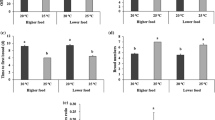
The effect of different stress factors on a depth and b temperature selection (mean ± s.d.) by Daphnia magna males and females. C–control; Cy–cyanobacteria, Cylindrospermopsis raciborskii; F–fish, crucian carp, Carassius carassius; 153–toxic polychlorinated biphenyl (PCB)153; and 52–PCB52. * P < 0.05 (significant difference between sexes in temperature selection within the same treatment; Tukey test, N = 4)
The depth of each animal in each tube was recorded at noon, for 2 d. From these data, the temperature experienced by individuals at particular depths was extrapolated (based on data shown in Fig. 1). The data were analyzed by using two-way nested ANOVA (with tube effect nested in treatment and sex as independent variables). Based on the mean temperature experienced by animals in the separate treatments, Levene’s test of homogeneity of variance (Levene, 1960) was applied to compare the variability of temperature-depth selection between males and females exposed to the experimental factors.
Analysis of HSP Levels in Males and Females
A single cohort of D. magna neonates from the pre-experimental culture was transferred to several 0.8 l glass jars (20 animals of unknown sex per jar) and cultured at 20°C under constant dim light until first reproduction. Animals were fed green algae, Scenedesmus obliquus, at a concentration of 1 mg C/l and the growth medium was changed daily. After reaching maturity, animals were transferred to twelve glass jars (each treatment replicated three times) containing 200 ml medium (5 females or 15 males per jar). The combined mass of 5 females was approximately equal to that of 15 males, so the total mass of animals in each jar was roughly equivalent. After 24 h, half of the medium was removed from each jar. The same medium was immediately pumped back into 6 control jars (3 with males, 3 with females), while the other 6 jars were filled with hot medium to produce a 10°C increase in the water temperature within a few seconds. These jars were transferred to a 30°C water bath for 1 h. The range of temperature change was selected based on Mikulski et al. (2009). The thermal gradient set in this experiment (and the resulting thermal shock experienced by the animals) was more severe than in those conducted previously. Our unpublished data show that coping with even milder temperature gradients, e.g., crossing the thermocline (from 20°C at the surface to 15°C at the preferable depth), generates significant costs for female D. magna in terms of their life history and through enhanced production of HSPs. In this experiment, the severity of the thermal stress was increased to highlight the impairment of HSP production in males.
After this period of incubation at 30°C, the jars were removed from the water bath and slowly (over about 20 min) cooled to 20°C. Six h after the start of the heat treatment, the D. magna from each jar were collected, immediately dried on a sieve, and then transferred to 1.5 ml microcentrifuge tubes and homogenized in 75 μl of RIPA buffer (0.15 M NaCl, 1% deoxycholate Na salt, 1% Triton X-100, 0.1% SDS, 0.01 M Tris-HCl, pH 7.2) containing Complete protease inhibitors (Roche, Basel, Switzerland, Cat. No. 11836153001). The homogenates were centrifuged for 1 min at 13.4 rcf, and 10 μl aliquots of each supernatant were used to measure protein concentration with a DC Protein Assay (Bio-Rad, Berkeley, CA, USA, Cat. No. 500–0001) to ensure equal protein loading.
The remaining supernatants were transferred to fresh microcentrifuge tubes, and an equal volume of SDS-PAGE loading buffer (Sigma-Aldrich, St. Louis, MO, USA, Cat. No. L4390) was added. After boiling for 5 min, samples containing equivalent amounts of protein (30 μg) were separated by electrophoresis on 25% SDS-polyacrylamide gels, and HSPs were immunodetected by western blotting with four different primary antibodies made by Stressgen (Victoria, British Columbia, Canada): Anti-Hsp60 (Stressgen, Cat. No. SPA-805 and Cat. No. SPA-807), Anti-Hsp70 (Stressgen, Cat. No. SPA-812) and Anti-Hsp90 (Stressgen, Cat. No. SPA-846). The blots then were incubated with alkaline phosphatase-conjugated secondary antibody (Anti-Rabbit IgG - Stressgen, Cat. No. 611–1502), and positive immunoreaction was identified by a color reaction with SIGMAFAST™ BCIP/NBT substrate (Sigma-Aldrich, Cat. No. B5655).
The processed blots were examined with a Bio-Rad Gel-Doc system, and the intensity of immunostained bands was determined by using Quantity One analysis software (Bio-Rad, Cat. No #170-9600).
The relative amounts of the individual HSPs in each sample were calculated as a percentage of the intensity of the corresponding HSP bands in a thermally stressed human HeLa cell positive control sample (Stressgen, Cat. No LYC-HL101) run alongside the test samples. Data were analyzed by using the ANOVA model and post-hoc Tukey test with Statistica (StatSoft, Tulsa, OK, USA).
The Anti-Hsp60 and Anti-Hsp70 antibodies revealed multiple immunoreactive protein bands of different molecular weights. These bands were treated as separate proteins, as described by Mikulski et al. (2009).
Results
Generally, the stress factors (toxic cyanobacteria, presence of fish, toxic polychlorinated biphenyls, or combinations of these cues) (ANOVA: F (11, 288) = 2.24, P = 0.013) and sex (ANOVA: F (1, 288) = 207.99, P < 0.001) influenced depth (temperature) selection of D. magna (Fig. 2). Males occupied colder (deeper) water strata than females (Tukey test: P < 0.001). There was a strong interaction between sex and treatment effects (ANOVA: F (11, 288) = 2.24, P < 0.001). The different types of environmental stress resulted in higher variability of depth (and temperature) selection in D. magna females than in males (Fig. 2; Table 1; Levene’s test: F (1,22) = 11.25, P = 0.003). Thus, males did not change their depth (temperature), but females varied their depth according to the nature of the environmental stress.
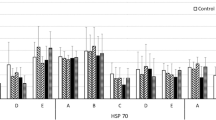
The pattern of production of different HSPs detected in D. magna (Mikulski et al., 2009) depended on the sex of the animal (Table 2, Fig. 3), and apart from the 79-kDa protein, the level of all HSPs was significantly higher in females than in males (Fig. 4). The level of the majority of HSPs (50 kDa, 55 kDa, 73–74 kDa, and 92 kDa) also was dependent on temperature (Table 2, Figs. 3 and 4). However, a significant increase in the level of these HSPs in response to raised temperature was only observed in females (see Fig. 3). There was a significant interaction between the effect of thermal shock and the effect of sex on the level of three HSPs: 50 kDa, 55 kDa, and 73–74 kDa (Table 2 and Fig. 3).

Stress proteins in male (Ma) or female (Fe) Daphnia magna in control (−C) and heat-shock (−S) conditions revealed by anti-HSP60 (a), anti-HSP70 (b) and anti-HSP90 (c) antibodies, against heat-shocked human HeLa cell lysates

The relative amounts (mean ± s.d.) of different HSPs expressed in control (white bars) and heat-shocked (shaded bars) male and female Daphnia magna. Optical density is expressed as a percentage of the density of the respective HSP bands in a thermally stressed human HeLa cell positive control sample run on the same gel; * P < 0.05 (significant effect of heat shock on protein level, Tukey test, N = 3)
Discussion
According to our hypothesis, depth-temperature selection is more dependent on environmental stress in D. magna females than in males (Fig. 2). To achieve reproductive success, females must grow and assimilate sufficient energy. Therefore, they undertake vertical migration and adjust their depth according to the availability of food and the level of environmental stress (Dawidowicz and Loose, 1992). In contrast, the fitness of adult males seems to be poorly correlated with assimilation rate; rather, it would be expected to depend on their longevity, which is related more to safety than growth rate (Roff, 1992). Albeit smaller than females, size of males renders them susceptible to fish predation (e. g., Eggers, 1982). Therefore, males assimilate less energy and grow more slowly while staying in deeper water (poor in food but thermally stable), waiting for the appearance of receptive females. Since D. magna females are exposed to greater variability in environmental (including thermal) conditions than males, their molecular anti-stress machinery should be adequately developed.
We found that D. magna females maintain generally higher levels of HSPs than males (Fig. 4). Furthermore, a significant increase in the level of these HSPs in response to thermal stress was observed solely in females (this inter-sex difference is supported by the significant interaction between the effect of thermal shock and the effect of sex on the level of the majority of heat shock proteins). Apparently, females that perform diel vertical migrations protect themselves against various stress factors associated with surface waters by maintaining a constitutively high level of HSPs and by greater responsiveness to temperature change. The only exception was HSP 79 kDa, which showed the opposite pattern, being more abundant in males than in females. It has been suggested that HSP 79 kDa represents a post-translational modification of HSP 73–74 kDa (Jolly and Morimoto, 2000). The intriguing physiological role of HSP70 in aquatic invertebrates is not well characterized. Higher levels of HSP70 in males compared with females previously have been described in Drosophila, although this apparently is not associated with greater survival in males (Dahlgaard et al., 1998). Only more detailed statistical analysis revealed a positive association between HSP70 level and survival after the induction of thermotolerance.
The results of our study show that behavioral and molecular defenses against environmental stress may be interrelated. Daphnia magna males can avoid dangers associated with surface waters by selecting the safe and stable (albeit cold with scarce food resources) environment of deep water strata. They may thus save energy by abandoning the expression of molecular defenses against stress. In contrast, D. magna females actively select an environment that is optimal for growth and reproduction, and, in consequence, are exposed to a broader range of environmental conditions that will unavoidably include stresses. For survival in these habitats, they require molecular mechanisms to protect their cells from adverse environmental conditions and rapid changes in stress levels. We have shown that these two strategies are adopted by genetically identical individuals of the same clone (sex in D. magna is environmentally determined; reviewed by Eads et al. 2008). The choice of both a behavioral strategy (migrating in the water column or permanently residing at certain depths) and a molecular strategy (e.g., activation of an inducible molecular anti-stress system) can thus constitute novel elements of phenotypic plasticity.
Although it is unwise to over-interpret the identification of these apparently linked defense mechanisms in a single clone (different genotypes are known to exhibit different stress responses), our study represents the first comprehensive analysis of sex-related differences in the levels of stress proteins in D. magna in an ecological and evolutionary context.
References
Andersson, M. 1994. Sexual Selection. Princeton University Press, Princeton, NJ.
Bernatowicz, P., and Pijanowska, J. 2010. Daphnia response to biotic stress is modified by PCBs. Ecotoxicol. Environ. Saf. 74:711–718.
Cairns, J., Heath, A. G., and Parker, B. C. 1975. The effects of temperature upon the toxicity of chemicals to aquatic organisms. Hydrobiologia 47:135–171.
Chen, C. Y., Sillett, K. B., Folt, C. L., Whittemore, S. L., and Barchowsky, 1999. Molecular and demographic measures of arsenic stress in Daphnia pulex. Hydrobiologia 401:229–238.
Csermely, P., Schnaider, T., Soti, C., Prohászka, Z., and Nardai, G. 1998. The 90-kDa molecular chaperone family: structure, function, and clinical applications. A comprehensive review. Pharmacol. Ther. 79:129–168.
Dahlgaard, J., Loeschcke, V., Michalak, P., and Justesen, J. 1998. Induced thermotolerance and associated expression of the heat-shock protein Hsp70 in adult Drosophila melanogaster. Funct. Ecol. 12:786–793.
Dawidowicz, P., and Loose, C. J. 1992. Metabolic costs during predator-induced diel vertical migration of Daphnia. Limnol. Oceanogr. 37:1589–1595.
Eads, B. D., Andrews, J., and Colbourne J. K. 2008. Ecological genomics in Daphnia: stress responses and environmental sex determination. Heredity 100:184–190.
Eggers, D. M. 1982. Planktivore preference by prey size. Ecology 63:381–390.
Gyllström, M., and Hansson, L. A. 2004. Dormancy in freshwater zooplankton: Induction, termination and the importance of benthic-pelagic coupling. Aquat. Sci. 66:274–295.
Hobæk, A. and Larsson, P. 1990. Sex determination in Daphnia magna. Ecology 71:2255–2268.
Jolly, C., and Morimoto, R. I. 2000. Role of the heat shock response and molecular chaperones in oncogenesis and cell death. J. Nat. Canc. Inst. 92:1564–1572.
Lampert, W. 1987. Feeding and nutrition in Daphnia. Mem. Ist. Ital. Idrobiol. 45:143–192.
Levene, H. 1960. Robust tests for equality of variances, pp. 278–292, in I. Olkin, S. G. Ghurye, W. Hoeffding, W. G. Madow, and H. B. Mann (eds.). Contributions to Probability and Statistics. Essays in Honor of Harold Hotelling. I. Stanford University Press, Palo Alto, CA.
Lindquist, S., and Craig, E. A. 1988. The heat-shock proteins. Annu. Rev. Genet. 22:631–677.
Mager, W. H., and De Kruijff, A. J. 1995. Stress-induced transcriptional activation. Microbiol. Rev. 59:506–531.
Mayer, M. P., and Bukau, B. 2005. Hsp70 chaperones: cellular functions and molecular mechanism. Cell. Mol. Life Sci. 62:670–684.
Mengel, M., and Clarck, C. W. 1988. Dynamic Modeling in Behavioral Ecology. Princeton University Press, Princeton, NJ.
Mikulski, A., Grzesiuk, M., Kloc, M., and Pijanowska, J. 2009. Heat shock proteins in Daphnia detected using commercial antibodies: description and responsiveness to thermal stress. Chemoecology 19:69–72.
Pauwels, K., Stoks, R., and De Meester, L. 2005. Coping with predator stress: interclonal differences in induction of heat-shock proteins in the water flea Daphnia magna. J. Evol. Biol. 18:867–872.
Pauwels, K., Stoks, R., Decaestecker, E., and De Meester, L. 2007. Evolution of heat shock protein expression in a natural population of Daphnia magna. Am. Nat. 170:800–805.
Pijanowska, J., and Stolpe, G. 1996. Summer diapause in Daphnia as a reaction to the presence of fish. J. Plankt. Res. 18:1407–1412.
Pijanowska, J., and Kloc, M. 2004. Daphnia response to predation threat involves heat-shock proteins and acid and tubulin cytoskeleton. Genesis 38:81–86.
Pockley, A. G. 2003. Heat shock proteins as regulators of the immune response. Lancet 362:469–476.
Ritossa, F. 1962. A new puffing pattern induced by temperature shock and DNP in Drosophila. Experientia 18:571–573.
Roff, D. A. 1992. The Evolution of Life Histories. Chapman & Hall, New York and London, pp. 207–208.
Shurin, J. B., and Dodson, S. I. 1997. Sublethal toxic effects of cyanobacteria and nonylphenol on environmental sex determination and development in Daphnia. Environ. Toxicol. Chem. 16:1269–127.
Spaak, P., and Boersma, M. 2001. The influence of fish kairomones on the induction and vertical distribution of sexual individuals of the Daphnia galeata species complex. Hydrobiologia 442:185–193.
Wetzel, R. G. 2001. Limnology: Lake and River Ecosystems, Academic Press, San Diego, CA.
Acknowledgments
This study was supported by the Polish Ministry of Science and Higher Education (NN304 094135). We are grateful to several anonymous reviewers for their helpful comments on earlier versions of the manuscript.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Mikulski, A., Bernatowicz, P., Grzesiuk, M. et al. Differential Levels of Stress Proteins (HSPs) in Male and Female Daphnia magna in Response to Thermal Stress: A Consequence of Sex-Related Behavioral Differences?. J Chem Ecol 37, 670–676 (2011). https://doi.org/10.1007/s10886-011-9969-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10886-011-9969-5