
Abstract
Beavers and their dams, once common in small streams throughout Eurasia and North America, are returning to their original range. Beaver dams are special habitats in the aquatic-terrestrial interface, but their macroinvertebrate fauna is hardly known. This study aims to qualify and quantify the invertebrate fauna of beaver dams, taking into account the maintenance status of the dams. Nine different areas covering the top, middle and base zones of eight maintained and eight abandoned beaver dams were sampled using a specially designed suction device. The invertebrate community of beaver dams proved to be diverse and predominantly rheophilic. Community composition reflects higher flow velocities in streamside habitats; this gradient is more pronounced in maintained than in abandoned dams. Shredders are the most abundant feeding type, but they are less frequent in streamside habitats, in particular in those of maintained dams, where passive filter feeders prevail. Terrestrial/Semiaquatic taxa colonising hygropetric areas were found in streamside habitats especially of abandoned dams. Our results underline that beaver dams provide a wide variety of environmental conditions and habitat types, which enhance biodiversity in small streams and their floodplains. Beaver dams in riverine landscapes should therefore be much more valued in floodplain conservation and management.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Beaver dams are among the animal kingdom’s architectural masterpieces, significantly modifying hydraulics, morphology and community composition of the streams where there are built (Müller-Schwarze, 2011). Beavers, Castor canadensis Kuhl, 1820 and C. fiber Linnaeus, 1758, construct dams exclusively in small streams up to 4th order to increase the water depth, thus enabling underwater access to their lodges (McDowell & Naiman, 1986; Collen & Gibson, 2001). River width, depth and slope, along with watershed area, floodplain morphology and vegetation cover, determine whether dams are built, their density, size and dimensions (Müller-Schwarze, 2011; Zavyalov, 2014). Due to individual site characteristics, the number of dams can vary between 0.1 dam/km (McComb et al., 1990) and 19 dams/km (Woo & Waddington, 1990).
Building materials for dams are mainly logs of varying lengths, stones, and mud (Müller-Schwarze, 2011; Rolauffs et al., 2001; Butler & Malanson, 1995). The beaver skilfully integrates these materials into a tightly interwoven structure, adapted to channel morphology (Kroes & Bason, 2015) and flow velocity (Andersen & Shafroth, 2010). For stabilisation, beavers seal the flow-facing side of the dams with mud (Müller-Schwarze, 2011); due to the reduced flow velocity, sediments accumulate in the ponds and the adjacent dam structure (Butler & Malanson, 1995; Kroes & Bason, 2015). Nevertheless, beaver dams retain their permeability, which is influenced by various environmental factors and the beavers’ maintenance activities (Ronnquist & Westbrook, 2021). The sizes and shapes of beaver dams varies greatly, with the usual pattern being concave-upstream arches (Warren, 1932; Dugmore, 1914). The range of recorded dam heights varies between 0.3 and 5 m (Müller, 2014), while the large majority is on average 1 m in height (Hafen et al., 2020). The length can measure 0.6 m in small canal dams (Townsend, 1953) up to several hundred metres (Thie, 2022), depending on the topography (Gurnell, 1998). Thereby, the construction activities of Castor canadensis and Castor fiber are very much the same, if both species are observed in the same areas with identical environments (Danilov et al., 2011). Danilov et al. (2011) demonstrate that the construction activity by beavers is exclusively determined by geomorphological and hydrographic settings in the habitat and that building behaviour has no species-specific features. Rolauffs et al. (2001) highlights the large internal surface area of beaver dams resulting in a network of interstitial spaces easily accessible to organisms. Furthermore, short-term flooding and drying within the dam causes rapid shifts between terrestrial and aquatic conditions (Rolauffs et al., 2001). Flow velocity within the dams can be very high and variable (Woo & Waddington, 1990), creating microhabitats with a lotic character in lowland streams, where otherwise lentic conditions prevail (Clifford et al., 1993). Beaver dams are constantly exposed to changing flow conditions and decomposition processes (Woo & Waddington, 1990; Ronnquist & Westbrook, 2021). Because of these dynamics, beavers repair and maintain their dams, and sometimes expand them to increase the size of the upstream pond (Richard, 1967; Wohl et al., 2019). These activities enhance habitat variability and flow diversity between the upper and lower parts of a dam (Rolauffs et al., 2001). Likewise, dams are regularly neglected or abandoned when they no longer serve any purpose or the beavers leave the area (Woo & Waddington, 1990; Bylak & Kukuła, 2018). This reduces dam stability and increases dam permeability, as fine material is washed out and the water is flowing more rapidly through the remaining skeleton of branches, without allowing for much habitat variability from top to base (Woo & Waddington, 1990). The construction and maintenance of dams is an ongoing process, thus there is a large variance in the lifetime and maintenance states of the dams within a beaver territory (Johnston, 2017). Thereby, single dams can withstand or be maintained from several months to decades (Johnston, 2017). As beavers rapidly increase their range in Europe (Halley et al., 2021; Wróbel, 2020), both maintained and abandoned beaver dams are increasingly common elements within the valleys of small streams.
From the perspective of macroinvertebrates, beaver dams, i.e. piles of dead wood in the aquatic-terrestrial interface, are very special habitats, but little is known on the communities colonising them. In general, beaver dams offer a wide variety of food sources to the invertebrate community: wood of different size (Barnes & Mallik, 1996; Blersch & Kangas, 2014), from different trees species (Barnes & Mallik, 1996; Blersch & Kangas, 2014), and in different decomposition stages (Woo & Waddington, 1990); tufts of grass, organic debris and accumulating leaves (Woo & Waddington, 1990; Blersch & Kangas, 2014); and plankton that develops in the upstream beaver ponds (Czerniawski et al., 2017). So far, primarily beaver ponds have been studied to investigate the influence of beaver activities on invertebrate diversity (Harthun, 1998, 1999; Naiman et al., 1988; Willby et al., 2018). Additionally, studies that compare stream sections upstream and downstream of beaver territories are found quite frequently (Fuller & Peckarsky, 2011a, b; Redin & Sjöberg, 2013; Smith et al., 1991). There are several studies on invertebrates in beaver created wetland complexes (e.g. Naiman et al., 1984; Nummi, 1989; Hood & Larson, 2014; Bush et al., 2016, 2019; Nummi et al., 2021), including studies focusing on fish communities (e.g. Hägglund & Sjöberg, 1999; Bylak & Kukuła, 2018; Renik & Hafs, 2020; Bylak et al., 2014; Fritz & Gangloff, 2022). Thereby, beaver dams are generally not considered as movement barriers for fish species, especially for native species (Lokteff et al., 2013; Cutting et al., 2018; Pollock et al., 2022). It has been shown that physical dam features such as height, upstream location and the presence of side channels (Lokteff et al., 2013), as well as breach status (maintenance condition) and hydrological links (Cutting et al., 2018) determine the passability of beaver dams for fish species.
However, macroinvertebrates within beaver dams have only been studied by Clifford et al. (1993) and Rolauffs et al. (2001). Clifford et al. (1993) sampled the dams qualitatively using pond nets (500 and 150 µm), either by thoroughly disturbing the substrates so that organisms drifted into the net, or by removing individual parts of the dam and rinsing them off. A first quantitative study of beaver dams was carried out by Rolauffs et al. (2001), in which the beaver dams were sampled with emergence traps. In both studies, an invertebrate fauna typical for fast-flowing streams was detected. Reflecting habitat variability, the heterogeneity of species composition, abundance and biomass were significantly higher in beaver dams compared to free-flowing stream sections and beaver ponds (Rolauffs et al., 2001). The composition of functional feeding types of the free-flowing stream segments and beaver dams was very similar and included filter feeders, shredders, gatherers, grazers and predators (Clifford et al., 1993). The available studies, however, did not lead to generalisable patterns of benthic invertebrate communities in beaver dams. Rolauffs et al. (2001) focussed on insect emergence and thus did not consider hololimnic organisms. Rolauffs et al. (2001) and Clifford et al., (1993) investigated beaver dams without distinguishing the states of maintenance (maintained vs. abandoned).
Here, we comparatively studied maintained and abandoned beaver dams (Fig. 1) with the aim to comprehensively characterise their macroinvertebrate fauna in different zones (Fig. 2). Individually, we expected:

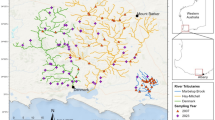
a Study area with the distribution of 48 mapped beaver dams, including the investigated dams (1–16). In the stream Thön, there are two territories, one near the mouth of the water reservoir (dams 4, 3, 2, 13); next further upstream (dams 12, 11, 10, 9). One territory is in the stream Weberbach (dams 1, 5, 6, 14, 15, 7, 16) and extends into the confluence of the Weiße Wehe stream (dam 8). Maintained dams are marked with a white circle and black outline, abandoned with a black cross. b Picture of a maintained beaver dam, spring 2018 in the stream Weberbach. c Picture of an Abandoned beaver dam, spring 2018 in the stream Thön

a Sample point distribution on a dam, showing sample number 1–8 (0.25 m2 each) at the streamside and sample number 9 consisting of four single samples that were pooled (1 m2) and measured dam dimensions, including the width and length of the top and the base of a dam and its height. b Sample combination for analysis, Combination 1, comparing top, middle and base areas of a dam, c sample Combination 2, comparing top, edge and middle areas of a dam
Hypothesis 1:
The fauna of maintained dams is spatially more variable than those of abandoned dams. If still in use, beavers maintain dam stability by adding and arranging branches, stones and mud, thus increasing habitat variability and flow diversity. In abandoned beaver dams, these differences dissolve, as fine sediment is washed out and therefore the variability from top to base diminishes (Woo & Waddington, 1990).
Hypothesis 2:
Consequently, we consider the fauna of abandoned beaver dams to be composed of more rheophilic species in all parts of the dam.
Hypothesis 3:
Following the descriptions of Rolauffs et al. (2001), we assume a flow gradient from the upper to the lower areas, so that the proportion of rheophilic species reaches its maximum at the base of a dam, where flow velocity is highest. Limnophilic or indifferent species are more likely to reside in the upper areas, especially of maintained dams, characterised by minimal flow velocities.
Hypothesis 4:
Concerning feeding types, the percentage of grazers and shredders is expected to be highest in the top area of a dam. Food for shredders (woody substrate, leaves, and wads of grass) is actively introduced by beavers or accumulates upstream of a dam, and biofilms (i.e. food for grazers) develop on the branches that are exposed to sunlight. Passive filter feeders are found exclusively in middle and lower areas of maintained dams, as a result of high flow velocity and high throughput of drifting food particles.
Hypothesis 5:
Terrestrial/semiaquatic species are found, occasionally, in the upper and marginal areas of beaver dams, especially if dams are still maintained. The marginal parts are in direct contact to the shore, and the upper parts of maintained dams are only infrequently overflowed, making these areas readily accessible to terrestrial or semiaquatic species.
Material and methods
Study area and study streams
The studied streams of 1st to 3rd order (Strahler, 1954) are part of the Inde-Rur-Maas catchment area (https://www.elwasweb.nrw.de) and inlets of a water reservoir (Wehebachtalsperre) in the Hürtgenwald (50° 44′ N, 6° 20′ E). This is a 120 km2 state forest (http://ww.wald-und-holz.nrw.de, 2022) in the western part of Germany and the federal state of North Rhine-Westphalia, adjacent to the Netherlands and Belgium, which belongs to the Rhenish Massif. The area, at an elevation of 240–450 m a.s.l., has a moderate atlantic climate with cool summers and mild winters. The average temperature in this area is 8.5 °C (https://cdc.dwd.de/portal), and the annual precipitation is 1062 mm (https://www.elwasweb.nrw.de). The area is characterised by a steep relief that facilitates the formation of a finely branched and dense network of streams (Sommerhäuser & Pottgiesser, 2002). Due to a reintroduction project from 1981 to 1989, the European beaver (C. fiber) has returned to this area (Naumann, 1991). In the subsequent decades, the beaver population increased and populated the wider surrounding. Overall population size in the federal state of North Rhine-Westphalia, of which the population originating from the Hürtgenwald is the main part, is estimated about > 1200 individuals or 0.2 beavers/km2 (Dalbeck, 2021). In the study area, the estimated average density is 4 dams/km (Dalbeck et al., 2014).
In winter 2017/2018, three streams with similar characteristics in terms of altitude, stream size and slope were selected and the dams within these streams were mapped. On this basis, 48 beaver dams in three territories were surveyed and classified into maintained and abandoned dams (Fig. 1). We characterised maintained beaver dams by freshly processed and installed branches, as well as an intact mud layer on the crest, which are both lacking in abandoned dams. We randomly selected in total eight maintained and eight abandoned dams for further examination (Fig. 1). The age of these dams ranged between 2 and 8 years. In our observations, even dams with the same age may vary in terms of morphology, substrate density, permeability and thus taxa composition. Therefore, we used a simple definition to distinguish two types of dams (regardless of the age) to generalise the effects of abandoned vs. maintained dams. All dams were intact (not breached) and were still impounding the stream with an upstream pond. The arrangement of the dams corresponds to a disjunct series (Kroes & Bason, 2015), which means that there was always a free-flowing section below the dams (≤ 2 m), so that no silt accumulated on this dam side. All territories are located in the middle reaches (rhithral) of the studied streams.
Study design
We measured height, width (Fig. 2a) and length of the 16 dams with a 50-m-long tape measure. We measured the total length of the dam, separating the “aquatic part” with contact to the watercourse and the “terrestrial part” connecting the dam with the shore. The terrestrial part becomes larger when the dam is abandoned and the impounding effect decreases.
Nine areas of each individual beaver dam (Fig. 2a) were sampled once in spring (March–April) 2018. The areas included the top of the dam adjacent to the pondside, as well as eight areas on the upper and lower sections on the streamside. We further refer to the samples as pondside or streamside samples, depending on their location of the dam, in reference to Rolauffs' (1999) designation. Sampling was conducted with a vacuum sampler, which was specifically designed for this purpose (Schloemer & Hoffmann, 2018). Sampling was standardised by area and time. Each sampling area was vacuumed for 3 min, while the area sampled differed between pondside and streamside samples. Only at the streamside, the dam has a clear three-dimensional structure with bulks and spikes, the cavities between which were vacuumed, while at the pondside, the dam has a smooth surface without cavities and was thus vacuumed only at the surface (Fig. 2a). Accordingly, we chose a smaller area per sample at the streamside as compared to the pondside samples. Each streamside sample accounted for 0.25 m2 (0.5 m * 0.5 m), while the single pondside sample accounted for 1 m2 (four individual areas of 0.25 m * 0.25 m at the top of a dam that were pooled together). Due to the highly variable morphology and surface of a dam, the dimensions of the sample areas were approximated.
Prior to sampling, depth, substrates, and gaps in the streamside sampling areas were examined by palpation. During the sampling time, all these gaps were sampled as deep as possible; if stones (≥ 10 cm) were detected and were loose enough, we removed and vacuumed them and put them back inside. The crest was vacuumed in a swabbing motion, and larger branches were removed, vacuumed and returned to prevent sampling pond substrate. The prime target of our sampling were aquatic taxa. Therefore, only gaps and parts with contact to water were sampled, with a focus on the inner structure of the beaver dams. However, the entire dam is in the transition zone of aquatic and terrestrial habitats, thus also offering habitats for terrestrial and semiaquatic organisms.
In total, 144 samples were taken (nine sampling areas on 16 dams). Every sample was filtered through a 500 µm sieve in the field and screened for species other than benthic invertebrates (e.g. fish). These were released back into the dam, while the rest of the sample was transferred into a container and preserved in 96% ethanol for further processing. In the laboratory, each sample was rinsed with water through a 2 mm mesh following the method of Meier et al. (2006) for macrozoobenthos collection and analyses, before the sample was divided into small amounts, which were ultimately transferred into a tray for sorting. Identification was on species level, except for most Diptera that were identified to genus or family level.
Data preparation and analysis
In a single sample originating from the streamside of a maintained dam, no specimens were detected. This we traced back to technical reasons and excluded the sample from the dataset. Accordingly, the total population of samples contained 71 samples for maintained dams (8 = pondside, 63 = streamside) and 72 samples for abandoned dams (8 = pondside, 64 = streamside). With the resulting table of the number of individuals per taxon and sample (ind./m2), we performed a taxonomic adjustment according to AQEM Consortium (2002) to ensure that taxonomic identification was consistent across instars. Furthermore, we removed all taxa that occurred just in one or two samples. To make sure we sampled invertebrate taxa adequately in each dam structure, we ran a species accumulation curve using the specaccum function of the vegan package (Oksanen et al., 2020) in Rstudio (R Core Team, 2021). The number of species and individuals in the different beaver dam areas and maintenance states was tested for significant differences with the Kruskal–Wallis test followed by the Dunn test (Holm-corrected) with the ggbetweenstats function of the package ggstatsplot (Patil, 2021) in Rstudio (R Core Team, 2021). With the same function, the mean values of the measured dam dimensions of abandoned and maintained dams were tested for significant differences with Welch’s t test.
We tested Hypothesis 1 (spatial variability of benthic invertebrate communities in maintained and abandoned dams) with the betadisper function of the vegan package (Oksanen et al., 2020) in Rstudio (R Core Team, 2021). The data was Hellinger transformed, and a Bray–Curtis dissimilarity matrix was created. In order to test the variability within the different dam habitats/categories, we performed a PCoA (betadisper) on the dissimilarity matrix. Since the samples had unequal sample size, we used the default adjustment option of the vegan package (Oksanen et al., 2020). The results of the PCoA were subjected to an ANOVA, calculating the between-group distance by the distances from the centroids, and were displayed with the effect-size Eta-squared. The means of the groups were compared with the TukeyHSD-test (“Honest Significant Difference” method) and confidence intervals at 95%. These calculations were also performed with the vegan package (Oksanen et al., 2020). The results are displayed in a biplot, showing the variability of the data and between each group. The distance between the centroids within each different group is displayed in boxplots with jittered points. Our null hypothesis is that there is no difference between the invertebrate communities of the dam categories. We used different sets of sample combinations for analysis: Combination 1 compared the top, the middle and the base of the dams (Fig. 2b), Combination 1 + m.s. (+ m.s. = including maintenance state) compared the same areas like Combination 1, but also regarding the maintenance state (maintained top, middle, base; abandoned top, middle, base).
Combination 2 compared the side/edge and the middle areas of the dams (Fig. 2c), Combination 2 + m.s. compared also side/edge and middle areas regarding the maintenance state (maintained edge, middle; abandoned edge, middle). Combination 3 compared the streamside to the pondside area (Combination 3 + m.s.: maintained stream-, pondside; abandoned stream-, pondside). Finally, we compared all samples of maintained with all samples from abandoned beaver dams (Combination 4).
To test Hypothesis 2 (rheophilic species are more abundant in abandoned dams), Hypothesis 3 (rheophilic species are more abundant at the dam’s basis) and Hypothesis 5 (terrestrial/semiaquatic species are more abundant in maintained dams), we calculated the percentages of species with different water velocity preferences (named current preferences in Schmidt-Kloiber & Hering, 2015) for the communities of the different dam areas. We used the tool “Perlodes-Online” (http://www.gewaesser-bewertung-berechnung.de) that is based on trait information compiled by Schmidt-Kloiber & Hering (2015); water velocity preferences were originally assembled by Schmedtje & Colling (1996). Hereby, a specific water velocity preference is assigned to every taxon and indicated by a numeric value. Terrestrial and semiaquatic taxa not included in Perlodes-Online were assigned to the additional category “terrestrial/semiaquatic”. We applied the same formula, which is implemented in “Perlodes-Online” and conducted the calculation in Excel to include the attribute “terrestrial/semiaquatic”. The formula is: \({P}_{VP}=\frac{\sum i ni}{N}*100\) (ni number of individuals of ith taxon with specific water velocity preference (PVP), N total abundance).
To test Hypothesis 4 (feeding types), we calculated the percentage of feeding types of the individual samples’ communities. Here, we also used the same method as the online tool “Perlodes-Online” but re-calculated in Excel to include the terrestrial/semiaquatic taxa. The formula is: \({P}_{ft}=\frac{{\sum }_{i} ft* ni}{N}*\frac{100}{10}\) (ft point value of the ith taxon of specific feeding type (Pft), ni number of individuals of ith taxon with specific feeding type, N total abundance). For each taxon, 10 points are distributed between feeding types corresponding to the taxon’s preferred diet. For the terrestrial/semiaquatic taxa, we added feeding types according to additional literature (Gepp, 2003; Speiser, 2001; van Stuivenberg, 1997). The percentage of feeding types and water velocity preferences in the different beaver dam areas and maintenance states was also tested for significant differences with the Kruskal–Wallis test followed by the Dunn test (Holm-corrected) with the ggbetweenstats function in Rstudio (Patil, 2021).
To calculate the preference of a taxon towards a specific dam category (Hypotheses 1–5), we computed the indicator value Index “IndVal” (Dufrêne & Legendre, 1997) using the multipatt function of the R package “indicspecies” (DeCáceres, 2020). The IndVal-Index is based on the mean abundance of a taxon as well as on its occurrence, and it is independent of the relative abundance of other taxa (Dufrêne & Legendre, 1997). The IndVal-Index ranges between 0 and 100 % and reaches its maximum when all individuals of a species are found in a single group of sites, and when the species occurs in all sites of that group (Dufrêne & Legendre, 1997). Results are expressed in value A—“specificity” (species occurrence within the different dam categories), and value B—“fidelity” (species occurrence within a specific dam category). The statistical significance of the association of a species with a certain dam category was tested with a permutation test, which is already default in the multipatt function (DeCáceres, 2020).
Results
Overview
On average, the studied beaver territories represented a pond cascade of 400 m length and consisted of 16 dams per colony (Fig. 1). Due to one territory that was completely abandoned (Fig. 1), 40% of the dams were in a “maintained” and 60% in an “abandoned” state, at the time of the study. On average, the distance between dams is 30 m (range: 10–36 m).
Beaver dam dimensions differ between the maintenance states (Table 1). The height, the length with contact to the water body (P-value = < 0.01) and the length of the dam base (P-value = < 0.01) was on average greater for maintained dams, while dam width (top and base: P-value = 0.02) and total length was on average greater for abandoned dams (Table 1).
We identified 57,716 individuals assigned to 130 taxa (Table S1) that were present in at least three sample sites. Additional 84 taxa with altogether 155 individuals (Table S1) were recorded in just one or two samples. The latter taxa were used to derive species numbers in abandoned and maintained dams, but excluded from the other analyses. 47 taxa exclusively occurred in abandoned dams, while 40 taxa exclusively occupied maintained dams (Table S1). In total, we detected an almost similar taxa richness in abandoned (174 taxa) and maintained beaver dams (167 taxa), while the overall number of individuals was higher in abandoned (33,417 individuals) than in maintained dams (24,454 individuals). The latter finding is related to the high abundance of the amphipod Gammarus fossarum Koch, 1835 in abandoned dams. The slope of species accumulation curves (Fig. S1) is approaching zero, indicating a sufficient sampling effort.
The median number of taxa differs between the pondside and streamside habitats of maintained and abandoned dams (Fig. S2a; Kruskal–Wallis test: 22.35, P-value = < 0.001). In abandoned dams, the numbers are higher (pondside = 28.5; streamside = 33) than in maintained beaver dams (pondside = 23.5; streamside = 24). When considering maintenance status and habitats, the numbers only differ significantly between the categories “streamside-abandoned” and “streamside-maintained” (Dunn test, Holm-corrected: P-value = 0.007) and between the categories “streamside-abandoned” and “pondside-maintained” (Dunn test, Holm-corrected: P-value < 0.001). The median abundance (ind./m2) of all four categories differ significantly (Fig. S2b; Kruskal–Wallis test: 33.03, P-value = < 0.001), and ranged between 387 ind./m2 (pondside-maintained) and 1584 ind./m2 (streamside-abandoned). Abundance differs most strongly between abandoned streamside areas and the pondside of maintained and abandoned dams (Dunn test, Holm-corrected: P-value = < 0.001).
Community analysis
Variance and diversity of the invertebrate community of the different habitats were further specified by a PCoA (betadisper), and an ANOVA (Table 2), alongside with the TukeyHSD-test (Table S2). The PCoA (Fig. 3a) shows the variability of the invertebrate communities of pondside and streamside samples of abandoned and maintained dams. The communities of each habitat are separate (Eta-squared = 0.08; P-value = < 0.009). Furthermore, the boxplot shows (Fig. 3b) that communities of maintained beaver dams are more diverse than those of abandoned beaver dams.

a PCoA shows the variability within and between each group of the pondside and streamside of abandoned and maintained dams “side and state” (Combination 3 + m.s.). b Boxplot with jittered points showing the distance between the centroids, within each different group, of the pondside and streamside of abandoned and maintained dams. po = pondside, str = streamside, ma = maintained, ab = abandoned. Illustration created with Rstudio (R core Team, 2021)
The maintenance state (Fig. S3a) most strongly impacts the diversity of the invertebrate community (Eta-squared = 0.09; P-value = < 0.001). The mean difference between the groups with the confidence intervals at 95% (TukeyHSD-test) is 0.087 for abandoned and maintained dams. Figures for all sample site combinations, next to “side and state” (Combination 3 + m.s., Fig. 3), are given in the supplementary material (Fig. S3a–f), as well as all results of the TukeyHSD-test (Table S2).
Water velocity preference
Rheophilic taxa represent the highest proportion of the community in all dam categories (Fig. 4) and range between a median of 48% (pondside-maintained) and 60% (streamside-abandoned). While the percentages differ between the four dam categories (Kruskal–Wallis test: X2 = 8.52, P-value = 0.04), the pairwise comparison resulted only in significant difference of the streamside of maintained and abandoned dams (P-value = 0.03). The second and third group with the highest percentage (Fig. 4) are indifferent taxa (Kruskal–Wallis test: X2 = 5.0, P-value = 0.17) and rheo-limnophilic (preferring slow water velocity) taxa (Kruskal–Wallis test: X2 = 3.77, P-value = 0.29). Despite these similarities, there are obvious differences between dam categories: The median percentages of rheobiont taxa, i.e. those only occurring in strong water velocity, differ the most between the four dam categories (Kruskal–Wallis test: X2 = 32.54, P-value = < 0.001). Rheobiont taxa occur almost exclusively in streamside habitats and have a higher percentage in maintained (5%) than in abandoned (3%) dams (Dunn test, Holm-corrected: P-value = 0.02). In addition, taxa preferring low water velocities, i.e. limno-rheophile taxa, barely occur (< 2%) with exception of the pondside of maintained dams (7%) (Kruskal–Wallis test: X2 = 17.80, P-value = < 0.001). Thus, pond- and streamside areas of maintained dams show the highest differences in water velocity preferences, from very low flow on top to very high flow in the middle and base areas of a dam (Fig. 4).

Percentage (%) of water velocity preferences of the community in the dam areas “side and state” (Combination 3 + m.s.). po = pondside, str = streamside, ma = maintained, ab = abandoned. Terr.-SemiAqu. = Terrestrial/Semiaquatic. Values below 1% were excluded; this concerns the preferences “limnophile” and “limnobiont”. Significant differences between categories are indicated with different letters. Boxplots show the median (middle line), quartiles (boxes), 1.5 × the interquartile range (whiskers) and extreme values (dots). Illustration created with Rstudio (R core Team, 2021)
Terrestrial/semiaquatic taxa occur almost exclusively in streamside habitats with an equally high median percentage in abandoned (2.5%) and maintained (2.2%) dams (Kruskal–Wallis test: X2 = 32.31, P-value = < 0.001). The differences between the streamside and the pondside categories are all significant (Dunn test, Holm-corrected: P-value = < 0.001). Occurring terrestrial or semiaquatic taxa include the beetle Dianous coerulescens (Gyllenhal, 1810) of the family Staphylinidae, the snail Vertigo antivertigo (Draparnaud, 1801) of the family Vertiginidae and the larvae of the Neuroptera Osmylus fulvicephalus (Scopoli, 1763) of the family Osmylidae (Table S1). Table S3 in the supplementary material contains detailed information on the water velocity preferences of the individual taxa.
Feeding types
Shredders are most abundant in all four dam categories (Fig. 5), but they are generally less frequently occurring in streamside habitats, in particular in those of maintained beaver dams (Kruskal–Wallis test: X2 = 33.07, P-value = < 0.001). Here, significant differences are detected between the streamside of maintained dams with every other category (Dunn test, Holm-corrected: streamside-abandoned P-value = 0.003; pondsides P-value = < 0.001). Passive filter feeders, which extract drifting material from the running water, prevail in the streamside of maintained dams (Fig. 5). Additionally, significant differences are detected between the streamside of maintained dams with every other category (Dunn test, Holm-corrected: P-value = < 0.001). In the streamside habitats of abandoned beaver dams, passive filter feeders are much less abundant, and they are almost absent in the pondside habitats of both maintained and abandoned beaver dams (Fig. 5). Overall, maintained streamside habitats seem to provide the most variable food sources leading to a balanced distribution of feeding type categories (Fig. 5). The significant differences of the median proportions of most feeding types clearly highlight the different nature of maintained-streamside habitats compared to the other three dam categories (Fig. 5). In contrast, there are no significant differences between the median proportions of feeding types between the pondside of maintained and abandoned dams (Fig. 5). Table S3 in the supplementary material contains detailed information of the feeding types of the individual taxa.

Percentage (%) of feeding types of the community in the dam areas “side and state” (Combination 3 + m.s.). po = pondside, str = streamside, ma = maintained, ab = abandoned. AFF active filter feeders, PFF passive filter feeders. The type “Others” combines xylophagous taxa, miners, parasites, other feeding types and not determined taxa. Significant differences between categories are indicated with different letters. Boxplots show the median (middle line), quartiles (boxes), 1.5 × the interquartile range (whiskers) and extreme values (dots). Illustration created with Rstudio (R core Team, 2021)
Indicator species
In total, 22 taxa were identified as indicators for the different dam categories (Table S4), which further characterise the respective habitat conditions. Indicators for the dam category abandoned-pondside (Table S4) include shredders like the Trichoptera Potamophylax cingulatus (Stephens, 1837) of the family Limnephilidae (IndVal = 82%, P-value = 0.001) and the mud-dwelling predator Sialis lutaria (Linnaeus, 1758) (Sialidae, Megaloptera) (IndVal = 46%, P-value = 0.044). Species characterising abandoned-streamside habitats include the two net-building Trichoptera larvae of the family Philopotamidae, Philopotamus montanus (Donovan, 1813) (IndVal = 68%, P-value = 0.01) and Wormaldia occipitalis (Pictet, 1834) (IndVal = 78%, P-value = 0.001), but also the wood-mining genus Lipsothrix (IndVal = 57%, P-value = 0.001) of the family Limoniidae (Diptera) and the riparian rove beetle Dianous coerulescens (Gyllenhal, 1810) (IndVal = 67%, P-value = 0.007). Maintained-pondside species include typical pond-dwellers, like the Gastropoda Radix balthica (Linnaeus, 1758) (IndVal = 58%, P-value = 0.022) of the family Limnaeidae and Physa fontinalis (Linnaeus, 1758) (IndVal = 47%, P-value = 0.031) of the family Physidae that graze on biofilms available on branches, but also shredders like various Trichoptera taxa (Table S4). Finally, taxa characterising the category maintained streamside exclusively include passive filter feeders preferring medium to strong flow velocities like Simuliidae larvae (e.g. Prosimulium sp.; IndVal = 77%, P-value = 0.004) and the Trichoptera Hydropsyche saxonica McLachlan, 1884 of the family Hydropsychidae (IndVal = 49%, P-value = 0.034).
Discussion
The invertebrate fauna of the studied beaver dams, regardless of the dam category, is typical for small streams of lower mountain areas. It is species-rich, with a high proportion of amphipods, Ephemeroptera and Trichoptera taxa, which prefer cool, well-oxygenated water. This confirms the findings of Rolauffs et al. (2001) and Clifford et al. (1993), who characterised beaver dams as lotic habitats with a rheophilic species community. Similar to the study of Rolauffs (1999), we discovered characteristic species of the headwater region (crenal), e.g. larvae of the caddisfly family Philopotamidae and Hydropsychidae (e.g. H. saxonica, Diplectrona felix McLachlan, 1878) were dominant. We even found rare and specialised headwater species, like the case bearing Tipulidae (Diptera) larvae Thaumastoptera calceata Mik, 1866, who is described as one of the most typical inhabitants of cold springs (Lenz, 1920) and the dragonfly larvae Cordulegaster boltonii Donovan, 1807 (Sternberg et al., 2000). This rheophilic community is common within the dams throughout the beaver territories, contrasting the communities typical of beaver ponds (Harthun, 1998, 1999; Naiman et al., 1988). Nevertheless, beaver dam communities have many peculiarities, which discriminate them from the general fauna of small mountain brooks. In particular, the high proportion of passive filter feeders is striking and most likely depending on the plankton community developing in the upstream beaver ponds in combination with the high flow velocities, especially in the streamside habitats. Clifford et al. (1993) also emphasised the high amount of passive filter feeders in beaver dams and characterised especially Simuliidae larvae as a typical fauna element of this habitat. Additionally, the high spatial discrimination of invertebrate communities renders the beaver dam a unique habitat within the framework of small mountain brooks.
Our results differ to those obtained by Rolauffs et al. (2001) and Clifford et al. (1993) in some regards, which is mainly rooted in the sampling methods applied. Rolauffs et al. (2001), who investigated one of the streams also addressed in our study, used emergence traps and thus missed hololimic taxa such as Gammarus, which is the most abundant taxon in the beaver dams. On the other hand, we may have missed several of the smaller Diptera taxa (larvae < 2 mm), which were dominant in the two other studies. This is particularly true for midge larvae (Chironomidae), who mainly emerged in summer according to Rolauffs et al. (2001) and might just have been too small when the sampling for the current study took place. Therefore, our sampling does not represent a full inventory of the dam fauna, but a spot check of the organisms ≥ 2 mm colonising the dam in spring, before emergence, at times when larval size of most merolimnic species is maximal.
Community analysis (Hypothesis 1)
The first hypothesis that the fauna of maintained dams is spatially more variable than that of abandoned dams, has been confirmed. The PCoA and ANOVA alongside the TukeyHSD-test clearly reveal the larger variability of the fauna of maintained dams, especially of streamside habitats (but regard that sample size was lower in pondside habitats). The analysis also confirms that the maintenance state is the greatest influencing factor on community composition. In addition, the distribution of water flow preferences and feeding types supports the conjecture that the habitat conditions are more variable in maintained dams, most likely due to the maintenance activity of beavers. Maintenance activities are required to prevent the flow from washing away dam material and weaken the whole structure (Woo & Waddington, 1990). Abandoned dams therefore become more permeable, allowing water to pass readily through the entire structure (Woo & Waddington, 1990); as a consequence, the extremes of both lentic and lotic habitats get lost. The effects on invertebrate communities are obvious, already on the level of coarse taxonomic entities. For example, one difference between maintained and abandoned dams is the high proportion of amphipods (consisting almost exclusively of the species G. fossarum). This highly mobile species accumulates primarily in dams with a species-specific food supply, e.g. rotting wood. We sampled in spring, at times when the availability of leaves in the stream is minimal—this might be an additional reason why G. fossarum accumulates in beaver dams. Abandoned dams are obviously more accessible to the species, as the number of gaps is higher, and more wood becomes available. Also, other hololimnic species are more abundant in the abandoned dams, e.g. the less mobile snail Ancylus fluviatilis Müller, 1774. It feeds on algal layers on stones and wood in fast-running streams (Arens, 1990) and was frequently observed on stones inside the dams (Table S3).
Water velocity preference (Hypotheses 2 and 3)
According to Hypothesis 2, we expected the fauna of abandoned beaver dams to be more rheophilic, as these dams are no longer repaired, and the water can flow more rapidly through the remaining structure. However, this was not the case. Regardless of the maintenance state, taxa depending on lotic conditions account for at least two-thirds of the invertebrate community in all dam categories. In the streamside of maintained dams, the percentage of taxa depending on high flow velocities is even greater than in abandoned dams. Also, the indicators for the category maintained streamside are exclusively specialists for fast-flowing conditions (Simulidae, Hydropsychidae). In particular, the high abundance of Simuliidae characterise the streamside of maintained dams as fast-flowing area. According to Harrod (1965), the larvae of Simulium ornatum (Meigen, 1818) requires a velocity of > 0.19 m/s to hold the cephalic fans open in the water. Feeding of Simulium larvae almost entirely depends on their cephalic fans (Zahar, 1951). In addition, Zahar (1951) states that the filtration efficiency of Simulium larvae increases with flow velocity, as more water can be filtered within a given period of time, while at the same time losses due to sinking of drifting material are prevented. In addition to flow velocity and food conditions, beaver dams provide excellent attachment substrates. In lotic systems, many sedentary or sessile filter feeders rely on solid substrates in local hydraulic environments, which guarantee a sufficient delivery of suspended materials (Hoffmann & Hering, 2000). The indicator species H. saxonica most likely needs similar conditions, which are provided by the streamside of maintained dams. Furthermore, Hydropsyche larvae bite out large depressions in wood structure, into which they construct the retreat portion of their shelter (Hoffmann & Hering, 2000). In the category “streamside-abandoned”, the indicator taxa mentioned before are replaced by the net-spinning caddis larvae W. occipitalis and P. montanus. These species also use coarse woody debris for net attachment and retreat building (Hoffmann & Hering, 2000); however, the nets of Philopotamid larvae are long tubular bags made out of very fine rectangular meshwork (Edington, 1968). These nets are common in places where water is percolating through piles of boulders or woody structures and where nets are hanging in a thin film of waters, in order to filter diatoms and fine detritus particles (Edington, 1968). Such trickle areas must therefore frequently exist in the streamside of abandoned dams. We assume that Philopotamid nets are generally occurring in less strong current than those of Hydropsyche and Simulium larvae and thus the conditions in abandoned dams are preferable. Also, the predacious beetle D. coerulescens, another indicator taxon of streamside-abandoned, requires trickle or hygropetric areas. There is little information about the hydraulic conditions in a beaver dam; experimental investigations, by Hart et al. (2020) could show that the discharge increases with porosity, which is congruent with our Hypothesis. The studied dams are made of wood, a few stones and little sediment, and this material composition leads to a comparatively high permeability (Ronnquist & Westbrook, 2021) regardless of the maintenance state. The diameter and the number of the pores/gaps are smaller and rarer in maintained dams, in the manner of our observations. According to the equation of continuity (“small diameter—faster flow velocity”), this would result in higher flow velocities within maintained dams. Referring to the work of Müller and Watling (2016), the hydraulic condition in a beaver dam is best described by Darcy’s law, where flow velocity is a linear function of head difference. For a 5-cm increase in head, the flow velocity increases by 0.05 m/s (Müller & Watling, 2016). In our study, maintained dams are on average 0.2 m higher than the abandoned ones, so assuming the relationship suggested by Müller and Watling (2016) flow velocity would on average increase by 0.2 m/s. Beaver dams primarily create lentic water bodies (beaver ponds), but are themselves a lotic water habitat supporting a lotic fauna. Our results revealed that the maintenance state has a direct influence on flow velocity. In combination with dam area, this results in a complex water flow pattern that leads to a variable dam colonisation with invertebrates.
According to Hypothesis 3, we expected a higher proportion of rheophilic species in the streamside of dams, where flow velocity is assumed to be highest, while limnophilic or indifferent species were expected to prefer the pondside, especially of maintained dams, which is characterised by minimal flow velocities. This hypothesis was supported. Community composition reveals that flow velocity increases from the pondside to the streamside. This gradient is most pronounced in maintained dams and dissipates in abandoned ones. Similar results were obtained by Rolauffs et al. (2001), whose measurements showed that flow velocity was lowest in the pondside (max. 0.2 m/s) and highest in the streamside area (on average: 0.4 m/s; max. 1.0 m/s). Accordingly, indicator taxa for the pondside include several lentic taxa, e.g. caddisflies of the family Limnephilidae like Limnephilus lunatus Curtis, 1834, but also lotic taxa such as Halesus radiatus (Curtis, 1834), H. digitatus/tesselatus and Potamophylax cingulatus (Stephens, 1837).
Feeding types (Hypothesis 4)
With Hypothesis 4, we expected the proportion of grazers and shredders to be highest in the upper areas of a dam, where food supply and flow velocity is consistent with their needs. This hypothesis was supported. Almost half of the invertebrate fauna of the category maintained-pondside is composed of shredders, with grazers comprising an additional 13%. The share of shredders and grazers in the other dam categories are lower. Furthermore, the indicator taxa of the category maintained-pondside include the snails R. balthica and P. fontinalis as well as the mayfly Baetis rhodani (Pictet 1843) of the family Baetidae that all feed on biofilms (Arens, 1920). Similar to Blersch & Kangas (2014), we have observed that the top of maintained dams is passively extended by washed up remains of the beavers’ gnawing activities or branches that have fallen into the stream. The former lead to the accumulation of smaller and debarked branches, whose smooth surface is often covered with an epixylic biofilm, which may be several millimetres thick. The crest of the beaver dam is usually fully exposed to the sun and the water of the beaver ponds is most likely nutrient-rich; therefore, the conditions for biofilm growth are ideal. In addition, the beavers repair the crest with muds of grass, floating leaves accumulate along the crest and of course the crest is also composed of woody structures; all these provide food to shredders. Accordingly, several of the indicator taxa are shredding species, e.g. representatives of the tribes Chaetopterygini and Stenophylacini and P. cingulatus. In general, shredders and gatherers were the main feeding types in beaver dams, with an exceptionally high proportion of passive filter feeders in the streamside, as mentioned earlier.
Terrestrial/semiaquatic taxa (Hypothesis 5)
Finally, we expected terrestrial or semiaquatic species predominantly in the upper and marginal areas of beaver dams, especially if dams are still maintained (Hypothesis 5). However, the contrary was the case, as the observed terrestrial/semiaquatic species predominantly occur in the streamside of the dams, especially of abandoned dams. As already outlined for Hypothesis 2, abandoned dams provide thin layers of water on woody structures, a habitat, which may enable the persistence of species using atmospheric oxygen, as the water depth is minimal. At the same time, prey availability is huge and mobile species, such as the beetle D. coerulescens, can access large parts of the dam, which is characterised by holes and gaps, when searching for prey. D. coerulescens is a typical species of the fauna hygropetrica which is linked to the splash zone of waterfalls and banks of fast-flowing streams with overflowed mosses and other vegetation (Koch, 1989; van Stuivenberg, 1997). The species was already recorded in a beaver dam in Bavaria (Schloemer & Hoffmann, 2018). V. antivertigo is described as a typical swamp inhabitant, living at the shore of waterbodies and in marshy meadows (Wiese, 2016). The larvae of O. fulvicephalus is another element of the riparian fauna that also likes to reside under water part-time (Moog, 2019). For such species living between aquatic and terrestrial conditions, beaver dams can provide ideal conditions.
Conclusions
Beaver dams provide a wide variety of environmental conditions and habitat types. Within a very small area, conditions can reach from lentic to lotic situations and microhabitats are available for species with all types of flow preferences and feeding types. Beaver dams undergo a distinct succession, especially when they are abandoned, which further enhance the variability of habitat types of a dam. Beaver dams in their various maintenance and decay states provide valuable habitats for a wide variety of organisms in small floodplains. Our results clearly outline that beaver dams are an important component of the numerous habitats created by beavers that increase invertebrate diversity in streams and adjacent floodplains. Therefore, the impact of beaver activities on biodiversity is underestimated, if the fauna of beaver dams is not taken into account.
Data availability
Data are available from the corresponding author upon reasonable request.
References
Andersen, D. C. & P. B. Shafroth, 2010. Beaver dams, hydrological thresholds, and controlled floods as a management tool in a desert riverine ecosystem, Bill Williams River, Arizona. Ecohydrology 3: 325–338. https://doi.org/10.1002/eco.113.
AQEM Consortium, 2002. Manual for the application of the AQEM system: a comprehensive method to assess European streams using benthic macroinvertebrates, developed for the purpose of the Water Framework Directive (No. Version 1.0). http://www.aqem.de.
Arens, W., 1990. Wear and tear of mouthparts: a critical problem in stream animals feeding on epilithic algae. Canadian Journal of Zoology 68: 1896–1914. https://doi.org/10.1139/z90-269.
Barnes, D. M. & A. U. Mallik, 1996. Use of woody plants in construction of beaver dams in northern Ontario. Canadian Journal of Zoology 9: 1781–1786. https://doi.org/10.1139/z96-197.
Blersch, D. M. & P. C. Kangas, 2014. Signatures of self-assembly in size distributions of wood members in dam structures of Castor canadensis. Global Ecology and Conservation 2: 204–213. https://doi.org/10.1016/j.gecco.2014.08.011.
Bush, B. M., S. A. Wissinger, D. Batzer, & D. Boix, 2016. Invertebrates in freshwater wetlands invertebrates in beaver-created wetlands and ponds. Springer, International Publishing Cham 411–449. https://doi.org/10.1007/978-3-319-24978-0_12
Bush, B., C. Stenert, L. Maltchik & D. Batzer, 2019. Beaver-created successional gradients increase ß-diversity of invertebrates by turnover in stream-wetland complexes. Freshwater Biology 64: 1265–1274. https://doi.org/10.1111/fwb.13302.
Butler, D. R. & G. P. Malanson, 1995. Sedimentation rates and patterns in beaver ponds in a mountain environment. Geomorphology 13: 255–269. https://doi.org/10.1016/0169-555X(95)00031-Y.
Bylak, A. & K. Kukuła, 2018. Living with an engineer: fish metacommunities in dynamic patchy environments. Marine and Freshwater 69: 883–893. https://doi.org/10.1071/MF17255.
Bylak, A., K. Kukuła & J. Mitka, 2014. Beaver impact on stream fish life histories: the role of landscape and local attributes. Canadian Journal of Fisheries and Aquatic Sciences 71: 1603–1615. https://doi.org/10.1139/cjfas-2014-0105.
Collen, P. & R. J. Gibson, 2001. The general ecology of beavers (Castor spp.), as related to their influence on stream ecosystems and riparian habitats, and the subsequent effects. Reviews in Fish Biology and Fisheries 10: 439–461.
Clifford, H. F., G. M. Wiley & R. J. Casey, 1993. Macroinvertebrates of a beaver-altered boreal stream of Alberta, Canada, with special reference to the fauna on the dams. Canadian Journal of Zoology 71: 1439–1447. https://doi.org/10.1139/z93-199.
Cutting, K., J. Ferguson, M. Anderson, K. Cook, S. Davis & R. Levine, 2018. Linking beaver dam affected flow dynamics to upstream passage of Arctic grayling. Ecology and Evolution 8: 12905–12917. https://doi.org/10.1002/ece3.4728.
Czerniawski, R., L. Sługocki & M. Kowalska-Góralska, 2017. Effects of beaver dams on the zooplankton assemblages in four temperate lowland streams (NW Poland). Biologia 4: 417–430. https://doi.org/10.1515/biolog-2017-0047.
Dalbeck, L., 2021. Bestandsentwicklung des Bibers am Beispiel des Kreises Düren. In Wald und Holz, N. R. W. (ed.), 40 Jahre Biber-Wiederansiedlung in Nordrhein-Westfalen: 14–17. Retrieved from http://www.wald-und-holz.nrw.de/.
Dalbeck, L., J. Janssen & L. S. Völsgen, 2014. Beavers (Castor fiber) increase habitat availability, heterogeneity and connectivity for common frogs (Rana temporaria). Amphibia-Reptilia 35: 321–329. https://doi.org/10.1163/15685381-00002956.
Danilov, P., V. Kanshiev & F. Fyodorov, 2011. Characteristics of North American and Eurasian beaver ecology in Karelia. In Sjöberg, G. & J. P. Ball (eds), Restoring the European beaver 2011: 50 Years of Experience. Section 2—Biology of two beaver species in Europe. Chapter 6: 55–72. Sofia. Pensoft.
DeCáceres, M., 2020. Package 'indicspecies'. Retrieved from http://cran.r-project.org/web/packages/indicspecies/vignettes/indicspecies.
Dufrêne, M. & P. Legendre, 1997. Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs 67: 345–366.
Dugmore, A. R., 1914. The Romance of the Beaver: Being the History of the Beaver in the Western Hemisphere, JB Lippincott, London:
Edington, J. M., 1968. Habitat preferences in net-spinning caddis larvae with special reference to the influence of water velocity. The Journal of Animal Ecology 37: 1–675. https://doi.org/10.2307/3081.
Fritz, S. & M. Gangloff, 2022. The effects of beaver impoundments on montane stream fish communities. Aquatic Conservation: Marine and Freshwater Ecosystems 32: 1618–1633. https://doi.org/10.1002/aqu.3876.
Fuller, M. R. & B. L. Peckarsky, 2011a. Does the morphology of beaver ponds alter downstream ecosystems? Hydrobiologia 1: 35–48. https://doi.org/10.1007/s10750-011-0611-x.
Fuller, M. R. & B. L. Peckarsky, 2011b. Ecosystem engineering by beavers affects mayfly life histories. Freshwater Biology 5: 969–979. https://doi.org/10.1111/j.1365-2427.2010.02548.x.
Gepp, J., 2003. Der Bachhaft Osmylus fulvicephalus—240 Jahre nach seiner Beschreibung durch Johannes Antonius Scopoli—Österreichs Insekt des Jahres (Osmylidae, Neuroptera). Carinthia II 193(113): 325–334.
Gurnell, A. M., 1998. The hydrogeomorphological effects of beaver dam-building activity. Progress in Physical Geography 22: 167–189. https://doi.org/10.1177/030913339802200202.
Hafen, K. C., J. M. Wheaton, B. B. Roper, P. Bailey & N. Bouwes, 2020. Influence of topographic, geomorphic, and hydrologic variables on beaver dam height and persistence in the intermountain western United States. Earth Surface Processes and Landforms 11: 2664–2674. https://doi.org/10.1002/esp.4921.
Halley, D. J., A. P. Saveljev & F. Rosell, 2021. Population and distribution of beavers Castor fiber and Castor canadensis in Eurasia. Mammal Review 51(1): 1–24. https://doi.org/10.1111/mam.12216.
Harrod, J. J., 1965. Effect of current speed on the cephalic fans of the larva of Simulium ornatum var. nitidifrons Edwards (Diptera: Simuliidae). Hydrobiologia 26: 8–12. https://doi.org/10.1007/BF00142248.
Hart, J., M. Rubinato & T. Lavers, 2020. An experimental investigation of the hydraulics and pollutant dispersion characteristics of a model beaver dam. Water 9: 1–22. https://doi.org/10.3390/w12092320.
Harthun, M., 1998. Biber als Landschaftsgestalter—Einfluß des Bibers (Castor fiber albicus Matschie, 1907) auf die Lebensgemeinschaft von Mittelgebirgsbächen. 1–199. Schriftenreihe der Horst-Rohde-Stiftung. Maecenata. München.
Harthun, M., 1999. Der Einfluss des Bibers (Castor fiber albicus) auf die Fauna (Odonata, Mollusca, Trichoptera, Ephemeroptera, Diptera) von Mittelgebirgsbächen in Hessen (Deutschland). Limnologica 29: 449–464. https://doi.org/10.1016/S0075-9511(99)80052-8.
Hägglund, A. & G. Sjöberg, 1999. Effects of beaver dams on the fish fauna of forest streams. Forest Ecology and Management 115: 259–266. https://doi.org/10.1016/S0378-1127(98)00404-6.
Hoffmann, A. & D. Hering, 2000. Wood-associated macroinvertebrate fauna in central european streams. International Review Hydrobiology 85: 25–48. https://doi.org/10.1002/(SICI)1522-2632(200003)85:1%3c25::AID-IROH25%3e3.0.CO;2-R.
Hood, G. A. & D. G. Larson, 2014. Beaver-created habitat heterogeneity influences aquatic invertebrate assemblages in boreal Canada. Wetlands 34: 19–29. https://doi.org/10.1007/s13157-013-0476-z.
Johnston, C., 2017. Beavers: Boreal Ecosystem Engineers, Springer, Cham:, 1–272. https://doi.org/10.1007/978-3-319-61533-2.
Koch, K., 1989. Die Käfer Mitteleuropas, Goecke & Evers, Krefeld:
Kroes, D. E. & C. W. Bason, 2015. Sediment-trapping by beaver ponds in streams of the Mid-Atlantic Piedmont and Coastal Plain, USA. Southeastern Naturalist 14: 577–595.
Lenz, F., 1920. Thaumastoptera calceata Mik. Eine Gehäusetragende Tipulidenlarve. Archiv Für Naturgeschichte 85: 114–136.
Lokteff, R., B. Roper & J. Wheaton, 2013. Do beaver dams impede the movement of trout? Transactions of the American Fisheries Society 142: 1114–1125. https://doi.org/10.1080/00028487.2013.797497.
Meier, C., P. Haase, P. Rolauffs, K. Schindehütte, F. Schöll, A. Sundermann & D. Hering, 2006. Methodisches Handbuch Fließgewässerbewertung zur Untersuchung und Bewertung von Fließgewässern auf der Basis des Makrozoobenthos vor dem Hintergrund der EG-Wasserrahmenrichtlinie. http://www.fliessgewaesserbewertung.de.
McComb, W. C., J. R. Sedell & T. D. Buchholz, 1990. Dam-site selection by beavers in an eastern Oregon basin. Great Basin Naturalist 50: 273–281.
McDowell, D. M. & R. J. Naiman, 1986. Structure and function of a benthic invertebrate stream community as influenced by beaver (Castor canadensis). Oecologia 68: 481–489.
Moog, O., 2019. Der Europäische Bachhaft—ein Ufer-Insekt mit Höhlenaffinität? Osmylus fulvicephalus—a shore insect with cave affinity? Höhlenkundliche Mitteilungen Des Landesvereins Für Höhlenkunde in Wien Und Niederösterreich. 5–6: 94–95.
Müller, G., 2014. Ingenieurtechnische Aspekte der Biberdämme. KW Korrespondenz Wasserwirtschaft 7: 158–163.
Müller, G. & J. Watling, 2016. The engineering in beaver dams. In Constantinescu, G., M. Garcia & D. Hanes (eds), A Balkema book, River flow 2016: Proceedings of the International Conference on Fluvial Hydraulics (River Flow 2016). St. Louis, USA. 11–14 July 2016 (pp. 2094–2099). CRC Press, Boca Raton. https://doi.org/10.1201/9781315644479-326.
Müller-Schwarze, D., 2011. The Beaver: Its Life and Impact, 2nd ed. Cornell University Press, Ithaca: 216.
Naiman, R. J., D. M. McDowell & B. Farr, 1984. The influence of beaver (Castor canadensis) on the production dynamics of aquatic insects. Internationale Vereinigung Für Theoretische Und Angewandte Limnologie: Verhandlungen 22: 1801–1810. https://doi.org/10.1080/03680770.1983.11897578.
Naiman, R. J., C. A. Johnston & J. C. Kelley, 1988. Alteration of North American Streams by Beaver. BioScience 11: 753–762. https://doi.org/10.2307/1310784.
Naumann, G., 1991. Aussetzen von Bibern in der Eifel—Ein Beispiel für die Problematik von Wiedereinbürgerungen. Naturschutz Im Rheinland, Rheinischer Verein Für Denkmalpflege Und Landschaftsschutz, Jahrbuch 1989–1991: 137–150.
Nummi, P., 1989. Simulated effects of the beaver on vegetation, invertebrates and ducks. Annales Zoologici Fennici 26: 43–52.
Nummi, P., W. Liao, J. van der Schoor & J. Loehr, 2021. Beaver creates early successional hotspots for water beetles. Biodiversity and Conservation 30: 2655–2670. https://doi.org/10.1007/s10531-021-02213-8.
Oksanen, J., F. G. Blanchet, M. Friendly, R. Kindt, P. Legendre, D. McGlinn, P. R. Minchin, R. B. O’Hara, G. L. Simpson, P. Solymos, M. H. Stevens, E. Szoecs & H. Wagner, 2020. Package ‘vegan’. Community Ecology Package. (Version R package version 2.5–7.) Cran.R-project.org: Cran.R-project.org. https://CRAN.R-project.org/package=vegan.
Patil, I., 2021. Visualizations with statistical details: the ggstatsplot approach. Journal of Open Source Software 6(61): 3167. https://doi.org/10.21105/joss.03167.
Pollock, M., S. Witmore & E. Yokel, 2022. Field experiments to assess passage of juvenile salmonids across beaver dams during low flow conditions in a tributary to the Klamath River, California, USA. PLoS ONE 17: 1–24. https://doi.org/10.1371/journal.pone.0268088.
R Core Team, 2021. R: A language and environment for statistical computing. Freeware. R Foundation for statistical computing. Vienna, Austria. http://www.R-project.org/.
Redin, A. & G. Sjöberg, 2013. Effects of Beaver Dams on Invertebrate Drift in Forest Streams. Sumarski List 11–12: 597–607.
Renik, K. M. & A. W. Hafs, 2020. Effect of Beaver on Brook Trout Habitat in North Shore, Lake Superior, Streams. North American Journal of Fisheries Management 40: 427–445. https://doi.org/10.1002/nafm.10422.
Richard, P. B., 1967. Le déterminisme de la construction des barrages chez le Castor du Rhône. Revue D`Ecologie, Terre Et Vie, Société Nationale De Protection De La Nature: 339–470.
Rolauffs, P., 1999. Untersuchungen zur Besiedlung von Biberdämmen durch Insekten unter besonderer Berücksichtigung der Emergenz. Diploma Thesis: 1–160. Faculty of Biology, University Essen.
Rolauffs, P., D. Hering & S. Lohse, 2001. Composition, invertebrate community and productivity of a beaver dam in comparison to other stream habitat types. Hydrobiologia 459: 201–212.
Ronnquist, A. L. & C. J. Westbrook, 2021. Beaver dams: how structure, flow state, and landscape setting regulate water storage and release. The Science of the Total Environment 785: 1–12. https://doi.org/10.1016/j.scitotenv.2021.147333.
Schloemer, S. & A. Hoffmann, 2018. The Beaver Dam Vacuum Sampler—a new sampling method to investigate the invertebrate fauna in beaver dams. Lauterbornia 85: 117–121.
Schmedtje, U. & M. Colling, 1996. Ökologische Typisierung der aquatischen Makrofauna (Informationsberichte des Bayerischen Landesamtes für Wasserwirtschaft No. 4). München. https://www.gewaesserbewertung-berechnung.de/files/downloads/perlodes/PerlodesOnline_Dokumentation_Teil_III_Beschreibung_Indizes.pdf.
Schmidt-Kloiber, A. & D. Hering, 2015. www.freshwaterecology.info—an online tool that unifies, standardises and codifies more than 20,000 European freshwater organisms and their ecological preferences. Ecological Indicators 53: 271–282. https://doi.org/10.1016/j.ecolind.2015.02.007.
Smith, M. E., C. T. Driscoll, B. J. Wyskoski, C. M. Brooks & C. C. Cosentini, 1991. Modification of stream ecosystem structure and function by beaver (Castor canadensis) in the Adirondack Mountains, New York. Canadian Journal of Zoology 69: 55–61. https://doi.org/10.1139/z91-009.
Sommerhäuser, M. & T. Pottgiesser, 2002. Fließgewässertypenatlas Nordrhein-Westfalens (Merkblätter Nr. 36). Essen.
Speiser, B., 2001. Food and Feeding Behaviour. In Barker G. M. (ed), The biology of terrestrial molluscs: 259–288. CABI Pub., Wallingford, Oxon, UK, New York, NY, USA.
Sternberg, K., R. Buchwald & U. Stephan, 2000. Cordulegaster boltonii. In: Sternberg, K. & R. Buchwald (eds), Die Libellen Baden-Würtembergs—Band 2: Großlibellen (Anisoptera): 191–208. Verlag Eugen Ulmer
Strahler, A. N., 1954. Statistical analysis in geomorphic research. The Journal of Geology 62: 1–25. https://doi.org/10.1086/626131.
Thie, J., 2022. Exploring Beaver Habitat in Canada: The Longest Beaver Dam in the World. https://www.geostrategis.com/p_beaver-longestdam.htm.
Townsend, J. E., 1953. Beaver ecology in western montana with special reference to movements. Journal of Mammalogy 34: 459. https://doi.org/10.2307/1375861.
Van Stuivenberg, F., 1997. Tabel en verspreidingsatlas van de Nederlandse Steninae (Coleoptera: Staphylinidae). Nederlandse Faunistische Mededelingen 6: 3–60.
Warren, E. R., 1932. Position of wood in beaver dams. Science (new York, N.y.) 75(1937): 194–195. https://doi.org/10.1126/science.75.1937.194-b.
Wiese, V., 2016. Die Landschnecken Deutschlands: Finden—erkennen—bestimmen (2., durchgesehene Auflage). Quelle & Meyer Bestimmungsbücher. Quelle & Meyer Verlag, Wiebelsheim.
Willby, N. J., A. Law, O. Levanoni, G. Foster & F. Ecke, 2018. Rewilding wetlands: beaver as agents of within-habitat heterogeneity and the responses of contrasting biota. Philosophical Transactions of the Royal Society, Series B, Biological Sciences 373: 1–8. https://doi.org/10.1098/rstb.2017.0444.
Wohl, E., D. N. Scott, S. E. Yochum, 2019. Managing for large wood and Beaver dams in stream corridors. General Technical Report-Rocky Mountain Research Station. USDA Forest Service, (RMRS-GTR-404):136.
Woo, M.-K. & J. M. Waddington, 1990. Effects of beaver dams on subarctic wetland hydrology. Arctic 43: 223–230.
Wróbel, M., 2020. Population of Eurasian beaver (Castor fiber) in Europe. Global Ecology and Conservation 23: 1–4. https://doi.org/10.1016/j.gecco.2020.e01046.
Zahar, A. R., 1951. The ecology and distribution of black-flies (Simuliidae) in South-East Scotland. The Journal of Animal Ecology 20: 33. https://doi.org/10.2307/1643.
Zavyalov, N. A., 2014. Beavers (castor fiber and castor canadensis), the founders of habitats and phytophages. Biology Bulletin Reviews 4: 157–180. https://doi.org/10.1134/S207908641402008X.
Acknowledgements
Special thanks to Willem Kaijser for his support in the data analysis and to Peter Rolauffs for the assistance in the identification of Ephemeroptera, William Y. Wrausmann, for assistance in the identification of Trichoptera, and the Aquatic Ecosystem Research Group of the University Duisburg-Essen for identifying “difficult species” via DNA Barcoding. Thanks for support in the field to Nicolaus Seefeld, Svenja Pfeifer and the Biologische Station im Kreis Düren e.V., especially Lutz Dalbeck. Many thanks to two anonymous reviewers, whose comments have greatly improved this article.
Funding
Open Access funding enabled and organized by Projekt DEAL. The authors have not disclosed any funding.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors did not receive support from any organisation for the submitted work and have no relevant financial or non-financial interest to disclose.
Additional information
Handling editor: Verónica Ferreira
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Schloemer, S., Hörren, T., Lorenz, A.W. et al. The macroinvertebrate fauna of maintained and abandoned beaver dams. Hydrobiologia 850, 1763–1778 (2023). https://doi.org/10.1007/s10750-023-05176-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-023-05176-9