
Abstract
Drained peatlands emit large amounts of greenhouse gases and cause downstream nutrient pollution. Rewetting aids in restoring carbon storage and sustaining unique biodiversity. However, rewetting for nature restoration is socio-economically not always feasible. Cultivation of Sphagnum biomass after rewetting allows agricultural production. In the short term, Sphagnum paludiculture is productive without fertilization but it remains unclear whether it sustains its functionality in the longer-term. We studied nutrient dynamics, organic matter build-up, and carbon and nutrient accumulation at a 16-ha Sphagnum paludiculture area in NW-Germany. Site preparation included topsoil removal and inoculation with Sphagnum and it was rewetted five and ten years ago and managed with mowing, irrigation, and ditch cleaning. The unfertilized sites were irrigated with (compared to bog conditions) nutrient-rich surface water and exposed to atmospheric nitrogen deposition of 21 kg N/ha/yr. Our data reveal that ten years of Sphagnum growth resulted in a new 30 cm thick organic layer, sequestering 2,600 kg carbon, 56 kg nitrogen, 3.2 kg phosphorus, and 9.0 kg potassium per ha/yr. Porewater nutrient concentrations were low and remained stable over time in the top layer, while ammonium concentrations decreased from 400–700 to 0–50 µmol/L in the peat profile over 10 years. Hydro-climatic fluctuations most likely caused the variation in ammonium in the top layer. We conclude that Sphagnum paludiculture enables rapid carbon and nutrient accumulation without active fertilization provided the biomass is not harvested, and provides perspective for bog restoration on agricultural peatlands. Large-scale application of Sphagnum paludiculture may mitigate environmental issues of unsustainable peatland-use.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Peatlands accumulate organic material under wet, anoxic conditions over decades to millennia (Joosten and Clarke 2002; Yu et al. 2010). While peatlands cover merely 3% of the land surface, they store over 30% of the world’s soil organic carbon (C) (Gorham 1991; Leifeld and Menichetti 2018; Temmink et al. 2022). Furthermore, they play a crucial role in nutrient storage and cycling, freshwater purification and retention, and habitat provision for unique biodiversity (Zedler and Kercher 2005; Lamers et al. 2015; Jurasinski et al. 2020). Consequently, draining peatlands and peat extraction result in large C emissions, land subsidence, nutrient release, and loss of unique biodiversity (Hooijer et al. 2012; Turetsky et al. 2015; Ahmad et al. 2020; Bianchi et al. 2021). The Paris agreement targets net-zero C emission by 2050, which requires halting and reversing peatland C emissions through the restoration of peat-forming ecosystems by rewetting and re-establishing peatland vegetation (Tanneberger et al. 2021).
Rewetting of drained peatlands often leads to socio-economic dilemmas, because many livelihoods depend on drained peatland-use (Ziegler et al. 2021). Crop production on rewetted peatlands, named paludiculture, allows both the mitigation of negative environmental impacts of peat drainage and economic gains (Wichtmann et al. 2016). With Sphagnum biomass produced in Sphagnum paludiculture, fossil peat can be substituted by sustainably produced biomass for horticultural growing media (Jobin et al. 2014; Gaudig et al. 2018). Even though in natural systems most Sphagnum species only grow under oligotrophic to mesotrophic conditions (Clymo 1973; Rydin and Jeglum 2013), some of them can accumulate large amounts of biomass and nutrients under nutrient rich conditions. This often requires an optimal supply of nutrients in a balanced ratio and proper management of water level and vascular plants (Temmink et al. 2017; Gaudig et al. 2018; Vroom et al. 2020).
Sphagnum paludiculture and peat formation require sustained organic matter accumulation (Rydin and Jeglum 2013; Gaudig et al. 2018). Sphagnum dry mass accumulation in Sphagnum paludiculture was determined as being 3.6–4.4 t/ha/yr in average, for 2.5 and 7.5 years old sites (Gaudig et al. 2018; Vroom et al. 2020). In most Sphagnum paludicultures in Europe, the macronutrients for plant uptake nitrogen (N), phosphorus (P), and potassium (K) typically originate from various sources including atmospheric deposition, irrigation water, and agricultural soil legacy (Smolders et al. 2008; Emsens et al. 2015; Temmink et al. 2017; Zak et al. 2018; Vroom et al. 2020). This is because of the former land use of drained peatlands by intensive agriculture or fringing mineral soils that led to nutrient loads and/or high N deposition rates (Temmink et al. 2023a). In areas that were less influenced by intensive agriculture the nutrient situation may differ. Optimal NPK ratios in the irrigation water stimulate high Sphagnum productivity, whereas unbalanced supply results in impaired growth (Limpens and Berendse 2003; Bragazza et al. 2004; Temmink et al. 2017). N in the form of ammonium (NH4+) negatively affects Sphagnum, as it can become toxic at high concentrations (Limpens and Berendse 2003; Breeuwer et al. 2009). Next to biomass, Vroom et al. (2020) studied nutrient dynamics in Sphagnum paludiculture and showed that porewater NH4+ and P seemed to accumulate gradually after 7.5 years. Furthermore, Sphagnum paludiculture showed to have accumulated and stored nutrients up to almost 50 kg N, 3 kg K and 0.5 P ha/yr in fields of 7.5 years old (Vroom et al. 2020). Yet, it remains unclear whether (i) nutrients accumulate in the porewater on a decadal time scale reaching levels that can hamper growth, and (ii) the rewetted peatland under Sphagnum paludiculture management in the long run accumulates C and nutrients in the newly formed organic matter.
In this paper, we discuss nutrient dynamics and carbon and nutrient accumulation rates on a rewetted peatland under long-term Sphagnum paludiculture. The peatland in NW-Germany encompasses two unharvested sites of 10 and 5 years old (established in 2011 and 2016, respectively). The comparison of these two sites enabled us to uncouple the effect of age from yearly weather fluctuations, while also showing the reproducibility of the farming strategy. As the newly formed organic matter had not yet been harvested, the study also provides insight for bog restoration on rewetted agriculturally used peatland. We hypothesized that (i) NH4+ and P accumulate in the porewater over time through sustained input by irrigation water, atmospheric N deposition and internal mobilization, and that K and Cl remain stable in the newly formed organic layer over time and in depth (Vroom et al. 2020), (ii) the organic matter layer continues to grow homogeneously depending on the distance of the source of irrigation (Temmink et al. 2017; Gaudig et al. 2018; Vroom et al. 2020), and (iii) the accumulated organic matter shows sustained longer term C, N, P, and K accumulation (Vroom et al. 2020).
Materials and methods
Study sites
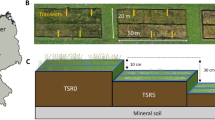
The studied rewetted peatland under Sphagnum paludiculture is located in NW Germany (53° 15.80′ N, 08° 16.05′ E). The area was a former raised bog with a current peat layer of 2–2.5 m that was used as a drained and intensively fertilized agricultural grassland since the 1950s. Intensity of grazing and fertilization decreased in the years prior to the conversion of fields into Sphagnum paludiculture. The surrounding peatlands remain drained and under drained agricultural use. To convert the area into a Sphagnum paludiculture, 30–50 cm of the strongly mineralised and nutrient enriched topsoil (> H8 after von Post 1924) was removed with an excavator to prepare plain production fields (10 m width) (Temmink et al. 2017, e.g. Quadra et al. 2023). Furthermore, irrigation ditches of 0.5 m wide and deep were dug, and levees were constructed to function as causeways. The Sphagnum founder material of S2011 (which originated from the Wieden National Park, the Netherlands, and the Sphagnum paludiculture site Ramsloh, Lower Saxony, Germany) was spread before rewetting (Fig. 1B, Gaudig et al. 2014), originally consisted of a mix of Sphagnum palustre and S. papillosum with some S. fallax. S2016 was installed with mainly S. fallax from other Sphagnum fields installed at the same site in 2011, where it had outcompeted S. palustre and S. papillosum. Sphagnum was spread with 80m3/ha to reach a cover of c. 70–80% (Gaudig et al. 2018). The Sphagnum paludiculture site was established on 4.5 ha in 2011 (named S2011) and extended with 9.5 ha in 2016 (named S2016, Fig. 1). The water table was maintained c. 5 cm below the Sphagnum surface (c.f., Gaudig et al. 2020) through actively supplying irrigation water from the adjacent stream ‘Schanze’ (average 2.900, min 450, max, 4.800 m3/ha/yr; calculated for the period 2013 till 2021; Brust et al. 2018, unpublished data) and an overflow outlet. The outlet can be adjusted to manage the water table. The water table was consistently stable and high (Brust et al. 2018). The nutrient concentrations of the irrigation water in the ditch are presented in Figure S2. Vascular plants, dominated by Juncus effusus, were mown 4–8 times during the growing season (May–October) and mown plant material remained on the production fields till 2016 and was always removed in the following years due to a change in the mowing method (Wichmann et al. 2020). The mowing of the sites changed over the years from using a brush cutter or strimmer to single-axle mowing with cutter bar and triple tyres (2011–2015) to using an excavator equipped with an extra-long arm and a mowing bucket, operating from a causeway (2016–2021, for photographs see Gaudig et al. 2018). The latter was used to prevent compaction of the peat layer.

A Overview of sampling locations. B Cross section of the Sphagnum paludiculture field in site S2011 with the sampling scheme of a transect (white line). C Timeline of the installation of the two sites (installation in 2011 = S2011, installation in 2016 = S2016) and the sampling events. Sampling of both sites is indicated by a bold S. White circles represent years. Map adapted from Vroom et al. (2020). Map background in A: Microsoft® BingTM Maps
Transects setup
In March 2014, we installed six transects perpendicular to the irrigation ditches up to half the width of the production fields on the then three year old site S2011 (5 m, Fig. 1) (Temmink et al. 2017). At the S2016 site, we installed nine similar transects one year after its setup (Vroom et al. 2020). Transects had measurement points at 0 (ditch surface water), 0.5, 1, 2.5, and 5 m distance from the nearest irrigation ditch, where we took pore water samples at a depth of 0–10 cm and biomass samples (Fig. 1B). At 5 m distance from the ditch, we additionally sampled pore water at a depth of 0.25, 0.5, and 1 m below the Sphagnum surface. We sampled the surface and pore water in March and November 2014 at S2011 only (Temmink et al. 2017), in June and November 2017, in March, June and November 2018 (Vroom et al. 2020), and in June 2020 and June 2021 at both sites (this study). We took the water samples on dry days to prevent dilution by heavy rainfall. To prevent disturbing and compaction of the peat, we worked on dedicated walking routes on the Sphagnum lawns and used snowshoes. We sampled the newly accumulated organic matter, which included Sphagnum, vascular plants and other mosses, in June 2021 (i.e., ten and five years after installation for S2011, and S2016, respectively).
Surface and pore water quality in the top and subsoil organic matter layer
To investigate whether Sphagnum lawns accumulate nutrients over time and depth, we determined the chemical composition of the pore water. During every field campaign (n = 9 for S2011 and n = 7 for S2016), ditch surface and pore water was extracted in each transect (Fig. 1). Furthermore, we collected surface water samples in the stream ‘Schanze’, which provides irrigation water for the Sphagnum paludiculture site. The pore water samples at 0–0.1 m depth (i.e., top layer of newly formed organic matter) and samples at 0.25, 0.5, and 1 m depth (i.e., subsoil layer) relative to the surface were taken using syringes under vacuum attached to a rhizon (Rhizosphere Research Products, Wageningen, The Netherlands) or ceramic cups, respectively. To prevent oxygenation of the samples, syringes were closed, stored at 4 °C, and processed within two days.
Organic matter accumulation and nutrient storage
To study how the proximity to the irrigation ditch affected the growth of Sphagnum and other plants, we quantified organic matter accumulation in all transects in June 2021. We cut out 10 by 10 cm cores from the Sphagnum lawn in the transect sampling points down to the old peat surface on which Sphagnum mosses were introduced at installation. We determined lawn thickness in-situ by measuring the distance of the Sphagnum surface to the old peat and calculated the volume using the newly formed organic matter thickness and surface area. Other mosses, vascular plants including roots, Sphagnum capitula, and other living and dead Sphagnum material were separated, weighed (fresh weight), dried for 72 h at 70 °C, and weighed again (dry weight). Dried organic matter was ground at 18 k rounds per minute for 2 min using 5 mm bullets (bullet grinder Mixer Mill MM 400, Retsch, Haan, Germany) to ensure sample homogeneity. Samples were stored dry and in the dark until chemical analyses.
Chemical analyses
Within one day after sampling, we determined the pH and alkalinity of the pore and surface water using an Ag/AgCl electrode (Orion Research, Beverly, MA, USA) and a TIM 840 Titration Manager (Radiometer Analytical SAS, Villeurbanne, France). Total inorganic C (TIC—HCO3− and CO2) was measured using an infrared C analyser (IRGA; ABB Analytical, Frankfurt, Germany), followed by pH-based calculation of CO2 and HCO3− concentrations (van Bergen et al. 2020). All water samples were divided and i) stored at 4 °C in vials (10 mL) containing 0.1 mL of 65% nitric acid (HNO3) or ii) frozen and stored at − 20 °C (20 mL) until further analyses.
Ammonium (NH4+), nitrate (NO3−), and chloride (Cl−) concentrations of pore and surface water subsamples stored at − 20 °C were determined by colorimetric methods (Auto Analyser III, Bran and Luebbe GmbH, Norderstedt, Germany). In the other subsamples, K and P were measured using inductively coupled plasma optical emission spectrometry (ICP-OES iCAP, Thermo Fischer Scientific, Bremen, Germany). Since 2020, ICP samples were measured with ARCOS (ICP-OES-ARCOS Spectro Analytical; Kleve, Germany), which was calibrated to the ICP-OES.
Total C and N content in dry and homogenized plant material (4–5 mg; Sphagnum capitula or other biomass, other mosses, and vascular plants) was determined using an elemental CNS analyser (Vario MICRO cube; Elementar Analysensysteme, Langenselbold, Germany). Total P and total K content of the biomass samples were determined by ICP-OES (see above) after adding 4 mL nitric acid (HNO3) (65%), 1 mL hydrogen peroxide (H2O2) (30%), and 95 mL milliQ water to 200 mg dried and homogenized plant material in Teflon vessels, followed by heating in a microwave oven (EthosD, Milestone, Sorisole Lombardy, Italy).
Statistical analyses
We analysed how nutrient concentrations, accumulated organic material, and nutrient accumulation (dependent variables) were influenced by time, depth, and site (independent variables) using linear mixed models (lme) and analyses of variances (ANOVAs). Statistics were performed using R version 4.03 (R Core Team 2020). All data are shown with their average ± Standard Errors (SE), and in all hypothesis testing procedures the significance level was pre-set at α = 0.05.
Differences in nutrient variables (dependent variables: NH4+, P, K, Cl−) in all transects and depth profiles were tested using linear mixed models with the ‘nlme’ package (Pinheiro et al. 2017) followed by ANOVAs according to Zuur et al. (2009). For the transects, main effects (independent variables) of ‘site’ and ‘date’ and their two-way interaction were included in the model. ‘Transect number’ and ‘distance to the ditch’ were included as random factor for only P, as it decreased the Akaike information criterion (AIC) value. For the depth profiles, main effects (independent variables) of ‘site’, ‘depth’, and ‘date’ and their two-way interaction were included in the model. Transect number was included in all models (see above). Non-significant interactions were removed from the model in a backwards stepwise analysis (Zuur et al. 2009). Specifically for P, the interactions ‘site*date’, and for K ‘site*date’ and ‘site*depth’ were dropped. Residual plots were created for visual assessment of normality and homogeneity. All data were log transformed to improve normality and homogeneity of residuals.
Differences in accumulated material (dependent variables: total weight, Sphagnum, other mosses, vascular plants, lawn thickness, volume) and ‘distance to the irrigation ditch’ (independent variable) after 5 or 10 years were analysed using one-way ANOVAs. We used Tukey-adjusted comparisons to test differences between distances to the irrigation ditch. Residual plots were created for visual assessment of normality and homogeneity.
Results
Nutrient dynamics in the top 10 cm of the Sphagnum paludiculture field layer
Porewater nutrient concentrations in the top 10 cm of accumulated organic matter were low for NH4+ and P in S2011, but increased in S2016 (Fig. 2; F1,447 = 39, p < 0.001, F1,384 = 148.7, p < 0.001 for NH4+ and P, respectively). Porewater K concentrations increased slightly in S2011 and strongly in S2016 over time (F1,447 = 46.3, p < 0.001). Cl concentrations increased marginally over time in S2011, and decreased over time in S2016 (F1,447 = 5.5, p = 0.02), but were generally lower than 2000 µmol/L. The largest deviation from the 10-year average for Cl occurred in 2018 coinciding with a temporarily lowering of the water table, which resulted in a porewater Cl concentration of 2800 ± 130 µmol/L at S2011. Across the sites and over 10 years, the porewater concentrations were 17 ± 2 µmol/L for NH4+, 10.5 ± 1 µmol/L for P, 43.5 ± 3 µmol/L for K, and 1040 ± 31 µmol/L for Cl. NO3− was virtually absent with 2 ± 0.3 µmol/L. The nutrient concentrations in the irrigation ditches are shown in Figure S1.

Nutrient dynamics in the top layer over time. Porewater (A–B) NH4+, (C–D) phosphorus, (E–F), potassium, and (G–H) chloride in the upper 10 cm over time. Red points depict site S2011 (n = 24 per sampling campaign) and blue S2016 (n = 36 per sampling campaign). Dark colours indicate data points overlap. Symbols represent distances to the irrigation ditch and show overlap between distance to the irrigation ditch. The vertical dotted lines depict the year of installation of the Sphagnum paludiculture
Nutrient dynamics in the sub layer
Porewater NH4+ concentrations were consistently low in S2011 and decreased strongly over time and depth in S2016. They were affected by site, depth, time since installation, and their interactions (Figs. 3 and 4). Specifically, porewater NH4+ concentrations were generally low with 20 ± 4 µmol/L at S2011 and higher with 130 ± 11 µmol/L at S2016 (F3,407 = 10, p < 0.001). NH4+ concentrations decreased over time at S2016 but not for S2011 where they were already low (F1,407 = 77.2, p < 0.001, Fig. 3). At S2016 at 50 cm depth, for example, porewater concentrations of NH4+ decreased from 280 ± 32 in year 1 to 14 ± 1 µmol/L year 5. Porewater K concentrations were similar in S2011 and S2016 (F1,407 = 2.9, p = 0.09) and decreased over time at 50 and 100 cm depth at both sites, while concentrations increased at 10 cm depth (F3,407 = 5.1, p = 0.02, Fig. 4). Porewater P concentrations were lowest at shallow depths and highest at deeper layers at S2011 and S2016 (F3,406 = 12.5, p < 0.001). Porewater P concentrations decreased over the study period at 25, 50, and 100 cm depth and increased at 10 cm depth (F3,406 = 19.6, p < 0.001). At S2016 at 100 cm depth, for example, porewater concentrations of P decreased from 15 ± 6 at year 1 to 3 ± 0.3 µmol/L at year 5 (Figure S1).

Porewater NH4+ dynamics up to 100 cm depth over time. Dynamics of porewater NH4+ concentrations at 10, 25, 50, and 100 cm depth relative to the land surface over time at the centre of the field (5 m) in S2011 (red, n = 6 per sampling campaign) and S2016 (blue, n = 9 per sampling campaign). Dark colours indicate data points overlap. The vertical dotted line depicts the year of installation of the Sphagnum paludiculture site

Porewater K dynamics up to 100 cm depth over time. Dynamics of porewater K concentrations at 10, 25, 50, and 100 cm depth relative to the land surface over time at the centre of the field (5 m) in S2011 (red, n = 6 per sampling campaign) and S2016 (blue, n = 9 per sampling campaign). Dark colours indicate data points overlap. The vertical dotted line depicts the year of installation of the Sphagnum paludiculture site
Organic matter accumulation and nutrient storage
During 10 years, a new organic matter layer with a dry weight of c. 6 kg/m2 formed at S2011 (Fig. 5). Specifically, the newly formed organic layer had a dry weight of 5.9 ± 0.3 kg/m2 formed at S2011, irrespective of the distance to the irrigation ditch (F1,22 = 0.05, p = 0.8, Fig. 5). Sphagnum, vascular plants, and other moss biomass were unaffected by the distance to the ditch (F1,22 = 3.1, p = 0.1, F1,22 = 0.1, p = 0.7, F1,22 = 1.5, p = 0.2 for Sphagnum, vascular plants, and other moss weight, respectively). However, Sphagnum biomass tended to be higher at 0.5 and 5 m (4 ± 0.6 and 4.9 ± 0.6 kg/m2 for 0.5 and 5 m, respectively) and lower at 1 and 2.5 m distance to the irrigation ditch (2.7 ± 0.8 and 3.1 ± 0.3 kg/m2 for 1 and 2.5 m, respectively; R2 = -0.64, p < 0.01, Fig. 5D, F). The newly formed organic layer was 27 ± 0.4 cm thick with a volume of 271 ± 4 L/m2 (F1,22 = 0.03, p = 0.9 and F1,22 = 0.03, p = 0.9 for thickness and volume, respectively, Figure S3). All newly formed biomass has accumulated nutrients over ten years and stored 26,000 kg C, 561 kg N, 31.5 kg P, and 89.9 kg K per hectare (Table 1). Organic matter accumulation and nutrient storage for S2016 can be found in Figure S4 and Table S1.

Organic matter accumulation after ten years post rewetting. New organic matter accumulated (A, kgDW/m2), (B) accumulated Sphagnum (kg/m2), (C) vascular plants (kg/m2), and (D) othermosses (kgDW/m2) at increasing distances to the irrigation ditch in S2011, n = 6. Boxplots show the median (middle line), quartiles (boxes), 1.5 times the interquartile range (IQR) (whiskers), and the individual data values (black circles). Black open circles outside the whiskers are extreme values
Discussion
Porewater nutrient accumulation was negligible and legacy nutrients rapidly declined
We did not observe an accumulation of NH4+ and P in the porewater of the newly upgrown Sphagnum lawn 10 years after rewetting in S2011, but only after 5 years in S2016. In the same area, Vroom et al. (2020) found indications of N accumulation in porewater after 7.5 years, but since then, we observed no further accumulation. Hydro-climatic fluctuations most likely caused the variation in NH4+ concentrations (i.e., the ratio between precipitation and surface water used for irrigation). For example, with 550 mm rain from April to September, the year 2021 was wet compared to the 10-year average of 380 mm. Higher rainfall may have diluted nutrients in the porewater or irrigation water and lowered the need for irrigation water, leading to lower nutrient loads. This is indicated by porewater Cl concentrations that were lower than the 10-year average in 2021 (Table S2), indicating dilution by rainwater. Moreover, the dry years of 2018 and 2019 required circa 3.000 m3/ha/yr irrigation water to maintain the water table (Brust et al. 2018, unpublished data). In the wet year of 2021, 5–10 times lower irrigation volumes in the magnitude of hundreds of m3/ha/yr were required to maintain this water table, resulting in lower surface water nutrient loads. Overall, our results reveal that an accumulation of porewater nutrients in the newly accumulated layer has not occurred and that hydro-climatic variation affected temporal porewater nutrient fluctuations.
One year after installation of S2016, we observed high concentrations of NH4+ and P in the porewater up to 1 m depth in the vertical profile, indicating an agricultural nutrient legacy of the former bog grassland use (Zak and Gelbrecht 2007; Zak et al. 2010; Temmink et al. 2017; Vroom et al. 2020). The agricultural legacy of NH4+ and P decreased rapidly over time and with depth to near-zero concentrations at 25 and 50 cm depth. This highlights the importance of long-term monitoring and warrants critical revisions of nutrient budget models and nutrient mobilisation estimates that are based on data collected during < five years after rewetting or restoration. As the sampling depth was related to the surface, the growth of the new organic layer resulted in a change in sampling depth. Yet, the porewater ammonium concentrations decreased more strongly over time and to levels lower than at the start of the measurements, highlighting the robustness of the change. Two years after removing the nutrient-rich topsoil at installation had resulted in legacy porewater K concentrations of near-zero in the top 10 cm layer. However, K slowly accumulated and reached levels comparable to the deeper layers after 2–5 years and subsequently remained stable. Most likely, K originated both from irrigation water with concentrations of 100 µmol/L and leakage of K from adjacent causeways (Vroom et al. 2020). Overall, hydro-climatic variation most likely controlled N and P concentrations in the newly accumulated layer, whereas time since rewetting controls concentrations of N, P, K in the subsoil. However, knowledge is lacking on nutrient dynamics the first 2 years after site installation (Temmink et al. 2017; Vroom et al. 2020), which warrants further study.
High organic matter and nutrient accumulation
Organic matter rapidly accumulated over ten years due to high and stable water table (Gaudig et al. 2018, 2020), optimal nutrient stoichiometry of the peat mosses (Temmink et al. 2017), and the frequent mowing of vascular plants (Gaudig et al. 2018). This resulted in a thick organic matter layer consisting mainly of peat mosses, other mosses, and vascular plants and because of adapted water management increased the water storing ability (i.e., water tables were raised with lawn height growth). Mean biomass and nutrient accumulation rates of/in this newly formed organic matter layer (6 t dry mass, 2,6 t C, 56 kg N, 3.2 kg P, and 9 kg K/ha/yr), are similar to rates earlier found at this area (Temmink et al. 2017; Vroom et al. 2020). However, these rates are considerably higher than those found in a pristine bog in Norway with 34 kg N/ha/yr (Ohlson and Halvorsen Økland 1998) and in Patagonia with 0.9 kg P ha/yr and 2.2 kg N ha/yr (Fritz et al. 2012).
The C accumulation rates found in our study are almost 2–3 times higher than in other productive Sphagnum systems (Nugent et al. 2018; Huth et al. 2022; Wilson et al. 2022). These studies differed with ours mainly in site management and irrigation water composition. For example, in the study by Nugent et al. (2018) and Wilson et al. (2022) the water table was not as strictly controlled, which is required to achieve high Sphagnum productivity (Gaudig et al. 2020). Furthermore, in our study, field sites were irrigated with surface water from a stream with higher nutrient concentrations than rainwater, which in turn can promote Sphagnum growth and thus C accumulation (Limpens et al. 2004; Gaudig et al. 2020). We argue that a balance between nitrogen, phosphorus and potassium is key to explain high C accumulation rates when water quantity management is close to optimal (see also Temmink et al. 2017). A comparison with Sphagnum systems in N-America should also take length of growing season, continentality index and intrinsic growth rates of Sphagnum species (and genotypes) into account. Additionally, the three studies highlighted give little information on establishment success of mosses in the first 18 months and to what extent repeated spreading of founder material (‘re-seeding’) was part of the management scheme, as performed for S2011 (Gaudig et al. 2014). We argue that the site studied here is managed close to optimal.
The C accumulation rate of 260–320 g C/m2/yr in Sphagnum paludiculture (5–10 years) is tenfold higher than the long-term C accumulation rates in natural bogs (> 100 years) (Yu et al. 2010; Young et al. 2019). It is known that recent C accumulation rates are higher than long-term accumulation rates due to sustained anaerobic decomposition (Yu et al. 2010; Young et al. 2019). Without organic matter harvest (i.e. not Sphagnum paludiculture, but carbon farming), we expect that anaerobic decomposition will increase leading to lower long-term cumulative C accumulation rates. Similarly, the adaptive water management in Sphagnum paludiculture (Brust et al. 2018) combined with very favourable nutrient rich irrigation water composition allowed rapid vertical growth of c. 2.7 cm/yr and we expect this growth can continue up until one of these conditions become less favourable, most probably water tables that cannot be increased (i.e. higher than the causeways or surrounding agricultural lands). Further research is needed to account for stochastic disturbance (e.g. drought, fire, disease of mosses) in C accumulation rates and whether this disturbance can largely be banned by management as provided at the current site. Furthermore, we would like to point out that optimal nutrient provision to mosses may be an important driver of system inherent high gross primary production and litter accumulation at this site. With the water table constantly managed upwards, biomass accumulation (in height) is not increasing the width/volume of aerobic decomposition.
Interestingly, Sphagnum grew well and sequestered considerably more N than the local atmospheric nitrogen deposition of 21 kg ha/yr. The remaining N originated most likely from irrigation water (Vroom et al. 2020), input from the nutrient rich causeways, internal N mobilization from the peat, and N fixation by the mosses and their microbiome (Vile et al. 2014, van den Elzen et al. 2018). Over ten years, the accumulated N deposition equals to 210 kg/ha and input through surface water irrigation adds another 40–60 kg N/ha in our area (Vroom et al. 2020). Sphagnum paludiculture is a N-efficient system by uptaking of large amounts of available N without leaching like in fertilized conventional bog grasslands (Van Beek 2007; Smolders et al. 2010).
Implications for Sphagnum paludiculture and bog restoration
Our results indicate that despite relatively high input of nutrients through atmospheric deposition and irrigation water but without additional fertilization, organic matter accumulation remained high over ten years, with NPK ratios optimal for good Sphagnum growth (Temmink et al. 2017). High N deposition rates may have reduced carbon accumulation by Sphagnum mosses (Granath et al. 2012; Temmink et al. 2017). Furthermore, the relatively high nutrient availability in our area resulted in substantial mowing costs to prevent vascular plant dominance (Wichmann et al. 2020). However, the rapid decline of agricultural nutrient legacy improves environmental conditions for Sphagnum growth, as vascular plants are known to outcompete Sphagnum at high nutrient availability (Lamers et al. 2000; Tomassen et al. 2003). The net carbon storage in the newly formed organic layer over a 100-year lifespan (the standard “permanence” criterion in carbon schemes) will determine to what degree and under which conditions (no harvest, partial harvest, full harvest) Sphagnum paludiculture can be included in carbon farming and carbon removal schemes (Tanneberger and Wichtmann 2011; Smith et al. 2020). In our study, we found a high proportion of carbon stored in recalcitrant pools of Sphagnum biomass that is known to be a recalcitrant C pool (Fig. 5) (Jassey et al. 2011; Pipes and Yavitt 2022). This biomass is likely to remain largely stable given the absolute increase of water table height (Brust et al. 2018), which protects the organic matter from aerobic breakdown (Brouns et al. 2014). We hypothesize that carbon pools accumulated by Sphagnum paludiculture exceed the stability of soil carbon pools found in mineral soil grassland (Smith 2014). Overall, Sphagnum paludiculture management results in substantially less C emission compared to drained grasslands on peat (Daun et al. 2023). Specifically, the Sphagnum production fields are C sinks (i.e. where we measured the organic matter accumulation), but the ditches and causeways are major C sources (Daun et al. 2023). Further optimization in irrigation, topsoil removal and causeway construction and lay-out is required to improve the C balance of the Sphagnum farm. We hypothesize that carbon pools accumulated by Sphagnum paludiculture exceed the stability of soil carbon pools found in mineral soil grassland (Smith 2014).
Our investigations confirm results from Huth et al. (2022) that bog restoration is possible on former agricultural peatlands and not only on cut-over bogs or bog remnants (Convention on Wetlands 2021; Temmink et al. 2021). Water storage and stability of the water table is supposed to be improved by the newly formed organic layer (Figure S3), which showed a low bulk density (Vroom et al. 2020) often associated with a high volume of large pores (Fritz et al. 2008; Lennartz and Liu 2019). The sponge water holding and storing capacity function of this secondary acrotelm-like structure may increase the resilience to hydro-climatic extremes, such as floods and droughts of early-stage Sphagnum lawns (Ivanov 1981, Nijp et al. 2017). Our results highlight that intensive paludiculture management to facilitate high Sphagnum growth and the formation of a secondary acrotelm-like structure in 10 years is possible with additional measures to improve Sphagnum growth, including stable- and high water level (Brust et al. 2018), optimal nutrient stoichiometry (this paper, Temmink et al. 2017; Vroom et al. 2020) and reduced competition with vascular plants through mowing (Gaudig et al. 2014), can yield rapid results in the first years after rewetting. This knowledge to create a secondary acrotelm-like structure in 10 years should inform bog restoration worldwide. Furthermore, Sphagnum grown in a paludiculture setting may become key as sustainably-produced donor material for revegetating restoration sites (Wichmann et al. 2020).
Conclusions
We conclude that (i) Sphagnum paludiculture can sustain high organic matter, C and nutrient accumulation on a decadal time scale with no other nutrient input than nutrient deposition from the atmosphere and surface water irrigation, provided biomass is not harvested, (ii) NH4+ and P accumulation in the porewater has not occurred after 10 years, and (iii) Sphagnum paludiculture effectively remediates the agricultural nutrient legacy in the original peat. Beyond paludiculture, our results provide insight that bog-like vegetation establishment is possible on former agricultural peatlands even in a nutrient rich environment under adequate management measurescorrect management. Interestingly, a relative low N input resulted in high C accumulation relative to mineral soil farming practices. More broadly, global degradation of peatlands with negative environmental impacts, including loss of biodiversity, greenhouse gas emissions, downstream pollution, and land subsidence, calls for large-scale restoration and the re-establishment of vital peatland functions. Therefore, we argue that the implementation of productive use of rewetted peatlands through Sphagnum paludiculture can aid in reaching targets set in the Paris Agreement and the UN Decade on Ecosystem Restoration.
Data availability
All data is available through Yoda, a research data management service by Utrecht University (https://public.yoda.uu.nl/geo/UU01/41QFC6.html): Temmink et al. (2023b).
References
Ahmad S, Liu H, Günther A, Couwenberg J, Lennartz B (2020) Long-term rewetting of degraded peatlands restores hydrological buffer function. Sci Total Environ 749:141571
Bianchi A, Larmola T, Kekkonen H, Saarnio S, Lång K (2021) Review of greenhouse gas emissions from rewetted agricultural soils. Wetlands 41:1–7
Bragazza L, Tahvanainen T, Kutnar L, Rydin H, Limpens J, Michal H, Grosvernier P, Tom H, Hajkova P, Hansen I, Iacumin P, Gerdol R (2004) Nutritional constraints in ombrotrophic Sphagnum plants under increasing atmospheric nitrogen deposition in Europe. New Phytol 163:609–616
Breeuwer A, Heijmans MPD, Gleichman M, Robroek BM, Berendse F (2009) Response of Sphagnum species mixtures to increased temperature and nitrogen availability. Plant Ecol 204:97–111
Brouns K, Verhoeven JTA, Hefting MM (2014) The effects of salinization on aerobic and anaerobic decomposition and mineralization in peat meadows: the roles of peat type and land use. J Environ Manage 143:44–53
Brust K, Krebs M, Wahren A, Gaudig G, Joosten H (2018) The water balance of a Sphagnum farming site in north-west Germany. Mires and Peat 20:1–12
Clymo RS (1973) Growth of Sphagnum - some effects of envionment. J Ecol 61:849–869
Convention on Wetlands (2021) Global guidelines for peatland rewetting and restoration, 11th edn. Secretariat of the Convention on Wetlands, Gland, Switzerland
Daun C, Huth V, Gaudig G, Günther A, Krebs M, Jurasinski G (2023) Full-cycle greenhouse gas balance of a Sphagnum paludiculture site on former bog grassland in Germany. Sci Total Environ 877:162943
Emsens W-J, Aggenbach CJS, Smolders AJP, van Diggelen R (2015) Topsoil removal in degraded rich fens: can we force an ecosystem reset? Ecol Eng 77:225–232
Fritz C, Campbell DI, Schipper LA (2008) Oscillating peat surface levels in a restiad peatland, New Zealand—magnitude and spatiotemporal variability. Hydrol Process 22:3264–3274
Fritz C, van Dijk G, Smolders AJP, Pancotto VA, Elzenga TJTM, Roelofs JGM, Grootjans AP (2012) Nutrient additions in pristine Patagonian Sphagnum bog vegetation: can phosphorus addition alleviate (the effects of) increased nitrogen loads. Plant Biol 14:491–499
Gaudig G, Fengler F, Krebs M, Prager A, Schulz J, Wichmann S, Joosten H (2014) Sphagnum farming in Germany: a review of progress. Mires and Peat 13:1–13
Gaudig G, Krebs M, Prager A, Wichmann S, Barney M, Caporn SJM, Emmel M, Fritz C, Graf M, Grobe A, Pacheco SG (2018) Sphagnum farming from species selection to the production of growing media : a review. Mires and Peat 13:1–30
Gaudig G, Krebs M, Joosten H (2020) Sphagnum growth under N saturation: interactive effects of water level and P or K fertilization. Plant Biol. https://doi.org/10.1111/plb.13092
Gorham E (1991) Northern peatlands: role in the carbon cycle and probable responses to climatic warming. Ecol Appl 1:182–195
Granath G, Strengbom J, ¥kan Rydin H (2012) Direct physiological effects of nitrogen on Sphagnum: a greenhouse experiment. Funct Ecol 26:353–364
Hooijer A, Page S, Jauhiainen J, Lee WA, Lu XX, Idris A, Anshari G (2012) Subsidence and carbon loss in drained tropical peatlands. Biogeosciences 9:1053–1071
Huth V, Günther A, Bartel A, Gutekunst C, Heinze S, Hofer B, Jacobs O, Koebsch F, Rosinski E, Tonn C (2022) The climate benefits of topsoil removal and Sphagnum introduction in raised bog restoration. Wiley Online Library
Ivanov, K. E. 1981. Water movement in mirelands. Academic Press, London England. 1981. Translated by Arthur Thomson and H. A. P. Ingram. 276.
Jassey VEJ, Chiapusio G, Gilbert D, Buttler A, Toussaint M, Binet P (2011) Experimental climate effect on seasonal variability of polyphenol/phenoloxidase interplay along a narrow fen–bog ecological gradient in Sphagnum fallax. Glob Change Biol 17:2945–2957
Jobin P, Caron J, Rochefort L (2014) Developing new potting mixes with Sphagnum fibers. Can J Soil Sci 94:585–593
Joosten, H., and D. Clarke. 2002. Wise use of mires and peatlands - background and principles including a framework for decision-making. International Mire Conservation Group and International Peat Society.
Jurasinski G, Ahmad S, Anadon-Rosell A, Berendt J, Beyer F, Bill R, Blume-Werry G, Couwenberg J, Günther A, Joosten H (2020) From understanding to sustainable use of peatlands: The WETSCAPES approach. Soil Systems 4:14
Lamers LPM, Bobbink R, Roelofs JGM (2000) Natural nitrogen filter fails in polluted raised bogs. Glob Change Biol 6:583–586
Lamers LPM, Vile MA, Grootjans AP, Acreman MC, van Diggelen R, Evans MG, Richardson CJ, Rochefort L, Kooijman AM, Roelofs JGM, Smolders AJP (2015) Ecological restoration of rich fens in Europe and North America: from trial and error to an evidence-based approach. Biol Rev 90:182–203
Leifeld J, Menichetti L (2018) The underappreciated potential of peatlands in global climate change mitigation strategies. Nat Commun 9:1071
Lennartz B, Liu H (2019) Hydraulic functions of peat soils and ecosystem service. Front Environ Sci 7:92
Limpens J, Berendse F (2003) Growth reduction of Sphagnum magellanicum subjected to high nitrogen deposition: the role of amino acid nitrogen concentration. Oecologia 135:339–345
Limpens J, Berendse F, Klees H (2004) How phosphorus availability affects the impact of nitrogen deposition on Sphagnum and vascular plants in bogs. Ecosystems 7:793–804
Nijp JJ, Metselaar K, Limpens J, Teutschbein C, Peichl M, Nilsson MB, Berendse F, van der Zee SE (2017) Including hydrological self-regulating processes in peatland models: Effects on peatmoss drought projections. Sci Total Environ 580:1389–1400
Nugent KA, Strachan IB, Strack M, Roulet NT, Rochefort L (2018) Multi-year net ecosystem carbon balance of a restored peatland reveals a return to carbon sink. Glob Change Biol 24:5751–5768
Ohlson M, Halvorsen Økland R (1998) Spatial variation in rates of carbon and nitrogen accumulation in a boreal bog. Ecology 79:2745–2758
Pinheiro, J., D. Bates, S. DebRoy, D. Sarkar, S. Heisterkamp, B. Van Willigen, and R. Maintainer. 2017. Package ‘nlme.’ Linear and nonlinear mixed effects models, version 3.
Pipes GT, Yavitt JB (2022) Biochemical components of Sphagnum and persistence in peat soil. Can J Soil Sci 102:785–795
Quadra GR, Boonman CCF, Vroom RJE, Temmink RJM, Smolders AJP, Geurts JJM, Aben RCH, Weideveld STJ, Fritz C (2023) Removing 10 cm of degraded peat mitigates unwanted effects of peatland rewetting: a mesocosm study. Biogeochemistry 163:1–20
R Core Team. 2020. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Rydin H, Jeglum JK (2013) The Biology of Peatlands. Oxford University Press
Smith P (2014) Do grasslands act as a perpetual sink for carbon? Glob Change Biol 20:2708–2711
Smith P, Soussana J, Angers D, Schipper L, Chenu C, Rasse DP, Batjes NH, Van Egmond F, McNeill S, Kuhnert M (2020) How to measure, report and verify soil carbon change to realize the potential of soil carbon sequestration for atmospheric greenhouse gas removal. Glob Change Biol 26:219–241
Smolders AJP, Lucassen EC, Bobbink R, Roelofs JGM, Lamers LPM (2010) How nitrate leaching from agricultural lands provokes phosphate eutrophication in groundwater fed wetlands: the sulphur bridge. Biogeochemistry 98:1–7
Smolders AJP, Lucassen ECHET, van der Aalst M, Lamers LPM, Roelofs JGM (2008) Decreasing the abundance of Juncus effusus on former agricultural lands with noncalcareous sandy soils: possible effects of liming and soil removal. Restor Ecol 16:240–248
Tanneberger F, Abel S, Couwenberg J, Dahms T, Gaudig G, Günther A, Kreyling J, Peters J, Pongratz J, Joosten H (2021) Towards net zero CO2 in 2050: an emission reduction pathway for organic soils in germany. Mires and Peat 27
Tanneberger F, Wichtmann W (2011) Carbon Credits from Peatland Rewetting: Climate - Biodiversity - Land Use. Schweizerbart Science Publishers, Stuttgart
Temmink RJM, Fritz C, van Dijk G, Hensgens G, Lamers LPM, Krebs M, Gaudig G, Joosten H (2017) Sphagnum farming in a eutrophic world: the importance of optimal nutrient stoichiometry. Ecol Eng 98:196–205
Temmink RJM, Cruijsen PMJM, Smolders AJP, Bouma TJ, Fivash GS, Lengkeek W, Didderen K, Lamers LPM, van der Heide T (2021) Overcoming establishment thresholds for peat mosses in human-made bog pools. Ecol Appl 31:e02359
Temmink RJM, Lamers LPM, Angelini C, Bouma TJ, Fritz C, van de Koppel J, Lexmond R, Rietkerk M, Silliman BR, Joosten H, van der Heide T (2022) Recovering wetland biogeomorphic feedbacks to restore the world’s biotic carbon hotspots. Science 376:eabn1479
Temmink RJM, Robroek BJM, van Dijk G, Koks AHW, Käärmelahti SA, Barthelmes A, Wassen MJ, Ziegler R, Steele MN, Giesen W (2023a) Wetscapes: Restoring and maintaining peatland landscapes for sustainable futures. Ambio. https://doi.org/10.1007/s13280-023-01875-8
Temmink RJM, Vroom RJE, van Dijk G, Käärmelahti SA, Koks AHW, Joosten H, Krebs M, Gaudig G, Brust K, Lamers LPM, Smolders AJP, Fritz C (2023b) Data from: restoring organic matter, carbon and nutrient accumulation in degraded peatlands: 10 years Sphagnum paludiculture. Yoda. https://doi.org/10.24416/UU01-41QFC6
Tomassen HBM, Smolders AJP, Lamers LPM, Roelofs JGM (2003) Stimulated growth of Betula pubescens and Molinia caerulea on ombrotrophic bogs: role of high levels of atmospheric nitrogen deposition. J Ecol 91:357–370
Turetsky MR, Benscoter B, Page S, Rein G, Van Der Werf GR, Watts A (2015) Global vulnerability of peatlands to fire and carbon loss. Nat Geosci 8:11–14
Van Beek C (2007) Nutrient losses from grassland on peat soil. Wageningen University and Research. PhD-thesis
van Bergen TJHM, Temmink RJM, van Tweel-Groot L, Bakker WJ, Rehlmeyer K, Koks AHW, Waajen AC, Roelofs JGM, Grootjans AP, van der Heide T, Lamers LPM (2020) Self-facilitation and negative species interactions could drive microscale vegetation mosaic in a floating fen. J Veg Sci 31:343–354
van den Elzen E, van den Berg LJL, van der Weijden B, Fritz C, Sheppard LJ, Lamers LPM (2018) Effects of airborne ammonium and nitrate pollution strongly differ in peat bogs, but symbiotic nitrogen fixation remains unaffected. Sci Total Environ 610:732–740
Vile M, Kelman Wieder R, Živković T, Scott K, Vitt D, Hartsock J, Iosue C, Quinn J, Petix M, Fillingim H, Popma JA, Dynarski K, Jackman T, Albright C, Wykoff D (2014) N2-fixation by methanotrophs sustains carbon and nitrogen accumulation in pristine peatlands. Biogeochemistry 121:317–328
Von Post, L. 1924. Das genetische system der organogenen bildungen schwedens. Memoires sur la nomenclature et la classification des sols. International Committee of Soil Science, Helsinki: 287–304.
Vroom RJE, Temmink RJM, van Dijk G, Joosten H, Lamers LPM, Smolders AJP, Krebs M, Gaudig G, Fritz C (2020) Nutrient dynamics of Sphagnum farming on rewetted bog grassland in NW Germany. Sci Total Environ 726:138470
Wichtmann W, Schröder C, Joosten H (2016) Paludiculture-productive use of wet peatlands. Schweizerbart Science Publishers, Stuttgart
Wichmann S, Krebs M, Kumar S, Gaudig G (2020) Paludiculture on former bog grassland: profitability of Sphagnum farming in North West Germany. Mires and Peat 20:1–18
Wilson D, Mackin F, Tuovinen J, Moser G, Farrell C, Renou-Wilson F (2022) Carbon and climate implications of rewetting a raised bog in Ireland. Glob Change Biol 28:6349–6365
Young DM, Baird AJ, Charman DJ, Evans CD, Gallego-Sala AV, Gill PJ, Hughes PDM, Morris PJ, Swindles GT (2019) Misinterpreting carbon accumulation rates in records from near-surface peat. Sci Rep 9:1–8
Yu Z, Loisel J, Brosseau DP, Beilman DW, Hunt SJ (2010) Global peatland dynamics since the Last Glacial Maximum. Geophys Res Lett 37:L13402
Zak D, Gelbrecht J (2007) The mobilisation of phosphorus, organic carbon and ammonium in the initial stage of fen rewetting (a case study from NE Germany). Biogeochemistry 85:141–151
Zak D, Wagner C, Payer B, Augustin J, Gelbrecht J (2010) Phosphorus mobilization in rewetted fens: the effect of altered peat properties and implications for their restoration. Ecol Appl 20:1336–1349
Zak D, Goldhammer T, Cabezas A, Gelbrecht J, Gurke R, Wagner C, Reuter H, Augustin J, Klimkowska A, McInnes R (2018) Top soil removal reduces water pollution from phosphorus and dissolved organic matter and lowers methane emissions from rewetted peatlands. J Appl Ecol 55:311–320
Zedler JB, Kercher S (2005) Wetland resources: status, trends, ecosystem services, and restorability. Annu Rev Environ Resour 30:39–74
Ziegler R, Wichtmann W, Abel S, Kemp R, Simard M, Joosten H (2021) Wet peatland utilisation for climate protection–an international survey of paludiculture innovation. Cleaner Engineering and Technology 5:100305
Zuur AF, Leno EN, Walker NJ, Saveliev AA, Smith GM (2009) Mixed effect models and extensions in ecology with R. Springer, New York
Acknowledgements
The authors thank all volunteers for supporting the practical work, and Sebastian Krosse, Germa Verheggen, Roy Peters and Peter Cruijsen for their help with chemical analyses. We highly appreciated the assistance of Keno Gerwing and Heinz Schmidt in managing the area.
Funding
Part of this research was funded by Interreg NWE Carbon Connects. C.F. received funding under Wet Horizons (GAP-10105684 Horizon Europe). We thank the German Federal Ministry of Food, Agriculture and Consumer Protection (BMELV), the Ministry for Environment, Energy and Climate Protection of Lower Saxony and the European Regional Development Fund for funding the research projects MOOSGRÜN, MOOSWEIT and OptiMOOS and the peat company Torfwerk Moorkultur Ramsloh Werner Koch GmbH & Co. KG, and Deutsche Torfgesellschaft mbH, for their financial and in-kind support of this study.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. GG and MK: supervised the set-up and performance of the Sphagnum paludiculture field trials. Material preparation and data collection were performed by RJMT, RV, GvD, SAK, AHWK, MK, GG, KB and CF. RJMT analyzed the data, created the figures and wrote the first draft of the manuscript. All authors commented on previous versions of the manuscript and read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Responsible Editor: Klaus Butterbach-Bahl.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Temmink, R.J.M., Vroom, R.J.E., van Dijk, G. et al. Restoring organic matter, carbon and nutrient accumulation in degraded peatlands: 10 years Sphagnum paludiculture. Biogeochemistry 167, 347–361 (2024). https://doi.org/10.1007/s10533-023-01065-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-023-01065-4