
Abstract
Very old trees harbour a diverse fauna of saproxylic insects, many of which are classified as threatened due to the scarcity of this kind of habitat. Parks, which often contain many old trees, are therefore considered to be important sites for this fauna. However parks are intensively managed and dead wood is often removed. Therefore this study compares if the saproxylic beetle fauna in parks is as diverse as it is in more natural stands. Eight ‘Park’ sites at manor houses around lake Mälaren, Sweden were compared with trees in wooded meadows: eight grazed sites, here termed ‘Open’, and 11 sites regrown with younger trees, termed ‘Re-grown’. The comparison was made on lime trees (Tilia spp.): one of the most frequent tree species in old parks which host a diverse beetle fauna. Beetles were sampled with window traps, which in total caught 14,460 saproxylic beetles belonging to 323 species, of which 50 were red-listed. When comparing all saproxylic species, ‘Park’ sites had significantly fewer species than ‘Open’ sites. However, for beetles in hollow trees and for red-listed species there was no significant difference, the number in ‘Park’ being intermediate between ‘Open’ and ‘Re-grown’. Species composition differed between sites, but only marginally so. Therefore, the conclusion is that old park trees on average are as valuable for faunal diversity as trees in more natural sites. Large conservation benefits can be obtained by combining cultural and conservational values in the management of park habitats.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Old trees is the habitat for a diverse fauna and flora. A large and well-known proportion of this fauna are beetles (Coleoptera) (Warren and Key 1991), among which are many red-listed or threatened species (Ranius and Jansson 2000; Speight 1989). Parkland, which often contains old trees, may therefore be a valuable resource for the conservation of these species (Carpaneto et al. 2010; Ehnström and Waldén 1986). Parkland, however, differs from other sites with old trees, as it is intensively managed in order to achieve the aesthetic effect of a large, tidy garden. Such intensive management is likely to be detrimental to saproxylic insects as it may often involve the removal of dead wood from the ground and tree crowns. Furthermore, old parks usually contain few bushes and small trees that might contribute to the habitat pool of dead wood. Nevertheless, studies conducted in parks and avenues have shown that they are used by threatened species (Gerell 2000; Jonsell 2004, 2008; Oleksa et al. 2006; Sörensson 2008). However, no quantitative comparisons between parks and other sites exist; this paper therefore aims to measure how parkland and more natural sites compare in their conservation value for saproxylic beetles.
The fauna of ancient trees is threatened because these trees have become increasingly rare in large parts of Europe, especially in the west (Emanuelsson 2009). Until relatively recently, old trees were widely distributed over large parts of the European landscape and were traditionally coppiced or pollarded at regular intervals to harvest wood for construction, fuel and for manufacturing wooden tools, and to provide fodder for cattle. Most of these woodlands have now been replaced by ‘high forests’ for timber production, or with modern agricultural lands. In Sweden, this decline of old trees is well documented for oak (Eliasson and Nilsson 2002). The ancient trees which remain until today were most often growing on land owned by the nobility, who could afford to keep them in parks or other semi-natural land. A century ago this land consisted of wooded meadows used for grazing, hay production and/or hunting. Today some of these areas are still kept open by grazing or they have regrown with young trees while the rest have been transformed to land without old trees. Land where the old trees still remain are highly prioritised in conservation work with protection and restoration.
In Europe, parks were often established around manor houses in the late 1600s or in the 1700s. Avenues of trees were an important feature of parks, with lime (Tilia spp.) being the most popular species at that time (Bengtsson 2005; Sernander 1926). In most of these old parks, at least in Sweden, some 300-year old trees still remain from the original plantings (Bengtsson 2005). A number of the original trees have died, but these have usually been individually replaced, so creating a continuous supply of trees that might grow into old age. As manor houses are relatively abundant in the countryside of the region where the present study was conducted, their parks probably harbour a considerable proportion of all the ancient trees present on a landscape-scale.
The tree species studied in this paper is lime (Tilia spp.), which hosts fewer saproxylic beetle species than, e.g. oak (Palm 1959). Compared to most other deciduous tree species, however, lime has a comparatively large assemblage of specialised saproxylic beetle species (Ehnström 2006; Palm 1959; Warren and Key 1991). But in general host specific differences in the fauna of ancient trees are not large because associated species are not usually confined to a single host species (Warren and Key 1991). Instead, the unique structures, such as hollows, dead parts of the trunk, dead branches, etc. are the important features. Because old lime trees are so frequent in parks, they might constitute an important proportion of habitat available at a landscape-scale, and so contribute to the long-term persistence of populations of saproxylic beetles.
The questions addressed in the present paper are:
-
(1)
Can park trees host a saproxylic beetle fauna as diverse as that found in trees of more natural stands?
-
(2)
Is there a difference if the natural sites are open grazed or re-grown?
Materials and methods
Study area
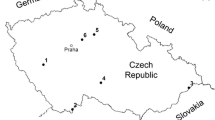
The study was conducted in an area of about 100 × 120 km2 situated around and north of lake Mälaren in Sweden (16°00′′–18°00′′E and 59°20′′–60°20′′N) (Fig. 1). The area lies within the hemiboreal zone (Ahti et al. 1968) which, although dominated by boreal tree species, is also characterised by a considerable element of southern deciduous tree species, e.g. oak Quercus robur, lime Tilia cordata, maple Acer platanoides, ash Fraxinus excelsior, elm Ulmus glabra, and hazel Corylus avellana, on richer soils and on sites with a warmer microclimate. All land with southern deciduous trees is much affected by present and former human land-use. Lime trees rarely dominate the stands, being rather scattered among other southern deciduous trees, mainly oak. Parks and a few other stands are exceptions. As in most of Europe, the older trees in the Mälaren area grew up in a landscape with large areas of hay meadows and grazing lands for cattle (Emanuelsson 2009), which are today either still grazed or regrowing with younger trees.

Map over the sampling sites. Characteristics for the sites are listed in Table 6
Lime trees were often pollarded to produce winter fodder for cattle, and wood, including the tough fibres in the bast, for a variety of uses. This practice was almost totally abandoned in the first half of the 1900s, but on many of the inventoried sites the trees have a conspicuous conformation from having been pollarded in earlier times. Lime trees in parks have also usually been pollarded, but for aesthetic reasons. On some of the natural stands however, there are no visible traces of pollarding. The limes in the natural sites are the small-leaved lime T. cordata, whereas most limes in parks are the common lime T. × europea, a hybrid between T. cordata and T. platyphyllos (Bengtsson 2005).
Around lake Mälaren there are many old estates that were built by the nobility. As described above, most of these estates had large parks established 250–350 years ago, an important feature of which were avenues of limes.
Selection of sites
Most study sites were selected for survey according to the criterion that they should contain lime trees that had the potential to host those species encompassed by an action plan for saproxylic beetles on lime (Ehnström 2006; Jonsell and Sahlin 2010) i.e. sites with old hollow lime-trees. The selection was mainly made by the county administrative boards in the respective county (three are included) based on information from inventories of valuable trees and on their personal knowledge. In addition, data from three other park inventories were included in this study (Andersson 2010; Jonsell 2004, 2008). In total, 27 sites were used and they were categorised as either ‘Open’ (8), ‘Re-grown’ (11) or ‘Park’ (8). The maximum area of a site was a few hectares, but was usually less than one. Each site was registered by GPS according to its Swedish national grid coordinates, RT90, where one unit = 1 m.
All ‘Open’ sites were grazed wooded meadows (Fig. 2a). Lime dominated only one site. In the other sites lime was mixed with other coarse trees, mainly oaks. The ‘Re-grown’ sites were similar to the ‘Open’ ones, differing only inasmuch that they had re-grown with young deciduous trees (or in one case, planted spruce) some 40–70 years ago (Fig. 2b). Two of the ‘Re-grown’ sites had been recently opened up as a nature conservation measure by thinning out the younger trees. As this was done only 1 year before sampling, they were still classified as ‘Re-grown’ since the fauna was assumed to need some years to respond to the opening-up of the habitat. On several of the ‘Re-grown’ sites it was evident that there had been many more old lime trees some decades before as there were circles of sprouting stems from the remnants of former stumps. The ‘Park’ sites were either avenues in parks (n = 6) (Fig. 2c), along roads (n = 1), or a mixture of these (n = 1) at manor houses in the countryside.

Three categories of sites were studied: a ‘Open’ sites, which were grazed wooded meadows, b ‘Re-grown’ sites, which were wooded meadows re-grown with forest 40–60 years ago, c ‘Park’, which were avenues in parks or along roads at manor houses in the countryside
The number of hollow lime trees in total was included in the analysis as a measure of the size of the sites. For 16 of the sites data on this were obtained from “the tree gateway” (www.tradportalen.se, on the 18th of March 2011) which is a web-based database for collecting reports on veteran trees and other trees worthy of protection. Inventories made by county administrative boards are usually included. For the remaining nine sites the number of trees was estimated from our field visits when doing the beetle inventory, in three of the baroque parks with some help of web-based satellite images on which crowns of alley trees are distinguishable. This data has a lot of apparent uncertainties as several persons have collected the data. Furthermore, somewhat different criteria seems to have been used for which trees to include. Therefore, the data was categorised in three classes (Table 1). Also the total number of hollow trees was counted, but not included in analyses because this measure had the same problem with uncertainties and was strongly correlated to the number of lime trees.
Sampling of beetles
At each site, four lime trees with a high potential to harbour a rich saproxylic beetle fauna were selected on which window traps were placed to catch beetles. Thus, selected trees should preferentially be coarse and hollow. If possible, trees of somewhat different types were selected, although choice was limited at sites where there were few trees to choose from.
On each tree a window trap was placed, as close to a hollow as possible. The window was 30 × 60 cm with a tray underneath, filled with 50% propylene glycol and 50% water. The traps were placed between 2.5 and 5 m above ground, mainly to avoid damage from cattle or people. The traps were active during the summer season between May and late August or early September in the years 2006–2008, except for Skokloster and Drottningholm, which were inventoried in 2001 and 2004, respectively. Year is included as a variable in the analyses since there might be variation among years.
Tree circumference at breast height was measured with a tape at most sites (Table 1). However, at six sites the circumferences were only estimated visually, by multiplying estimated diameter with pi. The average circumference of trees at all sites was 295 cm (range 189–465 cm per site). The corresponding maximum circumferences per site were 406 cm (range 235–628 cm).
All trapped saproxylic beetles were determined to species level according to the nomenclature of Lundberg and Gustafsson (1995). However, some difficult groups were only determined to genus: Cryptophagus, Euplectus, Atomaria, Corticaria and most species within the sub-family Aleocharinae. Species were categorised as saproxylic or non-saproxylic, and as being associated with hollows, wood and bark, or with sap-runs, according to published information (Hansen 1964; Koch 1989–1992; Palm 1959). Species living in nests of birds and hymenopterans were classified as being associated with hollows, while species living on the fruiting bodies of saproxylic fungi were classified as wood and bark living species. Red-listed species were defined according to Gärdenfors (2010).
Statistics
Among the three site-categories, the average numbers of species per site were compared in general linear regression models. All environmental variables (Table 1) were tested univariately, the most significant variable being added to the regression model by forward selection until no further variable could add significantly (P < 0.05) to the model if added last. As a check the selections were also made with automatic backward elimination. The software used was JMP for Mac ver 8.0.1.
Species composition was analysed by ordination. Species data, i.e. the numbers of individuals of each species, were square root transformed as recommended for count data (Leps and Smilauer 2003). The variable ‘type’ was transformed into two dummy variables, as the ordination technique used is only able to work with dichotomous categorical variables. Thus, the variable ‘Park’ became (‘Park’/‘not Park’), and ‘Open’ became (‘Open’/‘not Open’). The results are presented graphically using correspondence analysis (CA), with the effects of environmental parameters being shown with respect to an indirect gradient analysis, i.e. an analysis that shows environmental effects on an ordination that only takes species data into account. To test whether a variable might significantly explain species composition, a direct gradient analysis was used, canonical correspondence analysis (CCA), which maximises the explanatory power of environmental variables. The significance of the variables was tested with a Monte Carlo simulation, run with 499 iterations. The software used was CANOCO 4.5 (Braak and Smilauer 1998).
How much individual species were associated with a site ‘type’ was tested with indicator species analysis (IndVal) (Dufrene and Legendre 1997). This analysis gives a value of 100 for a perfect indicator which means a species that occur on all sites with in a category (type) and not on any other site. Bad indicators get a value near 0. With 15,999 permutations in a Monte Carlo test the statistical significance of the indicator values were calculated under the null hypothesis that the indicator value is not larger than would be expected by chance. Species present on four or more sites (n = 164) were analysed. PcOrd 6.0 was used for the calculations.
Results
In total 14,460 individuals of 323 saproxylic beetle species were found (Table 2). Of these, 56 were classified as living in hollows, and 259 as living in wood and bark. The eight remaining species live in sap-runs, but this category had too few species to allow further statistical analyses. Of all saproxylic species, 50 were red-listed (Table 2).
Number of species
‘Open’ sites had the highest average number of species per site for all combinations of red-listed and non-red-listed species and substrate associations (Fig. 3). However, it was significantly higher than another category ‘Park’ only when “all saproxylic species” and “all wood and bark species” were compared (Fig. 3a, c; Table 3). Regarding species associated with hollows and red-listed species, the number of species in ‘Park’ was intermediate between ‘Open’ and ‘Re-grown’ sites, although these differences were not statistically significant (Fig. 3b, d–f; Table 3).

The average number of beetle species in the three stand types under comparison: a all saproxylic species, b species living in hollows, c species living in wood and bark, d all red-listed saproxylic species, e red-listed species in hollows, f red-listed species in wood and bark. Significant differences were found in (a) and (c) (see Table 3). Number of sites were: ‘Open’ n = 8, ‘Re-grown’ n = 11, ‘Park’ n = 8
Species numbers in most categories decreased significantly with the variable ‘RT90N’, i.e. a northward decline in number of species (Table 3). Numbers of species associated with hollows declined in an eastward direction, although this was only marginally significant. ‘Year’ was a significant variable for all species and for all wood and bark associated species. This difference was mainly caused by there being few species present in 2004 compared to 2007. In 2004, a park (Drottningholm) was the only surveyed site, whereas in 2007 many sites in the southwestern part of the study region were surveyed. The two measures of trunk circumference did not, in five out of the six cases, significantly explain species number. The exception was red-listed species associated with hollows, which was significant when also the variable ‘RT90N’ was included (Table 3). The number of lime trees on a site had a significantly negative relationship to all species and all wood and bark species. ANOVA failed to show any significant association (df = 24: RT90N, P = 0.44; RT90E, P = 0.78) between the two coordinate variables and the ‘type’ of the locality (Fig. 1).
Species composition
Species composition was significantly affected by site ‘type’ (Fig. 4; Table 4). Both ‘Park’ and ‘Open’ were significantly correlated with species composition for all three tested groups of species. However, the north–south gradient had an even stronger explanatory power (Table 4). The tree circumference variables were significantly correlated with species composition in one case each (Table 4).

Ordination plots of a all saproxylic species, b species living in hollows, where the different sites are ordinated only due to species data (CA) and environmental variables assigned in an indirect gradient analysis. Statistical significances of variables are calculated in a CCA (Table 4)
Of the 164 species having the minimum number of occurrences for being tested (four or more), the IndVal analyses showed 16 species to be significantly associated with one of the site-‘types’, while 148 species showed no association (Table 5). When ‘Open’ and ‘regrown’ were pooled to ‘non-Park’ and compared to ‘Park’ nine species showed significant association and 155 no association (Table 5). Among the significantly associated species, three were living in hollows (Table 5) and all these three were mainly found in ‘Park’.
Discussion
For saproxylic beetle species living in tree hollows and for red-listed saproxylic beetles species, species numbers did not differ between parks and the more natural sites. Also for species associated with wood and bark rather high numbers were found in the ‘Park’ sites, but their numbers were significantly lower than in the ‘Open’ sites. This shows that the old trees in parks harbour a rich fauna in spite of the more intensive management. The removal of wood from parks probably explains the significantly lower number of species associated with wood and bark. However, even among them, the red-listed species showed no such pattern, indicating that they could be living within the dead wood still attached to the living parts of old park trees.
Although the ordination revealed the species composition in ‘Park’ sites to be significantly different from other sites, few species discriminated between the two types of sites. The nine species showing association to ‘Park’ versus ‘non-Park’ is just one more than the number expected from the mass-significance effect (5% of the 164 tested species = 8.2). Expanding the analysis to compare all three ‘types’ gave 16 associated species, which is still marginally more than expected from mass-significance. Thus, the analysis shows that parks can be useful sites for almost all the species encountered in this study.
Sverdrup-Thygeson et al. (2010) found parks to be species-rich sites for the saproxylic beetle fauna of hollow oaks. However, their definitions differed from those adopted in the present study since their ‘Park’ would have included the sites defined as ‘Open’ in this paper. Using a similar definition to that used in the present study, they found ‘Open’ sites to have the same numbers of red-listed species as ‘forest’ sites. However, their ‘Open’ sites had a higher proportion of species associated with hollows, which agrees with results from a study of Swedish oaks (Ranius and Jansson 2000). This suggests that regarding the hollow–dwelling species in the present study, the insignificantly higher numbers found in ‘Open’ sites compared to the ‘Re-grown’ sites was more likely to be due to the low power of the analysis, rather than any lack of a real difference. Conversely, since lime is a shade-tolerant tree it might be expected to harbour a fauna comprising fewer species adapted to sun-exposed habitats (Gärdenfors and Baranowski 1992). However, most species associated with hollows are not specific to certain tree species, and there are probably more species on lime that prefer exposed habitats than prefer shaded habitats. In the parks, the positive effect of openness seems to compensate for the negative effects from the removal of dead wood.
A problem with comparing sun-exposed sites to more shaded is that the catchability of beetles in open traps might be higher in sun-exposed sites as insect activity often is larger at higher temperature. Usually this effect is not considered at all (e.g. Sverdrup-Thygeson et al. 2010) or just assumed to be low with no reference to data (e.g. Ranius and Jansson 2000). However, Wikars et al. (2005) found that window trapping and methods sampling directly from the wood gave similar relations in species numbers in sun-exposed and shaded environments. Thus, the assumption of low difference in catchability seems true, but more studies would be valuable and could easily be conducted by analysing already collected data.
In this paper no sites were included that could be categorised as forest because old lime trees in the region almost always grow on sites that were part of an agricultural landscape a 100 years ago, i.e. wooded meadows. For trees that exhibit traces of having been pollarded, any other situation is extremely unlikely. But trees with no such traces might originally have grown in sites that resembled forest, but which were grazed by cattle, so keeping them more open than forests are today (Emanuelsson 2009).
Generally, tree circumference explained little in this study. This is somewhat surprising as tree diameter has previously been shown to be positively correlated with the number of species (Grove 2002; Ranius and Jansson 2000; Sverdrup-Thygeson et al. 2010). However, in the present study, the trap catches and the circumferences are estimates relevant on stand scale rather than on the scale of individual trees. Therefore, other variables might have confounded the results. Furthermore, all sites were characterised by trees that had reached a size and age defining them as ancient, and the degree of ancientness may be more important than diameter itself. Pollarding slows down growth and because of that, thin trunks may be ancient trees. In oaks, 50% of trees form hollows by about 250 years of age (Ranius et al. 2009). For lime trees, this age is probably lower, as lime rots faster than oak and especially so in pollarded trees as the formation of hollows is enhanced where branches are shed. However, hollowness need not imply a rich fauna if the trees are too young, as seen in the case of 80-year old hollow limes in the park at Drottningholm, which had fewer species, especially red-listed species, than the old limes in the same park (Jonsell 2008).
The amount of habitat, measured as number of hollow lime trees on each site (No. of trees), had significant relationship to species number for all wood and bark living species, and it was negative. This lack of relation, or relation opposite to what should be expected, could be due to that the variables no. of trees and type were confounded with somewhat more trees in parks than in the other type of sites (2.6 compared to 1.9 for the two others). Also problems with quantifying this variable may contribute. First the data collected for each locality had several uncertainties in itself (see “Materials and methods”). The numbers obtained also give just the present situation, totally disregarding the history of the site. In addition to that, the definition of where the borders for a locality should be drawn is also problematic. Most of these sites are found in regions where old hollow trees may occur here and there. Data on suitable trees for the whole landscape with estimates of connectivity related to distance to each of these occurrences should probably be more explanatory (Ranius et al. 2010). Such an analysis would probably suggest that the rich saproxylic beetle fauna on several sites in the Mälaren area is due to a dense patchwork of sites. The number of sites is high, there is a high connectivity between them, several sites are large and the individual trees in them are often a high quality habitat, all factors that contribute to a sustainable metapopulation system (Hanski 1994; Ranius 2007). Two practical conclusions from this is that every park with old trees constitutes a site that contributes to the sustainability of the metapopulation and that the number and quality of trees within each park also contributes to this.
The north–south coordinate was a strong explanatory variable both for species numbers and species composition. This is not surprising because, compared to the sites north of the lake, the area around lake Mälaren is both climatically favourable (Raab and Vedin 1995) and has a high density of sites with old trees. Mälaren has been identified as a diversity hot-spot for saproxylic beetles (Ehnström and Waldén 1986), with the western part of Mälaren regarded as being especially species-rich. This was only weakly supported by the results of the present study, as the variable RT90E (west–east coordinate) had low explanatory power.
Practical implications
The high conservation value of parks for saproxylic insects shown in this study is dependent on the retention of old trees. Thus, the total rejuvenation of trees, which is considered in some parks, would be fatal to the resident fauna. However, all trees will sooner or later die, or they have to be removed for safety or aesthetic reasons. If they individually and continuously are replaced when they die there will be a continuous supply of new trees growing into the ancient-tree age class which in turn means a continuous supply of suitable habitat for the saproxylic insects.
On a short term a good measure is to retain trees, or parts of trees, that are cut or fallen in a “tree-graveyard” situated in a remote part of the park, where it does not conflict with the aesthetic values. Such graveyards is both a chance for insects to finalise their development and a habitat patch that can be colonised (Aulén and Franc 2008). However, compared to the management aiming at a long term continuous supply of old trees, this is of minor importance, both because its’ short term effect and because most of the valuable contributions to the graveyard emanate from the old trees.
As almost all lime trees in parks, and many lime trees in the more natural sites, were originally pollarded, they are at risk of breaking apart when the shoots from the last pollarding are allowed to grow into large trees. This was observed on several of the sites in this study. The risk of breakage is especially great in re-grown sites where the closer canopy gives less light to the trees, which in turn decreases the production of carbohydrates needed for building a stable trunk. For keeping these old trees alive it is important to continue pollarding. However, old trees that has not been managed for a long time need careful treatment when management is resumed (Slotte 1997; Wisenfield 1995). A strong reduction of the crown by cutting all large branches may be fatal. As pollarding is an expensive measure, it is important that it should only be done on sites where there is the potential to retain the associated fauna and flora, i.e. where one can forecast a continuous supply of old trees in the future. Most of the parks in the present study do have this potential due to the continuous replacement of trees that die.
This study has shown that old trees in parks are a very valuable substrate for saproxylic beetles and thus, they may contribute to the long-term survival of these species. In the management of the parks, however, conservation of the biological values can be in conflict with measures undertaken to enhance or preserve historical and cultural values (Jonsell 2008) or measures for public safety (=risk for falling parts of trees) (Carpaneto et al. 2010). Such conflicts often arise because park managers are not always aware of the high biological value of elements within their domain and vice versa. The biologists may not reflect over the heavy cultural influence on the habitat of “their” species. Thus, effective communication between those with different interests is important in order to share knowledge about different sets of values so that both the cultural and biological value of a site can be maintained for the benefit of all.
References
Ahti T, Hämet-Ahti L, Jalas J (1968) Vegetation zones and their sections in north–western Europe. Annales Botanici Fennici 5:169–211
Andersson H (2010) Inventering av insekter i fem linddominerade lokaler i Västmanlands län 2006. Länsstyrelsens rapportserie. Rapport 2010:7, Västerås
Aulén G, Franc N (2008) Hänsynsyta på hygge, förstärkt med mer död ved, blev “nyckelbiotop” med 39 rödlistade skalbaggsarter (A consideration area on a felling site enhanced with dead wood, became a woodland key habitat with 39 redlisted beetle species (Coleoptera) species). Entomologisk Tidskrift 129:53–68
Bengtsson R (2005) Variation in common lime (Tilia × europaea) in Swedish gardens of the 17th and 18th centuries. PhD thesis, Swedish University of Agricultural Sciences, Alnarp
Braak CJFt, Smilauer P (1998) Reference manual and user’s guide to CANOCO for Windows: software for canonical community ordination (version 4). Microcomputer Power, Ithaca
Carpaneto GM, Mazziotta A, Coletti G, Luiselli L, Audisio P (2010) Conflict between insect conservation and public safety: the case study of a saproxylic beetle (Osmoderma eremita) in urban parks. J Insect Conserv 14:555–565
Dufrene M, Legendre P (1997) Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol Monogr 67:345–366
Ehnström B (2006) Åtgärdsprogram för skalbaggar på skogslind. Rapport 5552, Naturvårdsverket, Stockholm
Ehnström B, Waldén HW (1986) Faunavård i skogsbruket–den lägre faunan. (The protection and management of endangered and declining invertebrate species in Swedish woodlands). Skogsstyrelsen, Jönköping
Eliasson P, Nilsson SG (2002) ‘You should hate young oaks and young noblemen’. The environmental history of oaks in eighteenth- and nineteenth-century Sweden. Environ Hist 7:659–677
Emanuelsson U (2009) The rural landscapes of Europe. How man has shaped European nature. Formas, Stockholm
Gärdenfors U (2010) The 2010 red list of Swedish species. ArtDatabanken, SLU, Uppsala
Gärdenfors U, Baranowski R (1992) Skalbaggar anpassade till öppna respektive slutna ädellövskogar föredrar olika trädslag (Beetles living in open deciduous forests prefer different tree species than those living in dense forests). Entomologisk Tidskrift 113:1–11
Gerell R (2000) Alléernas betydelse för rödlistade vedlevande skalbaggar (The importance of avenues for threatened saproxylic beetles). Entomologisk Tidskrift 121:59–66
Grove SJ (2002) Saproxylic insect ecology and the sustainable management of forests. Annu Rev Ecol Syst 33:1–23
Hansen V (1964) Fortegnelse over Danmarks biller 1.og 2.del. (Catalogue of the Coleoptera of Denmark 1 and 2nd part). Entomologiske Meddelelser 33:1–507
Hanski I (1994) A practical model of metapopulation dynamics. J Anim Ecol 63:151–162
Jonsell M (2004) Old park trees: a highly desirable resource for both history and beetle biodiversity. J Arboric 30:238–244
Jonsell M (2008) Vedlevande skalbaggar i Drottningholms slottspark. Entomologisk Tidskrift 129:103–119
Jonsell M, Sahlin E (2010) Inventering av vedlevande skalbaggar på lindar Södermanlands, Uppsala och Västmanlands län. Rapport, länsstyreslen i Västmanlands län, Västerås
Koch K (1989–1992) Die Käfer Mitteleuropas. Ökologie. Band 1–3. Goecke & Evers, Krefeld
Leps J, Smilauer P (2003) Multivariate analysis of ecological data using CANOCO. Cambridge University Press, Cambridge
Lundberg S, Gustafsson B (1995) Catalogus Coleopterorum Sueciae. Naturhistoriska riksmuseet, Stockholm
Oleksa A, Ulrich W, Gawronski R (2006) Occurrence of the marbled rose-chafer (Protaetia lugubris Herbst, Coleoptera, Cetoniidae) in rural avenues in northern Poland. J Insect Conserv 10:241–247
Palm T (1959) Die Holz-und Rindenkäfer der süd-und mittelschwedischen Laubbäume. Opuscula Entomologica Supplementum 16:1–374
Raab B, Vedin H (1995) Climate, lakes and rivers. National atlas of Sweden, Bra Böcker, Höganäs
Ranius T (2007) Extinction risks in metapopulations of a beetle inhabiting hollow trees predicted from time series. Ecography 30:716–726
Ranius T, Jansson N (2000) The influence of forest regrowth, original canopy cover and tree size on saproxylic beetles associated with old oaks. Biol Conserv 95:85–94
Ranius T, Niklasson M, Berg N (2009) Development of tree hollows in penduculate oak (Quercus robur). For Ecol Manag 257:303–310
Ranius T, Johansson V, Fahrig L (2010) A comparison of patch connectivity measures using data on invertebrates in hollow oaks. Ecography 33:971–978
Sernander R (1926) Stockholms natur. Almqvist & Wiksells förlag, Uppsala
Slotte H (1997) Hamling–historisk tillbakablick och råd för naturvårdare (Pollarding - a look back on its history, and advice to nature conservationists). Svensk Botanisk Tidskrift 91:1–21
Sörensson M (2008) AHA–en enkel metod for prioritering av vedentomologiska naturvärden hos träd i sydsvenska park-och kulturmiljöer (AHA – a simple method for evaluating conservation priorities of trees in South Swedish parks and urban areas from an entomo-saproxylic viewpoint). Entomologisk Tidskrift 129:81–90
Speight MCD (1989) Saproxylic invertebrates and their conservation. Council of Europe, Strasbourg
Sverdrup-Thygeson A, Skarpaas O, Ødegaard F (2010) Hollow oaks and beetle conservation: the significance of the surroundings. Biodivers Conserv 19:837–852
Warren MS, Key RS (1991) Woodlands: past, present and potentials for insects. In: Collins NM, Thomas JA (eds) The conservation of insects and their habitats, 15th Symp. of R. Entomol. Soc. London. Academic Press, London, pp 155–211
Wikars L-O, Sahlin E, Ranius T (2005) A comparison of three methods to estimate species richness of saproxylic beetles (Coleoptera) in logs and high stumps of Norway spruce. Can Entomol 137:304–324
Wisenfield J (1995) Experience at Hatfield Forest, Essex, with restoration of old pollards and establishment of new ones. Biol J Linn Soc 56(Suppl):181–183
Acknowledgments
Thanks to Erik Sahlin, Rickard Sellberg, Karin Andersson and Markus Rehnberg for their help with managing the traps in the field. Thanks also to Gunnar Sjödin and Erik Sahlin who determined a large proportion of the trapped insects; to Thomas Ranius for his valuable comments on an earlier version of the manuscript; and to Jonas Victorsson for help with the IndVal analyses. The author gratefully acknowledges financial support from FORMAS for the analysis of the material and the writing of the paper.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Jonsell, M. Old park trees as habitat for saproxylic beetle species. Biodivers Conserv 21, 619–642 (2012). https://doi.org/10.1007/s10531-011-0203-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-011-0203-0