
Abstract
The little fire ant, Wasmannia auropunctata, is an invasive species known to negatively affect many native species in its introduced range. Here, we studied the effect of little fire ant presence on the biomass of soil fauna in rainforest of New Caledonia to assess if little fire ants have an impact on an endangered endemic flightless bird, the kagu, Rhynochetos jubatus. The average biomass of soil fauna (except for millipedes) was lower in 53 sample plots with little fire ants than in 597 sample plots without little fire ants and the biomass decreased with increasing density of little fire ant nests. Millipedes on the contrary seemed to be positively impacted as their average biomass was higher in areas with fire ants than in areas without, but even millipede biomass decreased with increasing nest density of fire ants. Kagu density decreased with increasing area of rainforest invaded by little fire ants, suggesting, given their observed negative impact on soil fauna biomass, that they may limit food resources for kagu. Little fire ants might therefore transform rainforest into a less suitable habitat for the kagu by reducing the biomass of their prey and thus have a negative impact for kagu conservation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Ants are one of the most ecologically successful and numerically dominant insect groups, especially in tropical terrestrial ecosystems, in which they play key roles as predators, saprophages, nutrient recyclers, soil engineers and regulators of plant growth and reproduction (Del Toro et al. 2012). Nonetheless, some ants are known as highly invasive species that interfere with many components of ecosystems (Holway et al. 2002; Clarke et al. 2021). The little fire ant, Wasmannia auropunctata, is considered as one of the worst invasive ant species (Jourdan 1997; Dunham and Mikheyev 2010; Vonshak et al. 2010), threatening native ants (Le Breton et al. 2003; Walker 2006; Bertelsmeier et al. 2015), crickets (Gasc et al. 2018; Anso et al. 2022), lizards (Jourdan et al. 2001), and even mammals (Walsh et al. 2004). Little fire ants are expected to also threaten other taxonomic groups (Lubin 1984; Duham and Mikheyev 2010; Anso et al. 2022) and to promote sap-sucking insects (Le Breton et al. 2005), but there has not yet been a comprehensive study of their impact on the entire soil fauna.
Little fire ants were unintentionally introduced to New Caledonia in the early 1960s and began spreading throughout the islands of the whole archipelago across most habitats including native rainforest (Jourdan 1997; Le Breton et al. 2003). As the species lacks an efficient nuptial flight and has clonal reproduction (Fournier et al. 2005), it has mainly been spread by human activity (Walsh et al. 2004). In New Caledonia, it has been spread by soil or firewood to rainforest reserves, in which their further colonisation speed is about 10–20 m per year (H. Jourdan, unpublished data). Since the introduction of little fire ants, research in New Caledonia has focussed on their biology (Jourdan 1997, 1999; Jourdan et al. 2001; Le Breton et al. 2003, 2004, 2005). Here, we use data collected during the study of an endangered flightless bird, the kagu, Rhynochetos jubatus (Eurypygiformes: Rhynochetidae). The kagu is the only species of its family, the only extant flightless bird of New Caledonia, and unique also due to its complex social organisation otherwise found only in human societies (Theuerkauf et al. 2018). We aim at demonstrating that little fire ants not only threaten other ant species but also change the soil fauna composition in general and potentially transform rainforest into a less suitable habitat for the kagu by reducing the biomass of their prey. This degradation of habitat may have severe conservation impacts for the kagu as human colonisation of New Caledonia 3000 years ago already drove kagu into suboptimal habitats and caused an about 30% reduction in body mass (Theuerkauf and Gula 2018). The focus in our kagu study was assessing food abundance in sample plots within home ranges of 34 kagu families in two protected sites of New Caledonia, which represent poor and productive kagu habitats (Theuerkauf et al. 2017). As the food of kagu includes nearly the whole soil fauna larger than about 5 mm of length (with the exception of ants and mites, authors’ unpublished data of cafeteria tests), our sampling design allowed us to also directly assess the impact of little fire ants on the soil fauna.
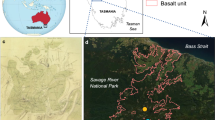
We conducted our research in two protected areas, the Parc Provincial de la Rivière Bleue (PPRB, 22°3–12’S, 166°33–46’E), to which little fire ants were introduced around 1997, and the Parc Provincial des Grandes Fougères (PPGF, 21°30–39’S, 165°39–50’E), to which the species was introduced around 1984. The PPRB is a 221-km² reserve on ultramafic soils with a mean annual rainfall of about 2900 mm and mean annual temperature of 21 °C. Ultramafic soils are extremely low productive and rich in heavy metals. We worked in rainforests at altitudes of 150–350 m asl. The PPGF is a 45-km2 reserve on oligotrophic schist soils, with a mean annual rainfall of about 1,600 mm and a mean annual temperature of 20 °C. We worked in rainforests at altitudes of 300–500 m asl.
To ensure that the sample plots were representative for each kagu family, we predefined straight transects dissecting the family’s home range and spaced sample plots at 25-m distances along the transect. From 2008 to 2021, we assessed the abundance of soil fauna larger than about 5 mm in 650 sample plots of 1 m2 (delimited by a 1 m × 1 m frame) by hand inspecting the entire litter transferred to white plastic bowls during the hot wet season (December to April, n = 120 sample plots), the cool wet season (April to August, n = 170) and the warm dry season (August to December, n = 370). Little fire ants were present in 10% of sample plots in the ultramafic site (n = 310) and 6.5% of sample plots in the non-ultramafic site (n = 340). The sample plots were distributed representatively, so these proportions correspond approximately to the area colonised by little fire ants in each study site.
Because there were seasonal differences in soil fauna abundance (Fig. 1a), we normalised biomass with seasonal correction factors. We calculated seasonal correction factors for each study site separately as the average abundance over the whole year divided by the average abundance in a given season. During inspection of sample plots, we counted the number of individuals in each taxonomic order to assess abundance. We weighed some of the captured individuals alive with electronic scales to the next 0.001 g and calculated average mass of an individual for each order (Table 1). We then calculated biomass of individual sample plots as the sum of products of average mass and number of individuals in each order. We estimated home ranges of the 34 kagu families as 100% Minimum Convex Polygons (MCP) of locations taken during our kagu radio-tracking study (Theuerkauf et al. 2018). We then calculated density as the number of kagu in a family divided by the home range size. Because our two study sites were of different productivity resulting in different kagu density, we standardised kagu density by dividing the density of each family by the average kagu density in the respective study site.

Average (with 95% confidence intervals) biomass of soil fauna (A) in sample plots during the wet hot (n = 120), the wet cool (n = 170) and the dry warm (n = 370) season in the ultramafic study site (grey bars) and the non-ultramafic study site (white bars), (B) seasonally normalised biomass of Julida (dotted bars) and non-Julida soil fauna in sample plots with little fire ant presence (n = 53) and absence (n = 597) in the ultramafic (grey bars) and the non-ultramafic study site (white bars), (C) seasonally normalised biomass of all soil fauna in relation to the number of little fire ant nests (dotted area represents the proportion of Julida) on sample plots colonised by fire ants (n = 53), and (D) exponential regression (y = 0.978e−1.452x, F = 6.213, R² = 0.163, P = 0.018) between the proportion of sample plots with little fire ants and relative kagu density (proportion of average in the respective study site) of 34 kagu families in the ultramafic (grey circles) and the non-ultramafic site (white circles)
In both study sites, the biomass of soil fauna (except for Julida) was at least twice as high in sample plots without little fire ants than in sample plots with little fire ants (Fig. 1b, Table 1) and the biomass decreased with increasing nest density of little fire ants (Fig. 1c). Phasmatodea, Orthoptera, Blattodea, Haplotaxida, Araneae and Amphipoda appeared to be the most negatively impacted by the presence of ants (Fig. 2). Julida on the contrary seemed to be positively impacted as their biomass was higher in areas with fire ants than in areas without (Figs. 1b, 2, Table 1). However, with increasing densities of little fire ant nests, Julida numbers started decreasing as well (Fig. 1c). There appeared to be a trend for lower relative kagu density with an increasing proportion of the home range invaded by little fire ants (Fig. 1d).

Ratio (with confidence interval) of abundance in sample plots with little fire ants (n = 53) divided by abundance in sample plots without little fire ants (n = 597) for each order of soil fauna (in parentheses total number of individuals in all plots). Negatively impacted orders in red (confidence interval entirely below 1), positively impacted in blue (confidence interval above 1) and neutral in grey (confidence interval including 1)
Most of the documented effects of invasive ants on native invertebrate species are direct predation or competition for reproductive sites (Holway et al. 2002). Due to the high abundances that invasive ants can achieve, they can cause population declines or local extinctions of invertebrates (Clarke et al. 2021). We demonstrated an important difference in the composition of soil fauna in areas colonised by little fire ants and in areas without, which has also been observed in New Caledonian canopy fauna (Jourdan 1999). As there might be confounding environmental factors and we did not use an experimental design with little fire ant exclusion to prove changes (Kumschick et al. 2014), our results are no direct proof for changes caused by little fire ants. However, we think, given the relatively large sample size and the repetition in two sites, that our study provides sufficient indication for a probable effect of fire ants on soil fauna composition. This change in community structure may even lead to cascading effects at higher trophic levels as pointed out by Jourdan et al. (2001) for endemic litter-dwelling skinks and some arboreal geckos. The negative effect on reptiles in our study was less obvious, probably because the most often observed reptiles were lizards of the genus Nannoscincus, which are burrowers and use the lower layer in the litter (Sadlier et al. 2014), whereas little fire ants use the upper layer for both foraging and nesting (Orivel et al. 2009). We might also have underestimated the negative effect if some lizards escaped the sample plots before we could spot them.
Our study demonstrates likely ant-associated declines in the abundance of various orders of soil fauna. This results in a decrease in soil fauna biomass that is somewhat mediated by the increase in Julida biomass. It is likely that the Julida biomass increase was caused by the suppression of other predators through little fire ants in early phases of the invasion. However, even Julida biomass decreased with increasing density of little fire ants, which in later phases of the invasion could lead to a reduction of food resources for kagu. The observed trend of higher relative kagu density in areas with a smaller proportion of area invaded by little fire ants indicates that the presence of the ants probably reduces resources for kagu. As the reproductive output of kagu is relative to resources (Theuerkauf et al. 2017), colonisation by little fire ants may indirectly limit kagu population growth and therefore may have a negative impact for kagu conservation as long as the little fire ants can maintain high abundance.
Data availability
The datasets generated during the current study are available from the corresponding author on reasonable request.
References
Anso J, Gasc A, Bourguet E, Desutter-Grandcolas L, Jourdan H (2022) Crickets as indicators of ecological succession in tropical systems, New Caledonia. Biotropica: Accepted for publication
Bertelsmeier C, Avril A, Blight O, Confais A, Diez L, Jourdan H, Orivel J, Saint Germès N, Courchamp F (2015) Different behavioural strategies among seven highly invasive ant species. Biol Inv 17:2491–2503
Clarke DA, Palmer DJ, McGrannachan C, Burgess TI, Chown SL, Clarke RH, Kumschick S, Lach L, Liebhold AM, Roy HE, Saunders ME, Yeates DK, Zalucki MP, McGeoch MA (2021) Options for reducing uncertainty in impact classification for alien species. Ecosphere 12:e03461
Del Toro I, Ribbons RR, Pelini SL (2012) The little things that run the world revisited: a review of ant-mediated ecosystem services and disservices (Hymenoptera: Formicidae). Myrmecol News 17:133–146
Dunham AE, Mikheyev AS (2010) Influence of an invasive ant on grazing and detrital communities and nutrient fluxes in a tropical forest. Divers Distrib 16:33–42
Fournier D, Estoup A, Orivel J, Foucaud J, Jourdan H, Le Breton J, Keller L (2005) Clonal reproduction by males and females in the little fire ant. Nature 435:1230–1234
Gasc A, Anso J, Sueur J, Jourdan H, Desutter-Grandcolas L (2018) Cricket calling communities as an indicator of the invasive ant Wasmannia auropunctata in an insular biodiversity hotspot. Biol Inv 20:1099–1111
Holway DA, Lach L, Suarez AV, Tsutsui ND, Case TJ (2002) The causes and consequences of ant invasions. Annu Rev Ecol Syst 33:181–233
Jourdan H (1997) Threats on Pacific islands: the spread of the tramp ant Wasmannia auropunctata (Hymenoptera: Formicidae). Pac Conserv Biol 3:61–64
Jourdan H (1999) Dynamique de la biodiversité de quelques écosystèmes terrestres néo-calédoniens sous l’effet de l’invasion de la fourmi peste Wasmannia auropunctata (Roger), 1863 (Hymenoptera: Formicidae). PhD Dissertation, Université Paul Sabatier, Toulouse, https://core.ac.uk/download/pdf/39848517.pdf
Jourdan H, Sadlier RA, Bauer AM (2001) Little fire ant invasion (Wasmannia auropunctata) as a threat to New Caledonian lizards: evidences from a sclerophyll forest (Hymenoptera: Formicidae). Sociobiology 38:283–301
Kumschick S, Gaertner M, Vilà M, Essl F, Jeschke JM, Pyšek P, Ricciardi A, Bacher S, Blackburn TM, Dick JTA, Evans T, Hulme PE, Kühn I, Mrugała A, Pergl J, Rabitsch W, Richardson DM, Sendek A, Winter M (2014) Ecological impacts of alien species: quantification, scope, caveats, and recommendations. Bioscience 65:55–63
Le Breton J, Chazeau J, Jourdan H (2003) Immediate impacts of invasion by Wasmannia auropunctata (Hymenoptera: Formicidae) on native litter ant fauna in a New Caledonian rain forest. Aus Ecol 28:204–209
Le Breton J, Delabie J, Chazeau J, Dejean A, Jourdan H (2004) Experimental evidence of large scale unicoloniality in the tramp ant Wasmannia auropunctata. J Insect Behav 17:263–271
Le Breton J, Jourdan H, Chazeau J, Orivel J, Dejean A (2005) Niche opportunity and ant invasion: the case of Wasmannia auropunctata in a New Caledonian rain forest. J Trop Ecol 21:93–98
Lubin YD (1984) Changes in the native fauna of the Galápagos Islands following invasion by the little red fire ant, Wasmannia auropunctata. Biol J Linn Soc 21:229–242
Orivel J, Grangier J, Foucaud J, Le Breton J, Andrès FX, Jourdan H, Delabie JHC, Fournier D, Cerdan P, Facon B, Estoup A, Dejean A (2009) Ecologically heterogeneous populations of the invasive ant Wasmannia auropunctata within its native and introduced ranges. Ecol Entomol 34:512–523
Sadlier RA, Bauer AM, Wood PL, Smith SA, Whitaker AH (2014) Cryptic speciation in the New Caledonian lizard genus Nannoscincus (Reptilia: Scincidae) including the description of a new species and recognition of Nannoscincus fuscus Günther. Zoologia Neocaledonica 8. Biodiversity studies in New Caledonia. Mém Mus Natl Hist Nat 206:45–68
Theuerkauf J, Gula R (2018) Indirect evidence for body size reduction in a flightless island bird after human colonisation. J Ornithol 159:823–826
Theuerkauf J, Haneda T, Okahisa Y, Sato NJ, Rouys S, Bloc H, Ueda K, Watanabe I, Kuehn R, Gula R (2017) Elevated concentrations of naturally occurring heavy metals inversely correlate with reproductive output and body mass of the Kagu Rhynochetos jubatus. Ibis 159:580–587
Theuerkauf J, Kuehn R, Rouys S, Bloc H, Gula R (2018) Fraternal polyandry and clannish spatial organisation in a flightless island bird. Curr Biol 28:1482–1488
Vonshak M, DayanT I-H, Freidberg A, Hefetz A (2010) The little fire ant Wasmannia auropunctata: a new invasive species in the Middle East and its impact on the local arthropod fauna. Biol Inv 12:1825–1837
Walker KL (2006) Impact of the little fire ant, Wasmannia auropunctata, on native forest ants in Gabon. Biotropica 38:666–673
Walsh PD, Henschel P, Abernethy KA, Tutin CEG, Telfer P, Lahm SA (2004) Logging speeds little red fire ant invasion of Africa. Biotropica 36:637–641
Acknowledgements
We thank Emilie Baby, Hugo Bourgogne, Alice Bousseyroux, José Brinon, Fabien Condamine, Hélène De Méringo, Mathieu Donnat, Marjorie Levant, Rose-Mai M’Boueri, Mathilde Méheut, Jean Roger, Noémie Saint Germès and Pierre-Louis Stenger for helping us during field work, and anonymous reviewers for useful comments. The Province Sud (New Caledonia) issued all permits.
Funding
This study was part of a research project financed by the Polish National Science Centre (grant NCN 2011/01/M/NZ8/03344 and 2018/29/B/NZ8/02312), the Polish Ministry of Science and Higher Education (grant 2P04F 001 29), Syndicat Mixte des Grandes Fougères (New Caledonia), Conservation des Espèces et Populations Animales (France), and La Fondation Nature et Découvertes (France). HJ received funds from the Agence Nationale de la Recherche (ERA-Net BiodivERsA Project FFII, JE 288/7-1) and CM from the IAC core budget.
Author information
Authors and Affiliations
Contributions
HJ, RG and JT contributed to the study conception and design. All authors contributed to data collection. HJ and JT performed data analysis and wrote the first draft of the manuscript. All authors revised and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jourdan, H., Bourguet, E., Mille, C. et al. Impact of invasive little fire ants Wasmannia auropunctata on rainforest soil fauna: implications for conservation of the endangered flightless kagu of New Caledonia. Biol Invasions 24, 3675–3680 (2022). https://doi.org/10.1007/s10530-022-02882-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-022-02882-8