
Abstract
The oak (Quercus robur L.) regeneration intensity was assessed in the core area of the Białowieża National Park (BNP) in Poland with respect to the selected ecological factors. The emphasis was placed on the response of oak regeneration to disturbances, including the large-scale dieback of spruce stands. Defining their effect could help predicting the role of oak in naturally developing lowland forest ecosystems in the European hemiboreal zone. The results of the study challenge the opinion that the ‘lime-oak-hornbeam forest’ is a ‘climax’ community, confirming a very poor regeneration represented by only two saplings taller than 0.5 m per hectare. By contrast, in spruce-dominated communities, from 49 to 848 taller saplings per hectare were found. The occurrence of saplings was associated with discontinuous canopy of late seral stage of stands, as well as with large gaps. Most of the best quality grown-up oak saplings developed in the immediate neighbourhood of spruce logs. The results of the research indicate that ‘lime-oak-hornbeam forest’ (Tilio-Carpinetum) should be rather perceived as a transient community, evolved from relict, culturally modified, oak woodlands. Because spruce had become a dominating species only after abandoning in mid-1800s the historical regime of anthropogenic disturbances (involving frequent forest fires), the observed phenomena related to the disintegration of spruce stands had probably no precedent over the last 500 years. To confirm whether the massive decline of spruce stands will finally result in the successful establishment of the new canopy oak generation, both in Białowieża and other forests of hemiboreal zone, further research is needed.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Unfavourable ecological conditions related to competitive abilities of oaks in the contemporary forests are often addressed as a threat to this genus of trees, represented only by Quercus robur and Q. petraea north from Alps and Carpathians. In particular, oak regeneration and recruitment have become one of the most topical issues in the debate of practitioners and forest researchers on the future of European forests. This is because in oak-rich old-growth stands representing ‘lime-oak-hornbeam forest’ (referred in phytosociology as Tilio-Carpinetum), with oak assumed to be a “climax” species (Falinski 1986; Ellenberg 1988; Sokołowski 1993), its regeneration is practically absent (Bernadzki et al. 1998; Zajączkowski 1999; Vera 2000; Götmark 2007; Brzeziecki 2008). One commonly accepted factor blamed for the shortage of oak in the new generation of trees is the strong competition and suppression by the shade-tolerant abundant undergrowth, particularly beech, lime and hornbeam (Vera 2000; Paluch 2005). Another factor considered responsible for the oak regeneration failure is grazing/browsing pressure referred to as ‘game damages’ (Paczoski 1930; Falinski 1986; Miścicki 1996; Kweczlich and Miścicki 2004; Palmer et al. 2004; Götmark et al. 2005; Paluch 2005).
The discussion over the role of ungulate herbivores in forest dynamics, particularly their effect on the oak regeneration, was revigorated by Frans Vera, propagating the theory of the savannah-like primeval landscape shaped by huge herds of large herbivores (Vera 2000). According to the author, unlike closed forest habitats favouring the regeneration of shade-tolerant species, the combination of groves, single trees and open areas maintained due to the very high grazing pressure must have secured favourable conditions for the successful regeneration of intolerant species, including oaks (Vera 2000). Although a long time ago there were forest ecologists who recognized oaks as skilled colonizers of abandoned fields (Watt 1919), it is only since recently when one has commonly perceived Quercus robur and Q. petraea as post-pioneer competitors (sensu Grime 1979), rather than constituents of close forests climax (Feijen 2003). It has been recognized that the oak regeneration benefits from large-scale disturbances (Packham et al. 1992; Danielewicz and Pawlaczyk 2006; Bobiec 2007), including the disturbances caused by man (Bradshaw et al. 2003; Kramer et al. 2003). Low-energy forest fires, retrograde extensive farming, in particular pastoralism leading to the soil degradation and acidification, have neutralized the competition of the climax shade-tolerant species, such as lime and hornbeam, and have given a strong competitive advantage to oaks being more tolerant of poorer soils (Peterken 1993; Mason 2000; Feijen 2003).
The new emerging paradigm of the evolution of forests in Europe is being subject to thorough palynological, archaeological and historical investigation. Will it downgrade the forests nowadays revered as windows to the past to mere artefacts of strict preservation as suggested Vera (2000)? The Białowieża Primeval Forest poses serious challenges to all parties attached either to the utilitarian perspective, claiming that the original character of the ecosystem can be restored through silvicultural intervention, or to the idealistic perception of an untouched relict of the primeval forest or to the ‘large herbivore theory’.
However, the choice between oak regeneration in ‘closed forests’, assumed by many as doomed, or regeneration in ‘savannah’ woodland pastures, ecotone zones, is not necessarily the only alternative. Although in contemporary European forests natural catastrophic disturbances occur much less frequently than those caused by man, they may be sufficient to secure successful replacement of generations regarding the longevity of oak. It has been suggested that under natural conditions, oak has developed a strategy of discrete regeneration through the waves triggered by large-scale disturbances (Kowalski 1993; Bobiec 2007). The problem is that throughout history, mankind has severely modified the disturbance regime of forests, replacing infrequent natural catastrophic disturbances with anthropogenic disturbances favouring oak regeneration (Feijen 2003).
Spontaneous regeneration of stands in the core zone of the Białowieża National Park (BNP), secured by the strict preservation, provides the opportunity to study the influence of natural large-scale disturbances on regeneration of trees, including oak. The progressing expansion of its major competitors, hornbeam and lime, at the expense of less shade-tolerant species has been observed since 1936 on ca. 15 hectares of permanent plots in the BNP core area (Bernadzki et al. 1998). Recently, a more detailed analysis of the stand development on one of these plots (1.2 ha) has been performed by Brzeziecki (2008). The author followed the stand development between 1936 and 2001 in the deciduous and mixed oak-pine forest, revealing conspicuous shifts in both stand densities and species composition. In the latter, the 65-year-long period resulted in the maturing of numerous spruce trees, already recorded in 1936, outcompeting younger oaks, and in the continuous decline of the then dominating old pines and almost total disappearance of birches being replaced by the dense hornbeam undergrowth. In the deciduous forest, the initially very sparse, oak-dominated stand (total density 129 trees ha−1) turned to the dense pole phase stand (density 966 trees ha−1) dominated by hornbeam (none in 1936, 475 ha−1 in 2001) and lime (4 in ha−1 1936, 371 ha−1 in 2001) (Brzeziecki 2008). The advanced regeneration of spruce in 1936 in the pine-oak forest compared to the delayed regeneration of broadleaved species in the deciduous forest community can be explained by the selective and very intense grazing pressure on the deciduous trees regeneration at the end of 1800s and the beginning of 1900s (Paczoski 1930; Falinski 1986; Jędrzejewska et al. 1997). It seems that the cited figures are a very good illustration of the ecosystem reaction to the release from the previous forms of anthropogenic pressures (Brincken 1826; Paczoski 1930; Hedemann 1939; Samojlik 2006). Perhaps the most obvious symptom of the natural disturbance regime nowadays in the Białowieża Forest is the massive decline of spruce (Picea abies) stands. In particular, the role of the spruce bark beetle, Ips typographus, should be taken into consideration.
The objective of this study was to investigate oak regeneration in a closed lowland forest, having been protected as a strict preserve for the last ninety years. The emphasis was placed on the response of oak regeneration to disturbances including the large-scale dieback of spruce stands. This can help finding an answer to the question whether oak has a strategy enabling the species to regenerate in closed forests and, in consequence, predicting the role of oak in naturally developing lowland forest ecosystems in the European hemiboreal zone. The most opportune research area for such project was the core zone (strict reserve) of the Białowieża National Park, Poland.
Materials and methods
Study area
The core area of today’s Białowieża National Park (N 52.74°, E 23.87°, 4,747 hectares = ca. 8% of the Polish part of the Białowieża Primeval Forest) was set aside as a nature reserve in 1916 from the intensive use of forest initiated in 1915, having been preserved ever since. Until World War I, the entire forest had a special protective status of a royal forest (from the 15th to the end of the eighteenth century) and served as the imperial hunting ground of tsars of Russia (during the nineteenth century until 1915). Abiotic conditions are primarily determined by the continental climate (average annual precipitation of 641 mm, ranging from 426 to 940 mm, 85% in rain; average annual temperature of 6.8°C, ranging from 5.1 to 8.8°C, with the average temperature in July ranging from 15.2 to 21.6°C and in January from −13.4 to 1.8°C; Olszewski 1986), the postglacial geological formation (vast plains with small altitude differences; altitude ranging from 134 to 202 m a.s.l.) and its location within the continental watershed that make the system dependant on precipitation (Falinski 1986).
Field sampling and data
Sampling lines of a total length of approximately 9,800 m were laid out through the stands of the BNP core area (strict reserve). Their location was arbitrarily aimed to avoid swamps and bogs. The reason for this exclusion was to avoid the damage that excessive trampling would induce to those vulnerable and locally rare habitats. Two L-shaped lines in the NE part of the BNP were set in order to improve the representation of meso-oligotrophic sites, less common in the BNP than eutrophic sites of deciduous forests (Fig. 1).

The study area (core zone of the Białowieża National Park) with sampling lines and four 50 × 50-m plots (a, b, c, d); magnified fragment of sampling line and 100 × 50-m sampling units; Dc deciduous, MDc mixed deciduous, MCn mixed coniferous, BF bog and swamp and RF riparian forest site type; H0 seedlings and saplings shorter than 20 cm, H1p saplings ≥20 cm; Ps Pinus sylvestris, Pa Picea abies; a digital map of the stand layers: the Białowieża National Park courtesy
The lines provided the basis for the collection of data on oak regeneration and selected characteristics of the local environment, including:
-
Height classes of regeneration: seedlings under the height of 0.2 m (H0), 0.2–0.5 m (H1), saplings of 0.5–1.3 m (H2), 1.3–2.5 m (H3), 2.5–5.0 m (H4) and the undergrowth taller than 5.0 m, but thinner than 0.1 m DBH (H5).
-
Sapling relative quality representing the future growth potential of H1-5 oaks, based on the qualitative evaluation of the health status regarding the development of foliage and stem/shoot damages as well as the relative level of suppression by the neighbouring vegetation: from 1 healthy and vigorous (Q1), through slightly damaged and/or suppressed (Q2), to seriously damaged and/or suppressed (Q3). Preliminary observations showed that using one joint category, expressing the influence of the effect of health status and competitive stress is fully justified. For instance, on the border zone between the ancient forest and abandoned fields/meadows, many saplings were classified as ‘bonsai’—although heavily browsed, in full sunlight, they vigorously respond by building up their well dense, strongly ramified crowns (A. Bobiec, K. Solecka, unpublished data). As most of their counterparts in small gaps or under the canopy hardly survive even minor injuries, competitive suppression and game damage act as one negative complex factor.
Young oaks were recorded and positioned with GPS up to 1 m on each side of the lines;
-
Assessment of the stand developmental phase within the radius of 10 m around the recorded oaks: young, pole, mature (optimal), old-growth (biostatic) and decay (degradation) phases as well as gaps were distinguished [nomenclature and definitions after Oldeman (1990); Bobiec et al. (2000); Bobiec (2007)];
-
Representing stand influence on the growing conditions of young oaks, four largest trees within the radius of 10 m around oaks were recorded and assigned to three DBH classes: 0.05–0.25 m, 0.26–0.50 m and more than 0.50 m;
-
The presence of lying logs larger than 0.10 m in diameter within the distance up to 1 m from the oak seedlings/saplings was recorded as well as their diameter at the oak nearest point;
-
The coverage degree of up to five major herbaceous species within the radius of 1 m around the young oaks was assessed in five-point scale: + (less than 1% of the area), 1 (1–5%), 2 (5–25%), 3 (25–50%), 4 (50–75%) and 5 (75–100%).
The same parameters were recorded along logs longer than 10 m crossing the sampling lines, used as additional, random sampling lines (within the 1-m-wide zone on each side of the logs); obviously, all the regeneration recorded this way represented seedlings/saplings with logs in their immediate neighbourhood; end points of logs were georeferenced (Fig. 1).
In order to estimate the volume of dead wood that might affect the regeneration, diameters of all lying coarse woody debris (CWD) thicker than 0.1 m at the line-crossing points were measured (“line intercept” method by Van Wagner 1968). The wood species (unless beyond recognition) and degree of decay in five-point scale (from 1- fresh to 5- entirely rotten) were determined and assessed (Bobiec 2002).
In order to better capture oak regeneration in the aftermath of large-scale natural disturbances, four 50 × 50-m plots were set in large canopy gaps: the result of massif spruce stand dieback. Two gaps originated between 2000 and 2004 (plots A and B) and the other two between 1994 and 1997 (plots C and D; A. Keczyński, pers. communication, Fig. 1). In each plot, the same parameters as listed earlier were measured and recorded along the 300-m-long grid of six 50-m-long lines as well as along randomly selected logs yielding a comparable total length. All data were collected in June–August, 2009.
Data management with GIS
All the georeferenced data (oaks, lines and logs) were transferred to shape files and referred to site and stand data extracted from the BNP numeric maps based on the 2001 BNP’s inventory. Although the real positions of recorded oaks or any other point objects (such as log ends) strictly correspond with certain environmental characteristics, their GPS-recorded coordinates were burdened with the inaccuracies of five to a dozen metres, depending on the acquired satellite signal. Even more serious sources of the spatial model deviation from reality are mapping errors underlying identification and demarcation of forest site and stand units. Therefore, in order to enable the correlation of regeneration variables with environmental factors regardless of the existing inaccuracies, 193 rectangular sampling units were created by the division of the 100-m-wide belt (50-m buffer each side of the sampling line) in 50-m-long sections.
The forest site types crossed by the sampling lines were generalized to the deciduous forest type Dc (combined mesic and wet deciduous forest on eutrophic soils with mull form of humus: mainly haplic cambisols, cambic luvisols and haplic epigleysols), the mixed-deciduous forest type MDc (combined mesic and wet mixed-deciduous forest on mesotrophic soils with moder form of humus: mainly cambic ochriarenosols, cryptic podzolsols and albic luvisols) and the mixed-coniferous forest type MCn (combined mesic and wet mixed-coniferous forest on meso-oligotrophic soils with mor form of humus: mainly albic ochriarenosols and haplic podzols) (Prusinkiewicz and Michalczuk 1998; Michalczuk 2001). Similarly, from the numeric layer of BNP stands, the subset layers of stands dominated by oak and spruce were generated. For each sampling unit, the length of sampling lines and ‘sampling logs’, as well as the area occupied by the particular site and stand categories were calculated.
While three 50 × 50 m plots (B, C and D) represented only MCn, plot A also partly covered MDc sites.
The GIS data management, length and area calculations were performed with ArcGIS ArcView 9.3, ESRI.
Calculations and statistical analyses
In order to detect major environmental factors influencing oak regeneration, a direct ordination technique, canonical correspondence analysis (CCA), was applied with CANOCO (Ter Braak 2003). The direct relationships between particular variables and the density of oak regeneration within 193 sampling units were researched with use of the Spearman rank correlation (Statistica v. 8, StatSoft). The G-test of goodness of fit was applied in order to find whether the number of young oaks reflected the proportions of the forest categories in the sampling units. Similarly, the distributions of particular oak regeneration classes in the forest categories were compared with the G-test of independence (McDonald 2009).
Results
The total length of regular sampling lines and sampling logs amounted to 16,655 m, referring to the area of 33,310 ha, where the oak regeneration was inventored. This does not include the lines and logs in four additional sampling areas A, B, C and D, totalling 2,538 m (0.5076 ha). While in the whole strict reserve, the area share of Dc, MDc and MCn sites is as 72:18:10, on the sampling scheme, the share of mixed sites in the sampling line/logs length was strengthened at the expense of the deciduous forest site, reflected by the proportion of 58:27:15 of Dc, MDc and MCn, respectively (Table 1). As the branching mode of fallen trees can substantially influence the conditions of seedling establishment and sapling recruitment (safe spots) in the immediate neighbourhood of logs, the sampling logs were divided into two categories: hardwood (all broadleaved species) and softwood (spruce 97% and pine 3%) (Table 1).
General data
Overall regeneration density
The average number of oak seedlings/saplings per one hectare varied from 56 in deciduous forests through 333 in mixed-deciduous to 360 in mixed-coniferous forest types. However, the numbers were much higher in the four plots representing gaps in the mixed-coniferous forest, ranging between 425 and 1,441 young oaks ha−1. Most of the saplings taller than 0.2 m belonged to Q1 category (healthy and vigorous), and only a minor part was classified as Q3 (seriously damaged/suppressed) (Tables 2 and 3).
Correlation between oak regeneration with environmental variables extracted from the numerical map
The density of oaks (N oaks 100 m−2, calculated on the basis of the ratio of the oak number and the total surface of 2-m-wide belts along lines and logs in a sampling unit) was correlated with a set of environmental variables in 100 × 50-m sampling units: the area occupied by oak and spruce stands, the relative share of the forest site types as well as the length of softwood and hardwood logs. As the frequency of sampling units against the average density of oaks [N · 100 m−2] revealed a geometric distribution (with 43% of the sampling units with no recorded oaks), the nonparametric Spearman rank correlations were calculated.
Interestingly, the share of oak stands, unlike positively correlated spruce stands, showed no significant correlation with oak densities. Similarly, the share of the mixed-coniferous forest site in sampling units was significantly positively correlated with the densities of oak regeneration representing all height and quality classes. Except for the youngest seedlings H0, the correlations of the mixed-deciduous site were weaker and less significant than in the former case, and the young oaks were clearly avoiding deciduous forest sites, the fact reflected by relatively strong, significant negative correlations. Analogically, the length of softwood (predominantly spruce) logs was significantly and positively correlated with the oak regeneration density, which was not observed in the case of hardwood logs (Table 4).
Direct detection of the environmental influence on oak regeneration
In order to detect major factors influencing oak regeneration in the bulk of data, the direct gradient analysis CCA was applied. The oak regeneration represented by dummy variables H0, H1, H2, H3 and H4, as well as Q1, Q2 and Q3 together with the ground vegetation species occurring in the neighbourhood of more than five young oaks (in total 27 species) were species variables. They were referred to nine environmental variables:
-
Presence of young developmental phase of stand, PY;
-
Presence of mature and well-stocked old-growth (biostatic) phase, MOG;
-
Presence of degradation phase and gaps, DG;
-
Close neighbourhood of softwood and hardwood logs, CnLogs and DcLogs, respectively, as well as the lack of logs, NoLog;
-
Diameter of the oak-neighbouring logs, Diam1;
-
Presence of two oak-neighbouring logs, 2Logs;
-
Degree of the tree canopy cover, Cpy.
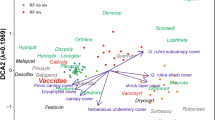
Altogether, 592 cases were subject to the analysis. No transformation of species data was performed, no weights were specified and no downweighing of rare species was made. The total inertia of axes (sum of all eigenvalues) was 5.138, and the eigenvalues of the two first displayed axes were 0.351 (species-environment correlation 0.752) and 0.036 (species-environment correlation 0.301). The sum of all canonical eigenvalues was 0.458. The unrestricted permutation Monte Carlo tests (499 permutations under reduced model) were performed: one of significance of the first canonical axis (eigenvalue = 0.351, F-ratio = 42.812, P = 0.002) and the other of significance of all canonical axes (Trace = 0.458, F-ratio = 7.128, P = 0.002).
The ordination results displayed on the biplot show a strong affinity of the higher oak categories (H1, H2, H3 and taller, symbolized H3p) with mainly coniferous forest ground vegetation as well as with the light-demanding species of gaps and clearings such as Molinia caerulea, Rubus idaeus, Pteridium aquilinum and Calamagrostis arundinacea. All these species have negative values on the first canonical axis but they are positively correlated with the presence of gaps and decay phase of stands, presence of softwood logs, double logs and to some extend with the log diameter. The youngest and shortest seedlings H0 together with mostly deciduous forest herb species have been arranged on the positive side of the first canonical axis, and they are positively correlated with the remaining environmental variables. The most distant species of the highest positive values on the second canonical axis (with Majanthemum bifolium and Stellaria holostea) are these which were the least frequent in the neighbourhood of young oaks (Fig. 2).

The biplot of the canonical correspondence analysis; environmental variables: DG decay stand developmental phase and gaps, MOG mature and well-stocked old-growth phase, PY pole and young phase, Cn/DcLogs presence of a softwood/hardwood log in the 1-m distance from a young oak, NoLog absence of a log in the 1-m distance from a young oak, 2Logs presence of two or more logs in the 1-m distance from a young oak, Diam1 diameter of an oak—neighbouring log, Cpy degree of the canopy cover; species: At_fi Athyrium filix-femina, Ca_pi Carex pilosa, La_ga Lamiastrum galeobdolon, Ma_bi Majanthemum bifolium, St_ne Stellaria nemorum, St_ho S. holostea, Eq_sy Equisetum sylvaticum, Ox_ac Oxalis acetosella, Ge_ro Geranium robertianum, Gy_dr Gymnopteris dryopteris, Ca_re Carex remota, Ur_di Urtica dioica, Ca_ar Calamagrostis arundinacaea, Ru_id Rubus idaeus, Va_my Vaccinium myrtillus, Pt_aq Pteridium aquilinium, Mo_co Molinia coerulea, Dr_ca Dryopteris cartusiana, Moss moss layer in general, BG vegetation-free soil, H0 oak seedlings shorter than 0.2 m, H1 oak seedlings 0.2–0.5-m tall, H2 oak saplings 0.5–1.3 m, H3p oak saplings >1.3 m, Q1 oak seedlings/saplings of the best quality, Q2 oak seedlings/saplings of intermediate quality, Q3 oak seedlings/saplings of the lowest quality
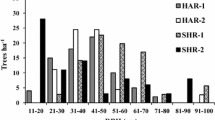
The comparison of stands composition in the nearest neighbourhood of young oaks (within 10-m radius) showed significant differences between oak seedling/sapling categories: H0, H1 and H2p (saplings taller than 0.5 m). Taller and older saplings were neighboured by smaller numbers of trees (DBH > 5 cm), ranging from 2.2 trees around seedlings and saplings H0, to 1.6 around saplings H1, to 1.2 around saplings taller than 0.5 m. This corresponds with the decreasing canopy cover and the importance of the stand decay phase and gaps for the oak recruitment. In particular, in the neighbourhood of taller saplings, there were considerably fewer young hornbeams (DBH from 0.05 to 0.25 m) and relatively more large and intermediate size spruces (Figs. 3, 4).

Stands composition in the immediate neighbourhood of oak seedlings/saplings. N average number of largest trees (N max = 4, DBH ≥ 5 cm) within 10-m radius around oak seedling/sapling; the G-test of goodness of fit showed significant difference between the % proportions of the largest trees neighbouring oaks H0 and H1 (5 d.f., G = 13.1905, P = 0.0217), H1 and H2p (5 d.f., G = 26.9669, P = 0.0001) as well as H0 and H2p (5 d.f., G = 49.9714, P = 0.0000); Qr Quercus robur, Cb Carpinus betulus, Tc Tilia cordata (few specimens of Ap Acer platanoides were added) and Pa Picea abies; the indices 1, 2 and 3 following the species symbols refer to the DBH classes: 1 (5–25 cm), 2 (26–50 cm), 3 (>50 cm). H2p—saplings taller than 0.5 m, for H0 and H1, see Fig. 2

Stand developmental phases as an environmental factor of the oak regeneration; the G-test of independence showed significant differences in the proportions of developmental phases between the locations of H0/H1 (d.f. 2, G = 37.3337, P = 0.0000), H1/H2p (d.f. 2, G = 12.3408, P = 0.0021) and H0/H2p (d.f. 2, G = 73.8801, P = 0.0000); PY young and pole phase, MOG mature and well-stocked old-growth phase, DG decay phase and gaps (following Bobiec et al. 2000); see Fig. 3 for H0, H1 and H2p
Coarse woody debris influence on oak regeneration
The average volume of CWD that was estimated along 9,793 m of the regular sampling lines was 88 m3 ha−1, ranging from 60 m3 ha−1 in the deciduous forest to 93 m3 ha−1 (significant difference, P = 0.0095) in the mixed-coniferous forest, to 112 m3 ha−1 (insignificant difference between the two latter figures, P = 0.2086, exact binominal test of goodness of fit) in the mixed-deciduous forest type. In four additional square plots set in the MCn forest gaps, the volume of dead wood was even higher, from 142 (A), to 186 (B), to 254 (D), to 285 m3 ha−1 (C). In all the three types of forest sites, most of the dead wood volume recruited from spruce decline; however, in mixed forest sites, the contribution of this species was overwhelming. As it could have been expected, the more productive the site the bigger the share of the larger diameter woody debris was (Table 5).
The comparison of the numbers of oak seedlings/saplings along the regular sampling lines with those found in the close vicinity of sampling logs showed that the latter either directly favoured or indicated other factors favouring the regeneration (Fig. 5). The difference was even sharper when the comparison was made in the four gap plots. The average density in plots A–D calculated on the basis of oaks found along 1,200 m of regular sampling lines was 407 ha−1 (the specimens growing within the distance of 1 m from a log of diameter > 0.1 m were removed from the calculation), while the average density calculated on the basis of the regeneration along the sampling logs was two-and-a half fold higher: 1,043 ha−1 (Fig. 6).

The percentage of young oaks along regular sampling lines (Line), hardwood/softwood sampling logs (Dc/Cn) versus the length proportion; the differences between the observed proportions of oak numbers along the three categories of sampling lines and those expected from the line length proportions were significant: H0 (d.f. 2, G = 48.2884, P = 0.0000), H1 (d.f. 2, G = 80.1830, P = 0.0000) and H2p (d.f. 2, G = 66.4633, P = 0.0000, G-test of goodness of fit); see Fig. 3. for H0, H1 and H2p

Average densities of oak seedlings/samplings in the four gap plots (A, B, C and D) calculated on the basis of oaks found near the regular sampling lines (1,200 m) and sampling (spruce) logs (1,338 m); H3- saplings 1.3–2.5 m, H4—2.5–5.0 m; see Fig. 2 for H0, H1 and H2
Assuming that spruce logs had a similar effect on the oak regeneration in Dc, MDc and MCn forest types, one could expect that the ratio of seedlings/saplings numbers along spruce logs in those forest types would reflect the ratio of the total length of those logs (respectively, 1,827:1,936:1,305 m). Although 36% of the total length of spruce “sampling logs” were located in the deciduous forest type, only 11% of all seedlings/saplings were found along spruce logs in this forest type. The deficit of the oak regeneration with respect to the expected number was even larger if only saplings taller than 0.5 m were considered (Fig. 7).

Discussion
Oak regeneration in different forest types
The results of the present study depict the current oak regeneration status in three site categories. The richest of them, where the longest section of the sampling line was located, the deciduous forest, is the site of mixed lime-oak-hornbeam (EEA 2009), a synonym of an oak-hornbeam forest (Ellenberg 1988) or Tilio-Carpinetum (Falinski 1986). Although single grown-up saplings are found here and there in the gaps representing the ‘catastrophic mode’ of disturbance (i.e. a type of disturbance giving the advantage to intolerant species over the renewal of climax species, after Veblen 1992), this is not the case in most of the Tilio-Carpinetum area with gaps being filled by the prolific shade-tolerant hornbeam and lime recruited from the pre-disturbance seedling, sapling and vegetative outshoot banks (Bobiec 2007).
The large-scale inventory of tree regeneration carried out in the BNP core area in 2005 in 460 circular plots gave comparable figures to the average densities from the present study. According to that investigation, there were on average 164 oaks ha−1, including 111 representing the lowest class of seedlings/saplings shorter than 0.3 m and 50 representing the 0.3–1.3-m class of oak undergrowth (S. Miścicki, unpublished data; Krasuska and Miścicki 2002). Weighing the regeneration densities resulting from the present study with the relative share of particular categories of forest types resulted in comparable figures: 36 small (0.2–0.5 m) saplings and 25 taller (>0.5 m) saplings ha−1. The regeneration intensity in the particular types of forest as well as the differences between them detected by the current study was similar to those reported by Paluch (2005) from his three-year observation series covering similar habitats of the Białowieża Forest (Dc- 110, MDc- 242, MCn- 585 ha−1).
If the rate of oak regeneration in the deciduous forest type continues at such a low level as revealed by the present study and earlier investigations (Miścicki 1996; Krasuska and Miścicki 2002; Paluch 2005; Bobiec 2007; Brzeziecki 2008; S. Miścicki personal comm.), it will not be able to sustain the share of oak that is believed typical for the lime-oak-hornbeam forest community. However, anticipating major compositional changes of stands, one cannot conclude that oak will disappear. Although the study could not directly detect factors preventing successful regeneration of oak at the scale of the entire biochore of the mesic deciduous forest, several ecological characteristics covered by the project point at patterns and dynamics responsible for oak recruitment, including light regime and relative competitive power.
CWD and spruce stands dieback as a factor of oak regeneration
As it was shown, MCn and MDc sites had almost fivefold more saplings per length unit of spruce logs than Dc sites. This indicates that oak regeneration density is not a linear function of the CWD volume. Instead, one can hypothesize that only if a certain necessary CWD load is locally attained, are there opportune conditions for prolific oak regeneration. Our study revealed almost twofold higher CWD load (per hectare) in MCn than in Dc sites on average and three to fivefold higher in four square plots representing MCn declined stands. This would explain strong correlations of the oak regeneration intensity with both the share of the MCn forest type (to lesser extend also MDc), the area occupied by spruce stands, and the length of spruce logs. Unless unpredictable catastrophic factors occur (e.g. fire and tornado), the potential for the oak regeneration depending on intensive disturbances will probably be limited to spruce-dominated stands. Unlike other communities, they are subject to sudden disintegration producing large gaps and open degradation phase with a very high load CWD. While in Dc sites, the spruce abundance is limited, this is the major canopy species in both MDc and MCn forest sites of the BNP core area. According to the last inventory of the BNP core zone in Dc forest type, 29% of the biochore was occupied by oak-dominated stands while only 10% by spruce-dominated stands, in MDc forest type 22 and 44%, respectively, and in MCn 4 and 41% were occupied by oak- and spruce-dominated stands, respectively.
An even bigger deficit of oak regeneration in the deciduous forest compared to poorer forest sites occurred when only taller saplings were considered. The CCA ordination has clearly separated them together with coniferous ground vegetation and forest clearing species (such as Rubus idaeus) from the youngest unlignified seedlings more related to the deciduous forest ground vegetation and more shade-tolerant species.
Therefore, if the presence of spruce-dominated stands was applied as a rough, preliminary predictor of oak regeneration, such potential would exist in MDc and MCn forests but not in Tilio-Carpinetum. The concentration of cropping oaks in deciduous and mixed-deciduous forests should not be perceived as a factor preventing intensive regeneration in mixed-coniferous and coniferous forests. Even if there are no mature oaks in certain parts of the BNP, they remain within the range of seed rain due to the efficient zoochoric dispersal secured by jay Garrulus glandarius (Bossema 1979; Mosandl and Kleinert 1998).
The strong association of the developing young oaks with the immediate neighbourhood of spruce logs could be explained by both the ‘barwire’ effect of logs modifying the browsing pressure (as suggested by Bobiec 2002, 2005; Keczyński 2002) and opportune habitats for the location of jay’s acorn caches (M. van de Loo, pers. communication).
Various factors responsible for spruce decline have been discussed in numerous papers pointing at unfavourable changes in the ground water regime (Alavi 2004), air pollution (Schulze 1989; Esch and Mengel 1998), climate change causing spring draughts and summer heats (Rehfuess 2007; Levanič et al. 2009), advanced age and weakening of natural mechanisms of the resistance to fungal infections and insect infestations. In the view of our findings, the bark beetle Ips typographus outbreak seems a very beneficial factor, shaping the final effect of spruce stands decline, providing necessary for successful oak regeneration the “catastrophic” characteristics of the disturbance.
The results of the present study are congruent with the former inventory of dead wood in terms of both the total volume (one-fifth to one-fourth of the above ground dendromass) and species composition (Bobiec 2002). The conspicuous disproportion between the present low share of spruce in the deciduous forests canopy and its dominating position in their dead wood resources (32% of the Dc downed CWD volume) indicates that further spruce withdrawal from Tilio-Carpinetum will result in a lower CWD “equilibrium” volume. Therefore, if the observed trend continues, oak will regenerate as a result of occasional and perhaps local waves rather than continuously throughout larger area (Bobiec 2007). Such strategy of regeneration based on considerable longevity and opportunistic use of infrequent disturbances triggering secondary succession has been ascribed to oak (Kowalski 1993; Danielewicz and Pawlaczyk 2006; Bobiec 2007).
Historical and management context of the observed processes
Could the described processes occur in all other lowland temperate forests or are they limited to the Białowieża Forest as a result of the combined effect of the unique local factors?
There is a strong historical evidence that the processes observed in this forest are the consequence of its historical management. According to the account of Julius von Brincken, the Białowieża Forest superintendent in early 1800s, as much as 80% of the forest area was dominated by pine stands and the deciduous forest habitat consisted of isolated enclaves dispersed throughout the vast, monotonous matrix of pine forests (Brincken 1826). Interestingly, the author emphasized that spruce belonged to the deciduous forest communities, not to the coniferous ones, absolutely dominated by pine. Another noticeable fact was the substantial share of pioneering species in deciduous stands (Brincken 1826) which would suggest that such stands were partly undergoing the recovery succession after serious natural disturbances or a certain form of anthropogenic pressure. It is a well-known fact that under the rules of the Polish-Lithuanian Commonwealth (Rzeczpospolita), i.e. from the turn of the 15/16th centuries untill the end of the eighteenth century, the local dignitaries as well as villagers recruited as the royal forest wardens were entitled to various forms of the forest use, including grazing, hay making, beekeeping and producing of tar and potash (Brincken 1826; Paczoski 1930; Hedemann 1939).
One of the most important factors influencing the dynamics, species composition and structure of the Białowieża forests in the past was frequent but low energy fires, mostly caused by man, either beekeepers, charcoal-tar-potash manufacturers, or herd wardens (Harnak 1764 after Hedemann 1939; Samojlik and Jędrzejewska 2004; Niklasson et al. 2010). According to Brincken (1826), fires in the Białowieża Forest occurred almost on the annual basis. This was the factor that confined fire-intolerant spruce either to the least flammable sites of riparian and swamp forest or to enclaves of rich deciduous stands.
When the radical ban on the use of fire was imposed and efficient fire suppression techniques developed in 1800s (Samojlik and Jędrzejewska 2004; Niklasson et al. 2010), it triggered the regeneration of shade-tolerant species in semi-open stands, the secondary succession of forest on abandoned clearings and progressing spruce encroachment on the area freed from the overriding fire regime. The spruce encroachment was even intensified after 1860s where big herds of cattle were allowed to graze in the forest and the game populations sharply increased due to the tzars’ hunting policy (Jędrzejewska et al. 1997).
As most of those spruce stands have begun to disintegrate only recently (the studied gaps emerged in mid-1990s), it is too early to predict the final result of the observed oak regeneration. Furthermore, because the resulting recovery of the potential vegetation occurs for the first time after four hundred years of various forms of human activity, substantially modifying forest characteristics (Samojlik and Jędrzejewska 2004), we do not have any historical analogy of what is observed with regard to oak regeneration. The processes captured in our study can be interpreted as the second stage of the continuous natural recovery lasting since the beginning of 1800s, triggered by shutting down the hundreds-years-long history of the ancient forms of forest use. This stage is characterized by the maturing of hornbeam-lime stands and progressing since the late 1900s massive decay of older spruce stands.
The recovery of natural dynamics is, however, mostly confined to the area of the Białowieża National Park, merely 17% of the Białowieża Forest Natura 2000 site (both Special Aarea of Conservation and Special Protected Area). This process is being seriously hampered outside the park both by silvicultural measures aimed to establish oak stands through replanting small clear-cuts (up to 2,000 m2) and “sanitary” removal of bark beetle-infested spruces (RDLP 2002).
Conclusions
-
Lime-oak-hornbeam forest also known as Tilio-Carpinetum with a substantial participation of oak in the canopy layer is a transient community evolved from relict culturally modified oak woodlands; such communities developed and formed under the historical regime of anthropogenic disturbances which provided favourable conditions for oak regeneration; therefore, the lime-oak-hornbeam forests should not be perceived as climax, self-replacing communities.
-
The importance of declining spruce stands for oak recruitment has been confirmed by the present study: the density of the oak regeneration in spruce-rich mixed-forest communities was six to sevenfold higher than in Tilio-Carpinetum with a weak spruce component; in large gaps formed as the result of bark beetle outbreak, this ratio was even higher: from eightfold to 25-fold.
-
Also on a microscale, the close neighbourhood of spruce logs favours establishment and development of new oak generation.
-
As the large-scale dieback of spruce stands in meso-oligotrophic sites began only at the end of the twentieth century, the observed oak regeneration in such areas has probably no precedent over the last 500 years.
-
Artificial oak regeneration in the managed part of the Białowieża Forest does not mimic the historical disturbance regime that has led to the establishment of the present old-growth lime-oak-hornbeam forest.
-
Due to the dependence of oak regeneration on the dieback of spruce stands and to the lack of beech in the Białowieża Forest, the reference applicability of the observed phenomena should probably be confined to the hemiboreal zone.
References
Alavi G (2004) The impact of soil moisture on stem growth of spruce forest during a 22-year period. Forest Ecol Manage 166:17–33
Bernadzki E, Bolibok L, Brzeziecki B, Zajączkowski J, Żybura H (1998) Compositional dynamics of natural forests in the Białowieża National Park, northeastern Poland. J Veg Sci 9:229–238
Bobiec A (2002) Living stands and dead wood in the Bialowieza Forest: suggestions for restoration management. Forest Ecol Manage 165:121–136
Bobiec A (2007) The influence of gaps on tree regeneration: a case study of the mixed lime-hornbeam (Tilio-Carpinetum Tracz. 1962) communities in the Białowieża Primeval Forest. Pol J Ecol 55:441–455
Bobiec A, Burgt H, van der Meijer K, Zuyderduyn C, Haga J, Vlaanderen B (2000) Rich deciduous forests in Bialowieza as a dynamic mosaic of developmental phases: premises for nature conservation and restoration management. Forest Ecol Manage 130:159–175
Bossema I (1979) Jays and oaks: an eco-ethological study of a symbiosis. Behaviour 70:1–117
Bradshaw RHW, Hannon GE, Lister AM (2003) A long-term perspective on ungulate-vegetation interactions. Forest Ecol Manage 181:267–280
Brincken von J (1826) Mémoire Descriptif sur la forêt impériale de Białowieża, en Lithuanie. Annoté et commenté par Daszkiewicz P, Jędrzejewska B, Samojlik T. Glücksberg, Warszawa (reprint 2004 by Editions Epigraf)
Brzeziecki B (2008) Long-term dynamics of natural stands on the example of two forest communities of the Białowieża National Park: Pino-Quercetum and Tilio-Carpinetum. Studia Naturae 54(2):9–22 (in Polish with English summary)
Danielewicz W, Pawlaczyk P (2006) Rola dębów w strukturze i funkcjonowaniu fitocenoz. In: Boratyński A, Bugała W (eds) Dęby. Bogucki Wydawnictwo Naukowe, Poznań, pp 474–563
EEA (2009) Mixed lime-oak-hornbeam forests, EUNIS biodiversity database—find species, habitats and sites across Europe. European Environment Agency. http://eunis.eea.europa.eu/habitats-factsheet.jsp?tab=4&idHabitat=4193. Accessed 20 November 2009
Ellenberg H (1988) Vegetation ecology of Central Europe. Cambridge University Press, Cambridge
Esch A, Mengel K (1998) Combined effects of acid mist and frost on the water status of young spruce trees (Picea abies). Chemosphere 36:645–650
Falinski JB (1986) Vegetation dynamics in temperate lowland primeval forests, ecological studies in Białowieża forest. Dr W Junk Publishers, Dordrecht
Feijen JM (2003) Oak, King or Villein? Is the common oak (Quercus robur and Q. petraea) an anachronism in the Dutch forests? Dissertation, Thesis nr AV 2003-17 FEM 80328, Wageningen Agriculutural University
Götmark F (2007) Careful partial harvesting in conservation stands and retention of large oaks favour oak regeneration. Biol Conserv 140:349–358
Götmark F, Berglund Å, Wiklander K (2005) Browsing damage on broadleaved trees in semi-natural temperate forest in Sweden with a focus on oak regeneration. Scand J Forest Res 20:223–234
Grime JP (1979) Plant strategies and vegetation processes. Wiley, New York
Harnak G (1764) Summaryusz z podatków łowieckich. SA 11575 Lietuvos valstybės istorijos archyvas, Vilnius (in Polish)
Hedemann O (1939) Dzieje Puszczy Białowieskiej w Polsce przedrozbiorowej (w okresie do 1798 roku) [L’histoire de la Forêt de Białowieża (jusqu’a 1798)]. Instytut Badawczy Lasów Państwowych, Warszawa (in Polish with French Resumé)
Jędrzejewska B, Jędrzejewski W, Bunevich AN, Miłkowski L, Krasiński ZA (1997) Factors shaping population densities and increase rates of ungulates in Białowieża Primeval Forest (Poland and Belarus) in the 19th and 20th centuries. Acta Theriol 42:399–451
Keczyński A (2002) The effect of spruce bark beetle gradation on tree stands in the strict protection area of Białowieża National Park. Kosmos 4(257):471–474 (in Polish with English summary)
Kowalski M (1993) O sukcesji ekologicznej w lasach Jasienia (On the ecological succession in the forests of Jasienia, in Polish). Sylwan 9:37–45
Kramer K, Groen TA, van Wieren SE (2003) The interacting effects of ungulates and fire on forest dynamics: an analysis using the model FORSPACE. Forest Ecol Manage 181:205–222
Krasuska M, Miścicki S (2002) Large-scale inventory of stands in the Białowieża National Park. Sylwan 146(3):73–90 (in Polish with English summary)
Kweczlich I, Miścicki S (2004) The impact of herbivorous ungulates on forest regeneration in the Białowieża National Park. Sylwan 148(6):18–29 (in Polish with English summary)
Levanič T, Gričar J, Gagen M, Jalkanen R, Loader NJ, McCarroll D, Oven P, Robertson I (2009) The climate sensitivity of Norway spruce [Picea abies (L.) Karst.] in the southeastern European Alps. Trees Struct Funct 23:169–180
Mason SLR (2000) Fire and Mesolithic subsistence—managing oaks for acorns in northwest Europe? Palaeogeogr Palaeocl 164:139–150
McDonald JH (2009) The handbook of biological statistics. http://udel.edu/~mcdonald/statintro.html. Accessed 10 November 2009
Michalczuk C (2001) Forest habitats and treestands of Białowieża National Park. Phytocoenosis, Supplementum Cartographiae Geobotanicae 13 N.S., Warszawa-Białowieża
Miścicki S (1996) Forest regeneration and its damage by herbivorous ungulates in the Białowieża National Park. In: Paschalis P, Zajączkowski S (eds) Biodiversity protection of Białowieża Primeval Forest. Fundacja Rozwój SGGW, Warszawa, pp 91–108
Mosandl R, Kleinert A (1998) Development of oaks (Quercus petraea (Matt.) Liebl.) emerged from bird-dispersed seeds under old-growth pine (Pinus sylvestris L.) stands. Forest Ecol Manage 106:35–44
Niklasson M, Zin E, Zielonka T, Feijen M, Korczyk AF, Churski M, Samojlik T, Jędrzejewska B, Gutowski JM, Brzeziecki B (2010) A 350-year tree-ring fire record from Białowieża Primeval Forest, Poland: implications for Central European lowland fire history. J Ecology 98:1319–1329
Oldeman RAA (1990) Forests: elements of Silvology. Springer, Berlin
Olszewski JL (1986) The role of forest ecosystems in modifying local climate of the Białowieża Primeval Forest as revealed by air temperature characteristics. Ossolineum, Wrocław—Łódź (in Polish with English summary)
Packham JR, Harding DJL, Hilton GM, Stuttard RA (1992) Functional ecology of woodlands and forests. Chapman & Hall, London
Paczoski J (1930) Lasy Białowieży (Die Waldtypen von Białowieża). Państwowa Rada Ochrony Przyrody, Kraków (in Polish with German summary)
Palmer SCF, Mitchell RJ, Truscott AM, Welch D (2004) Regeneration failure in Atlantic oakwoods: the roles of ungulate grazing and invertebrates. Forest Ecol Manage 192:251–265
Paluch R (2005) Natural regeneration of oak in the Białowieża primeval forest promotional forest complex—state, conditions and prospects. Sylwan 1:30–41 (in Polish with English summary)
Peterken GF (1993) Woodland conservation and management. Chapman & Hall, London
Prusinkiewicz Z, Michalczuk C (1998) Soils of the Białowieża National Park. Phytocoenosis, Supplementum Cartographiae Geobotanicae 10 N.S., Warszawa-Białowieża
RDLP Białystok (2002) Program ochrony przyrody i wartości kulturowych w Leśnym Kompleksie Promocyjnym Puszcza Białowieska na okres 1.01.2002-31.12.2011. Regional State Forests Directoriat Białystok, manuscript [The programme of nature conservation and protection of cultural values in the Białowieża Forest Promotional Complex for the period 1.01.2002-31.12.2011] www.bialystok.lasy.gov.pl/ftp/n71/POP_LKP.pdf
Rehfuess KE (2007) On the causes of decline of Norway spruce (Picea abies Karst.) in Central Europe. Soil Use Manage 1:30–31
Samojlik T (2006) The grandest tree—a history of Scots pine (Pinus sylvestris L.) in Białowieża Primeval Forest until the end of the 18th century. Rocznik Dendrologiczny 54:7–27
Samojlik T, Jędrzejewska B (2004) Utilization of Bialowieza Forest in the times of Jagiellonian dynasty and its traces in the contemporary forest environment. Sylwan 148:37–50 (in Polish with English summary)
Schulze ED (1989) Air pollution and forest decline in a spruce (Picea abies) forest. Science 244:776–783
Sokołowski AW (1993) Phytosociological characteristics of forest communities in the Białowieża National Park. Parki Narodowe i Rezerwaty Przyrody 12:5–190 (in Polish with English summary)
Ter Braak CJF (2003) CANOCO Version 4.5A February 2003 (C) 1988–2003. Biometris—quantitative methods in the life and earth sciences. Plant Research International, Wageningen University and Research Centre, Netherlands
Van Wagner CE (1968) The line intersect method in forest fuel sampling. Forest Sci 14(1):20–26
Veblen TT (1992) Regeneration dynamics. In: Glenn-Lewin DC, Peet RK, Veblen TT (eds) Plant succession: theory and prediction. Chapman & Hall, London, pp 152–187
Vera FWM (2000) Grazing ecology and forest history. CABI, Wallingford
Watt AS (1919) On the cause of failure of natural regeneration in British oakwoods. J Ecol 7:173–203
Zajączkowski J (1999) Odnowienie lasu naturalnego na przykładzie powierzchni badawczej w Białowieskim Parku Narodowym. Sylwan 7:5–14
Acknowledgments
We are endebted to Prof. Stanisław Miścicki for sharing the unpublished data from the large-scale tree regeneration inventory. Our great thanks are due to Prof. Jan Holeksa, Prof. Jerzy Szwagrzyk, Mr Markus Feijen and two Reviewers for their help and invaluable comments on the manuscript. The study was financed through the Polish public resources dedicated to research in 2008–2010 (Ministry of Science and Higher Education, grant NN3092955).
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by G. Brazaitis.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Bobiec, A., Jaszcz, E. & Wojtunik, K. Oak (Quercus robur L.) regeneration as a response to natural dynamics of stands in European hemiboreal zone. Eur J Forest Res 130, 785–797 (2011). https://doi.org/10.1007/s10342-010-0471-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-010-0471-3