
Abstract
The rules governing bird song sequences vary considerably across the avian phylogeny, and modifications to these rules represent one of the many ways in which bird song varies interspecifically. Cassin’s Vireo (Vireo cassinii) is one species that shows a highly structured syntax, with clearly non-random patterns of sequential organization in their songs. Here I present a description of Cassin’s Vireo song sequences from the Sierra Nevada Mountains in California and employ network analysis to quantify transition patterns within the songs. Repertoire sizes varied between 44 and 60 phrase types per bird for the 13 birds analyzed here. The repertoire was subdivided into ‘themes’ containing between two and seven phrase types. The birds sang the phrase types in a given theme for a time before eventually introducing a new theme; in this manner the repertoire was revealed relatively slowly over time. Theme composition within a bird’s repertoire did not change within or between singing bouts throughout the season. The tendency to sing in themes was corroborated by network analysis, which revealed small-world structure in the songs. Phrase types were widely shared within the population. I discuss these findings as they compare with the singing styles of other species, both closely and distantly related.
Zusammenfassung
Zusammensetzung und sequenzielle Organisation der Gesangsrepertoires von Cassinvireos ( Vireo cassinii )
Die Regeln, welche die Gesangssequenzen von Vögeln regulieren, variieren beträchtlich zwischen Arten, und Modifikation dieser Regeln repräsentiert eine von vielen Weisen, auf die Vogelgesang interspezifisch variiert. Der Cassinvireo (Vireo cassinii) ist eine Art, die eine stark strukturierte Syntax aufweist, mit eindeutig nicht-zufälligen Mustern sequenzieller Organisation in den Gesängen. Hier präsentiere ich eine Beschreibung von Gesangssequenzen des Cassinvireos aus dem Sierra Nevada-Gebirge in Kalifornien und wende Netzwerkanalyse an, um Überleitungsmuster innerhalb der Gesänge zu quantifizieren. Die Größe des Repertoires variierte von 44 bis 60 Phrasentypen pro Vogel für die 13 hier analysierten Vögel. Das Repertoire wurde in „Themen“untergliedert, die zwei bis sieben Phrasentypen enthielten. Die Vögel sangen die Phrasentypen in einem bestimmten Thema für eine gewisse Zeit, bevor sie schließlich ein neues Thema einführten; auf diese Weise wurde das Repertoire relativ langsam über die Zeit hinweg offenbart. Die Themenkomposition innerhalb des Repertoires eines Vogels veränderte sich nicht innerhalb von oder zwischen Gesangsperioden im Verlauf der Saison. Die Tendenz, in Themen zu singen, wurde durch die Netzwerkanalyse bestätigt, die eine Kleine-Welt-Struktur in den Gesängen aufzeigte. Phrasentypen wurden in der Population weithin gemeinsam benutzt. Ich diskutiere diese Befunde im Vergleich mit den Gesangsstilen anderer nah und entfernt verwandter Arten.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Complex syntax is a defining feature of the songs of many species of birds. Given its prevalence in bird songs, it is likely that syntax plays a role in the conveyance of information from sender to receiver (Kershenbaum et al. 2014). The syntax of most species of birds appears to follow sets of rules, the nature and complexity of which vary greatly between species (Bolhuis and Everaert 2013), and which are sufficiently variable that the particular structure of sequences could conceivably contain information about the sender or their surroundings. To fully understand the information contained within the songs of any species, it is first necessary to evaluate in detail the nature of their song repertoire and syntax.
Though most songbird species utilize song repertoires comprising multiple vocal units, and are therefore potential subjects of studies of syntax, research to date has overwhelmingly focused on variation in repertoire size and composition between species (e.g., Byers and Kroodsma 2009), while largely ignoring the question of how the vocal units are combined into sequences. To do so overlooks the possibility that the ordering of songs may convey biologically relevant information about individual fitness (Leitner et al. 2006) or external factors such as predators (Baker and Becker 2002). Furthermore, in some species investigations into song syntax have provided more insight into the nature of songs than subjective estimates of repertoire size alone. For example, re-arrangement of relatively few vocal units according to probabilistic rules gives rise to considerable variability in the songs of many species (e.g., Sedge Warbler, Catchpole 1976; Willow Warbler, Gil and Slater 2000; Brown-throated Wren, Sosa-López and Mennill 2013; Bengalese Finch, Jin and Kozhevnikov 2011).
The songs of birds are subject to selection based on various ecological variables, including habitat characteristics (Morton 1975; Anderson and Conner 1985; Slabbekoorn and Smith 2002; Boncoraglio and Saino 2007), mating systems (Catchpole 1987, but see Soma and Garamszegi 2011) and migratory distance (Read and Weary 1992; Mountjoy and Leger 2001). Detailed and accurate descriptions of the songs of a given species can greatly facilitate future efforts to understand the influence of these factors on the evolution of bird songs. In this study, I will lay such a foundation by describing the songs of Cassin’s Vireo (Vireo cassinii) with a focus on the syntactical patterns that emerge over the course of long bouts of song.
Cassin’s Vireo is a migratory songbird that breeds in the forests of western North America. Past work analyzing the songs of the Solitary Vireo complex included only brief analysis of the songs of Cassin’s Vireo (James 1981), but the recent elevation of Cassin’s Vireo to full species status warrants a revisitation of past efforts (American Ornithologists’ Union 1997). Cassin’s Vireo belongs to the Vireonidae, a family whose species display a striking variety of singing styles. Songs vary from the simple repeated phrases of Hutton’s Vireo, a species that possesses small repertoires of approximately three phrase types (Mountjoy and Leger 2001), to the highly variable songs of the Warbling Vireo, which combines subunits to produce a heretofore unquantified variety of songs (Howes-Jones 1985). Intermediate between these two extremes are a few close relatives of Cassin’s Vireo: Blue-headed Vireo repertoires have been estimated to contain 12 phrase types (James 1981), while Yellow-throated and White-eyed Vireos possess repertoires of 5 (James 1984) and 12 songs (Borror 1987), respectively. The intermediate repertoire sizes of these species appear to be representative of the genus: a review of 28 Vireo species by Mountjoy and Leger (2001) showed a mean repertoire size of 12.
This article aims to identify the repertoire sizes of Cassin’s Vireo individuals and outline the patterns of repertoire delivery that became apparent over the course of extensive recordings on a population of 13 individuals. I begin with an assessment of the repertoire sizes of the males in the focal population, using classification algorithms to confirm judgments of discrete phrase types. Then I illustrate patterns of song sharing among males. Finally, I describe the sequential arrangement of the songs and discuss the findings with respect to previously published work on the syntax of the songs of Vireos and other songbirds.
Methods
Study site
Fieldwork was conducted on private land 5 km north of the town of Volcano in Amador County, California, USA, in the foothills of the Sierra Nevada Mountains at an elevation of 750 m (10 S 706584 4262742, datum WGS 84). The forest is characterized by mixed evergreen and deciduous trees; the tree community is dominated by Incense Cedar (Calocedrus decurrens), Douglas Fir (Pseudotsuga menziesii), Live Oak (Quercus chrysolepis), Black Oak (Quercus kelloggii) and Ponderosa Pine (Pinus ponderosa). Fieldwork took place between 25 April and 28 June 2013 and between 5 May and 25 June 2014.
Field methods
I made recordings using a Marantz PMD661 solid-state digital recorder and a Sennheiser MKH20-P48 microphone with a Telinga parabolic reflector. Recordings were saved to wav files using a sampling rate of 44 kHz. Recordings were made opportunistically: I first located an individual aurally, then approached and recorded it for as long as possible, stopping only when the bird either ceased singing for a considerable period of time or moved too far away to permit a sufficiently high quality recording. Due to the unpredictable nature of the birds’ behaviors, recordings varied in the number of phrases they contained. Since the goal of the study was to investigate patterns of sequential delivery, I discarded recordings containing fewer than 50 phrases. The resulting data set contained 216 recordings, varying in length from less than 2 min to more than 2 h (mean 0:25:02, median 16:44) and containing between 50 and 1525 phrases per recording (mean 252, median 173).
Seven males were recorded in 2013. Five of these individuals returned to breed in the same area in 2014 and were thus recorded in both years; a further 6 males were recorded in 2014, for a total of 13 males in the focal population. During the 2013 season, individuals were identified based on their consistent association with non-overlapping territories, and identifications were later confirmed based on the phrase types in the recording, justified by the observation that each male possesses an individually distinct repertoire. To identify males between years, I again capitalized on the song characteristics and identified individuals using an ensemble of classifiers that has been shown to identify individuals with near perfect accuracy (J. Arriaga, unpublished). In the five instances where the algorithm identified the same male at the study site in both years, the male established a territory with roughly the same boundaries, providing further evidence that the identification was accurate (Online Resource 2). In early May of 2014, all males were banded with unique color combinations for subsequent visual identification. Individuals are here referred to by a unique letter combination corresponding to the colors of their band combination, with the exception of the two birds that were only recorded in 2013, which are referred to as ‘Meadow’ and ‘Gate’ individuals. The number of phrases recorded from each individual ranged from 1335 to 10,336 (mean 4193). The aggregate number of phrases annotated from all 13 birds was 54,515. All recordings and annotations used in this analysis are available online in a birdsong database at http://taylor0.biology.ucla.edu/birdDBQuery; links to specific recordings and their corresponding annotations are provided in Online Resource 1. In addition, the recordings used to produce the spectrograms in Figs. 1 and 7 are available from the Animal Sound Archive at the Museum für Naturkunde, Berlin (http://www.animalsoundarchive.org; see Electronic Supplementary Material).

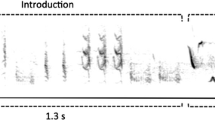
A spectrogram illustrating the typical singing style of Cassin’s Vireo. Four phrases of three phrase types are shown, forming the sequence aq, ar, dl, aq. Comparison of the two exemplars of aq demonstrates the strongly stereotyped delivery of phrases, with very little within-type variation. The third phrase in the sequence, dl, is a compound phrase type, comprised of two subunits that themselves are occasionally delivered independently as the phrase types dr and en (ESM_3_Online Resource 3 and ESM_4)
I plotted individual territories onto a map of the study area throughout each season by observing singing bouts and counter-singing interactions between neighbors (Online Resource 2). Some territories abutted one another, while others were separated by up to 700 m.
Terminology
Assessments of repertoire sizes and interpretations of song complexity can vary with the level of analysis; thus, it is important to explicitly define the elements of songs as they pertain to this study. Phrases are short bursts of sound less than 0.7 s in duration (mean 0.34 s) and usually separated from preceding and succeeding phrases by at least 1 s of silence (Fig. 1). Phrase types are categories of phrases that are considered equivalent based on shared spectrographic characteristics. A song bout was defined as a long string of phrases delivered in sequence.
Phrase identification and annotation
Cassin’s Vireos deliver songs in two audibly distinct ways: their typical song consists of phrases separated by ≥1 s of silence, while their chatter vocalizations are sequences of phrases and call notes delivered rapidly and continuously without pauses. Previous authors have referred to these latter vocalizations as ‘pre-copulatory songs’ (James 1978). For the purposes of this study, I focused exclusively on the typical song, which accounted for more than 99 % of the total vocal output of the birds. I have observed that phrase types found in the chatter vocalizations are often distinct from those in the typical song; individuals that do not deliver a certain phrase type in their typical repertoire may include it in their chatter songs. The structure and function of chatter songs remain as topics for future study.
I used the linguistic program Praat (Boersma and Weenink 2014) to annotate recordings. By visually inspecting spectrograms from 216 recordings of 13 individuals, I created a catalog containing the spectrograms of phrase types against which subsequent phrases could be compared (Online Resource 3). Each phrase type was assigned a unique two-letter code and was subjectively identifiable based on spectrographic characteristics and acoustic structure.
I marked the start and end times of phrases in each recording with approximately ±0.01 s accuracy, then identified the phrase types by visually comparing their spectrograms with the reference file. Phrases were highly stereotyped, such that there were few visible differences between any two exemplars of the same phrase type and phrase-type classification was unambiguous. Portions of some recordings were not annotated because of uncertainty regarding the identity of the bird. In such cases where identity was in doubt or recording quality suffered, I annotated only the recording either before or after the unusable portion.
The classification of phrase types by visually inspecting spectrograms is inherently subjective, but in my experience has proven to be the most noise-robust technique currently available for this species. Since my ultimate goal was to study the sequential arrangements of phrase types in the songs of Cassin’s Vireos, I could not sacrifice accuracy or noise robustness for the speed permitted by automated classification algorithms. Still, to ensure that my subjective classifications represented objectively identifiable and discrete phrase types, I verified a subset of my annotations using a supervised machine learning algorithm implemented by Tan et al. (2013); this is a sparse-representation algorithm that uses spectrographic features to perform classification. The algorithm was designed specifically for use on Cassin’s Vireo phrases and has been shown to perform at a high level on recordings with minimal background noise (Tan et al. 2012, 2013). I ran two tests with the classification algorithm. In both cases, I used only clean recordings for both training and testing, meaning I first manually inspected the annotated sound files and removed all instances of phrases with low signal-to-noise ratios. For the first test, I trained the algorithm using four recordings from four different individuals and tested on phrases from seven individuals at the study site. This tested the ability of the algorithm to generalize across individuals and tested whether my evaluation of the extent of phrase-type sharing between males was supported. In the second test, I trained the algorithm using a single recording from seven individuals and tested on 47 files from those same individuals. The algorithm used five exemplars of each phrase type in the training set, ignoring phrase types that occurred less than five times in the training files. As a result, the algorithm was not trained and tested on all phrase types used by the focal birds. In the first test, the algorithm was tested on 6863 examples of 83 phrase types, and in the second it was tested on 7145 examples of 90 phrase types.
Network analysis
I calculated the small-world coefficient for each of the 216 recordings. Networks that meet the criteria for small worldness are characterized by high local clustering and short path lengths between vertices; small-world structure has been identified in social networks, power grids and neural networks (Watts and Strogatz 1998). Measures of small worldness applied to bird song measure the tendency for phrase types to be clustered together within a bird’s repertoire, such that certain phrase types consistently appear near each other in sequences more than would be expected by chance. The methods I employed were identical to those of Sasahara et al. (2012), who identified small-world structure in the songs of California Thrashers. For more detail on small-world algorithms, see Watts and Strogatz (1998) and Humphries and Gurney (2008).
I first converted each recording into a transition graph, with each phrase type represented as a vertex and each observed transition between phrase types represented as an edge. Self-loops (in which a phrase type was repeated more than once) were removed from the graph, and the directionality was removed from the transitions. The small-world coefficient was calculated using the following formula:
where C is the average clustering coefficient of the transition graph from a given recording (a measure of the average number of connected edges with respect to the maximum number of edges possible) averaged for each phrase type in the recording. L is the characteristic path length (the average number of steps along the shortest paths between all vertices in the graph). C rand and L rand are the same measures calculated from randomly generated graphs with the same number of edges and vertices as the observed graph. For each recording 5000 random graphs were generated, and the C and L values for these graphs were averaged to arrive at estimates for C rand and L rand. The small-world coefficient, S, thus compares the clustering of phrase types from observed song sequences to randomly generated networks. A value of S greater than 1 indicates a tendency to organize phrase types into themes with more connections between phrase types within the theme than to phrase types outside of the theme (Humphries and Gurney 2008). Network analysis was conducted using the PajaroLoco package (Sanchez et al. 2015) in Mathematica version 9.0 (Wolfram Research Inc. 2012).
Results
Repertoires and sharing
In total, I identified 122 phrase types in the focal population and individual repertoire sizes for the 13 birds varied between 44 and 60 phrase types (mean 51.07). Most phrase types appeared as continuous traces on a spectrogram. Nineteen of the phrase types, however, were ‘compound,’ in that they were comprised of two subunits separated by a very short interval of silence. Some of the subunits of compound phrase types were themselves independent phrase types (Fig. 1), while other subunits were only delivered as part of a compound phrase type. Compound phrase types were considered single phrases because: (1) the two subunits were closely and consistently associated with one another with only very short periods of silence (~0.1 s) between subunits; (2) some individuals sang compound phrase types without ever singing either of the component subunits independently.
Phrase-type sharing was common, and pairwise comparisons between birds showed that males shared on average 25.4 ± 4.9 phrase types with any other male. A Mantel test conducted with the ‘ade4’ package in R version 3.1.2 (R Core Team 2014) comparing the pairwise repertoire similarity—calculated as \({\text{C/(}}\sqrt A \times \sqrt B )\), where C is the number of shared phrase types between two individuals, and A and B are their respective repertoire sizes—with the distance between territories showed no significant relationship between the two variables in either year (2013: r = −0.06, p = 0.61, n = 7 individuals; 2014: r = 0.1033, p = 0.26, n = 11 individuals; 9999 permutations).
Of the 122 phrase types encountered in the study, 107 were shared between at least 2 of the 13 focal individuals. Two phrase types were identified in the repertoires of all 13 individuals, while on average each phrase type was identified in the repertoires of 5.4 of 13 individuals. The total number of phrase types encountered in the population showed signs of leveling off as individuals were added to the sample (Fig. 2), an indication that few of the phrase types were unique to any individual.

Phrase-type accumulation curve showing the number of unique phrase types in a sample as individuals are added. The cumulative number of phrase types encountered (solid line) within a population increased as a function of the number of individuals sampled. The number of new phrase types encountered (dotted line) declined as more individuals were added to the sample, a result of the high levels of phrase-type sharing in the population
Classification results
In the first experiment, in which I trained the algorithm using exemplars from four individuals and tested the algorithm’s ability to generalize across individuals, the sparse representation algorithm agreed with my spectrogram inspection method on 99.4 % of the 6863 phrases tested. The second experiment, in which I trained the algorithm on a single sound file from each of seven individuals, showed similar results (99.4 % agreement on 7145 phrases). I conclude that the phrase-type categories, while subjectively determined by a human at first, did provide a consistent and objective basis for analyzing songs of the species.
Phrase type use
Phrase types were not used equally within an individual’s repertoire (chi-square goodness of fit test: p < 0.0001 for each of 13 individuals; Fig. 3). The most common phrase type in each individual’s repertoire comprised an average of 9.5 % (range 6.1–13.4 %) of the total phrases delivered. The rarest phrase types in each bird’s repertoire comprised an average of only 0.07 % (range 0.0009–0.5 %) of the total phrases.

Relative use of phrase types from recordings of ‘AOBu’ individual. Phrase types were arranged by rank from most common to least common. Analysis of all individuals yielded qualitatively similar results, showing strong biases favoring the delivery of some phrase types over others
Sequential arrangement of phrases
Consecutive phrases comprised distinct phrase types 93.4 % of the time, so song organization strongly tended toward immediate rather than eventual variety. The average small-world coefficient from all recordings was 3.73, reflecting a strong tendency toward grouping phrase types into themes. Sequences from only 1 of 216 recordings failed to meet the threshold for small worldness (Online Resource 1), and this was likely a result of the short length of the recording, which contained just 58 phrases. Supporting this idea, there was a significant positive correlation between the number of phrases in a recording and its small-world coefficient (Pearson: r 2 = 0.177, N = 216, p < 0.0001).
A representative sequence of phrases is illustrated in Fig. 4 and shows the typical singing style for the species. In this figure, phrase types were assigned numbers according to the order in which they were introduced. The male delivered phrase types in themes, such that groups of phrase types were consistently delivered in close succession. Each theme contained between two and seven phrase types, and themes are clearly evident as clusters of points within Fig. 4. The particular order of phrase types was variable within each theme, although the magnitude and nature of this variability remains to be investigated. The identity of the phrase types comprising the themes differed between individuals; within the songs of a given male, however, theme composition was consistent over the course of the season. Temporal conservation of theme composition is evident over a short timeframe in Fig. 4, where the bird repeated previously delivered themes after approximately 120 phrases had been delivered in the sequence; in each case, the phrase types in the themes were unchanged from previous instances of the same theme. After delivering each phrase type in a given theme one or a few times (mean 3.17, median 2), another theme was introduced with minimal overlap between themes. Themes were generally only revisited after a considerable portion of the repertoire had been delivered. All 13 individuals showed similar tendencies with regards to their arrangements of songs, organizing their phrase types into discrete themes that were consistently delivered together in sequence and were conserved over the course of the season.

Graphical representation of the sequential arrangement of a song bout from a single recording of the ‘Meadow’ individual. Phrase numbers were assigned based solely on the order of first appearance of each phrase type within the recording. Themes are clearly identifiable as clusters of phrase types regularly associated with one another. Phrase types au, ae, and en are denoted with horizontal lines, illustrating the distinctness of these three spectrographically similar phrase types in the bird’s repertoire. The style of singing illustrated here is typical of all individuals analyzed
A notable consequence of the use of themes is that once a given phrase type has been delivered, it is likely to recur again within the next few phrases because transitions between phrase types within a theme occur more frequently than transitions to other phrase types in the repertoire. A useful way of illustrating this is by calculating the recurrence interval, defined as the number of intervening phrases between two examples of a given phrase type. For all individuals, recurrence intervals up to eight were reasonably common, while intervals longer than eight were increasingly rare (Fig. 5). All individuals showed a similar trend, with peak recurrence intervals of one, two or three.

Distribution of recurrence intervals for all 13 individuals in the population, showing a strong tendency toward low recurrence intervals. Horizontal dashed lines show the mean for each value of the recurrence interval, and the solid line connects the mean values to illustrate the overall trend. Intervals greater than 8 occurred at low rates, but were not included here
Discussion
Sequential arrangement of phrases
The patterns of sequential phrase delivery described here illustrate the inadequacy of the categorization of birds as singing with either immediate or eventual variety. Within the broad category of ‘immediate variety,’ there are many possibilities for species-specific syntax; typically these have been overlooked in discussions of the evolutionary significance of song repertoires. As an example, some aspects of Cassin’s Vireo syntax run counter to the idea that large song repertoires are maintained by intersexual selection. First of all, the observation that some phrase types are exceptionally rare, being encountered only a few times in samples containing as many as several thousand phrases per individual, would not be expected under a female-choice model (Fig. 3). Under such a model—commonly invoked to explain large repertoires and immediate variety—males should deliver all phrase types approximately equally to increase their effective repertoire size (Byers and Kroodsma 2009). In the songs of Cassin’s Vireos, however, many phrase types in a male’s repertoire were very rarely delivered. Furthermore, under a female choice model, males should arrange their songs cyclically to advertise their repertoire size in an expedient manner. The tendency of Cassin’s Vireos to arrange their songs into themes that are often delivered at length before additional themes are introduced contradicts the patterns expected under an intersexually selected song repertoire. In the case of Cassin’s Vireo, at least, alternate explanations seem required to explain the large repertoires and complex syntax of the species.
The organization of phrase types into small-world themes is not unique to Cassin’s Vireos. The same finding has been identified in California Thrashers (Sasahara et al. 2012), Southern House Wrens (Deslandes et al. 2014) and Nightingales (Weiss et al. 2014). A similar hierarchical song structure has been qualitatively described in Rock Wrens (Kroodsma 1975), though the author did not employ the same network-based approach to quantify the strength of the pattern. Common to all of these species are large repertoires of song types, phrase types or syllable types. This may indicate a broad convergence to small-world structure among birds with complex song repertoires—at the very least it shows widespread non-random arrangements of song sequences.
Even within the family Vireonidae, the style of singing described here may not be unique. Black-capped Vireos (Vireo atricapilla) tend to group their songs into groups of three to seven phrase types, switching between groups in a fashion comparable to that described here for Cassin’s Vireo (Grzybowski 1995). Bell’s Vireos (Vireo bellii) tend to alternate between groups of two or three song types before abruptly switching to another group of song types (Kus et al. 2010). Similarly, Thick-billed Vireos (Vireo crassirostris) alternate between two song types for a period of time before switching to another pair of song types (M.R. Walker, unpublished). Although these patterns have not yet been formally analyzed, these singing styles appear to indicate a phylogenetically conserved tendency to group vocal units into themes. Syntax may therefore be influenced in part by the complexity of the repertoire, while also being strongly influenced by evolutionary history.
Repertoire size
The average repertoire size of 51 phrase types per individual is 325 % larger than the previously published repertoire size for Cassin’s Vireo. James (1981) recorded five individuals from a population in British Columbia and identified an average of 12 phrase types per individual, with a range of between 6 and 14 phrase types. Although he recorded a total of only 251 phrases, or approximately 50 phrases per bird, he maintained that his methodology approached the full repertoire size of the birds in his study population. Subsequent studies on Blue-headed Vireos (Vireo solitarius) have identified comparable repertoire sizes of approximately 15 types, lending some credibility to the reproducibility of James’ estimates (Martindale 1980).
The reasons for such a significant difference in repertoire size estimates are unclear. Incomplete sampling on the part of James (1981) most likely played a role; in my study population, new phrase types were still being encountered after thousands of phrases had been recorded from a single individual (Fig. 6). The singing style of the species, in which the bird occasionally delivers a single theme for an extended period of time, may have exacerbated this issue, by giving previous authors a false sense that the repertoire had been completely sampled. The possibility of geographic variation in repertoire sizes could also account for this and has been identified in other species (McGregor et al. 1981; Kroodsma 1983; Baker 1996; Irwin 2000; Peters et al. 2000).

Cumulative numbers of phrase types encountered in the songs of 13 focal individuals over the course of the study. Most phrase types were encountered within the first few hundred recorded phrases, but a more substantial recording effort was required to confidently estimate total repertoire sizes. Numbers in parentheses represent the final repertoire size estimate for each bird followed by the total number of phrases recorded from that bird
Another potential cause of the repertoire size differences is methodological: distinguishing between phrase types by inspecting spectrograms is inherently subjective. Martindale (1980) borrowed terminology from systematics when he framed the issue as one of ‘lumpers’ versus ‘splitters,’ wherein lumpers tend to categorize similar phrase types as being the same, and splitters would allow even slight differences to characterize distinct types, so long as those differences are consistent. My approach, justified by the remarkable stereotypy in the songs of Cassin’s Vireo, tended toward the ‘splitter’ approach. I believe this approach is generally vindicated by objective methods of supervised learning, discussed above.
The ultimate goal of either approach should be to accurately assess the phrase types as employed by the birds themselves. In the case of Cassin’s Vireos, the sequential arrangements of phrase types provide hints about individual- and population-level treatment of song types. In Fig. 7, for example, which depicts spectrographically similar phrases from the ‘Gate’ and ‘Meadow’ individuals from my study site, each phrase type has clearly distinct acoustic characteristics, and these differences are conserved between individuals. Inspection of Fig. 4, a depiction of a long sequence of the Meadow individual’s song, shows that these three phrase types were treated as distinct. Phrase type ae, labeled as phrase number 14 in the figure, was delivered twice: both times it was followed by the theme containing phrases numbered 15–19. In contrast, phrase types au and en, labeled as phrase numbers 6 and 28, respectively, were consistently delivered together in a theme with numbers 4, 5 and 7. Moreover, some individuals sang only one or two of the three types, but never strayed from the stereotyped features of the phrase types (Fig. 7). Distinctions between phrase types were confirmed by the sparse-representation algorithm, which showed almost complete agreement with my classifications of phrase types in the population. Taken together, my findings suggest that the repertoires of Cassin’s Vireos are much larger than has been previously appreciated.

Spectrograms of three similar phrase types from the repertoires of two different birds. Examples a, c and e are from the ‘Meadow’ individual, while examples b, d and f were recorded from the ‘Gate’ individual. Examples a and b show phrase type ae, c and d show phrase type au, and e and f show phrase type en. Despite obvious similarities among these three phrase types, variation in spectral characteristics between phrase types consistently exceeds within-type variation, even when exemplars from different individuals are considered. For the recordings and the data base, see ESM_4
Song sharing
Rates of phrase-type sharing between the focal individuals were high, with pairs of individuals sharing approximately half of their repertoires on average. Even more striking was the fact that the vast majority of phrase types were shared between at least two individuals. Only 15 of the 122 phrase types in the sample were unshared among individuals, and subsequent opportunistic recording efforts from elsewhere in the Sierra Nevada Mountains have identified a further 10 of these phrase types in the repertoires of other males (R. Hedley, unpublished data). This raises questions about whether the species is capable of improvising phrase types or whether individuals require tutoring for repertoire development. The near complete absence of unshared phrase types suggests a role for imitation, although the lack of a significant relationship between inter-territory distance and sharing rates leaves open the question of when and from whom they might acquire their songs. A further question, especially worthwhile for a species with such a distinct syntax, is: do individuals share common rules for the sequential delivery of shared phrase types? If not, are such differences in syntax meaningful to the birds? These questions and questions regarding the functional role of syntax will provide ample directions for future research.
References
American Ornithologists’ Union (1997) Forty-first supplement to the American Ornithologists’ Union check-list of North American Birds. Auk 114:542–552
Anderson ME, Conner RN (1985) Northern Cardinal song in three forest habitats in eastern Texas. Wilson Bull 97:436–449
Baker MC (1996) Depauperate meme pool of vocal signals in an island population of singing honeyeaters. Anim Behav 51:853–858
Baker MC, Becker AM (2002) Mobbing calls of black-capped chickadees: effects of urgency on call production. Wilson Bull 114:510–516
Boersma P, Weenink D (2014) Praat: doing phonetics by computer. Version 5.3.82. http://www.praat.org/
Bolhuis JJ, Everaert M (2013) Birdsong, speech, and language: exploring the evolution of mind and brain. MIT Press, Cambridge
Boncoraglio G, Saino N (2007) Habitat structure and the evolution of bird song: a meta-analysis of the evidence for the acoustic adaptation hypothesis. Funct Ecol 21:134–142
Borror DJ (1987) Song in the white-eyed vireo. Wilson Bull 99:377–397
Byers BE, Kroodsma DE (2009) Female mate choice and songbird song repertoires. Anim Behav 77:13–22. doi:10.1016/j.anbehav.2008.10.003
Catchpole CK (1976) Temporal and sequential organisation of song in the Sedge Warbler (Acrocephalus schoenobaenus). Behaviour 59:226–246
Catchpole CK (1987) Bird song, sexual selection and female choice. Trends Ecol Evol 2:94–97. doi:10.1016/0169-5347(87)90165-0
Deslandes V, Faria LRR, Borges ME, Pie MR (2014) The structure of an avian syllable syntax network. Behav Processes 106:53–59. doi:10.1016/j.beproc.2014.04.010
Gil D, Slater PJB (2000) Song organisation and singing patterns of the Willow Warbler, Phylloscopus trochilus. Behaviour 137:759–782
Grzybowski JA (1995) Black-capped Vireo (Vireo atricapilla). In: Poole A (ed) The birds of North America online. Cornell Lab of Ornithology, Ithaca. doi:10.2173/bna.181
Howes-Jones D (1985) The complex song of the Warbling Vireo. Can J Zool 63:2756–2766
Humphries MD, Gurney K (2008) Network “small-world-ness”: a quantitative method for determining canonical network equivalence. PLoS One 3:e0002051. doi:10.1371/journal.pone.0002051
Irwin DE (2000) Song variation in an avian ring species. Evolution 54:998–1010
James RD (1978) Pairing and nest site selection in solitary and yellow-throated Vireos with a description of a ritualized nest building display. Can J Zool 56:1163–1169
James RD (1981) Factors affecting variation in the primary song of North American Solitary Vireos (Aves: Vireonidae). Can J Zool 59:2001–2009
James RD (1984) Structure, frequency of usage, and apparent learning in the primary song of the yellow-throated vireo, with comparative notes on solitary vireos (Aves: Vireonidae). Can J Zool 62:468–472. doi:10.1139/z84-071
Jin DZ, Kozhevnikov AA (2011) A compact statistical model of the song syntax in Bengalese Finch. PLoS Comput Biol 7:e1001108. doi:10.1371/journal.pcbi.1001108
Kershenbaum A, Blumstein DT, Roch MA et al (2014) Acoustic sequences in non-human animals: a tutorial review and prospectus. Biol Rev. doi:10.1111/brv.12160
Kroodsma DE (1975) Song patterning in the Rock Wren. Condor 77:294–303
Kroodsma DE (1983) The ecology of avian vocal learning. Bioscience 33:165–171
Kus B, Hopp SL, Johnson RR, Brown BT (2010) Bell’s Vireo (Vireo bellii). In: Poole A (ed) The birds of North America online. Cornell Lab of Ornithology, Ithaca. doi:10.2173/bna.35
Leitner S, Marshall RC, Leisler B, Catchpole CK (2006) Male song quality, egg size and offspring sex in captive canaries (Serinus canaria). Ethology 112:554–563. doi:10.1111/j.1439-0310.2006.01195.x
Martindale S (1980) A numerical approach to the analysis of Solitary Vireo songs. Condor 82:199–211
McGregor PK, Krebs JR, Perrins CM (1981) Song repertoires and lifetime reproductive success in the Great Tit (Parus major). Am Nat 118:149–159
Morton ES (1975) Ecological sources of selection on avian sounds. Am Nat 109:17–34
Mountjoy DJ, Leger DW (2001) Vireo song repertoires and migratory distance: three sexual selection hypotheses fail to explain the correlation. Behav Ecol 12:98–102. doi:10.1093/oxfordjournals.beheco.a000385
Peters S, Searcy WA, Beecher MD, Nowicki S (2000) Geographic variation in the organization of song sparrow repertoires. Auk 117:936–942
R Core Team (2014) R: a language and environment for statistical computing
Read AF, Weary DM (1992) The evolution of bird song: comparative analyses. Philos Trans R Soc B Biol Sci 338:165–187. doi:10.1098/rstb.1992.0137
Sanchez HM, Vallejo EE, Taylor CE (2015) PajaroLoco: a suite of programs to study the grammatical structure of bird songs. In: Proceedings of the 20th international symposium on artificial life and robotics (AROB 20th 2015), Beppu, Japan, 21–23 Jan 2015, pp 148–153
Sasahara K, Cody ML, Cohen D, Taylor CE (2012) Structural design principles of complex bird songs: a network-based approach. PLoS One 7:e44436. doi:10.1371/journal.pone.0044436
Slabbekoorn H, Smith TB (2002) Habitat-dependent song divergence in the little greenbul: an analysis of environmental selection pressures on acoustic signals. Evolution 56:1849–1858
Soma M, Garamszegi LZ (2011) Rethinking birdsong evolution: meta-analysis of the relationship between song complexity and reproductive success. Behav Ecol 22:363–371. doi:10.1093/beheco/arq219
Sosa-López JR, Mennill DJ (2013) The vocal behavior of the brown-throated Wren (Troglodytes brunneicollis): song structure, repertoires, sharing, syntax, and diel variation. J Ornithol 155:435–446. doi:10.1007/s10336-013-1024-6
Tan LN, Kaewtip K, Cody ML, Taylor CE, Alwan A (2012) Evaluation of a sparse representation-based classifier for bird phrase classification under limited data conditions. Interspeech 2012:3–6
Tan LN, Alwan A, Kossan G, Cody ML, Taylor CE (2013) A sparse representation-based classifier for in-set bird phrase verification and classification with limited training data. ICASSP 2013:763–767
Watts DJ, Strogatz SH (1998) Collective dynamics of ‘small-world’ networks. Nature 393:440–442
Weiss M, Hultsch H, Adam I, Scharff C, Kipper S (2014) The use of network analysis to study complex animal communication systems: a study on nightingale song. Proc R Soc B 281:20140460
Wolfram Research Inc. (2012) Mathematica, version 9.0. Champaign, IL
Acknowledgments
This work was supported in part by National Science Foundation Award Number 1125423. Thanks to C. Taylor, M. Cody, J. Arriaga, H. Sanchez, E. Vallejo and L. Tan for providing constructive feedback throughout the duration of the study. All work presented here complied with the current laws in the USA.
Conflict of interest
The author declares that he has no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by O. Krüger.
Electronic supplementary material
Below is the link to the electronic supplementary material.
10336_2015_1238_MOESM1_ESM.xlsx
ESM_1_Online Resource 1: Spreadsheet of recordings used in this study, with relevant metadata and links to the online locations of all wav files and corresponding textgrid files for use in Praat (XLSX 35 kb)
10336_2015_1238_MOESM2_ESM.eps
ESM_2_Online Resource 2: Map of territories held by the 13 male Cassin’s Vireos recorded during the study. Individuals recorded in both years held overlapping territories that are depicted with the same color on the map. Territories drawn with thin borders were breeding territories in 2013, while those with thick borders were held in 2014 (EPS 3706 kb)
10336_2015_1238_MOESM3_ESM.pdf
ESM_3_Online Resource 3: Catalog of spectrographic images of phrase types sung by the 13 focal birds recorded at in 2013 and 2014. All images show spectrograms of the frequency range from 0 to 10 kHz, and the horizontal red dotted lines illustrate the 3-kHz frequency to facilitate visual comparisons between phrase types. Two-letter codes were assigned to each visually and audibly distinct phrase type (PDF 1251 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Hedley, R.W. Composition and sequential organization of song repertoires in Cassin’s Vireo (Vireo cassinii). J Ornithol 157, 13–22 (2016). https://doi.org/10.1007/s10336-015-1238-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-015-1238-x