
Abstract
Filtration capacity and feeding behaviour has been intensely studied for adult mussels (Mytilus edulis), but less information is available for juvenile mussels (1.5–25 mm, <1 year), especially in natural sea water. The recent introduction of mussel seed collectors in the Netherlands prompted the need for more detailed information on juvenile mussel behaviour. To estimate the impact of juvenile populations on ecosystem carrying capacity, information on clearance rate as well as usage of different prey items is essential. Clearance rates were measured in an experimental study, incubating juvenile mussels in natural sea water. Rates were related to isometrics as well as specified for the prey items bacteria, picophytoplankton (<3 μm), nanophytoplankton (3–20 μm), and ciliates. Results showed that the clearance rate of juvenile mussels depends on shell length2, while the relationship between clearance rate and weight was more variable. Length is thus a better parameter for estimating clearance rate than weight. Ciliates and nanophytoplankton were cleared at comparable, but variable rates, while picoalgae were cleared from the water at the rate of 11–64 % compared to nanophytoplankton. For bacteria, the clearance rate was on average 9 %. This study showed different retention of particles of similar size (picoalgae and bacteria) as well as variability in particle retention for the different prey items. This variable retention efficiency could not be related to seston concentration nor to dominance in cell size. The results from this study can be used to estimate the effect of mussel seed collectors on the carrying capacity of the Dutch Wadden Sea.
Similar content being viewed by others
Introduction
In estuarine ecosystems, suspension-feeding bivalves, like the blue mussel (Mytilus edulis), often occur in large numbers, affecting the surrounding ecosystem by filtering vast volumes of water, thereby removing different components of the plankton community (e.g. Verwey 1952; Cadée and Hegeman 1974; Cloern 1982; Dame 1996; Kreeger and Newell 1996; Wong and Levinton 2006). The recent introduction of mussel seed collectors in the Netherlands prompted the need for assessing the effect of large numbers of juvenile individuals on the carrying capacity of the surrounding ecosystem. Pelagic seed collectors facilitate the settlement of mussel larvae (300 µm). After settlement in June, the juveniles grow in <6 months to a maximum size of 25 mm at harvest (Jacobs et al. 2014). There have been numerous studies performed on the filtration capacity and feeding behaviour of mussels, but these studies were mainly confined to larger (>15 mm) individuals (e.g. Widdows 1978; Bayne and Widdows 1978; Møhlenberg and Riisgård 1979; Riisgård et al. 1980, 2014; Kiørboe and Møhlenberg 1981; Jones et al. 1992; Smaal et al. 1997), while smaller individuals have been studied far less intensively (but see Riisgård et al. 1980). Most studies on filtration rates of mussels were performed under controlled laboratory conditions using algal cultures. These experiments resulted in estimates of the maximum clearance rate, while it can be expected that under natural conditions, clearance rates will be lower. The need for information on actual realised clearance rates under natural conditions and the specific usage of natural plankton by these dense collections of juvenile mussels has been recognised (Bunt et al. 1992; Cranford et al. 2003, 2011; Trottet et al. 2008).
Mussel larvae are suspension feeders, utilising a ciliated velum to capture food particles (Riisgård et al. 1980). After settlement and during metamorphosis, the feeding modus changes from a velum to the ctenidium, which also serves as a respiratory organ (gills) (Cranford et al. 2011). Lateral cilia on the gill filaments create an inflow; water enters the inhalant chamber and flows through the gills towards the exhalant chamber. Particles in the water flow are captured when the frontal surfaces of the ctenidial filaments encounter and retain them. The size of particles efficiently retained depends on the size and complexity of the latero-frontal cilia of the filaments as well as the current produced by the cirri (Newell and Shumway 1993; Dame 1996; Ward and Shumway 2004).
Captured and retained particles are transported to the labial palps. Here, particles are either rejected as pseudofaeces or directed further to the mouth (Ward and Shumway 2004).
The assumption of isometric relationships between length, area, and volume (area~length2 and volume~length3) was more variable; this leads to the expectation that theoretically pumping or filtration rate (R F) scales with gill surface area, and gill surface area is expected to scale with length2, so R F = length2. Since weight scales with volume and volume scales with length3, gill area will scale with weight2/3 and filtration rate will thus also scale with weight2/3, so R F = weight2/3 (Jones et al 1992). For veliger and post-metamorphosed larvae, filtration rate was reported to scale with weight0.8–1 (Riisgård et al. 1980; Beiras and Camacho 1994). The high scaling factor was attributed to a high non-isometric growth of the gills.
In most studies, clearance rate (R C), which is the volume cleared of particles per unit time, is measured rather than the actual pumping or filtration rate. When particles are 100 % efficiently retained by the gills, the clearance rate equals the filtration rate. If the filtration efficiency is lower than 100 %, the clearance rate is thus lower than the pumping rate.
Numerous studies, starting with a study by Møhlenberg and Riisgård (1978), have reported on the particle size range that can be retained by adult mussels (see for overview Strohmeier et al. 2012). For a long time, it is was assumed that mussels do not efficiently retain smaller particles, with studies reporting on 90 % retention for 3-µm particles by Mytilus edulis, while 1-µm particles are retained with 50 % efficiency only (Møhlenberg and Riisgård 1978). Most studies were performed under controlled laboratory conditions using phytoplankton cultures. Results from experiments using natural plankton communities reported that retention efficiency might be more variable (Trottet et al. 2008; Strohmeier et al. 2012).
Mussels filter all kinds of particles from the water. Although phytoplankton was traditionally considered the main food source (Nielsen and Maar 2007), several studies have stated the importance of other food particles like dead organic material (Dame and Dankers 1988) and bacteria attached to this (Newell et al. 1989), microzooplankton (Horsted et al. 1988; Kreeger and Newell 1996; Trottet et al. 2008) and, for larger mussels (>22 mm; Horsted et al. 1988), mesozooplankton (Davenport et al. 2000; Wong and Levinton 2006; Lehane and Davenport 2006).
The aim of this study was to establish realised clearance rate of juvenile mussels (1.5–25 mm) in relation to both shell length and weight. Furthermore, clearance rates will be described for different prey items: bacteria (0.6 µm), picophytoplankton (<3 µm), nanophytoplankton (3–20 µm), and ciliates (10–200 µm). To establish the clearance rates of juvenile mussels, an experimental study was carried out for 3 years. Juvenile mussels were incubated in sea water originating from the western Wadden Sea. This study is one of the first describing grazing of dense populations of juvenile mussels in natural sea water. The results of this study can be used to estimate the effect of juvenile mussel cultures on the ecosystem of the western Wadden Sea.
Materials and methods
In order to measure the clearance rates of juvenile mussels and explore the planktonic prey items removed, an experimental study was carried out between 2010 and 2012. Clearance rates of juvenile mussels in natural sea water were calculated. Before and after the incubation, water samples were analysed for the presence of different prey items.
Study animals
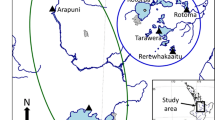
Each year, a small collector was placed in the Marsdiep (52°58′N, 4°49′E, Fig. 1). This collector consisted of filamentous ropes facilitating mussel settlement (Xmas tree ropes, Donaghys). After settlement around June, mussels increase in size up to approximately 25 mm when harvested in October. Mussel sizes used in this study were between 1.5 and 25 mm. The day before each incubation experiment, ropes with juvenile mussels were collected, transported in sea water, and stored at 4 °C. At the day of the experiment, mussels were acclimatised to ambient sea water temperature and pre-incubated.

Locations for the collector and experimental site (NIOZ harbour) in the Dutch Wadden Sea
After each experiment, the number of mussels used, average length (±0.01 mm), and dry weight (dried at 60 °C for 48 h, ±0.1 mg) were recorded. Weight included both shell and flesh. In 2012, separate tissue dry weights were determined for an additional series of mussels (7.5–20 mm). The relationship between total dry weight and tissue dry weight was used to construct the relation of clearance rate depending on tissue dry weight in 2012, allowing for a comparison with results reported in other studies.
Experimental set-up
Two types of experiments were designed. In 2010 and 2011, pieces of mussel ropes were incubated in mesocosms to calculate the clearance rate of a mussel community. These mussel assemblages on a rope consist of different-sized mussels, resulting in a relatively high variation in shell lengths (Table 1). In 2012, laboratory experiments were performed; in this set-up, the variation in shell length was greatly reduced by removing mussels from a piece of rope, measuring them, and sorting them by size. Clearance rates of these equally sized mussels were measured in smaller volumes (Table 1).
Mesocosm experiments
To measure the clearance rate of a population of juvenile mussels, pieces of rope were incubated in mesocosms (60–85 L) in 2010 and 2011. On each experimental date (Table 1), 4 or 5 mesocosms were filled with natural sea water by suspension and placed in the NIOZ harbour (Fig. 1). Both before and after the experiment, complete mixture of the water was checked by comparing the readings of the fluorescence probe (microFlu, TriOS) at different depths. 2 or 3 mesocosms were incubated with mussels, two served as control. Mussel ropes were placed in the mesocosm, and a rotator enabled gentle mixing of the water to avoid damage of the fragile microzooplankton community. The removal rate of phytoplankton biomass was monitored using a fluorescence probe. Experiments lasted 1–4 h and were terminated before plankton depletion was expected to have occurred. This assumption was checked at the end of each experiment by verifying the linearity of ln (fluorescence signal) over time.
Laboratory experiments
Mussels were gently removed from a piece of rope, measured, and sorted by size. 1–100 equally sized mussels (Table 1) were placed loosely in petticoat netting (0.5 × 0.5 cm mesh size). For each experiment, two glass beakers were filled with natural sea water (0.1–1 L). To one beaker, mussels were added, one beaker served as control. Water was gently stirred, and phytoplankton numbers at different depths were compared by means of flow cytometry to check for complete mixture of the water. Phytoplankton cell numbers were monitored throughout the experiment, and linearity of the natural logarithm of cell concentration over time was checked afterwards, to verify the absence of depletion. The experiments lasted between 0.75–1.5 h. On several occasions, mussels were reused again, repeating the experiment using a new water sample (Table 1).
Prey items
Bacteria
Triplicate subsamples (1 mL) for enumerating free-living bacteria were fixed with glutaraldehyde (0.5 % final concentration), mixed, and then stored at −80 °C until analysis. Analysis was always within 1 month.
Analyses were performed using a flow cytometer (C6, BD Accuri, excitation with 488 nm laser), and samples were diluted with 10 % TE buffer to lower the count rate below 3,500 events s−1, the maximum recording rate of the instrument. SYBR green I (Invitrogen) stain was added (fc 0.1 %), and samples were incubated in the dark for 15 min. The 530 nm laser (FL1) was used to detect the stained cells.
Pico- and nanophytoplankton
Phytoplankton cell counts were obtained by means of flow cytometry. Water subsamples (1 mL) in triplicates were processed freshly, immediately after collection. Fluorescence at wavelengths >670 nm (FL3) was ascribed to chlorophyll a. Forward scatter was used as an indication of cell size (e.g. Li 1995). Based on the relative fluorescence to size, a distinction between phytoplankton and debris was made. Phytoplankton cell counts were further divided into two size classes (<3 µm: pico and 3–20 µm: nano) using 3-µm beads (Polyscience). A minimum cell count of 1,000 per size class was applied. Within the picophytoplankton, two distinct groups could be identified: those with the pigment phycoerythrin (FL2: 585 nm) (‘picocyanobacteria’) and those without this pigment (‘others’).
To calculate an average size per prey item measured with the flow cytometer, additional beads (7–10 µm) were used to calibrate forward scatter with size.
Ciliates
For enumeration of ciliates, one subsample (0.5–1 L) was fixed in 4 mL acid Lugol and stored in brown glass bottles at 4 °C until analysis. Samples were concentrated (10–20×), and per sample, a minimum of 100 individuals were counted or, at very low abundances, all individuals in a maximum of 10 % of the concentrated sample. Ciliate cells were counted and divided into five size classes (<20, 20–40, 40–60, 60–80, and >80 µm) with an inverted microscope using the Utermöhl sedimentation technique (Verweij et al. 2010).
Calculation of clearance rates
Clearance rates (R C) for each parameter of interest were calculated following the equation (Coughlan 1969):
where V is the volume (L) cleared, t is the duration of the measurement (h), n is the number of mussels used in the experiment, C 0 is the concentration of a particulate parameter at the start of an experiment, and C t is the concentration at the end. C 0′ and C t ′ are the concentrations at the start and end, respectively, in the control. R C was expressed as litre per hour per individual mussel. At the end of each experiment, linearity of ln(C 0/C t ) was verified. This ‘clearance rate’ method is considered reliable when the above condition is met (Riisgård 2001).
Statistical analysis
To describe clearance rate as a function of either shell length or weight, the removal rate of nanophytoplankton cells was used. For this functional group, with an average size of 6.6 µm, 100 % efficient retention was assumed. The theoretical relationship between clearance rate and shell length or weight can be described by the following equations, for length
in which R C is the clearance rate in litres per hour and L the shell length in mm. For weight, the equation is given by
in which R C is the clearance rate (L h−1) and W is either the total dry weight (shell and tissue, 2010 and 2011) in grams or dry tissue weight (g) (2012).
Under the null hypothesis, that clearance rate scales with length to an exponent b = 2. The exponent for weight d is expected to be 2/3 (Jones et al. 1992).
To test the potential difference between years for the relationship between clearance rate and either length or weight, linear models of log10-transformed data were used (models 1–3). The same kind of models was used to test whether the coefficients b and d differed from their expected values, i.e. 2 and 2/3, respectively (model 4). model 1: \({ \log }\,R_{{{\text C}ij}} = { \log }\,a + b\,{ \log }\,x_{ij} + \varepsilon_{ij}\) (common slope and intercept for all years) model 2: \({ \log }\,R_{{{\text C}ij}} = { \log }\,a_{j} + b\,{ \log }\,x_{ij} + \varepsilon_{ij}\) (common slope for all years only) model 3: \({ \log }\,R_{{{\text C}ij}} = { \log }\,a_{j} + \, b_{j} \,{ \log }\,x_{ij} + \varepsilon_{ij}\) (slope and intercept differ between years) model 4: \(\log R_{{{\text C}ij}} = { \log }\,a + \, 2{ \log }\,x_{ij} + \varepsilon_{ij} \;or\;{ \log }\,R_{{\text{C}i}} = { \log }\,a + { \log }\,x_{ij} + \varepsilon_{ij}\) (slope equal to 2 or to 2/3, common intercept for all years) R C is the clearance rate, a is the intercept, b the slope, and ε the error term. The indices i and j refer to observation i in year j.
To quantify the clearance rate of picophytoplankton and bacteria relative to the clearance rate of nanophytoplankton, linear regression was applied using the individual clearance rates measured.
To test whether the clearance rate of juvenile mussels on nanophytoplankton differed from the clearance rate on ciliates, the individual rates were compared using a paired t test.
All data were analysed using R version 2.14.1 [(C) 2011, The R Foundation for Statistical Computing].
A significance level of α < 0.05 was used for all tests.
Results
Clearance rate of juvenile mussels depending on length and weight
The clearance rate of mussels depending on mussel shell length
There was no significant interaction of the factor year with the relationship between clearance rate and length (F 4,48 = 1.42, p = 0.24, models 1 and 3). Neither did the intercepts of this relation differ between the 3 years (F 2,50 = 2.88, p = 0.07 models 1 and 2).
The common slope, grouping the measurements of all 3 years together, did not differ from the theoretically expected value of 2 for b (F 1,52 = 2.25, p = 0.14, models 1 and 4). Using this fixed value for b, the intercept was estimated at (0.0004) (Fig. 2) with no significant differences between the 3 years (F 2,51 = 2.20, p = 0.12).

Clearance rate on nanophytoplankton cells, measured for three consecutive seasons for mussels varying in mean size from 1.5 to 25 mm. The clearance rate is expressed as the litres of water cleared of cells per hour per individual mussel. There were no significant differences in either the slope or the intercept between the 3 years (models 1–3). The data from the 3 years were combined, and it was further tested whether the regression coefficient different significantly from the expected value of 2 (model 4). The regression coefficient did not differ significantly from the expected value, and one regression line was fitted using a slope of two (black line) (log R C = log (−3.41 ± 0.04) + 2 log Length). The small insert at the left shows the clearance rates of the smallest mussels only (<10 mm). Both axes are on log scale
The clearance rate of mussels depending on mussel weight
The individual clearance rate of juvenile mussels can also be described in relation to the weight of a mussel according to R C = cWd. Weight here is defined as the weight of shell and tissue together (Fig. 3a).

a Clearance rate on nanophytoplankton cells, measured for mussels varying in mean size from 1.5 to 25 mm (corresponding to 0.5–700 mg DW of shell and tissue) for 3 years. The clearance rate is expressed as the litres of water cleared of cells per hour per individual mussel. Both axes are on log scale. b The clearance rate on nanophytoplankton cells as a function of the mean individual mussel tissue dry weight. The data were collected in 2012. The regression coefficient did not differ significantly from the expected value of two-thirds. Therefore, a regression line was fitted using a slope of two (black line). The relationship between clearance rate (L h−1) and tissue dry weight (g) is best described by the equation log R C = log (−0.13 ± 0.06) + 0.67 log W. Both axes are on log scale
The relation of clearance rate with mussel dry weight was not the same for each year (F 4,48 = 8.61, p = 2.547e−05, models 1 and 3). The intercepts differed between the 3 years (F 2,50 = 14.72, p = 9.43e−06, models 1 and 2), not the slope (F 4,48 = 1.94, p = 0.15, models 2 and 3).
Whether the slope differed from the expected value for d = 0.67 was tested for each year separately. Only for 2010, the model with a fixed b of 0.67 differed significantly from the estimated d based on the data (2010: F 1,13 = 5.18, p = 0.04, 2011: F 1,8 = 0.32, p = 0.59, 2012: F 1,27 = 0.04, p = 0.85). The intercepts for 2011 and 2012 are different (F 2,51 = 6.01, p = 0.005), so the best fitted lines are given for each year separately (Table 2).
To compare the results on the relationship between clearance rates and weight in the current study with results reported in the previous studies, the relationship between clearance rate and tissue dry weight was established (Fig. 3b). Only for 2012, tissue and shell dry weights were measured separately (methods 2.1). The relationship between tissue dry weight (W, g) and shell length (L, mm) can be described by the relation W = 1.7 × 10−5 L2.7 (r 2 = 0.98).
In 2012, the relationship between clearance rate and tissue dry weight did not differ from the expected value of 0.67 (F 1,27 = 0.02, p = 0.90). Clearance rate depends on tissue dry weight according to log R C = −0.13 ± 0.06 + 0.67 log W.
Clearance rate of juvenile mussels on different prey items
The R C of juvenile mussels on bacteria is on average 9 % of the clearance rate on the better retained nanophytoplankton cells (Fig. 4a).

Clearance rate (R C , L h−1) of juvenile mussels on bacteria (a, top left), picophytoplankton (b, top right), and ciliates (c, bottom left) relative to the clearance rate on the nanophytoplankton fraction. The clearance rate on bacteria, picophytoplankton, and ciliates was assumed to be proportional to the clearance rate on nanophytoplankton (e.g. R C bact. = a R C nano). The proportionality coefficient a was estimated by the antilog of the mean log ratio of R C bact, pico and ciliates and R C nano. The black dashed line (a, b) indicates the estimate for a (all years together) (bacteria: a = 0.09, R = 0.75, n = 28, picophytoplankton: a = 0.5, R = 0.95, n = 35, for mussels smaller than 10 mm. For ciliates, there was no significant difference in clearance rate compared to the clearance rate on nanophytoplankton. (y = x). For reasons of clarity, the lines y = x, y = 0.1x, and y = 0.01x are also indicated
Picophytoplankton is cleared from the water on average at half the rate of the nanophytoplankton cells (Fig. 4b). Based on both the auto fluorescence of chlorophyll and phycoerythrin, two groups of picophytoplankton could be distinguished: ‘others’ and ‘picocyanobacteria’. The average size of picophytoplankton was 0.7 µm for ‘picocyanobacteria’ and 1.2 µm for ‘others’. There was no difference in the clearance rates of juvenile mussels between the two groups of picophytoplankton (data not shown).
There was no significant difference between the clearance rate of juvenile mussels on nanophytoplankton and ciliates (t = 0.77, df = 17, p value = 0.45) (Fig. 4c).
Discussion
Clearance rate in relation to mussel shell length and weight
There are many studies reporting on clearance rates of mussels. Most of these studies were performed under controlled laboratory conditions, using cultured algal species, while other, more recent studies established clearance rates under natural conditions. There are large differences in the clearance rates reported, and there has been much debate about the causes for these differences. The main arguments to explain the differences between studies are the use of different methodologies (Riisgård 2001; Riisgård et al. 2014), differences in mussel condition index (Filgueira et al. 2008; Riisgård et al. 2014) or food type, with lower clearance rates measured when natural plankton is used (Doering and Oviatt 1986). Nowadays, there seems to be consensus on the concept of considering filtration rates determined in controlled laboratory experiments using cultured algal species and low mussel densities as maximum rates, while clearance rates established under field conditions can be regarded as realised clearance rates (Cranford et al. 2011; Riisgård et al. 2014).
In the current study, clearance rates were among the lowest reported (Table 3). Although during the experiments complete mixing of the water was aimed for and no gradient of phytoplankton concentration in the experimental units was measured, depletion of algal cells close to an individual mussel cannot be excluded; especially, since in the current study, large numbers of closely packed mussels were used in the experiments. Local depletion of food can result in re-filtration of the water. Re-filtration of water might thus provide an additional explanation for the low clearance rates measured in the current study. But it seems that re-filtration was not a constant factor. In 2012, for the smallest mussels, clearance rates were comparable to rates determined in controlled laboratory experiments on small post-metamorphosed larvae (Riisgård et al. 1980). With increasing mussel weight and concentration (Table 1), the difference got larger and it seems that the influence of re-filtration on the clearance rate becomes more importance (Fig. 5).
There is thus a difference in the scaling relationship between clearance rate and weight between the current study and the study performed by Riisgård et al. (1980). While in the current study clearance rate scaled with an exponent of two-thirds over the entire size range. Riisgård et al. found that clearance rate scaled with weight1 for small mussels (tissue dry weight <10 mg), decreasing to two-thirds with increasing weight (Fig. 5).
The difference in scaling exponent between the current study and the study by Riisgård et al. is not easy disclosed, but might be due to differences either in morphology or in condition. Clearance rates scaling with weight1 could also indicate that gill area does not scale with length2, representing ‘high non-isometric growth’ of the gills (Riisgård et al. 1980). Unfortunately, no data are available on the relationship between gill area and length, nor on the relationship between clearance rate and length. In the current study, weight scaled with length3 and clearance rate scaled with length2, making a high isometric scaling of the gills unlikely. However, due to the relative large variation between measurements in our study, we cannot rule out that for the maximum clearance rate and the relation with weight might be best described by weight1.
Clearance rate of juvenile mussels on different-sized prey items
The average diameter of bacteria in the current study was 0.6 µm. Bacteria were cleared from the water with an average efficiency of 9 % (Fig. 4a) of the clearance rate on nanophytoplankton, the most effectively cleared prey item (Fig. 6). This is somewhat higher than efficiencies reported in other studies. Trottet et al. (2008), using natural sea water, found clearance rates of adult mussels on bacteria to be close to zero. Nielsen and Maar (2007) found no removal of bacteria above a mussel bed. (Fig. 5).

Relationship between clearance rate (L h−1) and tissue dry weight (mg) of mussels. Both axes are on a log scale. b in the relation to log R C = log a + b log W was reported to be 1 in Riisgård et al. (1980) (∆), 2/3 in Riisgård and Møhlenberg (1979)/ Møhlenberg and Riisgård (1979) (+), and 2/3 for the current study (2012, o)

Summarising boxplot indicating the clearance rate of juvenile mussels on four prey items for all years together. Clearance rate is expressed as litre cleared of items per hour per mm2 shell length, to make the R C independent of shell length
The clearance rate on picophytoplankton was higher than the average clearance rate on bacteria (Fig. 6). The clearance rate on the picofraction of phytoplankton occurred on average at half the rate of the clearance on larger nanophytoplankton (Fig. 4b). The diameter of picophytoplankton was between 0.7 and 1.0 µm, and the retention efficiencies found in the current study fall within the range of reported efficiencies for 1 µm (unidentified) particles (e.g. 50 %: Møhlenberg and Riisgård 1978; 20 %: Riisgård et al. 1980; 14–64 %: Strohmeier et al. 2012).
The difference in diameter between bacteria (0.6 µm) and picophytoplankton (0.7–1.0 µm) is small, while the average retention is much higher for picophytoplankton compared to bacteria. This sharp decline in retention efficiency with decreasing particle size has been reported before (Lucas et al. 1987; Matthews et al. 1989; Ward and Shumway 2004). Preferential capture of picophytoplankton over bacteria must be based on properties other than cell size alone. Differences in stickiness between species of the same size, affecting capture efficiency by the ctenidium, has been suggested as a possible explanation for the variation in retention of equally sized particles (Ward and Shumway 2004).
The average diameter of nanophytoplankton cells was 6.6 µm, while ciliate were much larger, ranging in diameter roughly between 10 and 200 µm, with a weighted average of 28.6 µm (±7.9). Clearance rates on ciliates however were comparable to the clearance rates on nanophytoplankton (Figs. 4c, 6). Optimal retention thus reaches a plateau for particles larger than 6.6 µm in this study.
Variable retention
The retention efficiency for different prey items is not constant (Fig. 4a–c). For bacteria, the retention relative to the retention of nanophytoplankton varied between 1 and 26 % and for picophytoplankton, retention varied between 11 and 64 %.
Mussels can lower the retention efficiency for small particles to some extent by widening the interfilamentary distances of the ctenidium or by shifting the movement of the latero-frontal cilia to the side, so cilia no longer block the passage of smaller particles (Atkins 1937; Dral 1967; Barillé et al. 1993; Strohmeier et al. 2012).
There is a positive relationship between the size of a particle and its nutritional value (Ward and Shumway 2004). Assuming that mussels strive to maximize their energy intake, a trade-off is expected with regard to the distance between the filaments, either a wide interfilamentary distance, creating a low concentration (since abundance is negatively related to size) of large nutritious (Ward and Shumway 2004) particles, or a more narrow distance, resulting in a high concentration of particles, but including a large quantity of low-quality particles. A higher inflow of lower-quality particles is likely to increase the processing costs (e.g. pumping, handling, selecting, and rejection). It can thus be expected that the optimal interfilamentary distance at least balances the costs of processing of different quantity and quality particles with the benefits.
There are studies reporting on higher or lower retention efficiencies in response to variations in natural seston. Strohmeier et al. (2012) found that when total cell volume was dominated by small particles, the particle size most efficiently retained decreased (to 6–16 µm). At times when total cell volume was dominated by larger cells, capture efficiency increased to larger particles (20–30 µm). Calculating the carbon per size class for data published in Lucas et al. (1987) revealed a similar pattern; retention efficiency for 1.6-µm particles differed between two sites. The highest retention efficiency for these picoparticles corresponded to relative small (8 µm) particles dominating total carbon availability, while at the site with a lower retention the carbon availability was dominated by 12- to 16-µm particles. Trottet et al (2008) investigated the clearance rates on different phytoplankton species, heterotrophic flagellates, and ciliates. Relative clearance rates between species and taxa varied throughout the year. No consistent relationship between cell abundance and clearance rate per species/taxa was found. In the current study, seston concentrations varied considerably. During the experiments, the suspended matter concentration fluctuated between 16 and 50 mg L−1 with chlorophyll a concentration between 3 and 11 µg L−1 (data not shown). Variation in retention of the different prey items could however not be related to differences in either suspended matter or chlorophyll a concentrations. Neither could this variable retention efficiency be attributed to differences in dominant cell size. Whether mussels are able to control particle retention in response to variations in natural seston concentration remains a controversial topic, and according to Riisgård et al. (2013), the mechanism of modulation of the retention efficiency ‘lacks a physical explanation’.
Conclusion
The current study is one of the first describing realised clearance rates related to length and weight for juvenile mussels. Clearance rates scaled with length2 in the same way as adult mussels do. Scaling of clearance rate with weight was more variable. Weight is not only expected to fluctuate within a year, but also between years, effecting the relation with clearance rate. In other studies, it was already concluded that gill area generally scales well with length, and that therefore clearance rate estimates based on length can be considered the actual clearance rates (e.g. Filgueira et al. 2008; Riisgård et al. 2014).
Clearance rates in the current study were performed on densely populated pieces of ropes or large numbers per water volume. This might have resulted in re-filtration of water, leading to lower clearance rates compared to maximum rates determined in studies performed under controlled laboratory experiments. Extrapolating maximum rates to estimate the clearance rate exercised by dense populations of juvenile mussels, a field situation thus leads to an overestimation. The estimation of realised clearance rates in the current study, including re-filtration of the water, better represent the filtration pressure in a natural situation.
Juvenile mussels exercise comparable clearance rates on nanophytoplankton and ciliates. And, similar to adults, juvenile mussels expressed reduced clearance rates on potential food particles with a diameter <3 µm. Size selective removal, as shown by this study, might result in relative changes in plankton groups. Information on the potential effect of size-dependent clearance rates of juvenile mussels on the pelagic food web will provide a more realistic estimate of the effect of large populations of filter feeders on the carrying capacity of an ecosystem.
References
Atkins D (1937) Memoirs: on the ciliary mechanisms and interrelationships of lamellibranchs part III: types of lamellibranch gills and their food currents. Q J Microsc Sci 2315:375–421
Barillé L, Prou J, Heral M, Bourgrier S (1993) No influence of food quality, but ration-dependent retention efficiencies in the Japanese oyster Crassostrea gigas. J Exp Mar Biol Ecol 171:91–106
Bayne BL, Widdows J (1978) The physiological ecology of two populations of Mytilus edulis L. Oecologia 37:137–162
Beiras R, Camacho AP (1994) Influence of food concentration on the physiological energetics and growth of Ostrea edulis larvae. Mar Biol 120:427–435
Bunt CM, Maclsaac HI, Sprules WG (1992) Pumping rates and projected filtering impacts of juvenile zebra mussels (Dreissena polymorpha) in western Lake Erie. Can J Fish Aquat Sci 50:1017–1022
Cadée GC, Hegeman J (1974) Primary production of the benthic microflora living on tidal flats in the Dutch Wadden Sea. Neth J Sea Res 8:260–291
Cloern JE (1982) Does the benthos control phytoplankton biomass in south San Francisco Bay. Mar Ecol Prog Ser 9:191–202
Coughlan J (1969) The estimation of filtering rate from the clearance of suspensions. Mar Biol 2:356–358
Cranford PJ, Dowd M, Grant J, Hargrave B, McGladdery S (2003) Ecosystem level effects of marine bivalve aquaculture. In: Fisheries and oceans Canada. A scientific review of the potential environmental effects of aquaculture in aquatic ecosystems Vol 1. Can Tech Rep Fish Aquat Sci 2450:51–95
Cranford PJ, Ward JE, Shumway SE (2011) Bivalve filter feeding: variability and limits of the aquaculture biofilter. In: Shumway SE (ed) Shellfish aquaculture and the environment. Wiley-Blackwell, Oxford, pp 81–124
Dame RF (1996) Ecology of marine bivalves: an ecosystem approach. CRC Press, Boca Raton
Dame RF, Dankers N (1988) Uptake and release of materials by a Wadden Sea mussel bed. J Exp Mar Biol Ecol 118:207–216
Davenport J, Smith RJJW, Packer M (2000) Mussels Mytilus edulis: significant consumers and destroyers of mesozooplankton. Mar Ecol Prog Ser 198:131–137
Doering PH, Oviatt CA (1986) Application of filtration rate models to field populations of bivalves: an assessment using experimental mesocosms. Mar Ecol Prog Ser 31:265–275
Dral ADG (1967) The movements of the latero-frontal cilia and the mechanism of particle retention in the mussel (Mytilus edulis). Neth J Sea Res 3:391–422
Filgueira R, Labarta U, Fernández-Reiriz MJ (2008) Effect of condition index on allometric relationships of clearance rate in Mytilus galloprovincialis Lamarck, 1819. Rev Biol Mar Oceanogr 43:391–398
Horsted SJ, Nielsen TG, Riemann B, Pock-Steen J, Bjørnsen PK (1988) Regulation of zooplankton by suspension-feeding bivalves and fish in estuarine enclosures. Mar Ecol Prog Ser 48:217–224
Jacobs P, Beauchemin C, Riegman R (2014) Growth of juvenile blue mussels (Mytilus edulis) on suspended collectors in the Dutch Wadden Sea. J Sea Res 85:365–371
Jones HD, Richards OG, Southern TA (1992) Gill dimensions, water pumping rate and body size in mussel Mytilus edulis L. J Exp Mar Biol Ecol 155:213–237
Kiørboe T, Møhlenberg F (1981) Particle selection in suspension-feeding bivalves. Mar Ecol Prog Ser 5:291–296
Kreeger DA, Newell RIE (1996) Ingestion and assimilation of carbon from cellulolytic bacteria and heterotrophic flagellates by the mussels Geukensia demissa and Mytilus edulis (Bivalvia, Mollusca). Aquat Microb Ecol 11:205–214
Lehane C, Davenport J (2006) A 15-month study of zooplankton ingestion by farmed mussels (Mytilus edulis) in Bantry Bay, Southwest Ireland. Estuar Coast Shelf Sci 67:645–652
Li WKW (1995) Composition of ultraphytoplankton in the central North Atlantic. Mar Ecol Prog Ser 122:1–8
Lucas MI, Newell RC, Shumway SE, Seiderer LJ, Bally R (1987) Particle clearance and yield in relation to bacterioplankton and suspended particulate availability in estuarine and open coast populations of the mussel Mytilus edulis. Mar Ecol Prog Ser 36:215–224
Matthews S, Lucas MI, Stenton-Dozey JME, Brown AC (1989) Clearance and yield of bacterioplankton and particulates for two suspension-feeding infaunal bivalves, Donax serra Röding and Mactra lilacea Lam. J Exp Mar Biol Ecol 125:219–234
Møhlenberg F, Riisgård HU (1978) Efficiency of particle retention in 13 species of suspension feeding bivalves. Ophelia 17:239–246
Møhlenberg F, Riisgård HU (1979) Filtration rate, using a new indirect technique, in thirteen species of suspension-feeding bivalves. Mar Biol 54:143–147
Newell CR, Shumway SE (1993) Grazing of natural particulates by bivalve molluscs: a spatial and temporal perspective. In: Dame RF (ed) Bivalve filter feeders in estuarine and coastal ecosystem processes NATO ASI Series, vol G 33. Springer, Berlin, pp 85–148
Newell CR, Shumway SE, Cucci TL, Selvin R (1989) The effects of natural seston particle size and type on feeding rates, feeding selectivity and food resource availability for the mussel Mytilus edulis Linnaeus, 1,758 at bottom culture sites in Maine. J Shellfish Res 8:187–196
Nielsen TG, Maar M (2007) Effects of a blue mussel Mytilus edulis bed on vertical distribution and composition of the pelagic food web. Mar Ecol Prog Ser 339:185–198
Riisgård HU (2001) On measurement of filtration rates in bivalves–the stony road to reliable data: review and interpretation. Mar Ecol Prog Ser 211:275–291
Riisgård HU, Møhlenberg F (1979) An improved automatic recording apparatus for determining the filtration rate of Mytilus edulis as a function of size and algal concentration. Mar Biol 52:61–67
Riisgård HU, Randløv A, Kristensen PS (1980) Rates of water processing, oxygen consumption and efficiency of particle retention in veligers and young post-metamorphic Mytilus edulis. Ophelia 19:37–46
Riisgård HU, Pleissner D, Lundgreen K, Larsen PS (2013) Growth of mussels Mytilus edulis at algal (Rhodomonas salina) concentrations below and above saturation levels for reduced filtration rate. Mar Biol Res 9:1005–1017
Riisgård HU, Larsen PS, Pleissner D (2014) Allometric equations for maximum filtration rate in blue mussels Mytilus edulis and importance of condition index. Helgol Mar Res 68:193–198
Smaal AC, Vonck APMA, Bakker M (1997) Seasonal variation in physiological energetics of Mytilus edulis and Cerastoderma edule of different size classes. J Mar Biol Assoc UK 77:817–838
Strohmeier T, Strand Ø, Alunno-Bruscia M, Duinker A, Cranford PJ (2012) Variability in particle retention efficiency by the mussel Mytilus edulis. J Exp Mar Biol Ecol 412:96–102
Trottet A, Roy S, Tamigneaux E, Lovejoy C, Tremblay R (2008) Impact of suspended mussels (Mytilus edulis L.) on plankton communities in a Magdalen Islands lagoon (Québec, Canada): a mesocosm approach. J Exp Mar Biol Ecol 365:103–115
Verweij GL, van Wezel RM, van den Oever A, Fockens K, Mulderij G (2010) Biomonitoring van microzoöplankton in de Nederlandse zoute wateren 2009. Rapport 2010–006. Bureau Koeman & Bijkerk, Haren
Verwey J (1952) On the ecology of distribution of cockle and mussel in the Dutch Wadden Sea, their role in sedimentation and the source of their food supply. Arch Neerl Zool 10:171–239
Ward JE, Shumway SE (2004) Separating the grain from the chaff: particle selection in suspension- and deposit-feeding bivalves. J Exp Mar Biol Ecol 300:83–130
Widdows J (1978) Combined effects of body size, food concentration and season on the physiology of Mytilus edulis. J Mar Biol Assoc UK 58:109–124
Wong WH, Levinton JS (2006) The trophic linkage between zooplankton and benthic suspension feeders: direct evidence from analyses of bivalve faecal pellets. Mar Biol 148:799–805
Acknowledgments
This study was supported by the Ministry of Economic Affairs through the MZI project. The authors would like to thank Piet-Wim van Leeuwen, André Meijboom, Pepijn de Vries, and Catherine Beauchemin for their help in collecting mussels, conducting experiments and analysis of the samples; Ecological consultancy Koeman and Bijkerk bv and Alex Blin for microzooplankton counts; Elze Dijkman for constructing Fig. 1; Bert Brinkman and Santiago Alvarez Fernandez for valuable discussions during the writing of this manuscript. Comments made on earlier versions of this manuscript by Pauline Kamermans and 2 anonymous reviewers greatly improved this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.-D. Franke.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article

Cite this article
Jacobs, P., Troost, K., Riegman, R. et al. Length- and weight-dependent clearance rates of juvenile mussels (Mytilus edulis) on various planktonic prey items. Helgol Mar Res 69, 101–112 (2015). https://doi.org/10.1007/s10152-014-0419-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-014-0419-y