
Abstract
The distribution patterns of macrobenthic invertebrate assemblages at different spatial scales and in relation to natural and anthropogenic disturbance gradients were investigated in the Pialassa Baiona, a eutrophic and polluted brackish coastal lagoon located along the Italian coast of the northern Adriatic Sea. This coastal lagoon shows a complex physiography with several shallow water ponds and channels separated by discontinuous artificial embankments. Environmental variables and macrobenthic invertebrate assemblages showed higher heterogeneity at small spatial scale (i.e. within channels and ponds). Distinction between channels and ponds is only weakly supported by the distribution patterns of macrobenthic assemblages. Depth was the major factor in structuring benthic communities within ponds, while species distributions along the channels were strongly correlated with the anthropogenic disturbance gradient. Anthropogenic disturbance mainly affected species richness, which was inversely correlated with the organic carbon contents in the sediments and the water surface temperature, which is affected by the input of cooling water from two thermal power plants. Some opportunistic species, like the polychaetes Streblospio shrubsolii and Capitella capitata, were more abundant in the southern polluted areas. In particular, the abundance of S. shrubsolii significantly increased with organic carbon contents in the sediments and water surface temperature, while C. capitata was more abundant in the warmed sites and close to sewages. Conversely, the abundance of the amphipod Corophium insidiosum was inversely related to organic carbon contents.
Similar content being viewed by others

Introduction
Temperate coastal lagoons are dynamic and unpredictable transitional water systems characterised by sharp daily and seasonal variations of the chemico-physical variables (Fiala 1972; Barnes 1991; Pérez-Ruzafa et al. 2005; Specchiulli et al. 2008). These habitats are very heterogeneous in physiography and hydrology (e.g. in extension, water depth, water exchange with the sea, salinity range), which are usually considered key features explaining a relevant proportion of their environmental variability (Basset et al. 2006). These habitats are selective towards tolerant species, and patterns of species distribution are related to the synergetic interactions of the abiotic and biotic factors (Lardicci et al. 1997; Arvanitidis et al. 1999). In coastal lagoons, species distribution is affected by heterogeneity of the environmental variable, such as water depth, hydrodynamic conditions, sediment characteristics, as well as by sources of anthropogenic disturbance, which could vary at a range of spatial scales, particularly in habitats with complex physiography due to presence of channels, shallow water ponds and embankments (Lardicci et al. 1997; Arvanitidis et al. 1999). Furthermore, anthropogenic disturbance could interact with natural heterogeneity affecting patterns of species distribution, species diversity and ecosystem functioning. In quality assessment and in analysing human impacts in transitional waters, variability at small spatial scales and natural vs. anthropogenic disturbance gradient need to be carefully investigated (Cao et al. 1996; Weis et al. 2004; Ponti et al. 2008).
The Italian northern Adriatic coastal area mainly consists of lagoon–river delta systems, hosting a large number of transitional water bodies. These habitats range for the largest and most studied Lagoon of Venice, to wetlands, estuaries, embayment, ponds, differing in their extension and connection with the sea. The increasing pressures (e.g. eutrophication, chemical and thermal pollution, over-harvesting, dredging and habitat loss) inevitably lead to degradation of these ecosystems (Abbiati et al. 2010). The aim of present study was to investigate the distribution patterns of macrobenthic assemblages in the Pialassa Baiona, a northern Adriatic brackish lagoon, characterised by a complex network of channels connecting shallow water ponds and by several sources of industrial and urban pollution. The possible effects of natural (e.g. depth, grain size, distance from the sea) and anthropogenic disturbance gradients (e.g. sediment organic content, Hg contamination, water temperatures affected by thermal pollution, distance from sewage inputs) on the distribution patterns of macrobenthic assemblages were investigate using a hierarchical sampling design, which allow to analyse patterns of spatial variability of species assemblages at scales of sites (hundreds of metres apart), areas (separated by discontinuous embankments), and two putative habitats: channel and pond.
Materials and methods
Study area
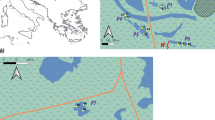
Pialassa Baiona is a eutrophic micro-tidal lagoon located along the northern Adriatic coast of Italy (Fig. 1). This area was declared wetland of international importance under the Ramsar Convention in 1981 and is included among the sites of Community interest (pSCIs, “Habitat” Directive 92/43/EEC). Nowadays, the lagoon is exploited for recreational fishing, Manila clams (Ruditapes philippinarum) harvesting, regulated hunting, leisure and cultural activities such as canoeing, guided walking tours and bird watching.

Map of Pialassa Baiona lagoon, showing sampling sites located in channels (circles) and ponds (triangles). Legend: VEN: Vena del Largo pond; POL: Pola Longa pond; RIS: Risega pond; MAG: Magni pond; BAC: Baccarini channel; TBF: Taglio della Baiona-Fossatone channel; BAI: Baiona channel; STA: Magni-Staggi channel (geographic grid UTM 32T, European Datum 1950)
The lagoon was originated in the eighteenth century. Nowadays, discontinuous artificial embankments divide the lagoon into several shallow water ponds with an average depth of 1 m. Main ponds are connected each other and to the sea by channels. In channels, the depth ranges from 1 m to 8 m in the landward and seaward sides, respectively. On average, the water covers an area of 10 km2, tidal range can exceptionally exceed 1 m, and usually vast shallow areas emerge during low tides. The lagoon receives water inputs from five main channels that drain a watershed of 264 km2, including urban (9%) and agricultural (87%) areas. The southern channel collects also the wastewater coming from urban and industrial sewage treatment plants and from two thermal power plants. Nutrient inputs are mainly located in the southern area (Ponti et al. 2005). Overall water turnover in the lagoon has been estimated in 3 days. Phytoplankton blooms and intense growth of seaweeds (Ulva sp., Enteromorpha sp., Gracilaria sp.) were frequently observed during the summer, especially in the southern part. Dystrophic crises were often recorded in the summer (Ponti and Abbiati 2004).
From 1958 to 1976, Pialassa Baiona was heavily impacted by industrial pollution. Mercury, polycyclic aromatic hydrocarbons (PAHs), and synthetic polymers were among the most important pollutants which nowadays contaminate the sedimentary compartment (Fabbri et al. 1998, 2000; McRae et al. 2000; Abbondanzi et al. 2005). Although the release of mercury ceased in the middle 1970s, superficial sediment are still contaminated, with a total mercury load ranging from 0.13 to 250 μg g−1 dry weight (Trombini et al. 2003). The pollution gradient decreases from south to north, in relation to the distance from the southern industrial area (Trombini et al. 2003). Previous studies on the Pialassa Baiona showed a correlation between total mercury concentrations in sediments and the overall level of contamination due to other pollutants, such as PAHs and synthetic polymers (Fabbri et al. 2000, 2001b). These results were ascribed both to the common history of the inputs of contaminants and to the transport processes of pollutants across the Pialassa Baiona lagoon.
Pialassa Baiona, as well as most temperate coastal lagoons, is a heterogeneous environment, where physical and biological variables change at different spatial and temporal scale. Thanks to the relatively small size and to the complex patterns of natural and anthropogenic stressors, it provides a natural mesocosm to test theories on models of species distribution in TW habitats.
Sampling and laboratory analyses
In the Pialassa Baiona lagoon, ponds and channels represent different putative habitats. The channels represent about 23% of the total water surface and contain almost half of the total water volume of the lagoon (Ponti et al. 2005). Within each habitat, macrobenthic assemblages were investigated at two spatial scales: areas, separated by discontinuous embankments, and sites, within each area, hundreds of meter apart. Four channel areas (BAC: ‘Baccarini’; TBF: ‘Taglio della Baiona-Fossatone’; BAI: ‘Baiona’; STA: ‘Magni-Staggi’) and four pond areas (VEN: ‘Vena del Largo’; POL: ‘Pola Longa’; RIS: ‘Risega’; MAG: ‘Magni’) were randomly chosen (Fig. 1). Five sampling sites were haphazardly located in each area (channels and ponds), and two replicate samples were collected at each site by means of a Wildco® box corer with a sampling area of 0.0225 m2. Samples were collected in June 2002. After a pre-sieving (0.5 mm mesh) in the field, samples were preserved using a buffered solution of 4% formaldehyde. Specimens were identified to the lowest possible taxonomic level, using a binocular microscope, and counted.
Several environmental variables were measured at each site, some of them related to the morphology and sedimentology of the lagoon (e.g. depth, grain size, pore water salinity, water flow speeds, distance from the sea), while the other are linked to anthropogenic disturbance (e.g. sediment organic content, Hg contamination, water temperatures affected by thermal power plants, distance from sewage inputs).
Water depths were measured by a metric tape and then referred to the mean lower low water (MLLW). A sample of superficial sediment (0–5 cm horizon) was collected at each site using a hand corer sampler to analyse sand and silt–clay percentage, organic carbon content, pore water salinity and mercury contamination. The apparent Redox Potential Discontinuity (RPD) was measured as depth of boundary between the light aerobic near-surface sediment and the underlying black hypoxic or anoxic sediment. Even though RPD is a qualitative measure of the redox status of the sediment, which could be little affected by the “wall effect” caused by core penetration into the sediment, it can be used as a proxy of the oxic/anoxic status of the sediments (Bona et al. 2000). Sand and silt–clay sediment content was measured as dry weight percentage after wet sieving (0.063 mm mesh), retaining the fine fraction by filter paper (particle retention 11 μm), and drying at 90°C for 24 h. Sediment organic content was determined as percentage loss of weight on ignition (LOI) at 500°C for 8 h after drying at 90°C for 24 h (King et al. 1998). Salinity of the pore water (Spw) was measured with a hand refractometer after the water extraction by a syringe and glass microfibre filters (grade GF/F, particle retention 0.7 μm).
Total mercury in the sediment (Hg) was analysed by Cold Vapour Atomic Absorption Spectrometry (CVAAS) after digestion with nitric and sulphuric acid (as in Fabbri et al. 2001a). Reagent blanks and certified reference materials, PACS-2, was analysed concurrently with sediment samples to validate the method used. Concentrations are expressed on a dry weight basis.
The water temperature changes rapidly during the day in function of solar radiation and tidal flow; therefore, synoptic measures of the surface temperature are needed to provide a reliable spatial pattern. Water surface temperatures (Ts) were measured during a summertime survey, carried out covering most of the lagoon, using a combined temperature and DGPS position recorder, in less then 2 h at the low tide stand. This survey provided a high spatial resolution and reliable measure of the thermal pollution gradient within the lagoon; even though these measures appeared as a snapshot, the results in term of ΔT were in well accordance with those carried out by six digital thermometers permanently placed around the lagoon (data stored every 30 min for 1 year). Average water flow speeds (Fm), for the sites located in the channels, were obtained simulating a complete lunar tide cycle with a hydrodynamic model (developed by the Hydraulic Laboratory of the University of Bologna, by courtesy of Prof. A Lamberti), based on the Full Equation model (FEQ, Franz and Melching 1997).
Distances of the sampling sites from the outlet into the sea (Dc) and from the main sewage inputs (Ds), located in the south-western part, were considered as possible explanatory environmental variables related to water renewal and pollution gradient, respectively. These distances were measured along the preferential water paths on the digital cartography according to the hydrodynamic model.
Data analyses
Macrobenthic assemblages were analysed in term of species abundance. Species richness (as number of species, S), species diversity (as Hill’s diversity number N1 = Exp H′, where H′ is the Shannon’s index based on natural logarithm) and the corresponding evenness component (as N10 = N1/S) were calculated for each replicate sample (Gray 2000).
Hierarchical analysis of variance (ANOVA) was applied to single species abundance, total abundance (A) and diversity indices (S, N1 and N10) to test for significant differences among Habitats (2 levels, fixed), Areas (4 levels, random, nested in Habitats) and Sites (5 levels, random, nested in Areas). Cochran’s C test was used to check the assumption of homogeneity of variances and, when necessary, appropriate transformations were applied. If variances were slightly heterogeneous even after transformations (Cochran’s C test P < 0.05), the analyses were run at α = 0.01 for significance test, while when variances were high heterogeneous (Cochran’s C test P < 0.01) only not significant results were retained (Underwood 1997). Bonferroni correction for multiple tests was not applied.
Patterns of environmental variables between habitats and areas were analysed by ANOVA. Differences in water flow speeds were analysed only among channels. Student-Newman-Kuels (SNK) post-hoc test was used for multiple comparisons of the means (Underwood 1997).
Spatial distribution of the assemblages were analysed using principal coordinate analysis (PCO, i.e. metric multidimensional scaling) based on Bray-Curtis dissimilarities of square root-transformed data (Anderson 2003b; Anderson and Willis 2003). Differences in community structures between habitats in different areas and sites were assessed by permutational non-parametric multivariate analysis of variance (PERMANOVA) following the same experimental design adopted for ANOVA (Anderson 2001, 2005). When less then 999 unique values in the permutation distribution were available, asymptotical Monte Carlo P-values were used instead of permutational P-values.
Relationships between the similarity distribution patterns of the assemblages and the environmental variables were investigated by a multivariate multiple regression based on the Bray-Curtis dissimilarities of square root-transformed species data, averaged among sites of the same area (McArdle and Anderson 2001). Multiple regressions were performed using the ‘DISTLM forward’ procedure (Anderson 2003a) that give a forward selection of the variable that better explain the observed assemblages patterns, with tests by permutation. Possible correlations between species abundances and measured environmental variables were investigated using Pearson’s linear correlation. Linear correlation among environmental variables and between them and species diversity indices were also investigated.
Results
A total of 81 taxa of macrobenthic invertebrates were recorded, 52 were identified to species level. The distribution of taxa among the forty sites is reported in the Appendix. Mean number of taxa (S) per site (n = 2) ranged from 8.5 (±1.5 SE) to 32.5 (±0.5 SE). Species richness showed high significant differences both among sites and areas, while it did not differ significantly among habitats (Fig. 2 and Table 1). Southern channel, where the anthropogenic disturbance sources are located, showed the lowers number of species (SNK: STA ≪ BAI = BAC = TBF). No significant differences were detected among habitats, areas or sites in species diversity (N1) (Fig. 2 and Table 1). Evenness (N10) varied from 0.09 ± 0.01 to 0.66 ± 0.04 (±SE, n = 2), and significant differences were found among areas and site, but not between habitats (Fig. 2 and Table 1).

Mean species richness (number of species, S), species diversity (Hill’s N1) and evenness (Hill’s N10) per sample (+SE, n = 5) at each study area
Species distribution
The amphipod Corophium insidiosum was the most abundant taxon, followed by nematodes, Hydrobia cfr. ventrosa, Corophium acherusicum, unidentified tubificid oligochaetes, Microdeutopus gryllotalpa, Polydora ciliata, Gammarus aequicauda, unidentified Actiniaria, Capitella capitata, Chironomus salinarius, Abra segmentum, Streblospio shrubsolii, Idotea balthica, Malacoceros cfr. fuliginosus, Musculista senhousia and Amphipholis squamata. The distribution of the 36 most abundant taxa was analysed by ANOVA in order to test for differences among putative habitats, sites and areas (Table 1). Most of the species showed significant differences among sites. Hediste diversicolor and Nebalia sp. were the only two taxa that did not show difference related to both site and area, while Cirrophorus furcatus is the only species showing significant differences among areas, but not among sites. Although the great variability observed at small spatial scales, Melita palmata, Mytilus galloprovincialis and Streblospio shrubsolii were significantly more abundant in the channels, the mud snail Hydrobia cfr. ventrosa and the polychaete Malacoceros cfr. fuliginosus were more abundant in the ponds.
Patterns of the macrobenthic assemblages
The first two axes of the PCO explained 21.13 and 19.76% of the variability of macrobenthic invertebrate assemblages, respectively (Fig. 3). The scatter plot of sampling points showed some degree of separation between the assemblages inhabiting channels and ponds, as well as among some areas (Fig. 3). These patterns were confirmed by the PERMANOVA test, showing significant differences among habitats, areas and sites (Table 2). Altogether, the assemblages in the channels appeared more heterogeneous than those of the ponds. Moreover, assemblages found in the southern channel and pond (i.e. STA = ‘Magni-Staggi’ channel and MAG = ‘Magni’ pond), which are close to the sources of anthropogenic disturbance, were well separated on the left of the plot (Fig. 3).

PCO unconstrained ordination (i.e. metric MDS) based on Bray-Curtis dissimilarities of square rout-transformed macrobenthic abundance data. Channels (BAC, TBF, BAI, STA) and ponds (VEN, POL, RIS, MAG) were indicated by open and solid symbols, respectively
The correlation of PCO Axes with species abundance, superimposed on the PCO ordination (Fig. 4), showed a high number of taxa related to the central and northern channels (right upwards rows), as well to the central and northern ponds (right downwards rows), while the abundance of only few species slightly increase towards southern ‘Magni-Staggi’ channel and ‘Magni’ pond (to the left in the plot). Species related to the southern channel and pond includes the polychaetes Streblospio shrubsolii, Capitella capitata, and Phyllodoce mucosa.

Correlation of PCO Axes with species abundance, represented by vectors superimposed to the PCO plot of Fig. 3 (only the names of the most important species are indicated)
Patterns of environmental variables
Average values of the environmental variables for each area were reported in Table 3. No significant differences were found in depth between areas within ponds or channels, while the average depth in channels (−1.75 ± 0.20 m ± S.E.) was significantly greater than in ponds (−0.38 ± 0.15 m ± S.E.; Table 4).
Sand contents in the sediments were extremely variable, varying from 12.1 to 89.5% in weight. These percentages are mainly related to the interaction between present sedimentation processes and the occurrence of relict sand dunes. Although hydrodynamic conditions are expected to differ between channels and ponds, no significant differences in sand contents were found between the considered areas or habitats (Table 4).
Content of organic carbon in sediment ranged between 3.5 and 25.0%. Significant differences were observed among areas (Table 4), due to the higher organic carbon content in ‘Magni’ pond compare to ‘Risega’ and ‘Vena del Largo’ ponds. On average, no significant differences in organic carbon contents between channels and ponds were found. Although sand percentage and organic carbon contents seem to have different patterns, a weak but significant negative correlation between them was found (R = −0.4293, P = 0.006; Table 5).
The apparent Redox Potential Discontinuity (RPD) varies from 0 to 85 mm of thickness. Significant differences were found between areas (Table 4), in particular the aerobic layer was thinner in the ‘Magni’ pond compared to the other. High, but not significant, differences between channels and ponds were found (Table 4) and, on average, the aerobic layer was thinner in ponds than in channels. This result is consistent with the weak negative correlation between RPD and depth (R = -0.3517, P = 0.026; Table 5).
Salinity of pore water (Spw) varies from 24 to 40 psu, no significant differences among areas or habitats were found (Table 4). A weak correlation between Spw and RPD was observed (R = 0.3662, P = 0. 020; Table 5).
Water surface temperature (Ts), measured during the specific survey, ranged from 25 to 34°C; significant differences between areas were fond (Table 4). ‘Magni-Staggi’ channel and the ‘Magni’ and ‘Risega’ ponds were warmer than other (SNK test: STA > BAI = TBF = BAC and MAG > RIS > POL = VEN). Water surface temperature was affected by the input of cooling water from two thermal power plants located in the southern side of the lagoon. A weak correlation between organic contents (LOI) and surface temperature was also observed (R = 0.4310, P = 0.005; Table 5), but this relationship could be simply explanted by spatial association of the power plants and sewage treatment plants inputs.
Total mercury found in the superficial sediments ranges between 0.10 and 40.57 μg g−1, and significant differences between areas were found (Table 4). ‘Magni-Staggi’ channel and the ‘Magni’ and ‘Risega’ ponds resulted more polluted than other. Hg concentration in the sediments appeared significantly correlated with organic contents (LOI), apparent Redox Potential Discontinuity (RPD), salinity of pore water (Spw) and water surface temperature (Ts) (Table 5).
The distance of the sampling sites from the outlet into the sea (Dc), ranges from 400 to 4880 m; no significant differences between areas, channels and ponds were found (Table 4). Only depth and sand percentage show a significant correlation with distance from the outlet (R = 0.4410, P = 0.004 and R = 0.3236, P = 0.042, respectively; Table 5). As expected, depth tends to increase seaward, while relict sand dunes are mainly located landward. The distance of the sampling sites from the main inputs of nutrients and pollutants (Ds), located in the south-western side of the lagoon, varies from 500 to 8380 m. Sampling areas significantly differ in mean Ds values (Table 4; SNK: STA < BAI = BAC = TBF and MAG < RIS = POL = VEN), since ‘Magni-Staggi’ channel and ‘Magni’ pond were closer to the inputs of the civil and industrial sewage treatment plants than the other channels and ponds. Depth, grain size, organic contents, surface temperature and mercury concentration were significantly correlated with the distance from the main inputs of pollutants (Table 5).
Mean water flow speed (Fm), estimated by hydrodynamic model only for the channels, varies from 0.026 to 0.273 m s−1. On average, no significant differences were found between channels (Table 4). Fm values were not significantly correlated with the distance from the sea, while they are slightly correlated with depth and surface temperature (R = 0.4736, P = 0.035 and R = −0.4485, P = 0.047, respectively; Table 5).
Correlations between specie distribution, assemblage’s similarity patterns and environmental variables
Some opportunistic species, like the polychaetes Streblospio shrubsolii and Capitella capitata, were more abundant in the southern polluted areas. In particular, the abundance of S. shrubsolii significantly increased along LOI and Ts gradients (r = 0.3608, P = 0.022 and r = 0.4129, P < 0.008, respectively), while C. capitata were more abundant in the warmed sites and close to sewages (r = 0.3973, P = 0.011 and r = −0.3340, P < 0.035, respectively). Conversely, the abundance of the amphipod Corophium insidiosum, which is considered a species sensitive to pollution and thermal stress (Prato et al. 2008), was negatively correlated with LOI (r = −0.3351, P = 0.035).
Correlation analyses between species diversity indices and environmental variables revealed that species richness was inversely correlated with water surface temperatures (Ts) and sediment organic content (LOI) (r = −0.5531, P < 0.001 and r = −0.3861, P = 0.014, respectively).
The structures of macrobenthic assemblages showed significant differences between the two considered habitats; therefore, possible relationships between assemblage structure and environmental variables were investigated separately for channels and ponds. The relationship between environmental variables and patterns of similarities of the assemblages in channels was graphically represented in Fig. 5a. Assemblages found in the ‘Magni-Staggi’ channel were well separated from the others and were associated to high temperatures of the water surface, high total mercury concentration in the sediment and short distances from the main sewage treatment plants inputs. Multivariate multiple regression showed a significant relationship between Ts, Ds, Hg and Dc, considered alone, and similarity of the assemblages. Using the forward selection procedure, Ts explained the greatest proportion (19.9%) of the variability of the assemblages, followed by Dc (11.7%), Spw (8.5%) and depth (7.1%). After this selection, the information from Hg and Ds, which are strongly correlated with Ts (Table 5), did not add any further significant proportion of explained variation.

Correlation of PCO Axes with environmental variables, represented by vectors superimposed to the PCO ordination of the assemblages found a in the channels and b in the ponds (LOI: organic carbon as Loss of weight on Ignition; RPD: apparent Redox Potential Discontinuity; Spw: pore water salinity; Ts: water surface temperature; Hg: total mercury; Dc: distance from the confluence point; Ds: distance from the main sewage input; Fm: mean water flow speed)
The correlation of environmental variables with the patterns of similarities of assemblages found in the ponds was graphically represented in Fig. 5b. Multivariate multiple regression showed a significant relationship of depth, LOI, Ts, Ds, considered alone, with the assemblage structure. Using the forward selection procedure, depth explained the greatest proportion (16.5%) of the variability of the assemblages; the other environmental variables did not add any significant proportion of explained variation.
Discussion
Benthic macroinvertebrate inhabiting soft bottoms are considered good indicators of environmental health because they are relatively sedentary, and therefore they cannot avoid deteriorating water and sediments quality conditions and respond to local environmental impacts; moreover, most species have relatively long lifespans, then they over time integrate disturbance events even in habitats showing seasonal and daily large variability, like temperate coastal lagoons and estuaries (Bilyard 1987; Dauer 1993; Salas et al. 2006). Even if this study was based on only one sampling date, in June, and did not take into account natural and human-induced seasonal variability, the distribution pattern of the macrobenthic invertebrate assemblages should be considered as the results of both current environmental status and previous disturbance events.
Pialassa Baiona, as well as most of the temperate coastal lagoons, is a complex and heterogeneous environment, where physical and biological variable change at different spatial scale. Most of the environmental variables measured in the Pialassa Baiona lagoon significantly differed among areas within the lagoon. The analyses of the benthic assemblage showed a high heterogeneity throughout the whole lagoon at small spatial scale (i.e. between sites, hundreds of metres apart, within areas). The distribution of single species, species richness and evenness also showed the higher heterogeneity at small spatial scale. Altogether channels and ponds physically differed only by the depth. The distribution of only five species significantly differed in relation to these two putative habitats. Furthermore, no differences in terms of species diversity indices were found between channels and ponds. Even the multivariate analysis of the overall benthic assemblages confirmed the high heterogeneity at small spatial scale, but it has also highlighted the presence of some minor differences between channels and ponds. Assemblages appeared more heterogeneous in channels compare to ponds. In ponds, depth appeared the leading factor determining the benthic community structure, while species distributions along the channels was strongly correlated with the anthropogenic disturbance, represented by water surface temperature affected by thermal power stations, and the land–seaward gradient, in terms of distance from the outlet into the sea. Altogether pollution and land–seaward gradients appeared more important compared to the habitat type (i.e. ponds VS channels) in controlling the multivariate distribution pattern of macrobenthos. Anthropogenic disturbance gradients overlap natural gradient, here as well as in most of the transitional waters, and this could represent a confounding factor. However, the multivariate analysis and multiple regression approaches improve the discrimination capability between natural and anthropogenic stress (Gamito 2008).
Major sources of anthropogenic disturbance in this coastal lagoon are the inputs of wastewater from urban and industrial sewage treatment plants and cooling waters from power plants, all located in the southern side of the lagoon. These water inputs provide nutrient enrichment, which lead to eutrophic condition (Ponti et al. 2005), as well as thermal and chemical pollution, which mainly affect southern ponds and channels (Trombini et al. 2003). The set of environmental variables analysed provides an overall picture of the different physical and chemical factors that could affect and threaten the Pialassa Baiona lagoon. Water surface temperature follows the dispersal of the thermal plume due to the two thermal power plants located on the southern side. Contents of organic carbon and RPD, although influenced by grain size distribution, could reflect the algal biomass accumulation related to eutrophication and dystrophic events. Intensity of the dystrophic crises, which often occur in the summer, as well as their effects on the benthic macroinvertebrates are higher in the southern areas of the lagoon (Ponti and Abbiati 2004). Southern areas (Magni-Staggi channel, Magni and Risega ponds) resulted more affected by thermal and chemical pollution and eutrophication, while northern areas showed reduced water flow. Water flow and the distance from the channels confluence could influence the water renewal, oxygenation, larval supply as well as the transport, resuspension and deposition of the pollutants.
In the Pialassa Baiona lagoon, mercury was considered a good tracer for the past chemical pollution of sediments in the lagoon (Trombini et al. 2003). The average concentration of total mercury in channel and ponds exceeded the Effects Range Median (ERM) benchmark, over which negative effects to biota might be frequently expected (ERM for Hg = 0.71 μg g−1, dry weight; Long et al. 1995).
In summary, although this coastal lagoon appeared as a complex system subdivided in several ponds and channels by discontinuous artificial embankments, environmental variables and macrobenthic invertebrate assemblages showed high heterogeneity at spatial scale smaller than a priori supposed homogeneous areas. Even the distinction between two putative habitats, channels and ponds, is weakly supported by the distribution patterns of macrobenthic invertebrate assemblages. Major forcing factors structuring the assemblages appeared the anthropogenic disturbance and land–seaward gradients.
Land–seaward gradient and the associated salinity gradient and confinement degree are widely recognised as the leading natural factors in structuring the macrobenthic invertebrate assemblages in coastal lagoons and estuaries (Guélorget and Perthuisot 1992; Sacchi and Occhipinti-Ambrogi 1992; Barnes 1994; Attrill 2002; Attrill and Rundle 2002; Gray et al. 2002; Sconfietti et al. 2003; Rossi et al. 2006; Gamito 2008). Nevertheless, models based on salinity as the main variable affecting patterns of distribution and abundance of benthic organisms at the scale of hundreds of meters within coastal lagoons, like the ‘paralic domain’ (Guélorget and Perthuisot 1992), were demonstrated to be over-simplistic (De Biasi et al. 2003). In some Italian lagoons, it has also been recognised that the increasing levels of anthropogenic disturbance can led to the homogenisation of the lagoons and the reduction in the land–seaward gradient effects on benthic assemblages (Benedetti-Cecchi et al. 2001; Lardicci et al. 2001; Sconfietti et al. 2003). For instance, the inner parts of the Venice and Comacchio lagoons had lost their distinctiveness, and their invertebrate assemblages appeared impoverished and more similar to those typical of harbours (Mistri et al. 2000, 2001; Sconfietti et al. 2003). In Comacchio lagoon, the largest non-tidal lagoonal complex (about 100 km2) of the Po River deltaic area, located just 10 km northern to Pialassa Baiona lagoon, intensely exploited for extensive aquaculture over the last 50 years, only 37 soft bottom macrobenthic taxa were identified in nine sampling stations quarterly investigated for 1 year (Mistri et al. 2000). In the Pialassa Baiona, despite the high level of anthropogenic disturbance, relatively high species diversity and a clear land–seaward gradient in the composition of macrobenthic invertebrate assemblages are still well recognisable, at least along the inner channels. Even if comparison between different studies could be affected by different sampling methods and effort (e.g. sampling surface, number of replicate), the total number of soft bottom specie recorded in Pialassa Baiona (81 taxa) is greater than that of the most of the Italian transitional waters, only Venice lagoon overcome Pialassa Baiona among the northern Adriatic coastal lagoons (Munari and Mistri 2008). Pollution mainly affects the distribution of sensitive species as well as the species richness, which could alter ecological functioning. Anthropogenic disturbance tend to reduce the natural variability of the benthic assemblages by increasing small spatial scale heterogeneity and strengthening in some way the land–seaward gradients.
In the transitional water ecosystems, understanding spatial heterogeneity of macrobenthic invertebrate assemblages, as well as their distribution patterns in response to natural and anthropogenic disturbance gradients, is of paramount importance, because this knowledge forms the basis for any management and monitoring program as well as for the environmental quality or impact assessment projects.
References
Abbiati M, Mistri M, Bartoli M, Ceccherelli VU, Colangelo MA, Ferrari CR, Giordani G, Munari C, Nizzoli D, Ponti M, Rossi R, Viaroli P (2010) Tradeoff between conservation and exploitation of the transitional water ecosystems of the northern Adriatic Sea. Chem Ecol 26. doi:10.1080/02757541003693193
Abbondanzi F, Campisi T, Focanti M, Guerra R, Iacondini A (2005) Assessing degradation capability of aerobic indigenous microflora in PAH-contaminated brackish sediments. Mar Environ Res 59:419–434
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecol 26:32–46
Anderson MJ (2003a) DISTLM forward: a FORTRAN computer program to calculate a distance-based multivariate analysis for a linear model using forward selection. Department of Statistics, University of Auckland, Auckland. Available at: www.stat.auckland.ac.nz/~mja/programs.htm
Anderson MJ (2003b) PCO: a FORTRAN computer program for principal coordinate analysis. Department of Statistics, University of Auckland, Auckland. Available at: www.stat.auckland.ac.nz/~mja/programs.htm
Anderson MJ (2005) PERMANOVA. Permutational multivariate analysis of variance. Department of Statistics, University of Auckland, Auckland. Available at: www.stat.auckland.ac.nz/~mja/programs.htm
Anderson MJ, Willis TJ (2003) Canonical analysis of principal coordinates: a new ecologically meaningful approach for constrained ordination. Ecology 84:511–525
Arvanitidis C, Koutsoubas D, Dounas C, Eleftheriou A (1999) Annelid fauna of a Mediterranean lagoon (Gialova Lagoon, south-west Greece): community structure in a severely fluctuating environment. J Mar Biol Assoc UK 79:849–856
Attrill MJ (2002) A testable linear model for diversity trends in estuaries. J Anim Ecol 71:262–269
Attrill MJ, Rundle SD (2002) Ecotone or ecocline: ecological boundaries in estuaries. Estuar Coast Shelf Sci 55:929–936
Barnes RSK (1991) European estuaries and lagoons, a personal overview of problems and possibilities for conservation and management. Aqua Conserv Marine Freshw Ecosyst 1:79–87
Barnes RSK (1994) A critical appraisal of the application of Guélorget and Perthuisot’s concepts of the paralic ecosystem and confinement to macrotidal Europe. Estuar Coast Shelf Sci 38:41–48
Basset A, Sabetta L, Fonnesu A, Mouillot D, Do Chi T, Viaroli P, Giordani G, Reizopoulou S, Abbiati M, Carrada GC (2006) Typology in Mediterranean transitional waters: new challenges and perspectives. Aqua Conserv Marine Freshw Ecosyst 16:441–455
Benedetti-Cecchi L, Rindi F, Bertocci I, Bulleri F, Cinelli F (2001) Spatial variation in development of epibenthic assemblages in a coastal lagoon. Estuar Coast Shelf Sci 52:659–668
Bilyard GR (1987) The value of benthic infauna in marine pollution monitoring studies. Mar Pollut Bull 18:581–585
Bona F, Cecconi G, Maffiotti A (2000) An integrated approach to assess the benthic quality after sediment capping in Venice lagoon. Aqua Ecosyst Health and Manag 3:379–386
Cao Y, Bark AW, Williams WP (1996) Measuring the responses of macroinvertebrate communities to water pollution: a comparison of multivariate approaches, biotic and diversity indices. Hydrobiologia 341:1–19
Dauer DM (1993) Biological criteria, environmental health and estuarine macrobenthic community structure. Mar Pollut Bull 26:249–257
De Biasi AM, Benedetti-Cecchi L, Pacciardi L, Maggi E, Vaselli S, Bertocci I (2003) Spatial heterogeneity in the distribution of plants and benthic invertebrates in the lagoon of Orbetello (Italy). Oceanol Acta 26:39–46
Fabbri D, Felisatti O, Lombardo M, Trombini C, Vassura I (1998) The Lagoon of Ravenna (Italy): characterisation of mercury-contaminated sediments. Sci Total Environ 213:121–128
Fabbri D, Tartari D, Trombini C (2000) Analysis of poly(vinyl chloride) and other polymers in sediments and suspended matter of a coastal lagoon by pyrolysis-gas chromatography-mass spectrometry. Anal Chim Acta 413:3–11
Fabbri D, Locatelli C, Snape CE, Tarabusi S (2001a) Sulfur speciation in mercury-contaminated sediments of a coastal lagoon: the role of elemental sulfur. J Environ Monitor 3:483–486
Fabbri D, Lombardo M, Trombini C, Vassura I, Zavoli E, Horvat M (2001b) Mercury contamination of a coastal lagoon (Pialassa Baiona, Ravenna, Italy). RMZ Mater Geoenviron 48:186–192
Fiala M (1972) Etudes physico-chimiques des eaux et sédiments de l’étang Bages Sigean (Aude). Vie Et Milieu 23:15–50
Franz DD, Melching CS (1997) Full equations (FEQ) model for the solution of the full, dynamic equations of motion for one-dimensional unsteady flow in open channels and through control structures. U.S. Geological Survey Water-Resources Investigations Report 96-4240
Gamito S (2008) Three main stressors acting on the Ria Formosa lagoonal system (southern Portugal): physical stress, organic matter pollution and the land-ocean gradient. Estuar Coast Shelf Sci 77:710–720
Gray JS (2000) The measurement of marine species diversity, with an application to the benthic fauna of the Norwegian continental shelf. J Exp Mar Biol Ecol 250:23–49
Gray JS, Wu RSS, Or YY (2002) Effects of hypoxia and organic enrichment on the coastal marine environment. Mar Ecol Prog Ser 238:249–279
Guélorget O, Perthuisot JP (1992) Paralic ecosystem. Biological organisation and functioning. Vie Et Milieu 42:215–251
King P, Kennedy H, Newton PP, Jickells TD, Brand T, Calvert S, Cauwet G, Etcheber H, Head B, Khripounoff A, Manighetti B, Carlos Miquel J (1998) Analysis of total and organic carbon and total nitrogen in settling oceanic particles and a marine sediment: an interlaboratory comparison. Mar Chem 60:203–216
Lardicci C, Rossi F, Castelli A (1997) Analysis of macro-zoobenthic community structure after severe dystrophic crises in a Mediterranean coastal lagoon. Mar Pollut Bull 34:536–547
Lardicci C, Como S, Corti S, Rossi F (2001) Recovery of the macrozoobenthic community after severe dystrophic crises in a Mediterranean coastal lagoon (Orbetello, Italy). Mar Pollut Bull 42:202–214
Long ER, MacDonald DD, Smith SL, Calder FD (1995) Incidence of adverse biological effects within ranges of chemical concentrations in marine and estuarine sediments. Environ Manage 19:81–97
McArdle BH, Anderson MJ (2001) Fitting multivariate models to community data: a comment on distance-based redundancy analysis. Ecology 82:290–297
McRae C, Snape CE, Sun C-G, Fabbri D, Tartari D, Trombini C, Fallick AE (2000) Use of compound-specific stable isotope analysis to source anthropogenic natural gas-derived polycyclic aromatic hydrocarbons in a lagoon sediment. Environ Sci Technol 34:4684–4686
Mistri M, Fano EA, Rossi G, Caselli K, Rossi R (2000) Variability in macrobenthos communities in the Valli di Comacchio, northern Italy, a hypereutrophized lagoonal ecosystem. Estuar Coast Shelf Sci 51:599–601
Mistri M, Fano EA, Rossi R (2001) Redundancy of macrobenthos from lagoonal habitats in the Adriatic Sea. Mar Ecol Prog Ser 215:289–296
Munari C, Mistri M (2008) Biodiversity of soft-sediment benthic communities from Italian transitional waters. J Biogeogr 35:1622–1637
Pérez-Ruzafa A, Fernández A, Marcos C, Gilabert J, Quispe J, García-Charton J (2005) Spatial and temporal variations of hydrological conditions, nutrients and. Hydrobiologia 550:11–27
Ponti M, Abbiati M (2004) Quality assessment of transitional waters using a benthic biotic index: the case study of the Pialassa Baiona (Northern Adriatic Sea). Aqua Conserv Marine Freshw Ecosyst 14:S31–S41
Ponti M, Giaquinta S, Abbiati M (2005) Pialassa Baiona lagoon, Ravenna. In: Giordani G, Viaroli P, Swaney DP, Murray CN, Zaldivar JM, Marshall Crossland JI (eds) Nutrient fluxes in transitional zones of the Italian coast. LOICZ reports & studies no 28. Texel, The Netherlands, pp 41–48
Ponti M, Pinna M, Basset A, Moncheva S, Trayanova A, Georgescu L, Beqiraj S, Orfanidis S, Abbiati M (2008) Quality assessment of Mediterranean and Black Sea transitional waters: comparing responses of benthic biotic indices. Aqua Conserv Marine Freshw Ecosyst 18:S62–S75
Prato E, Scardicchio C, Biandolino F (2008) Effects of temperature on the acute toxicity of cadmium to Corophium insidiosum. Environ Monit Assess 136:161–166
Rossi F, Castelli A, Lardicci C (2006) Distribution of macrobenthic assemblages along a marine gradient in Mediterranean eutrophic coastal lagoons. Marine Ecol Evol Perspec 27:66–75
Sacchi CF, Occhipinti-Ambrogi A (1992) Confinement et biocenoses paraliques. Vie Et Milieu 42:207–214
Salas F, Marcos C, Neto JM, Patricio J, Perez-Ruzafa A, Marques JC (2006) User-friendly guide for using benthic ecological indicators in coastal and marine quality assessment. Ocean Coast Manag 49:308–331
Sconfietti R, Marchini A, Ambrogi AO, Sacchi CF (2003) The sessile benthic community patterns on hard bottoms in response to continental vs. marine influence in northern Adriatic lagoons. Oceanol Acta 26:47–56
Specchiulli A, Focardi S, Renzi M, Scirocco T, Cilenti L, Breber P, Bastianoni S (2008) Environmental heterogeneity patterns and assessment of trophic levels in two Mediterranean lagoons: Orbetello and Varano, Italy. Sci Total Environ 402:285–298
Trombini C, Fabbri D, Lombardo M, Vassura I, Zavoli E, Horvat M (2003) Mercury and methylmercury contamination in surficial sediments and clams of a coastal lagoon (Pialassa Baiona, Ravenna, Italy). Cont Shelf Res 23:1821–1831
Underwood AJ (1997) Experiments in ecology. Cambridge University Press, Cambridge
Weis JS, Skurnick J, Weis P (2004) Studies of a contaminated brackish marsh in the Hackensack Meadowlands of Northeastern New Jersey: benthic communities and metal contamination. Mar Pollut Bull 49:1025–1035
Acknowledgments
This study was done within the research protocol among the Centre for Environmental Science of the University of Bologna (CIRSA), Ravenna Municipality, Local Health Agency (AUSL) and Environmental Protection Agency (ARPA-EMR). Hydrodynamic data were provided by Renata Archetti, Massimo Guerrero and Alberto Lamberti (from DISTART, University of Bologna), while chemical analyses were performed by Pilar Lopez, Mayte Llebrés, Roberta Guerra, Valentina Baravelli and Daniele Fabbri (CIRSA). We also thank Eleonora Torricelli for sediment grain size and organic matter analyses and Andrea Pasteris for constructive discussion on sampling design and analyses.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Luis Gimenez.
Appendix
Rights and permissions
About this article
Cite this article
Ponti, M., Casselli, C. & Abbiati, M. Anthropogenic disturbance and spatial heterogeneity of macrobenthic invertebrate assemblages in coastal lagoons: the study case of Pialassa Baiona (northern Adriatic Sea). Helgol Mar Res 65, 25–42 (2011). https://doi.org/10.1007/s10152-010-0197-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-010-0197-0