
Abstract
Biomonitoring of mussel bed assemblages can provide valuable information about the impact of pollution on hard substrate assemblages. This study of Mytilus galloprovincialis mussel beds in Thermaikos Gulf (northern Aegean Sea) deals with the spatial and temporal structure of the associated fauna. Samples were collected and abiotic factors were measured in two successive years. Common biocoenotic methods were employed to analyze the data. The samples could be separated into three groups, with summer and winter samples being clearly different. A total of 100 species were found: polychaetes and crustaceans were the most dominant taxa. The assemblage shows high diversity with respect to species abundance. Biotic interactions within the assemblage appear to influence its composition, although the total evenness remains unaffected in space and time. The M. galloprovincialis assemblages can be found in clean as well as in polluted waters and, therefore, are of great interest in biomonitoring studies.
Similar content being viewed by others


Introduction
Mussel bed (Mytilus galloprovincialis LMK) assemblages can develop in clean and moderately polluted as well as polluted waters (e.g. Bellan-Santini et al. 1994; Damianidis and Chintiroglou 2000). Therefore, biomonitoring of these assemblages can provide valuable information about the impact of pollution on hard bottom communities (e.g. Wenner 1988).
There is adequate information about the structure of M. galloprovincialis assemblages from various regions in the Mediterranean Sea, especially from the western Mediterranean coasts (Bellan 1969, 1980; Bellan-Santini 1969, 1981; Desrosiers et al. 1982, 1986; Hong 1983; Tursi et al. 1984; Tsuchiya and Bellan-Santini 1989), while information on the Aegean Sea is relatively limited (Kocatas 1978; Topaloglou and Kihara 1993; Lantzouni et al. 1998; Damianidis and Chintiroglou 2000). Many authors have put emphasis on the importance of these assemblages in biomonitoring studies (see Thiel and Ullrich 2002). Damianidis and Chintiroglou (2000) reported that the abiotic factors at all sampling sites within Thermaikos Gulf do not fluctuate in time. Therefore, any variation in the composition of M. galloprovincialis assemblages has to be attributed to biotic factors.
This study aims at the investigation of the spatial and temporal structure of the fauna associated with M. galloprovincialis assemblages on the eastern coast of Thermaikos gulf.
Methods
Study area
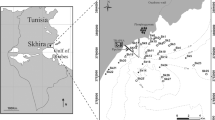
The sampling sites were selected on the basis of their historical background, as well as the exposure of the assemblages and the depth of their occurrence. Of all locations with artificial hard substrate along the east coast of Thessaloniki Bay, two appeared to be very similar in bathymetric distribution and exposure of M. galloprovincialis populations: the piers of Agia Triada (ST1) and Perea (ST2) (Fig. 1). These piers were constructed 25 years ago and are supported by concrete pillars, which comprise the substrate for the mussel beds. At these sampling sites, the M. galloprovincialis populations develop a uniform physiognomic aspect, with large numbers of mussels covering an area that extends from the lower infralittoral zone down to a depth of 2.5 m.

Map showing Thermaikos Gulf and the two sampling sites (ST1, ST2)
Physico-chemical factors
During this study, physico-chemical factors such as salinity (S‰), conductivity (μS/cm), water clarity (m), dissolved oxygen (mg/l), temperature (°C) and total hydrodynamics were measured. All measurements were made using WTW (Wissenschaftlich-Technische Werkstaetten, Weilheim, Germany) and Lovibond Checkit (Dortmund, Germany) micro-electronic equipment and water clarity was examined using the Secchi disc. These measurements were conducted monthly at each sampling site and the results have already been reported by Damianidis and Chintiroglou (2000).
Sampling methods
Sampling was carried out while scuba diving. Samples were taken as described by Chintiroglou and Koukouras (1992). The area covered by the quadrat sampler was 400 cm2 (Stirn 1981; Bakus 1990). Three replicates were taken each time. The samples, 24 in total, were collected during winter and summer of 1994 and 1995. After sampling, the specimens were preserved in 10% formalin and were transferred to the laboratory for further treatment. All samples were collected by the same scuba diver.
Data analysis
Common biocoenotic methods were employed to analyze the faunal composition of the M. galloprovincialis assemblages (Bellan-Santini 1981; Damianidis and Chintiroglou 2000). Hence, the numerical abundance (N) on a scale of 1 m2, the mean dominance (D) and the frequency (f) were estimated. Also Shannon-Weaver’s (H′), and Margalef (d) and Pielou’s Evenness (J′) were calculated on a log2 basis (Daget 1979).
Seasonal differences in the mean numbers of individuals were tested using one-way ANOVA and multiple comparisons. All data were then converted to logarithms. The Spearman rank correlation coefficient (r s) was employed to determine the relation of the number of mussels (Ab M) with faunal abundance (mAb) and richness (R).
The numerical abundance data, obtained per sampling station, were analyzed using cluster and multidimensional scaling (mds) techniques, based on the Bray-Curtis similarity, using the PRIMER package (see Clarke and Green 1988; Clarke and Warwick 1994). The square root transformation was applied in order to increase the contribution of the rare species (Clarke and Warwick 1994). The significance of the multivariate results was assessed using the ANOSIM test. SIMPER analysis was performed in order to identify the percentage contribution of each species to the overall similarity within a site and the dissimilarity among sites (Clarke 1993). The above were carried out to examine the similarity degree of samples, in both space and time.
Results
Composition of the assemblage
A total of 100 species were found associated with M. galloprovincialis assemblages in Thermaikos gulf (Table 1). The distribution of these species in major taxonomic groups is given in Table 2. The dominant groups are polychaetes (37.5%) and crustaceans (30.9%). As shown in Table 1, 17 species were distinguished as “very common” (f≥50%), 26 as “rare” (f<10%), and 57 as “common” (10%<f<50%).
Among the very common species, the polyclad Stylochus sp. and the decapods Pilumnus hirtelus and Pisidia longicornis have been described as the main predators of mussels (see Galleni et al. 1977; Damianidis and Chintiroglou 2000). The cirripeds Balanus perforatus and B. trigonus, the polychaete Serpula vermicularis and Bryozoa sp1, are well-known as organisms that often settle on mussel shells (Bussani 1983; Damianidis and Chintiroglou 2000). The rest of the very common species are free motile organisms which employ various modes of feeding. The tube-building peracarids are detritivores (Barnard 1958, 1963; Isaac et al. 1994), Ophiothrix sp. (Pérès 1976) is a suspensivore, and the polychaete Staurocephalus rudolphii is a carnivore (Fauchald and Jumars 1979).
Relationships between fauna and structure of mussel beds
According to Tsuchiya and Nishihira (1986), the morphology and relative age of the mussels in an assemblage can play a significant role in the composition of the associated fauna. Therefore, the correlations of mussel abundance with species richness and faunal abundance, respectively, were examined.
For both summer and winter samples, faunal abundance (mAb) and richness (R) were not correlated (P>0.5) with the mussel abundance (Ab M).
Diversity
A total of 17,090 individuals, representing 100 faunal species, were examined. As shown in Table 1, the number of species and the diversity indices (H′, d and J′), were determined for each sampling station and season (winter/summer). The number of species ranged from 37 to 49 in winter, and from 45 to 60 in summer. The diversity indices ranged proportionately as they appeared to be higher in the summer samples (Table 1).
Abundance
The comparison of the faunal abundance in time (within and between years) and in space (sampling sites) was based on the examination of the null hypothesis that the abundance of the fauna does not differ significantly. One-way ANOVA was used to detect the exact differences and showed an equal distribution of the abundance of the fauna in winter as well as in summer samples (F=0.58, df=1,3, P=0.64 in winter samples, and F=2.47, df=1,3, P=0.14 in summer samples).
Affinity of sampling in space and time
The affinity of all sampling sites is given in Fig. 2. Both analyses (cluster and non-metric mds) indicate a separation of samples into three main groups at about 50% similarity level. The winter samples from 1995 form group A, the winter samples from 1994 group B and the summer samples from both years (1994 and 1995) group C. Summer and winter samples are separated, indicating seasonality of the benthic assemblages. The two stations were not separated. Samples from successive years were only separated in winter. The stress value for the two-dimensional mds configuration is 0.01, indicating an excellent ordination of samples (Clarke and Warwick 1994). The performance of a one-way ANOSIM test gave global R=1 at a significance level of P<0.005, so the separation of the three groups (A, B and C) was confirmed. Group A showed an average similarity of 62%. As identified by SIMPER analysis, five species (M. galloprovincialis, B. perforatus, Elasmopus rapax, Pisidia longicornis and B. trigonus) were responsible for 60% of the average similarity, and 15 species for 90%. Group B reached an average similarity of 57%, with seven species (M. galloprovincialis, Pisidia longicornis, Ophiothrix fragilis, B. perforatus, B. trigonus, Pilumnus hirtelus and Athanas nitescens) covering 60% of this similarity, and 22 species 90%. Finally, group C reached an average similarity of 67%, with eight species (Corophium sp., M. galloprovincialis, B. trigonus, O. fragilis, Pisidia longicornis, Prionospio malmgreni, S. vermicularis and Pilumnus hirtelus) being responsible for 60% of the similarity, and 26 species for 90%. As regards the divergence between groups, we found that group A had an average dissimilarity percentage of 50% with group B (21 species contributed 60% and 55 species 90% of this value) and 57% with group C (16 species contributed 60% and 54 species 90%), while the dissimilarity percentage between groups B and C was 50% (22 species contributed 60%, and 58 species 90%). It is quite clear that only a few species are important for characterizing the groups, while most of them are important for differentiating the groups.

Affinity of the sampling sites according to cluster and non-metric multidimensional sampling analyses
Discussion
Photophilic soft algae communities probably have the highest faunal and flora diversity of all benthic biocoenoses in the Mediterranean (Pérès 1982; Bellan-Santini et al. 1994). The assemblage of mussel beds of M. galloprovincialis in Thermaikos Gulf belongs to this type of biocoenosis, showing high diversity with respect to species abundance. One hundred animal species were recorded during this study, 17 of which were characterized as very common (f≥50%), and 57 as common (10%<f<50%). The recorded species have been reported by many authors as members of the assemblages of sublittoral photophilic algae (Bellan-Santini 1969; Saldanha 1974; Kocatas 1978; Bellan 1980; Hong 1983; Desrosiers et al. 1986; Marinopoulos 1988; Chintiroglou and Koukouras 1992; Topaloglou and Kihara 1993; Bellan-Santini et al. 1994; Lantzouni et al. 1998; Baxevanis and Chintiroglou 2000; Damianidis and Chintiroglou 2000).
The biodiversity of M. galloprovincialis assemblages in Thermaikos Gulf differs from those reported for other Mediterranean and northern European sites (Table 3). Most authors have used similar methods for the study of hard substrates (based mostly on work by Bellan-Santini 1969) (Table 3), yet data from different geographical areas are difficult to compare. The highest species abundance (R=131) was found in the infralittoral zone of the Portuguese coast. The clean waters of Marseilles, Ismir and Thermaikos Gulf also show high species abundance, while species abundance was relatively low in the Bosporus, on Danish coasts, in the midlittoral zone of the Portuguese coast and in the polluted waters of Marseilles. The respective values in Japan and Chile ranged around 65, while 89 species were counted in Italy and 56 in Aberffraw (N Wales).
Table 3 also shows that polychaetes and crustaceans are the most important taxonomic groups of the assemblage, contributing almost 50% of the total faunal species abundance. The abundance of these groups, however, varies from one area to another and also seems to be dependent on the specific features of each study area (polluted/non-polluted; midlittoral/supralittoral), a fact that was also reported by Thiel and Ullrich (2002). Saldanha (1974), for instance, recorded fewer polychaetes and more crustacean species on the coasts of Portugal, while D’Anna et al. (1985) recorded exactly the opposite for Sicily. Kocatas (1978) found 35 polychaete and 32 crustacean species in the Izmir Bay, whereas Topaloglou and Kihara (1993) reported 10 polychaete and 22 crustacean species for the Bosporus. Seed and Suchanek (1992) drew the same conclusions, even though the Mediterranean mussel assemblages were not the focus of their work. The total diversity of mussel assemblages in the studied areas shows no significant variations. Furthermore, significant similarities exist at the taxonomic level. It should also be noted that, according to Damianidis and Chintiroglou (2000), there is no significant difference in the composition of the dominant species: this implies that the structure of the polychaete fauna of M. galloprovincialis assemblages in the Mediterranean Sea is largely homogeneous.
An important part of the study of mussel assemblages is the study of interactions between the structure of mussel populations and the associated fauna (Tsuchiya and Nishihira 1986; Lintas and Seed 1994). These studies have produced contradictory results. According to Tsuchiya and Nishihira (1986), the structure of mussel beds in the Pacific Ocean (Japan) has a direct effect on the diversity of the assemblages. Lintas and Seed (1994) suggested that the fauna associated with M. edulis appear to be related largely to mussel density. Damianidis and Chintiroglou (2000) reported similar results on the polychaete fauna of mussel beds in Thermaikos Gulf. In the present study, however, no relationship was found between mussel density and the abundance and diversity of the assemblage. As regards the structure (= distribution of size classes of the populations) of the M. galloprovinicialis populations, there is certain information in Le Breton and Chintiroglou (1998) indicating an uneven distribution in space and time. Although information is still limited, it appears that biotic interactions have a strong effect on the composition of the assemblage, which seems to decrease when the assemblage is studied as a whole. In this case, the composition of the assemblages is largely even in space and time. Nevertheless, attention should be paid to any variations in order to understand the biotic interactions in hard substrate assemblages (see Damianidis and Chintiroglou 2000).
The evenness of the studied assemblage showed some variation in space, and particularly in time, while the total homogeny of the faunal composition remained unaffected. The separate analyses conducted by Lantzouni et al. (1998) and Damianidis and Chintiroglou (2000) produced similar results.
In conclusion, there is now adequate information about M. galloprovincialis assemblages, although there are still open questions. For biomonitoring of marine benthic assemblages, knowledge of the structure and function of M. galloprovincialis assemblages can play an important role. The main advantage of studying such assemblages, on a smaller or wider scale, is their similar physiognomic appearance (Reish 1971; Wenner 1988). As these assemblages can be found in clean as well as in polluted waters, they are of great interest in biomonitoring studies (see Wenner 1988).
References
Bakus JG (1990) Quantitative ecology and marine biology. Balkema, Rotterdam
Barnard JL (1958) Amphipoda crustaceans as fouling organisms, in Los Angeles-Long Beach harbors, with reference to the influence of seawater turbidity. Calif Fish Game 44:161–170
Barnard JL (1963) Relationship of benthic Amphipoda to invertebrate communities of inshore sublittoral sands of Southern California. Pac Nat 3:439–467
Baxevanis A, Chintiroglou C (2000) Peracarida crustacean of the artificial hard substratum in N. Michaniona (N. Aegean). Belg J Zool 130:11–16
Bellan G (1969) Polychètes des horizons supérieurs de l’étage infralittoral rocheux dans la région Provençale. Tethys 1:349–366
Bellan G (1980) Relationship of pollution to rocky substratum polychaetes on the French Mediterranean coast. Mar Pollut Bull 11:318–321
Bellan-Santini D (1969) Contribution à l’ étude des peuplements infralittoraux sur substrat rocheux. Trav Stat Mar Endoume Bull 31:1–294
Bellan-Santini D (1981) Influence des pollutions sur le peuplement des amphipodes dans la biocénose des algues photophiles. Tethys 10:185–194
Bellan-Santini D, Lacaze JC, Poizat C (1994) Les biocénoses marines et littorales de Méditerranée, synthèse, menaces et perspectives. Muséum National d’ Histoire Naturelle Paris
Bussani M (1983) Guida pratica di Mitilicoltura. Edagricole, Bologna
Chintiroglou C, Koukouras A (1992) A population of the sea anemone Anemonia viridis (Forskal, 1775) and its associated flora and fauna in the North Aegean Sea. Int Rev Hydrobiol 77:483–495
Clarke KR (1993) Non-parametric multivariate analyses of changes in community structure. Aust J Ecol 18:117–143
Clarke KR, Green RH (1988) Statistical design and analysis for a ‘biological effects’ study. Mar Ecol Prog Ser 46:213–226
Clarke KR, Warwick MR (1994) Change in marine communities: an approach to statistical analysis and interpretation. Natural Environment Research Council, UK
Daget J (1979) Les modèles mathématiques en écologie. Masson, Paris
Damianidis P, Chintiroglou C (2000) Structure and function of Polychaetofauna living in Mytilus galloprovincialis assemblages in Thermaikos Gulf (north Aegean Sea). Oceanol Acta 23:323–339
D’Anna, Giaccone G, Riggio S (1985) Lineamenti bionomici dei banchi di Mitili di Balestrate (Sicilia Occidentale). Oebalia 11:389–399
Desrosiers G, Bellan-Santini D, Brethes JC (1982) Évolution spatio-temporelle des peuplements de substrats rocheux superficiels dans un golfe soumis à de multiples pollutions (Golfe de Fos, France). Tethys 10:245–253
Desrosiers G, Bellan-Santini D, Brethes JC (1986) Organisation trophique de quatre peuplements de substrats rocheux selon un grandient de pollution industrielle (Golfe de Fos, France). Mar Biol 9:107–120
Fauchald K, Jumars P (1979) The diet of worms: a study of polychaete feeding guilds. Oceanogr Mar Biol Annu Rev 17:193–284
Galleni L, Ferrero E, Salghetti V, Tongiorgi P, Salvadego P (1977) Ulteriori osservazioni sulla predazione di Stylochus mediterraneus (Turbellaria Polycladida) sui Mytili e suo orientamento chemiotattico. Atti IX Congr Soc Ital di Biol Mar, Lacco Ameno d’Ischia, pp 259–261
Hong JS (1983) Impact of the pollution on the benthic community. Bull Korean Fish Soc 16:279–290
Isaac MJ, Makings P, Naylor E, Smaldon G, Withers RG (1994) Crustacean II: Malacostraca Peracarida. In: Hayward PJ, Ryland JS (eds) The marine fauna of the British Isles and North-West Europe, vol I. Introduction and Protozoans to Arthropods. Oxford Science, pp 362–488
Kocatas A (1978) Contribution à l’étude des peuplements des horizons supérieurs de substrat rocheux du Golfe D’Izmir (Turquie). Ege Univ Fen Fak Monogr Ser 12:1–93
Lantzouni M, Voultsiadou E, Chintiroglou C (1998) Preliminary observations on amphipod assemblages associated with Mytilus galloprovincialis Lam. beds from Thermaikos Gulf (Aegean Sea). Rapp Comm Int Mer Médit 35:458–459
Le Breton JM, Chintiroglou C (1998) Etude préliminaire de la structure de trois populations de moules Mytilus galloprovincialis LMK sur la côte est de la baie de Thessaloniki. Rapp Comm Int Mer Médit 35:558–559
Lintas C, Seed R (1994) Spatial variation in the fauna associated with Mytilus edulis on a wave-exposed rocky shore. J Molluscan Stud 60:165–174
Marinopoulos J (1988) Etude des peuplements infralittoraux de substrats rocheux de la region de Marseille et des facteurs abiotiques (lumière, hydrodynamique) les influençant. Thesis. Université d’Aix Marseille
Pérès JM (1976) Précis d’océanographie biologique. Presses Universitaires de France
Pérès JM (1982) General features of organismic assemblages in pelagial and benthal. Structure and dynamics of assemblages in the benthal. Major benthic assemblages. Specific benthic assemblages. In: Kinne O (ed) Marine ecology, vol 5(1). Wiley, Chichester, pp 47–66, 119–186, 373–522, 523–581
Reish DR (1971) The effect of pollution abatement in Los Angeles Harbor, California. Mar Pollut Bull 2:71–74
Saldanha L (1974) Estudo do povoamento dos horizontes superiores da rocha litoral da costa da Arrabida (Portugal). Publ Mus Lab Zool Anthropol Fac Cien, Lisboa
Seed R, Suchanek TH (1992) Population and community ecology of Mytilus. In: Gosling EM (ed) The mussel Mytilus: ecology, physiology, genetics and culture. Elsevier, Amsterdam, pp 87–169
Stirn J (1981) Manual of methods in aquatic environment research. Part 8. Ecological assessment of pollution effects (Guidelines for the FAO GFCM/UNEP Joint Coordinated Project on Pollution in Mediterranean). FAO Fisheries Technical Paper
Svanne I, Setyobudiandi I (1996) Diversity of associated fauna in beds of the blue mussel Mytilus edulis L.: effects of location, patch size and position within a patch. Ophelia 45:39–53
Thiel M, Ullrich N (2002) Hard rock versus soft bottom: the fauna associated with intertidal mussel beds on hard bottoms along the cost of Chile, and considerations on the functional role of mussel beds. Helgol Mar Res 56:21–30
Tiganus V (1979) Observations sur la structure qualitative et quantitative de la biocenose des moules de rocher du littoral roumain de la Mer Noire. Rapp Comm Int Mer Medit 25/26:4
Topaloglou B, Kihara K (1993) Community of Mediterranean mussel Mytilus galloprovincialis Lamark, 1819 in the Bosphorus strait. J Tokyo Univ Fish 80:113–120
Tsuchiya M, Bellan-Santini D (1989) Vertical distribution of shallow rocky shore organisms and community structure of mussel beds (Mytilus galloprovincialis) along the coast of Marseille, France. Mésogée 49:91–110
Tsuchiya M, Nishihira M (1986) Islands of Mytilus edulis as a habitat for small intertidal animals: effect of Mytilus age structure on the species composition of the associated fauna and community organization. Mar Ecol Prog Ser 31:171–178
Tursi A, Matarrese A, Scalera A, Liaci L, Montanaro C, Cecere E (1984) Seasonal factors in mussel facies settlement on annual fouling panels. Oebalia 10:69–84
Wenner AM (1988) Crustacean and other invertebrates as indicators of beach pollution. In: Soule DF, Kleppel GS (eds) Marine organisms as indicators. Springer, Berlin Heidelberg New York, pp 199–229
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.-D. Franke
Rights and permissions
About this article
Cite this article
Chintiroglou, CC., Damianidis, P., Antoniadou, C. et al. Macrofauna biodiversity of mussel bed assemblages in Thermaikos Gulf (northern Aegean Sea). Helgol Mar Res 58, 62–70 (2004). https://doi.org/10.1007/s10152-003-0169-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-003-0169-8