
Abstract
Toll-like receptors (TLRs) recognize distinct microbial components and induce innate immune responses. TLR5 has been shown to recognize bacterial flagellin. Unlike other TLRs, TLR5 is not expressed on conventional dendritic cells or macrophages. By contrast, TLR5 is mainly expressed on intestinal CD11c+ lamina propria cells (LPCs), which do not express TLR4. These cells detect pathogenic bacteria and secreted proinflammatory cytokines, mainly in a TLR5-dependent manner. Notably, transport of pathogenic Salmonella typhimurium from the intestinal tract to mesenteric lymph nodes was impaired in Tlr5 −/− mice, suggesting that CD11c+ LPCs expressing TLR5 are used by S. typhimurium for systemic infection. CD11c+ LPCs consist of four subsets distinguished by differential expression patterns of CD11c and CD11b. CD11chiCD11bhi LPDCs have been identified as TLR5-expressing cells. In response to flagellin, these LPDCs induce the differentiation of naive B cells into IgA+ plasma cells via a mechanism independent of gut-associated lymphoid tissue (GALT), and trigger the differentiation of antigen-specific Th17 and Th1 cells. These LPDCs have unique properties in that they can induce acquired immunity as well as innate immunity via TLR5 in the intestine.
Similar content being viewed by others


Introduction
Toll-like receptors (TLRs) recognize a variety of microbial components and induce innate immune responses [1]. They are abundantly expressed on ‘professional’ antigen-presenting cells (APCs) such as macrophages and conventional dendritic cells (DCs) and serve as an important link between the innate and adaptive immune responses. To date, 13 TLRs have been identified in mammals. Bacterial flagellin is a structural protein that forms the main portion of flagella and promotes bacterial chemotaxis and bacterial adhesion to and invasion of host tissues [2]. TLR5 recognizes the conserved domain in flagellin monomers and triggers proinflammatory as well as adaptive immune responses [3, 4]. TLR5 is expressed on the basolateral surfaces of intestinal epithelial cells and is thought to be key in the recognition of invasive flagellated bacteria at the mucosal surface [5]. Human intestinal epithelial cell lines produce chemokines in response to flagellin, leading to subsequent migration of immature DCs [6]. Furthermore, TLR5 is highly expressed in the human lung and a common TLR5 polymorphism in humans causes susceptibility to legionellosis [7, 8]. Although accumulating evidence suggests that TLR5 is critical for host defense against enterobacterial infection, the in vivo function of TLR5 has not been fully elucidated.
Identification of TLR5-expressing cells in the intestine
Unlike other TLR family members, TLR5 is not expressed on macrophages and conventional DCs in mice. Instead, TLR5 is highly expressed in the small intestine [9]. In contrast to the findings of a previous report, TLR5 expression was not high in intestinal epithelial cells (IECs). IECs could not produce any inflammatory cytokines in response to flagellin, although they did produce defencin-β3 and CCL27. On the other hand, TLR5 was highly expressed in lamina propria cells (LPCs). TLR family members are preferentially expressed on APCs. In the LP of the mouse small bowel, DCs have been thought to be the dominant APCs. Interestingly, a considerable amount of TLR5 mRNA was detected in CD11c+ LPCs, but none was detected in CD11c− LPCs. Thus, CD11c+ LPCs were identified as TLR5-expressing cells in the intestine (Fig. 1) [9].

TLR5 is highly expressed on CD11c+ LPCs. Confocal microscopy of frozen tissue sections of the small intestine and Peyer’s patch of C57BL/6 mice, fixed and stained with antibodies specific for CD11c (red) and TLR5 (green) [9]
Innate immune responses by CD11c+ LPCs
Because the intestine is constantly exposed to food antigens (Ags) and commensal bacteria, it is the consensus that most mucosal DCs induce tolerance, but not inflammation. In the Peyer’s patches and mesenteric lymph nodes, conventional DCs consist of CD11chiCD11b+CD8a–, CD11chiCD11b–CD8a+ and CD11chiCD11b−CD8a− subsets. In addition, CD11cint plasmacytoid DCs exist in these sites [10]. DCs in the Peyer’s patch produce interleukin 10 (IL-10) rather than IL-12, polarize naive T cells toward T helper type 2 (Th2) or regulatory T (Treg) cells, and induce the differentiation of immunoglobulin A (IgA)+ plasma cells [11]. Flagellin-stimulated CD11c+ LPCs do not produce IL-10 and tumor-necrosis factor (TNF)-α, but instead produce IL-6 and IL-12 [9]. Furthermore, CD11c+ LPCs express chemokines, prostaglandins, antimicrobial peptides and molecules involved in cellular adhesion, cytoskeletal organization and intracellular transport in response to flagellin [9]. Accordingly, CD11c+ LPCs have a tendency to induce inflammatory responses rather than tolerance when exposed to flagellin.
Recognition of bacteria by CD11c+ LPCs
TLR4, which is abundantly expressed on innate immune cells such as conventional DCs and macrophages, recognizes lipopolysaccharide (LPS) of Gram-negative bacteria [1]. However, CD11c+ LPCs do not produce any inflammatory cytokines in response to LPS owing to their low expression of TLR [49]. Conventional DCs, which express TLR4 but not TLR5, recognize Gram-negative flagellated bacteria, mainly via TLR4, while TLR4−TLR5+CD11c+ LPCs produce inflammatory cytokines after exposure to such bacteria [9]. The unique profile of TLR expression in CD11c+ LPCs seems to be closely related to the specific environment in the intestine. Most commensal bacteria in the intestine are Gram-negative anaerobic rod bacteria, which contain LPS in their cell walls. The low expression of TLR4 may allow CD11c+ LPCs to avoid inducing excessive immune responses to commensal bacteria. Instead, CD11c+ LPCs may induce inflammatory responses to pathogenic flagellated bacteria, which are able to invade the LP, via TLR5. However, some commensal bacteria also have flagella. Recently, it was reported that α- and ε-proteobacteria can change the TLR5 recognition site in flagellin without losing flagellar motility [12]. Furthermore, some commensal bacteria in the intestine suppress flagellin expression [13]. Unlike pathogenic bacteria, commensal bacteria may have mechanisms to escape TLR5-mediated host detection in the intestine.
Role of TLR5 in Salmonella typhimurium infection
S. typhimurium is a facultative intracellular pathogenic bacterium that causes typhoid-like disease in mice. In oral infection, S. typhimurium are captured by sub-epithelial DCs after transport through M cells in Peyer’s patches (PPs) or intra-epithelial DCs in LPs [14, 15]. After internalization, S. typhimurium inject effector proteins into the cytoplasm via a type III secretion system and inhibit phagosome-lysosome fusion. S. typhimurium can safely replicate in Salmonella-containing vacuoles (Fig. 2) [16]. On the other hand, bacteria-laden DCs undergo maturation and migrate to the T-cell zones of PPs or draining mesenteric lymph nodes (MLNs). These mature DCs are also thought to be responsible for the dissemination of S. typhimurium via the blood stream to the liver and spleen [13, 17]. Although CD11c+ LPCs express inflammatory cytokines after exposure to S. typhimurium in vitro, via a TLR5-dependent mechanism, Tlr5 −/− mice showed resistance to oral S. typhimurium infection. The transport of S. typhimurium from the LP to MLNs was impaired in Tlr5 −/− mice. As S. typhimurium could not fully activate and induce maturation of Tlr5 −/− CD11c+ LPCs, migration of bacteria–laden CD11c+ LPCs may be inefficient in Tlr5 −/− mice. S. typhimurium may use CD11c+ LPCs as carriers during systemic infection by inversely utilizing the host defense activity of TLR5 (Fig. 3) [9].

Type III secretion system of S. typhimurium. S. typhimurium inject effector proteins into the cytoplasm via a type III secretion system and inhibit phagosome-lysosome fusion. S. typhimurium can safely replicate in Salmonella-containing vacuoles

Impaired transport of S. typhimurium in Tlr5 −/− mice. S. typhimurium are captured by sub-epithelial DCs after transport through M cells in PPs or intra-epithelial DCs in LPs. Bacteria-laden DCs undergo maturation and migrate to the T-cell zones of PPs or draining MLNs. These maturate DCs are also responsible for the dissemination of S. typhimurium via the blood stream to the liver and spleen. The transport of S. typhimurium from the LP to MLNs was impaired in Tlr5 −/− mice. As S. typhimurium could not fully activate and induce maturation of Tlr5 −/− CD11c+ LPCs, migration of bacteria–laden CD11c+ LPCs may be inefficient in Tlr5 −/−mice
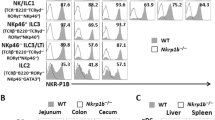
Four subsets of CD11c+ LPCs in the small intestine
CD11c+ LPCs in murine small intestine consist of four subsets distinguished by differential expression patterns of CD11c and CD11b: two subsets of DCs (CD11chiCD11blow and CD11chiCD11bhi), macrophages (CD11cintCD11bint) and eosinophils (CD11cintCD11bmid) (Fig. 4a). The CD11chiCD11blow and CD11chiCD11bhi subsets have a DEC-205+ major histocompatibility complex (MHC) class II-high CD80+CD86+CD103+ surface phenotype. In addition, the CD11chiCD11bhi subset is moderately F4/80 positive, suggesting that this subset expresses both DC (DEC-205) and macrophage (F4/80) markers. The CD11cintCD11bint subset consists of F4/80+DEC-205− MHC class II+ phagocytic macrophages. The CD11cintCD11bmid subset consists of eosinophils with uniquely shaped nuclei and eosinophilic granules, which express CD80 but not MHC class II (Fig. 4b) [18]. Among the four subsets of CD11c+ LPCs in the mouse small intestine, CD11chiCD11bhi lamina propria dendritic cells (LPDCs) specifically express TLR5 (Fig. 4c). This subset produces IL-6 and IL-12 in response to flagellin in a TLR5-dependent manner. However, they did not produce IL-10, TNF-α or IL-23. CD11chiCD11bhi LPDCs also express TLR9, and produce IL-6 and IL-12 in response to CpG DNA. In contrast to CD11chiCD11bhi LPDCs, other CD11c+ subsets in the LP do not produce such cytokines in response to flagellin. Thus, CD11chiCD11bhi LPDCs are responsible for TLR5-mediated innate immune responses in the intestinal LP [18].

Four subsets of CD11c+ LPCs. a Flow cytometry of intestinal low-density LPCs stained for CD11b and CD11c, before and after sorting. b May-Grunwald-Giemsa staining of four leukocyte subsets (gated in a) from the lamina propria. Scale bars 10 μm. c RT-PCR of Tlr5 expression in the four leukocyte lamina propria subsets. Actb encodes β-actin [18]
TLR5 and intestinal IgA production
The intestine is known as an organ that produces large amounts of secretory IgA [19]. Intestinal gut-associated lymphoid tissues (GALTs) such as PPs, ILFs and MLNs are major sites for generation of IgA+ plasma cells in the intestine. IgA+ plasma cells are induced there via a mechanism dependent on Ag, CD4+ follicular B helper T cells and the formation of germinal centers (GCs) [20–22]. Suppressive Foxp3+CD4+ T cells in PPs can differentiate efficiently into cells with characteristics of follicular B helper T cells, which then participate in the induction of GCs and IgA production in the gut[23]. CD103+ GALT DCs produce retinoic acid (RA), which induces the selective expression of gut homing receptors, such as integrin α4β7 and CCR9, on differentiated IgA+ plasma cells for gut homing [24, 25]. However, differentiation of IgA+ cells does not necessarily require T cell help and GC formation. GALT DC-derived RA and cytokines synergistically act on naïve B cells, leading to the generation of T cell–independent IgA+ cells [25, 26]. Furthermore, some IgM+ B cells, especially peritoneal B1 cells, directly migrate to the gut LP in a sphingosine 1-phosphate (S1P)-dependent manner and differentiate into IgA+ plasma cells in the LP with the help of stroma cells [27]. Commensal bacteria induce natural secretory IgA, and this process is mediated by commensal bacteria-laden DCs [28]. Furthermore, DCs in the small intestinal LP send protrusions into the lumen through an intercellular gap between epithelial cells via a mechanism dependent on the chemokine receptor CX3CR1, and actively sample luminal bacteria [29]. Although accumulating evidence suggests the involvement of DCs in the generation of IgA+ plasma cells in the LP, it is not known what subset of DCs is responsible for this event. Interestingly, CD11chiCD11bhi LPDCs are responsible for the generation of IgA+ plasma cells in the LP. Flagellin-stimulated CD11chiCD11bhi LPDCs efficiently induced the differentiation of B220− IgA+ plasma cells in the absence of T cells in a TLR5-dependent manner. Similar to CD103+ DCs in GALTs, CD11chiCD11bhi LPDCs specifically express Aldh1a2 mRNA, which encodes retinal dehydrogenase 2 (RALDH2). CD11chiCD11bhi LPDC-derived RA synergistically acts on naïve B cells with TLR5-mediated inflammatory cytokines, leading to the induction of IgA+ plasma cells. Furthermore, TLR5 is critical for IgA synthesis in vivo. Mice lacking the transcription factor Id2 do not develop GALTs, yet they retain intestinal IgA production. In wild-type mice, around 20% of IgA+ plasma cells exist in the small intestinal LP. Interestingly, about 10% of IgA+ cells are found in the LP of Id2 −/− mice, which confirms that gut IgA can be generated without GALTs. Although Tlr5 −/− mice did not have fewer IgA+ B cells, Id2 −/− Tlr5 −/− mice had far fewer IgA+ cells in the LP [18]. Thus, TLR5 signaling in CD11chiCD11bhi LPDCs is essential for GALT-independent IgA synthesis.
IgA class-switch recombination (CSR) is severely impaired in inducible nitric oxide synthase (iNOS)-deficient mice [30]. iNOS regulates the expression of transforming growth factor-β receptor (TGF-βR) II and plays a critical role in T-cell-dependent IgA CSR. iNOS is also involved in T-cell-independent IgA CSR through the production of a proliferation-inducing ligand (APRIL, also called Tnfsf13) and a B-cell-activating factor of the TNF family (BAFF, also called Tnfsf13b). iNOS is preferentially expressed in DCs in both GALTs and LPs. The number of iNOS+ DCs was reduced in the mucosa-associated lymphoid tissues (MALTs) of germ-free mice, Myd88 −/− mice and Tlr2 −/− 4 −/− 9 −/− mice, suggesting that this DC subset is induced via a TLR-dependent mechanism. These iNOS+ DCs have a TNF-α+CD11b+MHC class II+CD80+CD86loLy6C+Ly6G−Gr-1+Mac3+ surface phenotype, revealing that they are presumably a TNF-α/iNOS-producing DC (TipDC) subset, originally identified in mice infected with Listeria monocytogenes [31, 32]. Although both iNOS+ DCs and CD11chiCD11bhi LPDCs are important for IgA synthesis in the intestine, their phenotypes and functions are completely different (Fig. 5a). Based on the expression of surface molecules, iNOS+ DCs are similar to CD11cintCD11bint macrophages in the LP, which were recently shown to induce Foxp3+ regulatory T cells via IL-10 and TGF-β production [33]. Whereas iNOS+ DCs produce TNF-α, CD11chiCD11bhi LPDCs produce IL-6 and IL-12, but not TNF-α. Although iNOS+ DCs supply BAFF and APRIL, CD11chiCD11bhi LPDCs can not. By contrast, the expression of RALDH isoforms were ambiguous in iNOS+ DCs. Thus, CD11chiCD11bhi LPDCs and iNOS+ DCs have separate roles in IgA CSR and may work cooperatively with each other in the generation of IgA+ plasma cells (Fig. 5b) [18, 30].

iNOS+ DCs and CD11chiCD11bhi LPDCs. a Expression of surface molecules and phenotypes in iNOS+ DCs. b CD11chiCD11bhi LPDCs and iNOS+ DCs cooperatively act on naïve B cells to induce IgA+ plasma cells [30]
CD11chiCD11bhi LPDC-mediated CD4+ helper T cell (Th) response
CD11chiCD11bhi LPDCs are critical for the induction of Th responses as well as IgA production in the intestine [18, 33]. Th17 cells have recently emerged as a third T cell subset that produces IL-17, plays an essential role in protection against certain extracellular pathogens and induces inflammation and severe autoimmunity [34]. Differentiation of Th17 cells is initiated by TGF-β and IL-6 (Fig. 6) [35–37]. IL-6 signaling activates STAT3 and the lineage-determining transcription factor RORγt [38]. RORα, another member of the ROR family, is induced in a STAT3-dependent manner and may act synergistically with RORγt to induce Th17 cells [39]. IL-21 expression is induced in developing Th17 cells by IL-6. IL-21 acts autocrinely on Th17 cells to amplify this population [40, 41]. IL-23 serves to expand the previously differentiated Th17 cell population [35]. In humans, IL-1 has been reported to be involved in the generation of Th17 cells [42]. In addition, recent reports have shown that interferon regulatory factor (IRF) 4, Runt-related transcription factor 1 (Runx1) and aryl hydrocarbon receptor (AHR) are essential for Th17 cell differentiation [43–46]. Interestingly, Th17 cells preferentially exist in the intestinal LP of healthy mice [38]. Whereas conventional DCs induce Th1 cells in response to TLR ligands, CD11chiCD11bhi LPDCs induce RORγt+ functional Th17 cells as well as Th1 cells from naïve CD4+ T cells in response to flagellin in vitro. In addition, these DCs induce the generation of both Th17 cells and Th1 cells in an antigen (Ag)-specific manner in vivo. CD11chiCD11bhi LPDCs produce IL-6 in response to flagellin, and this process is essential for the induction of Th17 cell differentiation [18]. However, it remains unclear why CD11chiCD11bhi LPDCs, but not conventional DCs, have the ability to induce Th17 cells.

Differentiation of Th17 cells. Th17 cell differentiation is initiated by TGF-β and IL-6. IL-6 signaling activates Stat3 and the lineage-determining transcription factor RORγT. IL-21 is induced by developing Th17 cells in response to IL-6 and autocrinely acts on Th17 cells to amplify this population. IL-23 serves to expand the previously differentiated Th17 cell population
A series of recent studies showed that RA negatively regulates Th17 cell differentiation [47–49]. In every study, RA effectively inhibited in vitro differentiation of Th17 cells induced by IL-6 plus TGF-β in a dose-dependent manner. Although CD11chiCD11bhi LPDCs express RALDH2, they specifically induce Th17 cells [18]. Interestingly, the effect of RA on DC-mediated Th17 cell differentiation differs according to its concentration. RA at high concentration (10 μM) inhibits the differentiation of Th17 cells as well as Th1 cells, suggesting that high concentrations of RA inhibit both Th17 and Th1 cell differentiation induced by LPDCs. On the other hand, the RAR inhibitor LE540 inhibits the differentiation of Th17 cells, but not Th1 cells, suggesting that RA from LPDCs is necessary for Th17 cell differentiation. Interestingly, LPS-stimulated SPDCs can induce Th17 cells to the same extent as flagellin-stimulated LPDCs (R2) following the addition of RA at low concentration (1 nM). Furthermore, 10 μM RA abolished Th1 cell differentiation induced by LPS-stimulated SPDCs [18]. Thus, RA at low concentrations (1 nM) may have a positive effect on DC-mediated Th17 cell differentiation. According to a previous report, the plasma RA level is usually on the order of 10 nM [24]. It is well known that GALT DCs, but not DCs from other central lymphoid tissues, such as the spleen or peripheral lymph nodes (PLNs), produce RA, which upregulates expression of the chemokine receptor CCR9 and the integrin α4β7 on CD8+ precursors, Ag-specific CTLs, CD4+ T cells (Th1 and Th 2) and B cells [24, 25, 50–52]. Iwata et al [24] clearly showed that RA effectively enhanced α4β7 expression on CD8+ T cells even at concentrations in the range 0.1–1 nM. Because CD11chiCD11bhi LPDCs could induce B220−IgA+ plasma cells from naïve B cells and CCR9 expression on differentiated plasma cells effectively in vitro, the RA released from LPDCs is functional [18]. Although it is difficult to determine the local concentrations of retinoic acid secreted by CD11chiCD11bhi LPDCs, LPDC-derived RA acts as a positive regulator of Th17 cell differentiation. Thus, we have to reconsider the effects of RA on Th17 responses more cautiously.
CD11chiCD11bhi LPDCs and regulatory T cells
Recent reports show that CD103+ DCs in MLNs and LPs have a role in regulating immunity [53, 54]. CD103+ DCs migrate from the LP to MLNs in a CCR7-dependent manner and induce FoxP3+ regulatory T (Treg) cells via their derived RA [55, 56]. CD103+ DCs seem to be the same population as CD11chiCD11bhi LPDCs, because CD11chiCD11bhi LPDCs specifically express CD103 and RALDH2 [18]. CD11chiCD11bhi LPDCs may have opposite functions, inducing both immunological tolerance and protective immune responses. When CD11chiCD11bhi LPDCs take up food antigens under physiological conditions, they may induce immunological tolerance by promoting the development of T reg cells. When CD11chiCD11bhi DCs take up pathogenic bacteria during infection, they are activated and induced to maturate via innate immune receptors, leading to the activation of acquired immunity through the induction of proinflammatory cytokines and costimulatory molecules. Thus, CD11chiCD11bhi DCs may play critical roles in the decision to mount tolerant or protective immune responses in intestine by using innate immune receptors such as TLR5 as switches.
Conclusion
In this review, we have summarized the unique characteristics of TLR5-expressing LPDCs. These DCs induce innate immune responses by recognizing flagellated pathogenic bacteria via TLR5. They also work against bacterial infection by inducing ‘local’ IgA secretion and ‘systemic’ T helper responses through TLR5 stimulation (Fig. 7). Because IL-17 influences various kinds of cells to produce proinflammatory cytokines and induces the activation and migration of neutrophils, CD11chiCD11bhi LPDCs and Th17 cells may be involved in the pathogenesis of intestinal bowel diseases, such as Crohn’s disease. In addition, CD11chiCD11bhi LPDCs might be useful targets of mucosal vaccination based on their ability to induce Th1 responses and IgA synthesis. Future studies on CD11chiCD11bhi LPDCs will lead to new medical treatments.

Roles of CD11chiCD11bhi LPDCs in the small intestine. CD11chiCD11bhi LPDCs recognize invasive flagellated bacteria via a TLR5-dependent mechanism and induce innate immune responses. They also work against bacterial infection by inducing ‘local’ IgA secretion and ‘systemic’ T helper responses through TLR5 stimulation
References
Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801.
Macnab RM. Genetics and biogenesis of bacterial flagella. Annu Rev Genet. 1992;26:131–58.
Hayashi F, Smith KD, Ozinsky A, Hawn TR, Yi EC, Goodlett DR, et al. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature. 2001;410:1099–103.
Salazar-Gonzalez RM, McSorley SJ. Salmonella flagellin, a microbial target of the innate and adaptive immune system. Immunol Lett. 2005;101:117–22.
Gewirtz AT, Navas TA, Lyons S, Godowski PJ, Madara JL. Cutting edge: bacterial flagellin activates basolaterally expressed TLR5 to induce epithelial proinflammatory gene expression. J Immunol. 2001;167:1882–5.
Sierro F, Dubois B, Coste A, Kaiserlian D, Kraehenbuhl JP, Sirard JC. Flagellin stimulation of intestinal epithelial cells triggers CCL20-mediated migration of dendritic cells. Proc Natl Acad Sci USA. 2001;98:13722–7.
Sebastiani G, Leveque G, Lariviere L, Laroche L, Skamene E, Gros P, et al. Cloning and characterization of the murine toll-like receptor 5 (Tlr5) gene: sequence and mRNA expression studies in Salmonella-susceptible MOLF/Ei mice. Genomics. 2000;64:230–40.
Hawn TR, Verbon A, Lettinga KD, Zhao LP, Li SS, Laws RJ, et al. A common dominant TLR5 stop codon polymorphism abolishes flagellin signaling and is associated with susceptibility to legionnaires’ disease. J Exp Med. 2003;198:1563–72.
Uematsu S, Jang MH, Chevrier N, Guo Z, Kumagai Y, Yamamoto M, et al. Detection of pathogenic intestinal bacteria by Toll-like receptor 5 on intestinal CD11c+ lamina propria cells. Nat Immunol. 2006;7:868–74.
Kelsall BL, Leon F. Involvement of intestinal dendritic cells in oral tolerance, immunity to pathogens, and inflammatory bowel disease. Immunol Rev. 2005;206:132–48.
Iwasaki A, Kelsall BL. Freshly isolated Peyer’s patch, but not spleen, dendritic cells produce interleukin 10 and induce the differentiation of T helper type 2 cells. J Exp Med. 1999;190:229–39.
Andersen-Nissen E, Smith KD, Strobe KL, Barrett SL, Cookson BT, Logan SM, et al. Evasion of Toll-like receptor 5 by flagellated bacteria. Proc Natl Acad Sci USA. 2005;102:9247–52.
Niedergang F, Didierlaurent A, Kraehenbuhl JP, Sirard JC. Dendritic cells: the host Achille’s heel for mucosal pathogens? Trends Microbiol. 2004;12:79–88.
Hopkins SA, Niedergang F, Corthesy-Theulaz IE, Kraehenbuhl JP. A recombinant Salmonella typhimurium vaccine strain is taken up and survives within murine Peyer’s patch dendritic cells. Cell Microbiol. 2000;2:59–68.
Rescigno M, Urbano M, Valzasina B, Francolini M, Rotta G, Bonasio R, et al. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat Immunol. 2001;2:361–7.
Patel JC, Rossanese OW, Galan JE. The functional interface between Salmonella and its host cell: opportunities for therapeutic intervention. Trends Pharmacol Sci. 2005;26:564–70.
Vazquez-Torres A, Jones-Carson J, Baumler AJ, Falkow S, Valdivia R, Brown W, et al. Extraintestinal dissemination of Salmonella by CD18-expressing phagocytes. Nature. 1999;401:804–8.
Uematsu S, Fujimoto K, Jang MH, Yang BG, Jung YJ, Nishiyama M, et al. Regulation of humoral and cellular gut immunity by lamina propria dendritic cells expressing Toll-like receptor 5. Nat Immunol. 2008;9:769–76.
van Egmond M, Damen CA, van Spriel AB, Vidarsson G, van Garderen E, van de Winkel JG. IgA and the IgA Fc receptor. Trends Immunol. 2001;22:205–11.
Suzuki K, Ha SA, Tsuji M, Fagarasan S. Intestinal IgA synthesis: a primitive form of adaptive immunity that regulates microbial communities in the gut. Semin Immunol. 2007;19:127–35.
Tseng J. A population of resting IgM-IgD double-bearing lymphocytes in Peyer’s patches: the major precursor cells for IgA plasma cells in the gut lamina propria. J Immunol. 1984;132:2730–5.
Tseng J. Transfer of lymphocytes of Peyer’s patches between immunoglobulin allotype congenic mice: repopulation of the IgA plasma cells in the gut lamina propria. J Immunol. 1981;127:2039–43.
Tsuji M, Komatsu N, Kawamoto S, Suzuki K, Kanagawa O, Honjo T, et al. Preferential generation of follicular B helper T cells from Foxp3+ T cells in gut Peyer’s patches. Science. 2009;323:1488–92.
Iwata M, Hirakiyama A, Eshima Y, Kagechika H, Kato C, Song SY. Retinoic acid imprints gut-homing specificity on T cells. Immunity. 2004;21:527–38.
Mora JR, Iwata M, Eksteen B, Song SY, Junt T, Senman B, et al. Generation of gut-homing IgA-secreting B cells by intestinal dendritic cells. Science. 2006;314:1157–60.
Fagarasan S, Kinoshita K, Muramatsu M, Ikuta K, Honjo T. In situ class switching and differentiation to IgA-producing cells in the gut lamina propria. Nature. 2001;413:639–43.
Kunisawa J, Kurashima Y, Gohda M, Higuchi M, Ishikawa I, Miura F, et al. Sphingosine 1-phosphate regulates peritoneal B-cell trafficking for subsequent intestinal IgA production. Blood. 2007;109:3749–56.
Macpherson AJ, Harris NL. Interactions between commensal intestinal bacteria and the immune system. Nat Rev Immunol. 2004;4:478–85.
Niess JH, Brand S, Gu X, Landsman L, Jung S, McCormick BA, et al. CX3CR1-mediated dendritic cell access to the intestinal lumen and bacterial clearance. Science. 2005;307:254–8.
Tezuka H, Abe Y, Iwata M, Takeuchi H, Ishikawa H, Matsushita M, et al. Regulation of IgA production by naturally occurring TNF/iNOS-producing dendritic cells. Nature. 2007;448:929–33.
Serbina NV, Pamer EG. Monocyte emigration from bone marrow during bacterial infection requires signals mediated by chemokine receptor CCR2. Nat Immunol. 2006;7:311–7.
Serbina NV, Salazar-Mather TP, Biron CA, Kuziel WA, Pamer EG. TNF/iNOS-producing dendritic cells mediate innate immune defense against bacterial infection. Immunity. 2003;19:59–70.
Denning TL, Wang YC, Patel SR, Williams IR, Pulendran B. Lamina propria macrophages and dendritic cells differentially induce regulatory and interleukin 17-producing T cell responses. Nat Immunol. 2007;8:1086–94.
Korn T, Bettelli E, Oukka M, Kuchroo VK. IL-17 and Th17 cells. Annu Rev Immunol. 2009;27:485–517.
Veldhoen M, Hocking RJ, Atkins CJ, Locksley RM, Stockinger B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity. 2006;24:179–89.
Bettelli E, Carrier Y, Gao W, et al. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature. 2006;441:235–8.
Mangan PR, Harrington LE, O’Quinn DB, Helms WS, Bullard DC, Elson CO, et al. Transforming growth factor-beta induces development of the T(H)17 lineage. Nature. 2006;441:231–4.
Ivanov II, McKenzie BS, Zhou L, Tadokoro CE, Lepelley A, Lafaille JJ, et al. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell. 2006;126:1121–33.
Yang XO, Pappu BP, Nurieva R, Akimzhanov A, Kang HS, Chung Y, et al. T helper 17 lineage differentiation is programmed by orphan nuclear receptors ROR alpha and ROR gamma. Immunity. 2008;28:29–39.
Korn T, Bettelli E, Gao W, Awasthi A, Jager A, Strom TB, et al. IL-21 initiates an alternative pathway to induce proinflammatory T(H)17 cells. Nature. 2007;448:484–7.
Zhou L, Ivanov II, Spolski R, Min R, Shenderov K, Egawa T, et al. IL-6 programs T(H)-17 cell differentiation by promoting sequential engagement of the IL-21 and IL-23 pathways. Nat Immunol. 2007;8:967–74.
Acosta-Rodriguez EV, Napolitani G, Lanzavecchia A, Sallusto F. Interleukins 1beta and 6 but not transforming growth factor-beta are essential for the differentiation of interleukin 17-producing human T helper cells. Nat Immunol. 2007;8:942–9.
Brustle A, Heink S, Huber M, Rosenplanter C, Stadelmann C, Yu P, et al. The development of inflammatory T(H)-17 cells requires interferon-regulatory factor 4. Nat Immunol. 2007;8:958–66.
Zhang F, Meng G, Strober W. Interactions among the transcription factors Runx1, RORgammat and Foxp3 regulate the differentiation of interleukin 17-producing T cells. Nat Immunol. 2008;9:1297–306.
Veldhoen M, Hirota K, Westendorf AM, Buer J, Dumoutier L, Renauld JC, et al. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature. 2008;453:106–9.
Quintana FJ, Basso AS, Iglesias AH, Korn T, Farez MF, Bettelli E, et al. Control of T(reg) and T(H)17 cell differentiation by the aryl hydrocarbon receptor. Nature. 2008;453:65–71.
Mucida D, Park Y, Kim G, Turovskaya O, Scott I, Kronenberg M, et al. Reciprocal TH17 and regulatory T cell differentiation mediated by retinoic acid. Science. 2007;317:256–60.
Elias KM, Laurence A, Davidson TS, Stephens G, Kanno Y, Shevach EM, et al. Retinoic acid inhibits Th17 polarization and enhances FoxP3 expression through a Stat-3/Stat-5 independent signaling pathway. Blood. 2008;111:1013–20.
Schambach F, Schupp M, Lazar MA, Reiner SL. Activation of retinoic acid receptor-alpha favours regulatory T cell induction at the expense of IL-17-secreting T helper cell differentiation. Eur J Immunol. 2007;37:2396–9.
Mora JR, Bono MR, Manjunath N, Weninger W, Cavanagh LL, Rosemblatt M, et al. Selective imprinting of gut-homing T cells by Peyer’s patch dendritic cells. Nature. 2003;424:88–93.
Johansson-Lindbom B, Svensson M, Pabst O, Palmqvist C, Marquez G, Forster R, et al. Functional specialization of gut CD103+ dendritic cells in the regulation of tissue-selective T cell homing. J Exp Med. 2005;202:1063–73.
Johansson-Lindbom B, Svensson M, Wurbel MA, Malissen B, Marquez G, Agace W. Selective generation of gut tropic T cells in gut-associated lymphoid tissue (GALT): requirement for GALT dendritic cells and adjuvant. J Exp Med. 2003;198:963–9.
Annacker O, Coombes JL, Malmstrom V, Uhlig HH, Bourne T, Johansson-Lindbom B, et al. Essential role for CD103 in the T cell-mediated regulation of experimental colitis. J Exp Med. 2005;202:1051–61.
Sun CM, Hall JA, Blank RB, Bouladoux N, Oukka M, Mora JR, et al. Small intestine lamina propria dendritic cells promote de novo generation of Foxp3 T reg cells via retinoic acid. J Exp Med. 2007;204:1775–85.
Jang MH, Sougawa N, Tanaka T, Hirata T, Hiroi T, Tohya K, et al. CCR7 is critically important for migration of dendritic cells in intestinal lamina propria to mesenteric lymph nodes. J Immunol. 2006;176:803–10.
Coombes JL, Siddiqui KR, Arancibia-Carcamo CV, Hall J, Sun CM, Belkaid Y, et al. A functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-beta and retinoic acid-dependent mechanism. J Exp Med. 2007;204:1757–64.
Acknowledgments
We thank our colleagues in our lab for helpful discussion and E. Kamada for secretarial assistance. This work is supported in part by grants from the Special Coordination Funds of the Japanese Ministry of Education, Culture, Sports, Science and Technology.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Uematsu, S., Akira, S. Immune responses of TLR5+ lamina propria dendritic cells in enterobacterial infection. J Gastroenterol 44, 803–811 (2009). https://doi.org/10.1007/s00535-009-0094-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00535-009-0094-y