
Abstract
This paper examines if shell oxygen isotope ratios (δ18Oar) of Unio sp. can be used as a proxy of past discharge of the river Meuse. The proxy was developed from a modern dataset for the reference time interval 1997–2007, which showed a logarithmic relationship between discharge and measured water oxygen isotope ratios (δ18Ow). To test this relationship for past time intervals, δ18Oar values were measured in the aragonite of the growth increments of four Unio sp. shells; two from a relatively wet period and two from a very dry time interval (1910–1918 and 1969–1977, respectively). Shell δ18Oar records were converted into δ18Ow values using existing water temperature records. Summer δ18Ow values, reconstructed from δ18Oar of 1910–1918, showed a similar range as the summer δ18Ow values for the reference time interval 1997–2007, whilst summer reconstructed δ18Ow values for the time interval 1969–1977 were anomalously high. These high δ18Ow values suggest that the river Meuse experienced severe summer droughts during the latter time interval. δ18Ow values were then applied to calculate discharge values. It was attempted to estimate discharge from the reconstructed δ18Ow values using the logarithmic relationship between δ18Ow and discharge. A comparison of the calculated summer discharge results with observed discharge data showed that Meuse low-discharge events below a threshold value of 6 m3/s can be detected in the reconstructed δ18Ow records, but true quantification remains problematic.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Meuse is one of the major rivers in the Netherlands. It is a rain-fed river characterised by a pronounced rainfall-evaporation regime causing low discharge in summer and high discharge in winter (De Wit et al. 2007). Its 33,000 km2 basin drains the northeast of France and eastern Belgium. The average discharge at Borgharen (Fig. 1) is 274 m3/s; highest peak discharges exceed 3000 m3/s and low-flow events can be less than 4 m3/s. Floods (e.g. 1993 and 1995) and droughts (e.g. 1976 and 2003) do occur, and both are expected to become more frequent due to an increase in precipitation extremes caused by climate change (Bürger 2002; Gregory et al. 1997; Pfister et al. 2004; Ward et al. 2008). Large flood events mainly occur during the winter season when freshwater mussels do not grow. Severe drought in the Meuse hinterland generally occurs during the summer-autumn time interval, which results in low Meuse river discharge. These droughts limit water availability for many purposes, including agriculture and the cooling of power plants. In addition, water quality deteriorates during episodes of droughts, threatening drinking water supplies and impacting on river ecology (Van Vliet and Zwolsman 2008). Reduced discharge, and the associated changes in water chemistry, can potentially be recorded by freshwater bivalve shells.

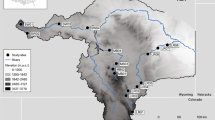
Map of the river Meuse in the Netherlands. Shell collection sites are black dots; Rijkswaterstaat gauging stations are circles. (Made with Online Map Creation: http://www.aquarius.geomar.de/)
In order to improve our understanding of river dynamics and to predict impacts of future climate change, it is important to gain insight into past discharge variability and the frequencies of floods and droughts. The instrumental Meuse discharge record only goes back to the early twentieth century. Therefore, it is crucial to develop proxies for river discharge prior to the period of instrumental record.
A likely candidate for such a proxy is the stable oxygen isotope ratio of the shells of unionid freshwater bivalves growing in rivers. These records have been previously demonstrated to be a useful proxy for past rainfall patterns, water source, or river discharge (Gajurel et al. 2006; Kaandorp et al. 2005; Ricken et al. 2003). In this study, the possibilities and limitations of using unionid aragonite oxygen isotope ratios (δ18Oar values) as a proxy for past river discharge in the river Meuse are investigated. Shell aragonite is precipitated in annual growth increments, which are clearly visible in dorso-ventral sections of the shell. Shell aragonite of many freshwater bivalve species is deposited in oxygen isotopic equilibrium with the ambient water (Dettman et al. 1999; Goewert et al. 2007; Kaandorp et al. 2003). Analysing growth increments at high-spatial resolution can thus reveal seasonal patterns in δ18Oar. Unionids cease growing below a certain temperature threshold (Dettman et al. 1999; Dunca and Mutvei 2001; Goewert et al. 2007), which is ~12–13°C for the species used here (Negus 1966; Versteegh 2009). Therefore, only summer conditions can be recorded in the shell, and winters are represented by hiatuses.
It has previously been established that freshwater bivalve δ18Oar values can be used as a proxy for past water compositions during summer by showing that aragonite in modern shells is precipitated in isotopic equilibrium with the ambient water (Dettman et al. 1999; Dettman et al. 2004; Goewert et al. 2007). However, no attempt has yet been made to calculate water δ18O (δ18Ow) values from δ18Oar values with the aim of reconstructing river discharges. Therefore, this study aims to: (1) investigate whether δ18Ow values can be used as a proxy for discharge using a dataset of measured δ18Ow values and discharge values from the time interval 1997–2007; (2) reconstruct δ18Ow values for a wet time interval (1910–1918) and a very dry time interval (1969–1977) on the basis of molluscan δ18Oar records; (3) use reconstructed δ18Ow (δ18Owr) values to estimate river discharge values and to compare these with measured river discharge during the same two selected time intervals.
Materials and methods
Freshwater mussels
Freshwater mussels of the genus Unio are abundant in freshwater bodies in the Netherlands. They form shells up to 12.5 cm in length and can live for up to 15 years (Gittenberger et al. 1998). Three species are studied here: U. crassus, which became extinct in the Netherlands in 1968 (Gittenberger et al. 1998); U. Pictorum; and U. tumidus.
Shell collection
Four twentieth-century shells from the river Meuse were taken from museum and private collections. The mussels were all collected alive, and therefore these shells can be temporally aligned with the available instrumental records of water temperature and discharge of the river Meuse. Two sets of shells were collected in different locations: two specimens were collected in 1918 in Beegden and Bergen; and two specimens were collected in 1977 in Vierlingsbeek and Waalwijk. For specifications of all specimens and collection sites, the reader is referred to Fig. 1 and Table 1. Unfortunately, any shells collected during the reference time interval (1997–2007) proved unsuitable for further analysis, because they originated not from the river itself, but a connected lake, or growth increments were too narrow to be identified up to the ventral margin.
Collection of river-data
Water temperature and discharge variability (river discharge: Fig. 2) have been measured since the beginning of the twentieth century by Rijkswaterstaat (Dutch Directorate for Public Works and Water Management; http://www.waterbase.nl/). Temperature data for the δ18Ow reconstruction were taken from the gauging stations closest to the collection sites. For the Bergen 1918 shell, the closest gauging station is Borgharen; for the Vierlingsbeek 1977 shell this is Sambeek; and for the Waalwijk 1977 shell temperature data from Heesbeen were used (Fig. 1). Discharge data were used from the gauging station at Borgharen.

Discharge (Q) of the Meuse for the time interval November 1911 to Dec 2007. Considerable long-term variation in both maxima and minima is visible. Blown-up figures show discharges for the time intervals in which the shells grew. The time interval 1912–1918 has several high-discharge summers (e.g. 1916 and 1917), whereas the time interval 1969–1977 generally has very dry summers and exhibits several severe summer droughts (e.g. 1971 and 1976)
The only continuous multi-year δ18Ow record for the river Meuse has been measured at Eijsden during the time interval 1990–2007 (Fig. 1). This dataset was obtained from the Centre for Isotope Research, University of Groningen (Fig. 3). Comparison of this dataset with Meuse δ18Ow values measured at Lith, which lies ~ 160 km downstream from Eijsden, during one year (June 2006-July 2007), showed that δ18Ow values measured at Eijsden and Lith during the same time period correlate well (Versteegh 2009).

Discharge (Q) measured at Borgharen and seasonal δ18Ow values sampled at Eijsden (raw data in grey and re-sampled data (simple cubic spline, 30 day resolution) in black) for the river Meuse during the time interval 1997–2007
Sampling and analysis of shells
Shells were embedded in epoxy resin and sections of 300 μm were cut perpendicular to the growth lines, along the dorso-ventral axis of the shell (Fig. 1a–b). The nacreous layer of the shell was sampled with a Merchantek Micromill microsampler. Drill bit diameter was ~ 800 μm and sampling resolution was 100–500 μm corresponding to a time span of between 6 days and > 2 months, depending on growth rate. Drilling depth was ~ 250 μm. Samples were analysed for δ18Oar values either on a Finnigan MAT 252 mass spectrometer equipped with a Kiel II device or a Finnigan Delta + mass spectrometer with a GasBench II. On both systems, the long-term standard deviation of a routinely analysed in-house CaCO3 standard was < 0.1 ‰. This CaCO3 standard is regularly calibrated to NBS 18, 19, and 20 (National Institute of Standards and Technology). Typical sample size for the MAT 252 system lies at 10–20 μg. For the Delta + system, samples of 20–50 μg are required. Occasional duplicate analyses confirmed that these two systems gave comparable results.
Calculation of δ18Owr values
For the calculations of δ18Owr, the equation of Grossman and Ku (1986) as modified by Dettman et al. (1999) was used:
where T is the water temperature in degrees Kelvin and α is the fractionation between water and aragonite described by:
where ar is shell aragonite and w is water.
δ18Ow values are reported relative to VSMOW, whereas δ18Oar values are reported relative to VPDB (Coplen 1996). δ18Oar (VSMOW) values were converted to δ18Oar (VPDB) using the equation of Gonfiantini et al. (1995):
Results
River data
The complete discharge record from Borgharen, covering the time interval November 1911–2007, is shown in Fig. 2. Both the maximum discharge values and the minimum discharge values show marked seasonal dry and wet intervals. The shells studied here grew in either an interval with relatively high summer discharge values (1910–1918) or a time interval with exceptionally low-discharge values (1969–1977). These two time intervals are shown in the insets of Fig. 2.
To facilitate qualitative comparison with the δ18Owr output, the mean, minimum and maximum discharges for the shell growing season (April–October; Versteegh 2009) of each year were calculated. Subsequently, the years were classified into five discharge classes by applying k-means cluster analysis on the natural logarithm of these values. These classes were labelled “very high”, “high”, “normal”, “low” and “very low” (Fig. 4). The two lowest minimum discharge classes (“low” and “very low”) represent summers in which the minimum discharge was ≤ 6 m3/s. For the time intervals 1912–1918 and 1969–1977, this classification is shown in Fig. 4.

Discharges categorised in five classes with respect to mean, minimum and maximum discharges during the season April–October 1912–1918 and 1969–1977 at Borgharen. For 1910 and 1911, no discharge data were available
From the discharge record, it appears that discharges were high during the time interval 1910–1918. In the years 1916, 1917 and 1918, the Meuse experienced very high mean discharges and very high minimum discharges; the latter was also the case for the years 1913, 1914 and 1915 (Fig. 4). In contrast, during the time interval 1969–1977, the Meuse experienced several extremely dry summers with minima ≤ 2 m3/s: 1969, 1971, 1973, 1974, 1976 and 1977, whereby during the year 1976 the seasonal mean and maximum was also very low. The year 1976 was exceptionally dry and hot (Können and Fransen 1996) and had the longest time interval with extremely low discharges of the entire record for 1912–2007 (Fig. 4). The reference time interval for which δ18Ow values are available (1997–2007) is mainly characterised by low mean summer discharges, with average minimum discharges. The summer of 2003 was the second driest in the 1912–2007 record. This resulted in very low discharge of the Meuse. In spite of this, both discharge and δ18Ow values have a sufficient range to calculate their relationship from this record (see discussion).
Measured δ18Oar values in shells
The δ18Oar records of the shells show the truncated sinusoidal pattern typical for seasonal growth (Fig. 5a–d). One year of growth is represented by a wide summer trough and a narrow positive peak. This narrow peak corresponds to δ18Oar values precipitated during slow growth just prior to and shortly after the winter growth cessation (Dettman et al. 1999; Goodwin et al. 2003; Versteegh et al. 2009). In these shells, δ18Oar values vary between −4.7 and −7.5 ‰ (VPDB), which is representative for the river Meuse (Verdegaal et al. 2005; Versteegh et al. 2009). The total number of growing seasons recorded in the shells varies between two in Waalwijk 1977 (Fig. 5d) and over 14 in Beegden 1918 (Fig. 5a).

δ18Oar records of the four shells: a Beegden 1918 (Unio tumidus), b Bergen 1918 (U. crassus), c Vierlingsbeek 1977 (U. pictorum), d Waalwijk 1977 (U. pictorum). All shells show a truncated seasonal pattern with sharp upward pointing peaks reflecting winter growth stops and low values reflecting summer growth seasons. δ18Oar data of the Bergen 1918 shell have previously been published by Verdegaal et al. (2005)
Discussion
In order to test the applicability of unionid δ18Oar values as a proxy for river δ18Ow values and, ultimately, discharge, it is necessary to first determine if there is a detectable relationship between these two variables. Subsequently, river δ18Ow values during the lifetime of the shell are reconstructed and finally an estimation river discharge is attempted.
Empirical relationship between δ18Ow values and discharge
The measured δ18Ow record for the period 1990–2007 contains several hiatuses during the years 1990–1997, which inhibit a comprehensive comparison of the relationship between δ18Ow and discharge. However, the time interval 1997–2007 is suitable for such an exercise. The focus therefore lies on the interval 1997–2007, for which the data are complete, to model the relationship between δ18Ow and discharge values. The 1997–2007 record shows that the Meuse had an average δ18Ow value of −7.1 ‰ (VSMOW) with summer maxima of −6.0 to −6.5 ‰ (VSMOW) and winter minima of −7.7 to −8.4 ‰ (VSMOW; Fig. 3). Some extreme high and low summer discharge events can be recognised in the δ18Ow record: the relatively high discharge time intervals during the summers of 2000 and 2001 coincided with the two lowest summer δ18Ow peaks, and the very dry summer of 2003 resulted in extremely high peak δ18Ow values (Fig. 3).
To evaluate whether a quantifiable relationship between δ18Ow and discharge exists, these variables were used for constructing a correlation diagram (Fig. 6). The relationship between δ18Ow and discharge (Q) is logarithmic, because the differences between mean and minimum and mean and maximum discharge within one year are both about an order of magnitude. In addition, discharge cannot have a negative value. The logarithmic relationship has the following equation (full data set):

Correlation of discharge (Q) and δ18Ow values for the Meuse during the years 1997 to 2007 for all months (circles) with a logarithmic fit (light grey line) with equation: \( \delta^{ 1 8} {\text{O}}_{\text{w}} = - 0.315 \cdot \ln Q - 5.592 \) (R 2 = 0.60; p < 0.0005) and for the growing season of the shells (squares) with a logarithmic fit (dark grey line) with equation \( \delta^{ 1 8} {\text{O}}_{\text{w}} = - 0.291 \cdot \ln Q - 5.659 \) (R 2 = 0.44; p < 0.0005)
This relationship is particularly interesting for the summer time interval, since the mussels studied precipitate their shells only during summer (April–October; Negus 1966; Versteegh 2009). For the summer data, this relationship has the following equation:
The logarithmic regressions approximate the data points well using the full dataset or only the summer data (for statistics of linear and quadratic regressions see Table 2). However, the variance of data points is ~ 1 ‰ for a given discharge value. The data used to construct Fig. 6 have a minimum discharge value of 9 m3/s and a maximum of 842 m3/s (during this time interval lower discharges (≥ 5.7 m3/s) occurred, but not on the dates that δ18Ow measurements were taken, whereas the total discharge variation during the summers from November 1911 to 2007 ranged from < 2 to 2000 m3/s. Because the relationship between discharge and δ18Ow is logarithmic, δ18Ow values will only differ slightly between the normal to extremely high summer discharge situations. Therefore, it is not possible to detect and reconstruct high summer discharges reliably. In the low to extremely low-discharge situations, however, δ18Ow values will show a significant shift towards higher values, enabling detection of summer droughts in the river Meuse (Fig. 6).
Reconstructed δ18Ow records
For reconstruction of δ18Ow values and subsequent linking of the δ18Owr patterns to known river discharge variation, first a calendar year needed to be assigned to each individual growing season in the shell δ18Oar records. Due to the ontogenetic decrease in the growth rate, growth increments were narrower in the adult shell and in some cases could not be resolved with the Micromill sampling technique used here. This was the case for the Beegden 1918 shell (Fig. 5a). In this shell, the growth seasons could not be identified up to the ventral margin and thus calendar years could not be assigned to the seasons. Therefore, this shell was excluded from the δ18Ow reconstructions. The other three shells exhibited high growth rates throughout their lives, which made them useful for the δ18Ow reconstructions presented here.
δ18Ow values were calculated from measured δ18Oar values and water temperatures, using Eqs. 1–3. It has previously been established that growth starts when water temperature rises above 13.5°C and ceases when water temperature falls below that temperature (Versteegh 2009). These temperatures were used to determine the dates of onset and cessation of growth for each year. It is likely that intra-seasonal growth is non-linear. However, since no robust growth model is available yet for these species, intra-seasonal growth was assumed to be linear. In order to clearly visualise the range of δ18Owr values, these data were plotted per season by means of a box-whisker diagram (Fig. 7a–c). From the 1997–2007 δ18Ow dataset, measurements of the months April to October (the growth season of the shells) were selected. The 2, 5, 25, 75, 95, and 98% percentiles of these data were calculated (Fig. 7a–c), in order to compare instrumental δ18Ow data with the δ18Owr values.

Box-whisker diagrams showing reconstructed δ18Ow records per season for three shells from the Meuse. Colours of the boxes indicate if discharge was very high (dark blue), high (light blue), normal (white), low (light yellow) or very low (dark yellow) (see Fig. 4). In the background, dark to light grey areas indicate 2, 5, 25, 75, 95 and 98% percentiles of δ18Ow values during the growing season (April–October) of the 1997–2007 record. a Bergen 1918 (Unio crassus), b Vierlingsbeek 1977 (U. pictorum), c Waalwijk 1977 (U. pictorum)
Molluscan δ18Oar as a recorder of low summer discharge
As described earlier, low summer discharges can be recognised in river δ18Ow values and are potentially recorded by unionids; this is not the case for high summer discharges. The focus therefore lies on the low to extremely low-discharge events that occurred during the lifetime of the 1998 and 1977 shells (the 2 lowest classes in the minimum discharge record).
In both the Vierlingsbeek 1977 and the Waalwijk 1977, the majority of δ18Owr values range above the 95% percentile of 1997–2007 data (representing 5% highest values), and in 1975, 1976 and 1977 δ18Owr values even range above the 98% percentile of 1997–2007 (representing 2% highest values) (Fig. 7b–c). This suggests that δ18Owr values from unionid shells are a useful proxy for summer low-discharge events. However, the proxy does not appear to work in all years: during 1972 δ18Owr values range above the 95% percentile, whereas minimum discharge was normal during that year; the year 1974 did have a very low minimum discharge, but δ18Owr values do not range above the 95% percentile (Fig. 7b). Possible causes for the failure to detect low-discharge events can be: (1) this low-discharge summer was not accompanied by high δ18Ow values in the river or (2) the shell experienced a temporary growth shutdown due to an environmental disturbance. If this type of temporary growth shutdown is indeed the case, and occurs often, this poses a serious problem for the reliability of the proxy. Erroneous recording of a low-discharge season might happen when a shell grew in an environment relatively isolated from the riverbed, experiencing a large influence of local evaporation. The use of a larger set of shells would enable us to distinguish between these possibilities.
Quantitative reconstruction of summer discharge
To investigate if past Meuse discharges can be quantified, reconstructed discharges were calculated using Eq. 5. Because of the focus on the low-discharge situation, reconstructed and measured discharges are plotted on a logarithmic scale (Fig. 8a–c). Correlations between measured and reconstructed discharge are very poor in all three shells (Table 3). Visual comparison in Fig. 8 confirms that high-discharge events cannot be reconstructed. In the cases of the years 1917 and 1918 reconstructed discharge values are much higher than ever recorded in the Meuse. The quantitative reconstruction of low-discharge appears accurate in the years 1970, 1972 and 1973 in the Vierlingsbeek shell. The other dry summers appear to show the correct lower boundary of discharge values but not with the correct timing (e.g. 1969 and 1976 in the Vierlingsbeek shell and 1976 and 1977 in the Waalwijk shell). This timing discrepancy probably occurs because of the assumption of linear intra-seasonal growth, which is not likely for Unionidae (Howard 1922; Negus 1966; Versteegh 2009). Another complicating factor is that there is no absolute certainty about the assignment of calendar years to parts of the shell δ18Oar records. The reconstruction and true quantification of low-discharge events in the Meuse remains problematic for two main reasons: the variance of the discharge-δ18Ow relationship is considerable, because multiple factors (such as source area of the precipitation or local evaporation) may play a role; and δ18Oar values might not always reflect δ18Ow values in the main river channel, because local habitats may differ in, for example, their connection to the river or the influence of evaporation. Sampling a larger number of shells would possibly clarify this latter issue.

a–c Reconstructed (grey) and actual measured discharges (black line) per season. High-discharge summers cannot accurately be reconstructed and result in large errors (e.g. 1917 and 1918 in the Bergen shell). All low-discharge events in the 1969 to 1977 time interval can be reconstructed from shell δ18Oar values. The timing of these events is not always accurate; this is due to the linear intra-seasonal age model used here
Conclusions
Due to global warming, extreme precipitation events are expected to become more frequent in the Meuse basin, probably leading to extremely low and high discharges of this river. In order to better predict impacts of future climate change, knowledge of the past is essential. Therefore, the utility of freshwater mussel δ18Oar values as a proxy for past δ18Ow and extreme discharges of the river Meuse was investigated.
It has been found that in the Meuse there is a logarithmic relationship between discharge and δ18Ow values. Furthermore, unionid freshwater mussels recorded ambient δ18Ow values in the δ18Oar values of growth increments in their shells, suggesting that past δ18Ow values and Meuse discharge can be reconstructed.
However, due to the logarithmic relationship between discharge and δ18Ow values, only low-discharge summers could be detected qualitatively. Meuse low-discharge events below a threshold value of 6 m3/s could be detected in the δ18Owr records. True quantification of summer discharges was complicated by noise in both the relationship between discharge and δ18Ow values, and between unionid δ18Oar values and those of the river water. Quantitative reconstructions of past δ18Ow values and Meuse discharge might be realised by analysing many more samples (e.g. 30) from the same time interval than the three specimens presented here.
References
Bürger G (2002) Selected precipitation scenarios across Europe. J Hydrol 262(1–4):99–110
Coplen TB (1996) New guidelines for reporting stable hydrogen, carbon, and oxygen isotope-ratio data. Geochim Cosmochim Acta 60(17):3359–3360
De Wit M, Van den Hurk B, Warmerdam P, Torfs P, Roulin E, Van Deursen W (2007) Impact of climate change on low-flows in the river Meuse. Clim Change 82(3):351–372
Dettman DL, Reische AK, Lohmann KC (1999) Controls on the stable isotope composition of seasonal growth bands in aragonitic fresh-water bivalves (Unionidae). Geochim Cosmochim Acta 63(7–8):1049–1057
Dettman DL, Flessa KW, Roopnarine PD, Schone BR, Goodwin DH (2004) The use of oxygen isotope variation in shells of estuarine mollusks as a quantitative record of seasonal and annual Colorado river discharge. Geochim Cosmochim Acta 68(6):1253–1263
Dunca E, Mutvei H (2001) Comparison of microgrowth pattern in Margaritifera margaritifera shells from south and north Sweden. Am Malacol Bull 16(1/2):239–250
Gajurel AP, France-Lanord C, Huyghe P, Guilmette C, Gurung D (2006) C and O isotope compositions of modern fresh-water mollusc shells and river waters from the Himalaya and Ganga plain. Chem Geol 233(1–2):156–183
Gittenberger E, Janssen AW, Kuijper WJ, Kuiper JGJ, Meijer T, van der Velde G, de Vries JN (1998) De Nederlandse zoetwatermollusken - Recente en fossiele weekdieren uit zoet en brak water. Nationaal Natuurhistorisch Museum Naturalis - KNNV Uitgeverij, Leiden
Goewert A, Surge D, Carpenter SJ, Downing J (2007) Oxygen and carbon isotope ratios of Lampsilis cardium (Unionidae) from two streams in agricultural watersheds of Iowa, USA. Palaeogeogr Palaeoclimatol Palaeoecol 252(3-4):637–648
Gonfiantini R, Stichler W, Rozanski K (1995) Standards and intercomparison materials distributed by the International Atomic Energy Agency for stable isotope measurements. I.A.E.A.
Goodwin DH, Schöne BR, Dettman DL (2003) Resolution and fidelity of oxygen isotopes as paleotemperature proxies in bivalve mollusk shells: models and observations. Palaios 18(2):110–125
Gregory JM, Mitchell JFB, Brady AJ (1997) Summer drought in northern midlatitudes in a time-dependent CO2 climate experiment.). pp 662–686
Grossman EL, Ku T-L (1986) Oxygen and carbon isotope fractionation in biogenic aragonite: temperature effects. Chem Geol Isotope Geosci Sect 59:59–74
Howard AD (1922) Experiments in the culture of fresh-water mussels. Bull Bureau Fish 38:63–89
Kaandorp RJG, Vonhof HB, Del Busto C, Wesselingh FP, Ganssen GM, Marmol AE, Romero Pittman L, van Hinte JE (2003) Seasonal stable isotope variations of the modern Amazonian freshwater bivalve Anodontites trapesialis. Palaeogeogr Palaeoclimatol Palaeoecol 194(4):339–354
Kaandorp RJG, Vonhof HB, Wesselingh FP, Pittman LR, Kroon D, van Hinte JE (2005) Seasonal Amazonian rainfall variation in the Miocene Climate Optimum. Palaeogeogr Palaeoclimatol Palaeoecol 221(1–2):1–6
Können GP, Fransen W (1996) De toestand van het klimaat in Nederland 1996. KNMI, De Bilt
Negus CL (1966) A quantitative study of growth and production of unionid mussels in the river Thames at Reading. J Anim Ecol 35(3):513–532
Pfister L, Kwadijk J, Musy A, Bronstert A, Hoffmann L (2004) Climate change, land use change and runoff prediction in the Rhine-Meuse basins. River Res Appl 20(3):229–241
Ricken W, Steuber T, Freitag H, Hirschfeld M, Niedenzu B (2003) Recent and historical discharge of a large European river system - oxygen isotopic composition of river water and skeletal aragonite of Unionidae in the Rhine. Palaeogeogr Palaeoclimatol Palaeoecol 193(1):73–86
Van Vliet MTH, Zwolsman JJG (2008) Impact of summer droughts on the water quality of the Meuse river. J Hydrol 353(1–2):1–17
Verdegaal S, Troelstra SR, Beets CKJ, Vonhof HB (2005) Stable isotopic records in unionid shells as a paleoenvironmental tool. Neth J Geosci 84(4):403–408
Versteegh EAA (2009) Silent witnesses—freshwater bivalves as archives of environmental variability in the Rhine-Meuse delta. Dissertation, VU University
Versteegh EAA, Troelstra SR, Vonhof HB, Kroon D (2009) Oxygen isotopic composition of bivalve seasonal growth increments and ambient water in the rivers Rhine and Meuse. Palaios 24(8):497–504. doi:10.2110/palo.2008.p08-071r
Ward PJ, Renssen H, Aerts JCJH, van Balen RT, Vandenberghe J (2008) Strong increases in flood frequency and discharge of the River Meuse over the late Holocene: impacts of long-term anthropogenic land use change and climate variability. HESS 12(1):159–175
Acknowledgments
This research was supported by BSIK-Climate changes Spatial Planning. We would like to thank Rob Molenbeek at the Zoological Museum of Amsterdam for providing unionids. We also thank Willem Mook at the Centre for Isotope Research, University of Groningen for providing river oxygen isotope data, Suzan Verdegaal for allowing us to incorporate data of one of the shells she investigated and Frank Peeters for helping with data analysis. This manuscript was greatly improved by the constructive comments of Alan Wanamaker and an anonymous reviewer. This is NSG publication n° 20090501.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Versteegh, E.A.A., Vonhof, H.B., Troelstra, S.R. et al. Can shells of freshwater mussels (Unionidae) be used to estimate low summer discharge of rivers and associated droughts?. Int J Earth Sci (Geol Rundsch) 100, 1423–1432 (2011). https://doi.org/10.1007/s00531-010-0551-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00531-010-0551-0