
Abstract
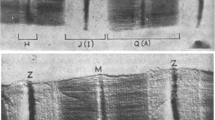
The sonic muscle of type 1 male midshipman fish produces loud and enduring mating calls. Each sonic muscle fiber contains a tubular contractile apparatus with radially arranged myofibrillar plates encased in a desmin-rich cytoskeleton that is anchored to broad Z bands (~1.2 μm wide). Immunomicroscopy has revealed patches of myosin-rich “flares” emanating from the contractile tubes into the peripheral sarcoplasm along the length of the fibers. These flares contain swirls of thick filaments devoid of associated thin filaments. In other regions of the sarcoplasm at the inner surface of the sarcolemma and near Z bands, abundant ladder-like leptomeres occur with rungs every 160 nm. Leptomeres consist of dense arrays of filaments (~4 nm) with a structure that resembles myofibrillar Z band structure. We propose that flares and leptomeres are distinct filamentous arrays representing site-specific processing of myofibrillar components during the assembly and disassembly of the sarcomere. Recent reports that myosin assembles into filamentous aggregates before incorporating into the A band in the skeletal muscles of vertebrates and Caenorhabditis elegans suggest that sonic fibers utilize a similar pathway. Thus, sonic muscle fibers, with their tubular design and abundant sarcoplasmic space, may provide an attractive muscle model to identify myofibrillar intermediates by structural and molecular techniques.






Similar content being viewed by others

References
Auerbach D, Bantle S, Keller S, Hinderling V, Leu M, Ehler E, Perriard JC (1999) Different domains of the M-band protein myomesin are involved in myosin binding and M-band targeting. Mol Biol Cell 10:1297-1308
Bader D, Masaki T, Fischman DA (1982) Immunochemical analysis of myosin heavy chain during avian myogenesis in vivo and in vitro. J Cell Biol 95:763–770
Bass AH (1989) Evolution of vertebrate motor systems for acoustic and electric communication: peripheral and central elements. Brain Behav Evol 33:237–247
Bass AH, Baker R (1990) Sexual dimorphisms in the vocal control system of a teleost fish: morphology of physiologically identified neurons. J Neurobiol 21:1155–1168
Bass AH, Marchaterre MA (1989) Sound–generating (sonic) motor system in a teleost fish (Porichthys notatus)—sexual polymorphism in the ultrastructure of myofibrils. J Comp Neurol 286:141–153
Benian GM, Tinley TL, Tang X, Borodovsky M (1996) The Caenorhabditis elegans gene unc–89, required for muscle M–line assembly, encodes a giant modular protein composed of Ig and signal transduction domains. J Cell Biol 132:835–848
Bogusch G (1975) Electron microscopic investigations on leptomeric fibrils and leptomeric complexes in the hen and pigeon heart. J Mol Cell Cardiol 7:733–745
Brantley RK, Marchaterre MA, Bass AH (1993a) Androgen effects on vocal muscle structure in a teleost fish with inter– and intra–sexual dimorphism. J Morphol 216:305–318
Brantley RK, Tseng J, Bass AH (1993b) The ontogeny of inter– and intrasexual vocal muscle dimorphisms in a sound-producing fish. Brain Behav Evol 42:336–349
Brantley RK, Wingfield JC, Bass AH (1993c) Sex steroid levels in Porichthys notatus, a fish with alternative reproductive tactics, and a review of the hormonal bases for male dimorphism among teleost fishes. Horm Behav 27:332–347
Du A, Sanger JM, Linask KK, Sanger JW (2003) Myofibrillogenesis in the first cardiomyocytes formed from isolated quail precardiac mesoderm. Dev Biol 257:382–394
Ehler E, Rothen BM, Hammerle SP, Komiyama M, Perriard JC (1999) Myofibrillogenesis in the developing chicken heart: assembly of Z–disk, M–line and the thick filaments. J Cell Sci 112:1529–1539
Harris AJ, Duxson MJ, Fitzsimons RB, Rieger F (1989) Myonuclear birthdates distinguish the origins of primary and secondary myotubes in embryonic mammalian skeletal muscles. Development 107:771–784
Hill CS, Duran S, Lin ZX, Weber K, Holtzer H (1986) Titin and myosin, but not desmin, are linked during myofibrillogenesis in postmitotic mononucleated myoblasts. J Cell Biol 103:2185–2196
Holtzer H, Hijikata T, Lin ZX, Zhang ZQ, Holtzer S, Protasi F, Franzini–Armstrong C, Sweeney HL (1997) Independent assembly of 1.6 microns long bipolar MHC filaments and I–Z–I bodies. Cell Struct Funct 22:83–93
Hosokawa T, Okada T, Kobayashi T, Hashimoto K, Seguchi H (1994) Ultrastructural and immunocytochemical study of the leptomeres in the mouse cardiac muscle fibre. Histol Histopathol 9:85–94
Ibara RM, Penny LT, Ebeling AW, Dykhuizen G van, Cailliet G (1983) The mating call of the plainfin midshipman fish, Porichthys notatus. In: Noakes DGL, Lindquist DG, Helfman GS, Ward JA (eds) Predators and prey in fishes. Junk, The Hague, pp 205-212
Karlsson U, Andersson-Cedergren E (1968) Small leptomeric organelles in intrafusal muscle fibers of the frog as revealed by electron microscopy. J Ultrastruct Res 23:417–426
Katz B (1961) The terminations of the afferent nerve fiber in the muscle spindle of the frog. Philos Trans R Soc Lond B 243:221–240
Kelly AM, Zacks SI (1969) The histogenesis of rat intercostal muscle. J Cell Biol 42:135–153
Knapp R, Wingfield JC, Bass AH (1999) Steroid hormones and paternal care in the plainfin midshipman fish (Porichthys notatus). Horm Behav 35:81–89
Lewis MK, Nahirney PC, Chen V, Adhikari BB, Wright J, Reedy MK, Bass AH, Wang K (2003) Concentric intermediate filament lattice links to specialized Z–band junctional complexes in sonic muscle fibers of the type I male midshipman fish. J Struct Biol 143:56–71
Lindholm MM, Bass AH (1993) Early events in myofibrillogenesis and innervation of skeletal, sound–generating muscle in a teleost fish. J Morphol 216:225–239
Martinez AJ, Hay S, McNeer KW (1976) Extraocular muscles: light microscopy and ultrastructural features. Acta Neuropathol (Berl) 34:237–253
Miller JB, Crow MT, Stockdale FE (1985) Slow and fast myosin heavy chain content defines three types of myotubes in early muscle cell cultures. J Cell Biol 101:1643–1650
Moncman CL, Rindt H, Robbins J, Winkelmann DA (1993) Segregated assembly of muscle myosin expressed in nonmuscle cells. Mol Biol Cell 4:1051–1067
Mukuno K (1966) The fine structures of the human extraocular muscles. (1). A laminated structure in muscle fibers. J Electr Microsc 15:227–236
Oliphant LW, Loewen RD (1976) Filament systems in Purkinje cells of the sheep heart: possible alterations of myofibrillogenesis. J Mol Cell Cardiol 8:679–688
Ono S (2001) The Caenorhabditis elegans unc–78 gene encodes a homologue of actin-interacting protein 1 required for organized assembly of muscle actin filaments. J Cell Biol 152:1313–1319
Ono S, Benian GM (1998) Two Caenorhabditis elegans actin depolymerizing factor/cofilin proteins, encoded by the unc–60 gene, differentially regulate actin filament dynamics. J Biol Chem 273:3778–3783
Ono S, Baillie DL, Benian GM (1999) UNC–60B, an ADF/cofilin family protein, is required for proper assembly of actin into myofibrils in Caenorhabditis elegans body wall muscle. J Cell Biol 145:491–502
Ono T, Yamamoto N, Yasuda K (1978) Fine structure of the "leptomere myofibrils" in the cardiac muscle of the mouse. Okajimas Folia Anat Jpn 55:69–92
Ovalle WK Jr (1972) Fine structure of rat intrafusal muscle fibers. The equatorial region. J Cell Biol 52:382–396
Ross JJ, Duxson MJ, Harris AJ (1987) Formation of primary and secondary myotubes in rat lumbrical muscles. Development 100:383–394
Ruska H, Edwards GA (1957) A new cytoplasmic pattern in striated muscle fibers and its possible role in relation to growth. Growth 21:73–88
Seiler SH, Fischman DA, Leinwand LA (1996) Modulation of myosin filament organization by C–protein family members. Mol Biol Cell 7:113–127
Shimizu T, Dennis JE, Masaki T, Fischman DA (1985) Axial arrangement of the myosin rod in vertebrate thick filaments: immunoelectron microscopy with a monoclonal antibody to light meromyosin. J Cell Biol 101:1115–1123
Srikakulam R, Winkelmann DA (1999) Myosin II folding is mediated by a molecular chaperonin. J Biol Chem 274:27265–27273
Srikakulam R, Winkelmann DA (2004) Chaperone-mediated folding and assembly of myosin in striated muscle. J Cell Sci 117:641–652
Thornell LE, Eriksson A (1981) Filament systems in the Purkinje fibers of the heart. Am J Physiol 241:H291–H305
Thornell LE, Sjostrom M, Andersson KE (1976) The relationship between mechanical stress and myofibrillar organization in heart Purkinje fibres. J Mol Cell Cardiol 8:689–695
Vikstrom KL, Rovner AS, Saez CG, Bravo-Zehnder M, Straceski AJ, Leinwand LA (1993) Sarcomeric myosin heavy chain expressed in nonmuscle cells forms thick filaments in the presence of substoichiometric amounts of light chains. Cell Motil Cytoskeleton 26:192–204
Walker SM, Schrodt GR, Currier GJ (1975) Evidence for a structural relationship between successive parallel tubules in the SR network and supernumerary striations of Z line material in Purkinje fibers of the chicken, sheep, dog and rhesus monkey heart. J Morphol 147:459–473
Wang K (1996) Titin/connectin and nebulin: giant protein rulers of muscle structure and function. Adv Biophys 33:123–134
Welikson RE, Fischman DA (2002) The C–terminal IgI domains of myosin–binding proteins C and H (MyBP–C and MyBP–H) are both necessary and sufficient for the intracellular crosslinking of sarcomeric myosin in transfected non–muscle cells. J Cell Sci 115:3517–3526
Winegrad S (1999) Cardiac myosin binding protein C. Circ Res 84:1117–1126
Author information
Authors and Affiliations
Corresponding author
Additional information
This work was supported by the Intramural Research Program of the NIAMS, NIH, HHS (KW).
Rights and permissions
About this article
Cite this article
Nahirney, P.C., Fischman, D.A. & Wang, K. Myosin flares and actin leptomeres as myofibril assembly/disassembly intermediates in sonic muscle fibers. Cell Tissue Res 324, 127–138 (2006). https://doi.org/10.1007/s00441-005-0110-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00441-005-0110-3