
Abstract
The objectives of the present study were to standardize a reproducible infection procedure with Cryptocaryon irritans and to examine the effects of infectious dose level on the immune protection in Mozambique tilapia (Oreochromis mossambicus). This study demonstrated that direct enumeration of trophonts on the pectoral fin was useful to quantitatively assess immune protection against C. irritans. The number of trophonts on a pectoral fin was positively correlated with infectious dose of live theronts. Fish immunized by direct exposure under controlled laboratory conditions allowed for in depth examination of the effects of the degree of infectious dose on immune response. There was no significant positive correlation between the initial infectious dose and degree of immune responses. Mozambique tilapia initiated a strong immune protection by direct exposure with even a small number of parasites (e.g. 300 theronts per fish). Moreover, as the result of the protein analysis, we identified 28 kD proteins that could be responsible for the immobilizing antigen.
Similar content being viewed by others


Introduction
Severe infection of marine fish by the ciliated protozoan parasite, Cryptocaryon irritans, is a major problem in commercial aquaculture in tropical and sub-tropical regions because it causes high mortalities of fish and leads to heavy economic loss (Rigos et al. 2001). Despite the increasing incidence of outbreaks, current control methods for cultured food fishes have been limited to chemical treatments, fresh water dip, or transference of those infected fish from the culture (Hirazawa et al. 2003). However, these methods have several drawbacks for large-scale commercial aquaculture so the development of effective vaccines has been proposed as a means of alternative control (Rigos et al. 2001).
Immobilization antigens (i-antigens) are important candidates for subunit vaccines against C. irritans. The i-antigen is defined by the fact that specific antibodies against this antigen interfere with movement of cilia and cause immobilization of ciliated protozoan parasites (Lin et al. 1996). Yoshinaga and Nakazoe (1997) demonstrated for the first time that sera from immunized fish immobilized live theronts in vitro. Serum and cutaneous mucus from immunized fish contain immobilization antibodies (Yambot and Song 2006; Luo 2007).
Standardizing quantitative measurements for evaluating immune protection in fish is essential to determine the efficacy of immunizations against C. irritans. Specific antibody level, mortality, and infection level are used to evaluate immune protection in host. Although the methods for measuring antibody level and mortality are already established, measuring the infectious level is not standardized yet. Since C. irritans is an ectoparasite, it is possible to measure the infectious level numerically by counting parasite burden on fish. When the infection level is measured for evaluating immune protection, it is important to outline a procedure for standardizing both the exposure of fish to C. irritans and the subsequent parasite density on the fish. In previous studies, successful attempts at experimentally inducing Cryptocaryoniasis have been reported but no one has validated experimental models resulting in well-controlled and quantitatively reproducible effects. Recent improvements in propagation methods for C. irritans (Dan et al. 2006) have made it possible to produce theront suspensions with nearly constant viability and to complete direct exposures in fish using accurately adjusted infectious doses.
In this study, measurement of the infectious level of C. irritans was defined with Mozambique tilapia (Oreochromis mossambicus) as a model species. There are several advantages of using Mozambique tilapia for the study related to C. irritans. First, raising Mozambique tilapia is easy and inexpensive with high fecundity rates. The Mozambique tilapia is one of the most euryhaline of teleost fish, so fresh water reared naïve tilapia have great utility as a negative control to study C. irritans infection. Moreover, Mozambique tilapia is very susceptible to C. irritans infection compared to other marine fish (Burgess and Matthews 1995b).
There is also a lack of basic study that examines the effects of the degree of infection on immune responses in fish. Such studies that define the baseline of immune responses are necessary to form a basis for evaluating the immunity of fish against C. irritans. Moreover, those fundamental studies are helpful when the efficacy of a vaccine is evaluated after challenge experiments.
Materials and methods
Fish
All fish used in this study were kept in 60-L glass aquaria that received continuous aeration and flow-through seawater (120 L/h). Seawater was continuously pumped from Kāne‘ohe Bay, filtered with 5 μm filters, and treated with UV before use in aquaria. The water temperature was 27.5 ± 0.5°C, and the salinity of seawater was 34‰. Broodstocks of Mozambique tilapia raised in fresh water were provided by Dr. Gordon Grau at the Hawaii Institute of Marine Biology (HIMB), University of Hawaii at Manoa. Tilapia broodstocks were initially kept in fresh tap water in the SEA lab at HIMB for 2 days and acclimated to seawater by gradually increasing the salinity up to 34 ppt over a 2 week period. All fish were fed approximately 2% of their body weight once daily with Silver Cup Trout Chow (Nelson and Sons, Murray, UT). All experimental fish used for the study were same age and bred from the same parents. Hatched tilapia larvae (7 days old) were separated from their parents into another aquarium and reared in UV-treated seawater as described above to maintain a C. irritans-free environment. All experiments were conducted in accordance with the principles and procedures approved by the Institutional Animal Care and Use Committee, University of Hawaii at Manoa.
Parasite isolation
Laboratory cultures of C. irritans obtained initially from Sergeant Major damselfish (Abudefduf abdominalis) from Kāne‘ohe Bay were maintained in UV-treated seawater at 27.5 ± 0.5°C in 60-L fish aquaria by serial passage to naïve Mozambique tilapia. A naïve tilapia was added and replaced twice a month for propagation of tomonts in the aquarium. The health of the fish was observed daily and any dead fish or fish with lesions were removed from tanks. Protomonts were collected from heavily infected fish with a 1,000 μl micropipette using a stereo-microscope (American Optical, Model 41) to visualize individual parasites. One hundred protomonts were collected and washed four times and placed in a well of 24-well culture plate with filtered (0.2 μm pore-size filter) seawater and supplemented with 100 I.U. per ml penicillin G potassium and 100 μg per ml streptomycin sulfate (Yoshinaga et al. 2007). After confirmation of encystment of the tomonts, cultures were incubated at room temperature. Newly hatched theronts from the culture were used for infection experiments, immobilization assay, and future propagation of C. irritans and the remaining parasites were stored in sterile phosphate-buffered saline (PBS) at −80°C until processed.
Experimental design
A total of 210 tilapia (mean weight 30.1 ± 9.3 g / length 12.3 ± 1.4 cm) were used for this study. They were divided into seven groups of 30 fish. There was no statistically significant difference in weight and body length of fish between groups. Blood was collected from the caudal vein from fish anesthetized by 0.2 ppm of tricaine methanesulfonate (MS 222: Finquel, Redmond, WA). Blood samples were drawn using a needle and syringe and allowed to clot overnight at 4°C. Serum was obtained by centrifugation of blood at 1,000 × g for 10 min at 4°C. Serum samples from each individual fish were distributed in aliquots of 20 μl in 200 μl microcentrifuge tubes and stored at −20°C for later analysis. Seven days after the first blood collection, newly hatched theronts with concentrations of 0 (0 K), 333 (0.3 K), 3,333 (3.3 K), 10,000 (10 K), 20,000 (20 K), 30,000 (30 K), and 56,667 (57 K) theronts per fish were transferred to each aquarium. This primary exposure was run in duplicate (15 fish per tank). Two days after the initial exposure, the number of infected trophonts on the left pectoral fin was counted under an inverted microscope (Olympus IX; Olympus Optical, Tokyo) in order to measure the infection level for each fish. There were two major reasons for using pectoral fins: (1) it could be regenerated completely in a month, and (2) its transparency was ideal for counting trophonts under the microscope. All fish were transferred into clean tanks every 5 days to avoid accidental reinfection by C. irritans. Blood was collected 21 days post initial exposure and serum extracted and stored as described above. Twenty eight days after the initial exposure, 15 fish from each group were randomly selected, mixed with fish from the other groups, placed into clean aquaria and exposed to 10,000 (10 K) theronts per fish. The remaining 15 fish from each group were mixed in a clean tank but not exposed to any C. irritans. Two days after the secondary exposure, the number of infected trophonts on the right pectoral fin of each fish was counted using an inverted microscope. Blood was collected 49 days post initial exposure and serum extracted and stored as described above.
Immobilization assay
Each serum sample from individual fish was tested for immobilizing activity against live theronts in vitro. Assays were done as described by Clark et al. (1987) with modifications. Serum was heat inactivated at 56°C for 30 min to inactivate complement. Serum samples from individual fish were serially diluted with filtered seawater and 20 μl from each dilution were added into each well of a 96-well plate (Corning Costar, Corning, NY) containing 100 μl of theront suspension (approximately 200 theronts per well) in filtered seawater. After 30 min incubation at room temperature, the number of immobilized theronts was counted using the inverted microscope (Olympus IX; Olympus Optical, Tokyo) to find the percentage of the parasites that were no longer motile. The immobilization titre was defined as the reciprocal value of the dilution at which 95% of the parasites were no longer motile.
SDS-PAGE and Western blot analysis
Serum from immunized fish directly exposed to live theronts twice was analyzed by Western blot analysis for reactivity against any antigenic proteins in the homogenates of C. irritans theronts. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed as described by Laemmli (1970). Briefly, sonicated theront proteins (0.5 μg per lane measured using the BCA protein assay) were resolved on 10% SDS-PAGE gels under non-reducing conditions and transferred to a polyvinylidene difluoride (PVDF) membrane using a Mini-Protean three transblot apparatus (BioRad, California, USA). Precision Plus protein standards (Bio-Rad) were used as molecular weight markers. Protein loading was confirmed in a duplicated gel visualized with silver stain (Wray et al. 1981). The PVDF membranes were blocked with Tween-PBS containing 1% (w/v) BSA overnight at 4°C or for 1 h at room temperature with mixing on an orbital shaker. The membranes were washed 3× with Tween-PBS. Three appropriate hyperimmune fish sera were pooled together and used to probe the Western blot. Pooled serum from 3 naïve fish that had never been exposed to C. irritans was used as the negative control. Pooled fish serum samples (diluted 1/75 in PBS), rabbit anti-tilapia IgM antibody (1:3,000 in PBS; Takemura 1993), and goat anti-rabbit IgG horseradish peroxidase conjugate (1:5,000 in PBS; Jackson Immuno Research Laboratories, West Grove, PA) were applied in sequential order to the PVDF membranes and incubated for 1 h at room temperature. Immunoreactive bands were visualized with West Dura chemiluminescent substrate (Pierce, Rockford, IL).
Twenty eight kD antigen protein isolation
Theront proteins homogenized by an ultrasonic sonicator were size fractionated on a 10% SDS gel and the 28 kD protein band visualized using a reversible zinc stain kit (Bio-Rad). The band excised from the gel was destained using 25 mM Tris containing 192 mM glycine. The 28 kD protein was recovered from the SDS band using a Centrilutor Micro-Electroeluter (Millipore, MA) at 200 V for 2 h according to the manufacturer“s instructions. Eluted protein was ultrafiltered by the Centricon-10 cartridge (Millipore) at 3,000 × g for 2 h at 4°C. PBS was added to dilute the concentrated protein and the resuspended sample was dialyzed to remove SDS and other impurities for 2 h at 4°C. Purity of the 28 kD protein was confirmed in a gel stained with silver stain.
Enzyme-linked immunosorbent assay (ELISA)
ELISA was conducted to measure the level of specific antibodies in the serum against the purified 28 kD C. irritans antigen protein. The 28 kD protein was diluted to 5 μg/ml in carbonate coating buffer (0.05 M Na2CO3 and 0.05 M NaHCO3, pH 9.6), and 50 μl of the homogenate was added into each well of a 96-well ELISA plate (Corning Costar, Corning, NY) and incubated overnight at 4°C. All subsequent steps were performed at room temperature. Wells were washed 3× with low salt wash buffer (20 mM Tris, pH 7.3; 380 mM NaCl; 0.05% Tween 20). Non-specific binding sites were blocked with PBS (250 μl/well) containing 1% (w/v) bovine serum albumin (BSA) for 2 h. After three washes with low salt wash buffer, diluted fish serum (1:500) in PBS were added to the plate at 50 μl/well and incubated for 3 h. PBS alone in the well served as the negative control. The plate was washed 5× with high salt wash buffer (20 mM Tris, pH 7.3; 500 mM NaCl; 0.1% Tween 20). At sequential 1 h intervals, wells were incubated with a rabbit anti-tilapia IgM (1:2,500 in PBS) at 50 μl/well and commercial goat anti-rabbit IgG horseradish peroxidase conjugate (1:2,500 in PBS) at 50 μl/well. Between additions of each new reagent, the ELISA plate was washed 5× with high salt wash buffer. Finally, 50 μl of 2,2′-azino-bis 3-ethylbenzthiazoline-6-sulfonic acid (ABTS) peroxidase substrate (KPL, Gaithersburg, MD) was added to each well to allow for chromogenic development. After 20 min, the absorbance at 405 nm was read using a microplate reader (Molecular Devices Co., Sunnyvale, CA). In this study, ELISA results were reported as optical density (OD) values of given optimal dilutions of serum and mucus due to the large number of samples. Optimal dilutions of serum and mucus were determined by the preliminary ELISA with serial dilutions (twofold) of samples collected from naïve and hyperimmune fish. The optimal dilutions allowed for maximum OD for positive controls with minimal background reading in negative controls.
Statistical analysis
Data are expressed as mean ± S.E.M. (standard error of mean). Parametric statistical tests (the Student's t-test and ANOVA) or non-parametric statistical tests (the Mann–Whitney U-test and the Kruskal–Wallace test) were used for statistical comparisons of data. Either Student–Newman–Keuls multiple range test or Dunn's test was used to determine which treatments were differed significantly from others. Statistical analyses were performed using Sigma Plot 11 software (Systat Software Inc., San Jose, CA). Measurements were considered significant when P values of both parametric and non-parametric tests were <0.05.
Results
Effect of exposure dose on parasite burden
To examine the relationship between the exposure doses and infectious levels, fish were directly exposed with different doses of live theronts. The infectious level was measured for each group by counting the number of trophonts on a pectoral fin (Fig. 1). Fish exposed to higher concentrations of theronts were burdened with more trophonts. The number of trophonts on a pectoral fin was dose dependent following the primary exposures (Fig. 1c). Fish without C. irritans exposure (0 K) developed no signs of disease and had no mortality (data not shown). All experimentally exposed fish were colonized by trophonts, excluding 50% of fish (15 fish) in the infection group that were dosed of 0.3 K theronts per fish, which did not show any sign of trophont infection on a pectoral fin. Two fish in the infection group at a dose of 57 K theronts per fish died 4 days after the primary exposure. There was a clear infective dose effect, which was significant overall (Student–Newman–Keuls test, P < 0.05). Indeed, each group was significantly different from the others (Student–Newman–Keuls test, P < 0.05). There was no significant difference between the exposure results of the two replicates (Mann–Whitney U-test, P > 0.05).

Comparison of the number of trophont on a pectoral fin 2 days after the primary exposure at different infectious dose of live theronts. a A pectoral fin clipped from tilapia which was exposed to 3 K theronts per fish. b A pectoral fin clipped from tilapia which was exposed to 57 K theronts per fish (scale bar = 400 μm). c The number of trophont is the mean (S.E.M.) for each experimental group (n = 30). Mean values followed by the different superscript letters are statistically different (Student–Newman–Keuls test, P > 0.05)
Immobilization assay after the primary exposure
Immobilization assays were conducted to examine the immune response of fish following the exposure with different doses of theronts (Fig. 2). All individual sera from immune fish exposed to any doses of parasite showed immobilization activity against live theronts in vitro. Serum from naive fish that were never exposed to the parasite did not immobilize live theronts at all (data not shown). Statistical analysis did not reveal any significant differences in the immobilization titres between groups (Dunn's test, P > 0.05) except the group exposed to the lowest infectious dose (0.3 K) (Dunn's test, P < 0.05). Although the immobilization titre of the medium-dose (10 K) group looked higher, the difference was not statistically significant from others (Dunn's test, P > 0.05). The lowest immobilization titre occurred at the lowest infectious dose (0.3 K). There was a significant difference in immobilization titre between two replicates of all groups (Mann–Whitney U-test, P < 0.05) except the highest dose group (57 K) (Mann–Whitney U-test, P > 0.05).

Immobilization titres following the primary exposure at different infectious doses of theronts. Antisera were collected 21 days after the primary exposure. Immobilization titre was the inverse of the highest dilution in which 95% of theronts were immobilized in vitro. The values (n = 28–30) are mean (±SEM) for each experimental group. Columns that have the same superscripts are not significantly different (Dunn's test, P > 0.05)
Western blotting analysis and 28 kD protein purification
SDS-PAGE and Western blotting analyses were conducted to determine if antiserum from immunized fish would detect any immunoreactive proteins in theronts. Pooled serum from hyperimmune fish following the secondary exposure with live theronts were used to probe Western blots of whole proteins of theronts. As shown by Western blot analyses (Fig. 3a), the antiserum antibodies recognized one specific band (approximately 28 kD). No signals were detected when the proteins were probed with pooled serum from naïve fish (data not shown). Following silver staining, a protein profile for total theront proteins revealed a heterogeneous mixture of molecular weight proteins with three prominent protein bands (Fig. 3b; lane 1). The molecular weight of the one of the three prominent bands was approximately 28 kD. Molecular weights of other two bands could not be measured because those were out of measurable range (between 10 and 250 kD) of the protein molecular weight standard. The 28 kD band was excised from another SDS-PAGE gel following zinc staining and recovered from the gel using the Centrilutor Micro-Electroeluter. The eluted 28 kD protein was verified after the purification by the silver staining under non-reducing conditions (Fig. 3b; lane 2).

a Western blot probed with pooled sera from immunized fish following the secondary direct exposure with C. irritans theronts. Lane 1: molecular weight standards; lane 2: sonicated total proteins of C. irritans theronts. b Silver stained SDS–PAGE. Lane 1: molecular weight standards; lane 2: sonicated total proteins of C. irritans theronts; lane 3: Isolated 28 kD protein using the Centrilutor Micro-Electroeluter
Specific antibody responses against 28 kD protein after the primary exposure
ELISA was carried out to measure the level of specific antibodies against the purified 28 kD protein before and after the primary exposure with different doses of C. irritans theronts. The level of specific antibodies increased significantly 21 days after the primary exposure at each infective dose compared to levels before the exposure (Mann-Whitney U-test, P < 0.05; Fig. 4). There was no statistically significant difference in the ELISA value between groups prior to the primary exposure (Dunn's test, P > 0.05). However, the ELISA value after primary exposure peaked at the dose of 10 K theronts per fish and decreased significantly with increasing the number of infectious dose (Dunn's test, P > 0.05). There was no significant difference in ELISA value before the exposure between groups of the two replicates (Mann-Whitney U-test, P > 0.05). There were significant differences in ELISA value after the primary exposure between groups of the two replicates (Mann-Whitney U-test, P < 0.05) except with the highest dose group (57 K) (Mann-Whitney U-test, P > 0.05).

Specific antibody response against the purified 28 kD antigenic protein before and after the primary exposure at different infectious doses of theronts. Antisera were collected 21 days after the primary exposure. ELISA results were reported as OD values of given optimal dilutions of serum (1:500). The values (n = 28–30) are mean (±SEM) for each experimental group. Significant differences between before and after exposure are denoted * (P < 0.05, Mann-Whitney U-test). Columns that have the same superscripts are not significantly different (Student–Newman–Keuls test or Dunn's test, P > 0.05)
Effect of the secondary exposure on parasite burden
Fish were re-exposed to parasites (10 K theronts per fish for all groups) 28 days after the primary exposure in order to examine the effect of infective doses on the immune protection. When fish were primarily exposed to and recovered from C. irritans infection, these immunized fish developed complete protection against the secondary exposure irrespective of initial parasite dose (Table 1; group 2–7). However, 0.9 ± 0.2 trophonts per pectoral fin were detected from fish that primarily received the lowest dose (0.3 K) although the number of trophont was much lower than the primary number of trophont (24.83 ± 1.79 trophonts per a pectoral fin) at a dose of 10 K theronts per fish. No parasites were found in fish without the secondary exposure (Table 1; group 8 through 14).
Immobilization assay after the secondary exposure
Immobilization assay was conducted to examine the immune response after the secondary exposure (Fig. 6). The immobilization titres of antisera after the secondary exposure (Table 1; group 2–7) were significantly higher than those of antisera from fish without the secondary exposure (group 1 and 8–13) (Mann-Whitney U-test, P < 0.05). There was no statistically significant difference in the immobilization titre between groups that did not undergo the secondary exposure (group 8–13) (Dunn's test, P > 0.05) except at the highest dose of exposure (group 14) (Dunn's test, P < 0.05). The immobilization titre in fish with the secondary exposure peaked at dose of 10 K theronts per fish and decreased significantly with increasing the number of infectious dose (Student–Newman–Keuls test, P < 0.05). There was no replication for this assay.
Specific antibody responses against 28 kD protein after the secondary exposure
ELISA was carried out to measure the level of specific antibodies against the purified 28 kD protein after the secondary exposure. The level of specific antibodies in fish (Table 1; group 2–7) that underwent the secondary exposure was always significantly higher than that in fish (group 9–14) that did not undergo the secondary exposure at each infective dose (Mann-Whitney U-test, P < 0.05). The level of specific antibodies in fish (group 2–7) that underwent the secondary exposure decreased with increasing the number of infectious dose (Dunn's test, P < 0.05). There was no replication for this assay.
Discussion
This study demonstrated that examination of the pectoral fin is a useful method for direct enumeration of the prevalence of trophonts on an infected fish. Previous studies (Burgess and Matthews 1995a; Yambot and Song 2006) counted the numbers of protomonts or tomonts to quantify degree of infection. Such approaches, however, should be used with caution because there is variability in timing of trophonts exiting from the host (3 ± 7 days after attachment) (Colorni 1985; Colorni and Diamant 1993). Moreover, we observed that fish were eating protrophonts or tomonts, so there is a possibility of losing protomonts or tomonts before measuring the infectious level. Therefore, in this study, we attempted to count the number of trophonts on a pectoral fin 2 days after exposures. Some studies cautioned that such an approach was difficult to be used because trophonts of both C. irritans and I. multifiliis often exist in aggregates (Ewing et al. 1988; Burgess and Matthews 1995a). However, in our study, even when aggregated trophonts existed, they were easily identified and counted under the microscope. In the case of I. multifiliis, Clayton and Price (1988) also attempted to quantify parasite intensity on the basis of the numbers of white spots visible on fish body and found a significant linear relationship between infection and response. Houghton and Matthews (1986) indicated that the average density of infected I. multifiliis differs on different parts of the host and found more trophonts on the periphery of the fins. The high density of trophonts on fins may be related to impaired immune responses at the sites of reduced blood flow (Burgess and Matthews 1995a).
Doses received by tilapia in the current experimental exposures to C. irritans were almost equivalent to those in a prior study using grouper (Epinephelus coioides; 20 ± 2 g in weight and 10–12 cm in length) by Luo et al. (2007). However, our infectious dose level resulted in significant host mortalities in previous studies using juvenile mullet (Chelon labrosus; average weight 4 g) conducted by Burgess and Matthews (1995a, b). The reason the lethal doses differed between those studies is not clear. One possible explanation is that the smaller fish size could simply increase the parasite density, and another reason is that the variation in susceptibility to C. irritans could be species dependent.
There was a need to address which life stage of C. irritans would be best used for exposure to standardize challenge method. Previous studies used tomonts (Yambot and Song 2006) or theronts (Luo et al. 2007; Bai et al. 2008) for challenge exposures. However, in the case of using tomonts, since each tomont has variable excystment time, occurring between 3 and 72 days (Colorni and Burgess 1997), experimental fish must be maintained under continual exposure to C. irritans until all tomonts excyst. The possibility could not initially be ruled out that immune protection would be established in the fish under the challenge exposure with tomonts. Using theronts for the challenge exposure is more accurate and reliable for assessing of the immune protection against C. irritans.
In this study, we examined effects of initial infectious doses on immune responses in tilapia infected with C. irritans. The direct exposure induced immobilization activities and specific antibody responses against C. irritans in fish at all infective doses between 0.3 and 57 K theronts per fish. The parasite burdens 2 days post second exposure were significantly lower than 2 days post primary exposure. These differences in parasite burden between first and second exposure must be due to acquired immunity in fish. The protective immunity was developed regardless of dose of original exposure between 0.3 and 57 K theronts per fish.
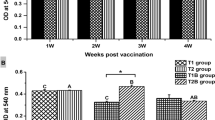
Duration of immunological memory seemed not to be dependent on infectious doses with the primary exposure. High levels of immobilization activity and specific antibodies against the 28 kD antigen were produced 21 days after initial exposure at all dose levels. However, the apparent decreases in immobilization titres and the specific antibody levels were observed in fish that were not re-exposed to C. irritans for 49 days after the initial exposure (Fig. 7). The decline in immobilization titres and the specific antibody level in those fish probably resulted from a loss of antigenic stimulus. Very similar results were obtained by Luo et al. (2007) using grouper (Epinephelus coioides).
One of the most interesting findings in the present study was that there was no significant positive correlation between the initial infectious dose and immune responses after the primary exposure. Previous similar studies demonstrated that the level of antibody production and the degree of protection were a positive relationship with immunization dose. Burgess and Matthews (1995a) reported that the degree of protection against C. irritans depended upon the levels of infection established, as assessed from the mean and range of numbers of the trophonts released. Yambot and Song (2006) found that the antibody titres increased with an increase in the number of parasite used in the exposure. However, present study (Fig. 2) showed that there was no significant difference in immobilization titre at infective doses between 3 and 57 K theronts per fish. That is, immobilization activity already reached a threshold at low level of infectious dose between 0.3 and 3 K and was not further activated by higher dose of theronts.
On the other hand, the ELISA value of specific antibodies against the 28 kD antigen reached a peak in the medium dose group (10 K) and decreased with increasing initial doses (Fig. 4). Moreover, a negative correlation was found between primary infectious dose and the ELISA value after the secondary exposure (Fig. 5). These ELISA results indicate that infection with high numbers of C. irritans changes characteristics of immune responses in fish. One possibility to explain these results is that stress responses induced by heavy infection may modulate immune functions including antibody production and the memory performance in fish. Normally, the stress response of the fish is assessed by measuring plasma cortisol level because the stress response is characterized by activation of the hypothalamus-pituitary-interrenal (HPI) axis (Wendelaar-Bonga 1997). Although the plasma cortisol level was not measured in this study, we observed that heavily infected tilapia had darker skin color compared to healthy and tranquil fish which had pale silver skin color. Since dark skin pigmentation reflect a stressed state with higher plasma cortisol levels (Gilham and Baker 1985; Hoglund et al. 2000; Ruane et al. 2005), our observation suggests that heavily infected fish by C. irritans may be under stressed conditions. Poole et al. (2000) and Rauane et al. (2000) also demonstrated that prolonged infection by a large number of the sea lice (Lepeophtheirus salmonis) led to chronic stress in salmonid fish. Moreover, Bowers et al. (2000) indicated an association between heavy infection of sea lice and a significant increase in cortisol and glucose level in fish. Although the complete mechanisms of the immune suppression induced by cortisol are poorly understood (Varsamos et al. 2006), many studies report direct effects of cortisol on adaptive immunity in fish, such as induction of apoptosis in B-cells (especially plasma B-cells) (Weyts et al. 1998; Weyts et al. 1999) and suppression of lymphocyte proliferation (Espelid et al. 1996). Those direct effects of cortisol on lymphocytes could lead to suppression of plasma IgM levels in fish (Carlson et al. 1993; Nagae et al. 1994; Iguchi et al. 2003; Varsamos et al. 2006). Several studies reported that cortisol administration caused susceptibility of fish to infectious diseases (Wiik et al. 1989; Houghton and Matthews 1990). However, it is still controversial whether parasites act as a stressor and whether their abundance is correlated to levels of cortisol. Some studies reported that parasites do not generate typical stress responses (Laidley et al. 1988; Grutter and Pankhurst 2000). Further study is needed to elucidate the mechanism of the immune suppression mediated via heavy infection with ectoparasites such as C. irritans.

Immobilization titres in fish with or without the secondary exposure (10 K theronts per fish) at different primary infectious doses. Immobilization titre was the inverse of the highest dilution in which 95% of theronts were immobilized in vitro. The values (n = 13–15) are mean (±SEM) for each experimental group. Significant differences between non-boosted fish and boosted fish are denoted * (P < 0.05, Mann-Whitney U-test). Columns that have the same superscripts are not significantly different (Dunn's test, P > 0.05)
The present study demonstrated that serum isolated from tilapia immunized by direct exposure immobilized the live theronts irrespective of initial infectious dose. The immobilization titres were significantly boosted after the secondary exposure in all groups. These results suggest that levels of i-antigen specific antibody were significantly increased following the secondary exposure because the i-antigen specific antibody interferes with movement of cilia and causes immobilization of C. irritans (Hatanaka et al. 2007).
However, Fig. 6 showed contradictory results. The level of specific antibodies did not increase in a similar manner as the immobilization titres post secondary infection except in the 0.3 K exposed group, compared between Figs. 4 and 6. Sigh and Buchmann (2001) also reported similar results that immobilization activity and the level of specific antibody (detected by ELISA) were not clearly correlated. That is, unlike immobilization titre, the specific antibody response was not boosted in fish post secondary exposure. Thus, our results suggest that the potential involvement of innate immunity in the immobilization activities should also be considered, even when the serum was heat-inactivated. Colorni et al. (2008) reported that piscidin three, an antimicrobial polypeptide, isolated from mast cells of hybrid striped bass was involved in immobilization responses and lethal to C. irritans theronts. It is not yet clear if other humoral factors such as complement, antimicrobial polypeptides, lysozyme, and lectins were boosted after the secondary exposure and resulted in higher immobilization titre compared to those post primary exposure. Further studies are necessary to describe the mechanism of immobilization mediated by innate immunity.

Specific antibody response against the purified 28 kD antigenic protein following the secondary exposure at different infectious doses of theronts. ELISA results were reported as OD values of given optimal dilutions of serum (1:500). The values (n = 13–15) are mean (±SEM) for each experimental group. Significant differences between non-boosted fish and boosted fish are denoted * (P < 0.05, Mann-Whitney U-test). Columns that have the same superscripts are not significantly different (Student–Newman–Keuls test or Dunn's test, P > 0.05)
SDS-PAGE and Western blotting analysis were conducted to identify proteins that were responsible for the i-antigen. Hatanaka et al. (2007) identified a predominant 32 kD polypeptide on the surface of the cilia of theronts as an i-antigen designated as G32. In subsequent studies, fish antiserum against 37 kD i-antigen purified from another C. irritans isolate was found not to immobilize G32 theronts, indicating that multiple i-antigen serotypes exist among C. irritans isolates (Hatanaka et al. 2008). Similarly, our results indicated that it was highly likely that the 28 kD protein was the i-antigen of C. irritans. Although our i-antigen was smaller in size compared to the 32 or 37 kD i-antigens of other C. irritans isolates, our finding has implications for the existence of the different serotype in Hawaii.
In conclusion, counting parasites on the pectoral fin is a useful method for direct enumeration of the prevalence of C. irritans on infected fish. One of the major findings in this study was that immunized fish showed a strong immune protection by direct exposure with even a small number of parasites. There was no significant positive correlation between the initial infectious dose and degree of immune responses. Moreover, we identified 28 kD protein that could be responsible for the immobilizing antigen. Although further study is needed to confirm that the 28 kD protein is, in fact, an i-antigen, there is the possibility of existence of different serotypes in Hawaii. Since an effective vaccine must provide cross-protection against all C. irritans, if there are several serotypes in C. irritans, the identification of antigenic proteins that are common in all serotypes is needed for future vaccine development.
References
Bai JS, Xie MQ, Zhu XQ, Dan XM, Li AX (2008) Comparative studies on the immunogenicity of theronts, tomonts and trophonts of Cryptocaryon irritans in grouper. Parasitol Res 102:307–313
Bowers JM, Mustafa A, Speare DJ, Conboy GA, Brimacombe M, Sims DE, Burka JF (2000) The physiological response of Atlantic salmon, Salmo salar L., to a single experimental challenge with sea lice, Lepeophtheirus salmonis. J Fish Dis 23:165–172
Burgess PJ, Matthews RA (1995a) Cryptocaryon irritans (Ciliophora)—Acquired protective immunity in the thick lipped mullet, Chelon labrosus. Fish Shellfish Immunol 5:459–468
Burgess PJ, Matthews RA (1995b) Fish Host-Range of 7 Isolates of Cryptocaryon irritans (Ciliophora). J Fish Biol 46:727–729
Carlson RE, Anderson DP, Bodammer JE (1993) In vivo cortisol administration suppresses the in vitro primary immune response of winter flounder lymphocytes. Fish Shellfish Immunol 3:299–312
Clark TG, Dickerson HW, Gratzek JB, Findly RC (1987) In vitro Response of Ichthyophthirius multifiliis to sera from immune channel catfish. J Fish Biol 31:203–208
Clayton GM, Price DJ (1988) Ichthyophthirius multifiliis—standardization of the infection response model in Ameca splendens (Miller and Fitzsimons). J Fish Dis 11:371–377
Colorni A (1985) Aspects of the biology of Cryptocaryon irritans, and hyposalinity as a control measure in cultured gilt head sea bream Sparus aurata. Dis Aquat Organ 1:19–22
Colorni A, Burgess P (1997) Cryptocaryon irritans Brown 1951, the cause of ‘white spot disease’ in marine fish: an update. Aqua Sci Conserv 1:217–238
Colorni A, Diamant A (1993) Ultrastructural features of Cryptocaryon irritans, a ciliate parasite of marine fish. Eur J Protistol 29:425–434
Colorni A, Ullal A, Heinisch G, Noga EJ (2008) Activity of the antimicrobial polypeptide piscidin 2 against fish ectoparasites. J Fish Dis 31:423–432
Dan XM, Li AX, Lin XT, Teng N, Zhu XQ (2006) A standardized method to propagate Cryptocaryon irritans on a susceptible host pompano Trachinotus ovatus. Aquaculture 258:127–133
Espelid GBL, Steiro K, Bogwald J (1996) Effects of cortisol and stress on the immune system in Atlantic Salmon (Salmo salar L.). Fish Shellfish Immunol 6:95–110
Ewing MS, Ewing SA, Kocan KM (1988) Ichthyophthirius (Ciliophora)—population studies suggest reproduction in host epithelium. J Protozool 35:549–552
Gilham ID, Baker BI (1985) A black background facilitates the response to stress in teleosts. J Endocrinol 105:99–105
Grutter AS, Pankhurst NW (2000) The effects of capture, handling, confinement and ectoparasite load on plasma levels of cortisol, glucose and lactate in the coral reef fish Hemigymnus melapterus. J Fish Biol 57:391–401
Hatanaka A, Umeda N, Yamashita S, Hirazawa N (2007) Identification and characterization of a putative agglutination/immobilization antigen on the surface of Cryptocaryon irritans. Parasitology 134:1163–1174
Hatanaka A, Umeda N, Hirazawa N (2008) Molecular cloning of a putative agglutination/ immobilization antigen located on the surface of a novel agglutination/immobilization serotype of Cryptocaryon irritans. Parasitology 135:1043–1052
Hirazawa N, Goto T, Shirasu K (2003) Killing effect of various treatments on the monogenean Heterobothrium okamotoi eggs and oncomiracidia and the ciliate Cryptocaryon irritans cysts and theronts. Aquaculture 223:1–13
Hoglund E, Balm PH, Winberg S (2000) Skin darkening, a potential social signal in subordinate arctic charr (Salvelinus alpinus): the regulatory role of brain monoamines and pro-opiomelanocortin derived peptides. J Experim Biol 203:1711–1721
Houghton G, Matthews RA (1986) Immunosuppression of juvenile Carp (Cyprinus carpio L) to the parasite Ichthyophthirius multifiliis induced by a corticosteroid. Dev Comp Immunol 10:142–142
Houghton G, Matthews RA (1990) Immunosuppression in juvenile carp, Cyprinus carpio L.: the effects of the corticosteriods triamcinolone acetonide and hydrocortisone 21-hemisuccinate (cortisol) on the acquired immunity and humoral antibody response to Ichthyophthirius multifiliis Fouque. J Fish Dis 13:269–280
Iguchi K, Ogawa K, Nagae M, Ito F (2003) The influence of rearing density on stress response and disease susceptibility of ayu (Plecoglossus altivelis). Aquaculture 220:515–523
Laemmli U (1970) Cleavage of structural proteins during assembly of head of bacteriophage-T4. Nature 227:680–685
Laidley CW, Woo PTK, Leatherland JF (1988) The stress response of rainbow trout to experimental infection with the blood parasite Cryptobia salmositicakatz, 1951. J Fish Biol 32:253–261
Lin TL, Clark TG, Dickerson HW (1996) Passive immunization of channel catfish (Ictalurus punctatus) against the ciliated protozoan parasite Ichthyophthirius multifiliis by use of murine monoclonal antibodies. Infect Immun 64:4085–4090
Luo XC, Xie MQ, Zhu XQ, Li AX (2007) Protective immunity in grouper (Epinephelus coioides) following exposure to or injection with Cryptocaryon irritans. Fish Shellfish Immunol 22:427–432
Nagae M, Fuda H, Ura K, Kawamura H, Adachi S, Hara A, Yamauchi K (1994) The effect of cortisol administration on blood plasma immunoglobulin M (IgM) concentrations in masu salmon (Oncorhynchus masou). Fish Physiol Biochem 13:41–48
Poole WR, Nolan D, Tully O (2000) Modelling the effects of capture and sea lice [Lepeophtheirus salmonis (Krøyer)] infestation on the cortisol stress response in trout. Aquacult Res 31:835–842
Rigos G, Pavlidis M, Divanach P (2001) Host susceptibility to Cryptocaryon sp infection of Mediterranean marine broodfish held under intensive culture conditions: a case report. Bull Eur Assn Fish Pathol 21:33–36
Ruane NM, Makridis P, Balm PHM, Dinis MT (2005) Skin darkness is related to cortisol, but not MSH, content in post-larval Solea senegalensis. J Fish Biol 67:577–581
Sigh J, Buchmann K (2001) Comparison of immobilization assays and enzyme linked immunosorbent assays for detection of rainbow trout antibody titres against Ichthyophthirius multifiliis Fouquet, 1876. J Fish Dis 24:49–51
Takemura A (1993) Changes in an immunoglobulin M (IgM)-like Protein during larval stages in tilapia, Oreochromis mossambicus. Aquaculture 115:233–241
Varsamos S, Flik G, Pepin JF, Wendelaar-Bonga SEW, Breuil G (2006) Husbandry stress during early life stages affects the stress response and health status of juvenile sea bass, Dicentrarchus labrax. Fish Shellfish Immunol 20:83–96
Wendelaar-Bonga SE (1997) The stress response of fish. Physiol Rev 77:591–625
Weyts FAA, Flik G, Rombout JHWM, Verburg-van Kemenade BML (1998) Cortisol induces apoptosis in activated B cells, not in other lymphoid cells of the common carp, Cyprinus carpio L. Dev Comp Immunol 22:551–562
Weyts F, Cohen N, Flik G, Verburg-van Kemenade BML (1999) Interactions between the immune system and the hypothalamopituitary-interrenal axis in fish. Fish Shellfish Immunol 9:1–20
Wiik R, Andersen K, Uglenes I, Egidius E (1989) Cortisol-induced increase in susceptibility of Atlantic salmon, Salmo salar to Vibrio salmonicida, together with effects on blood cell pattern. Aquaculture 83:201–215
Wray W, Boulikas T, Wray VP, Hancock R (1981) Silver staining of proteins in polyacylamide gels. Anal Biochem 118:197–203
Yambot AV, Song YL (2006) Immunization of grouper, Epinephelus coioides, confers protection against a protozoan parasite, Cryptocaryon irritans. Aquaculture 260:1–9
Yoshinaga T, Nakazoe J (1997) Acquired protection and production of immobilization antibody against Cryptocaryon irritans (Ciliophora, Hymenostomatida) in mummichog (Fundulus heteroclitus). Fish Pathol 32:229–230
Yoshinaga T, Akiyama K, Nishida S, Nakane M, Ogawa K, Hirose H (2007) In vitro culture technique for Cryptocaryon irritans, a parasitic ciliate of marine teleosts. Dis Aquat Organ 78:155–160
Acknowledgment
This research was supported by a grant from the Center for Tropical and Subtropical Aquaculture #2002-38500-12039 (USDA CREES) to Teresa Lewis and a grant from the National Science Foundation #EPS02-37065 to Jo-Ann Leong.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Misumi, I., Leong, JA.C., Takemura, A. et al. Immune protection of Mozambique tilapia (Oreochromis mossambicus) exposed to different infectious doses of ectoparasite (Cryptocaryon irritans). Parasitol Res 110, 363–372 (2012). https://doi.org/10.1007/s00436-011-2500-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-011-2500-7