
Abstract
Thirty-eight samples recovered from a latrine in the rural market town of Hadersdorf am Kamp (Lower Austria) and filled during the early 17th century were analysed and compared to the current bryophyte vegetation there and in its surroundings. Twenty-eight moss species from the latrine could be identified, and four taxa could be determined only to genus level because of missing sporophytes. The resulting data set is a valuable contribution to the state of knowledge of the bryophyte flora of that historic period, as none of the traditional methods such as herbaria date back to this early era. The most frequent species in the pit, Leucodon sciuroides, Antitrichia curtipendula and Neckera complanata, could hardly be found or not at all in the current bryophyte flora of the investigated area. A comparison of Ellenberg’s indicator values of the historic and current moss taxa revealed significant differences for temperature, humidity and continentality. Temperature values were much lower for the taxa found in the latrine. Furthermore, the historic mosses reflect a more humid and less continental climate than the ones growing nowadays. Many of the mosses found in the latrine are epiphytic or epilithic and could not be found in the investigated area today. These results indicate a different climate during the time of the Little Ice Age in the investigated area. According to the low total biomass of mosses in the pit (1–2 g per 10 l) we suggest that those found in the latrine arrived there mainly by chance, as in sweepings.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Bryophytes (liverworts, hornworts and mosses) are an important part of the vegetation both in terms of biodiversity and cover (Glime 2017). According to their morphology and physiology, they reflect changes in environmental conditions quickly. Therefore, they are generally accepted as good indicators for environmental monitoring (for example, Zechmeister et al. 2003; Tuba et al. 2011; Ares et al. 2012; Nickel et al. 2017).
Bryophytes from peat cores have been investigated to assess climatic changes, mainly in the Holocene period (Dickson 1986; Blundell and Holden 2015). However, there have hardly been any investigations into changes in bryophyte vegetation within the last 300 years. It is assumed that major changes in bryophyte diversity have been caused by changes in agricultural land use during the last three centuries (Vanderpoorten and Goffinet 2009). Since the beginning of the industrial revolution a loss in species diversity can be assigned to atmospheric air pollution too (Zvereva and Kozlov 2011). A loss or increase in bryophyte species diversity as a result of climate change has hardly been assessed by any bryological investigation covering a period before the last 50 years. There is little to no knowledge of changes in bryophyte vegetation dating back to the Little Ice Age in the 17th century. This could be a consequence of the lack of scientific bryophyte records from this period. Most herbarium collections of bryophytes start with samples of the late 18th century, as in the natural history museums in London and in Vienna. Therefore, the data set of this present study gives valuable insights into historic bryophyte vegetation.
There was a wide use of bryophytes (mainly mosses) in historical times for such purposes as medicine, toiletry, wound dressing material or fillings of mattresses (Richardson 1981). Only as a foodstuff, there was obviously no use for them (Glime 2017). Mosses were collected from the immediate vicinity and often ended up in the rubbish after use, which was deposited on muckheaps, in rivers and also in latrines. Therefore, bryophytes in these rubbish pits seem to be a valuable indicator for historic habits and/or historic environments. Nevertheless, with the exception of wet soils or specific circumstances of preservation such as in latrines, bryophytes are not preserved in most cases due to their decomposition in dry soils or disintegration in the predominantly charred archaeobotanical material (Jacomet and Kreuz 1999).
By investigating the filling of a latrine deposited in the 17th century in a historical building in Hadersdorf am Kamp, Austria, we tried to address the question whether there is any evidence for a change in bryophyte species diversity due to climatic changes. Furthermore, we attempted to solve the question of where the filling material came from and if any kinds of use for this material could be detected.
Methods
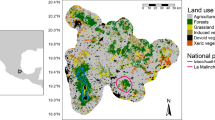
The rural village of Hadersdorf am Kamp is located in Niederösterreich (Lower Austria) (48°45′ N, 15°72′ E; 202 m a.s.l.; Fig. 1). It is situated at the western edge of the Pannonian climate thermophilic vegetation zone, and vineyards dominate the landscape today.

Location of the investigated area around Hadersdorf am Kamp, Niederösterreich (Lower Austria); mapping from Geoland Basemap Orthofoto (2016)
During restoration work in the town hall, a well or latrine was discovered and excavated in 1991/92 in a rescue excavation by Bundesdenkmalamt Österreich, the national heritage agency. The feature was 4.5 m deep and 1.5 m in diameter on average, as the pit was slightly conical. The core was built as a well and had been reused as a latrine in the 17th century for unknown reasons. Latrines of this period were mostly used both as toilets as well as rubbish pits. Plenty of household rubbish and other artefacts were found (Sauer 1992). Beside archaeological artefacts (bricks, ceramics, glass and metallic objects) and archaeozoological remains (animal bones and fish remains), leather (fragments of shoes), wood and other plant remains (seeds/fruits, mainly uncharred) were also visible to the naked eye. Moreover, crystals of struvite (magnesium ammonium phosphate) were found in the Hadersdorf latrine, which usually derive from excrement and urine. The lowermost 70 cm of the complex was waterlogged. For sampling, the pit fill was divided into eight artificial strata. The deposited material had a volume of 3.5 m3. Whereas big artefacts and objects were sorted out during digging, the whole sediment and smaller remains were sampled and processed in 207 sediment samples each of 10 l and wet sieved into size fractions using a smallest mesh size of 0.8 mm. Subsequently all fractions were preserved in Tego 51B.
Investigations for the present study started in 2016 with an emphasis on 38 samples (15% of the overall material) taken from the various strata. Bryophyte samples were washed with water and transferred into a mixture of ethanol, glycerol and distilled water with added phenol and stored in small glass vials (Scharrer-Liška and Popovtschak 2017). Specimens were identified to species level if possible. The amount of moss was approximately 1–2 g per 10 l sediment. The state of preservation was very good and bryophytes were not decomposed or only slightly. Nomenclature follows Köckinger et al. (2018).
For a comparison of the historic material with the current flora of the Hadersdorf area, we tried to record all bryophytes (mosses and liverworts) growing in the vicinity of the town hall. These investigations covered the market place of Hadersdorf itself (with parks, very old trees and alleys) and the surrounding areas including woods, vineyards, dry meadows and a valley, the Straßertal.
A comparison of ecological requirements of the historic and the current bryophytes was made using Ellenberg’s indicator values (Ellenberg et al. 1992). We used two ways to calculate them, unweighted, in which each taxon is represented once, and weighted calculation. We weighted the values according to the frequency of their occurrence. For the historic samples, the frequency of occurrences in the 38 samples was taken, and for the recent samples the distribution as given, both in Table 1.
Statistical tests using these values were the F test for variance, a Kolmogorov–Smirnov test for comparing the distribution of these two data sets and a Mann–Whitney test for comparing the mean indicator values (p < 0.05).
Results
Bryophyte diversity in the latrine
The examined bryophytes were in a good state of preservation and could be identified to species level in most cases, resulting in 28 species and four taxa which could be identified only to genus level because of missing sporophytes. In Table 1 the bryophyte taxa, their number of occurrences in the various samples and their main habitats are listed.
All of these historic taxa can be ranked in the class of mosses (Bryopsida) and none of the bryophytes belong to liverworts (Marchantiopsida) or hornworts (Anthoceratopsida).
The distribution of mosses in the various samples is uneven. A small number of taxa could be found in many of the samples, of which the most frequent was Leucodon sciuroides, occurring in 34 out of 38 samples. In Austria, this species is a vigorously growing trunk epiphyte on free-standing trees, and in Pannonian areas it can also be found on rocks. Next most frequent was Neckera complanata, which grows in similar but slightly moister sites than L. sciuroides. Surprisingly high was the frequency of Antitrichia curtipendula (14 samples). In pre-industrial times, this species grew mainly as a trunk epiphyte in central Europe (Frahm 2011), but nowadays it is rare and found mainly on shaded rocks in Austria.
Generally, most of the less frequent mosses found in the latrine grow on dry, sunny soils in dry grasslands, vineyards or pastures. Among these only Homalothecium lutescens (8 samples) and Abietinella abietina (6 samples) were more common. Many taxa occurred only once or in a very few samples. Among these Scleropodium tourettii is most notable, now a Mediterranean species, which has no recent occurrence and has never been found in Niederösterreich (Lower Austria) before.
Distribution within strata
Comparing the eight investigated strata, no differences were found for bryophyte distribution. Frequent taxa (n > 5) could be found from the deepest to the uppermost strata. This reflects either a quick filling of the pit and/or an even distribution (and use) of these mosses within the area.
Comparison of historic and recent bryophyte distribution
During the search for current bryophytes, 50 species (47 mosses and three liverworts) could be found (Table 1) compared to 28 moss species and 24 specimens identified only to genus level in the historic samples. In both the historic and recent samples we found epiphytes (growing on plants), epilithic mosses (on stone) and ones growing on soils. In the historic samples vigorously growing taxa dominated, whereas smaller bryophytes dominated the recent samples.
The most frequent species in the historic records, Leucodon sciuroides, could be found only once in the recent sampling. It did not grow in the market town itself, but close by, in the Straßertal. Other vigorously growing, weft forming species like Antitrichia curtipendula, Neckera crispa and N. pumila could either not be found in the recent sampling or only with a very low frequency, for example Leucodon sciuroides, Neckera complanata and Anomodon attenuatus, although substrates for their growth were still available, such as single trees, alleys, low growing oak woods or shaded siliceous rocks. The only species occurring in both collections more frequently was the ubiquitous Hypnum cupressiforme.
Although several mosses of open soils were detected both in the historic and current samples, such as Abietinella abietina, Oxyrrhynchium hians, Homalothecium lutescens and H. sericeum, a change in taxa and frequency could be observed. Some historic mosses which need moist conditions could not be found in the recent investigation (Calliergonella cuspidata, Hylocomium splendens, Rhytidiadelphus triquetrus), whereas recent bryophytes had an emphasis on dryer conditions (Aloina spp., Pottia spp., Syntrichia ruralis).
Ecology of bryophytes
The trends of both calculations (weighted and unweighted) were the same. By using unweighted calculations significant differences could be shown for temperature, whereas weighted calculations showed significant differences for all parameters except for the pH value. Weighting according to frequencies minimises the influence of rare or neglected taxa. Therefore, rare bryophytes in the recent sampling and very small ones, which might have been missed in the historic samples, did not strongly influence the results. In consequence we present only the results of the weighted calculation.
Figure 2 shows mean Ellenberg indicator values, both for mosses from the latrine as well as for those from the recent sampling. These values are given for light, temperature, continentality, humidity and soil pH value (Ellenberg et al. 1992).

Comparison of Ellenberg’s indicator values for all moss taxa found in the latrine samples and collected in recent fieldwork. L light indicator value, T temperature indicator value, C continentality indicator value, H humidity indicator value, R substrate pH indicator value, h historic samples, r recent collection
Light value for the historic bryophytes was 6.3 (± 1.9) compared to 6.9 (± 1.7) for current ones, which reflects semi-shaded to partially shaded conditions, and is typical for Neckera species, for example. Most epiphytic taxa can be ranked within these values. The difference is weakly significant (p = 0.015).
The average temperature value for the latrine mosses was 3.8 (± 0.9) compared to 4.4 (± 1.2) for current bryophytes. The difference is strongly significant (p = 0.0003). Historic values reflect a cool temperate climate, which comes as a surprise, as Hadersdorf is situated in the Pannonian area now. Values for taxa from the recent collection were markedly higher.
Climatic continentality for the latrine mosses was 4.9 (± 0.6) compared to 5.2 (± 0.6) for taxa growing nowadays. The difference is significant (p = 0.006). The present-day value reflects conditions between a suboceanic and a subcontinental climate and is typical for the central European flora with a slight eastern emphasis—the historical one is slightly more oceanic.
The average humidity indicator value for the mosses in the latrine was 3.9 (± 1.1) compared to 3.4 (± 1.4) for current bryophytes. The difference is strongly significant (p = 0.0007). Current mosses reflect dryer site conditions whereas the historic ones indicate damper conditions.
Average reaction indicator (soil pH value) was 6.2 (± 1.2) for latrine mosses compared to 6.2 (± 1.3). There is no statistically significant difference between these two values. Since both investigated periods showed comparable values, site pH and therefore substrates are comparable in both periods. The value reflects weakly acid conditions, which can be found on tree trunks or on the gneissic rocks in the surroundings of the investigated market town.
Discussion
Most of the taxa which were abundant in the latrine samples are rare or absent in the area of Hadersdorf today (Antitrichia curtipendula, Leucodon sciuroides, Neckera complanata, N. crispa, Hylocomium splendens or Rhytidiadelphus triquetrus) although their required substrates are sufficiently available in the modern environment. All of them favour a more humid and cooler climate than today’s. Temperature values of historic samples (3.8) were significantly lower than the mean temperature value of the recent collection (4.4), the same goes for light, humidity and in consequence also continentality.
Most remarkable is the frequent occurrence of Antitrichia curtipendula in the past, as nowadays it cannot be found in a wide area around Hadersdorf (Zechmeister et al. 2013). The nearest places where it grows now are situated either 50 km north of Hadersdorf at higher elevations with a cooler climate, or in areas with extremely high precipitation in the Alps. A comparable extinction of this species is also reported from other parts of central Europe (Frahm 2011). It is concluded that A. curtipendula disappeared mainly due to industrial pollution in many countries since the beginning of the industrial revolution (Frahm and Wiethold 2004). In fact, air pollution had detrimental effects on the distribution of many bryophytes (Franzen 2001; Zvereva and Kozlov 2011; Dittrich et al. 2016). A decrease or extinction of bryophytes could be mainly attributed to SO2 emissions, one of the most important pollutants over the last 150 years, but which have decreased dramatically in recent decades in Europe (Umweltbundesamt 2018); a return of bryophytes into formerly heavily polluted areas has been observed (Sergio et al. 2016). At present, pollutants like NOx or NH4 still have strong effects on bryophyte diversity, facilitating nitrophiles, among these luxuriant growing species like L. sciuroides or Hypnum cupressiforme while discouraging others (Zechmeister et al. 2007). Recolonisation after disappearance is also a matter of diaspore dispersal and establishment. Ahrens (2007) shows for Leucodon sciuroides that the frequency of sexual reproduction decreased since historical times. It now occurs only in a temperate valley in the investigated area and not in the near vicinity of Hadersdorf, where the climate is slightly warmer and drier. The sensitivity of L. sciuroides to atmospheric pollution is discussed controversially (Frahm 1998; Sauer 2000). Assuming that it is sensitive to pollution, diaspore dispersal could have been an intrinsic factor, otherwise climate change (temperature, humidity) could have caused its absence. The fact that L. sciuroides as well as A. curtipendula can be found in areas nowadays which were heavily polluted before, as in northwestern Niederösterreich, but which have a substantially cooler climate than the investigated area, suggests that the climate theory is correct. Therefore, we assume that at least L. sciuroides would occur more frequently in the Hadersdorf area if the climate was suitable for it there.
Landscape structure is another important cause of species diversity. One could assume that the structure of the historic landscape was different from today’s, which would have led to a different bryophyte vegetation (Vanderpoorten and Engels 2003; Pharo and Zartman 2007). The area around Hadersdorf am Kamp is still a small scale structured landscape offering appropriate habitats for those species which have vanished since the 17th century. Beside vineyards, arable land and open broad-leaved woodlands are the most common elements of landscape structure today, even alleys and siliceous rocks can be found. All these would still provide plenty of adequate substrates for the frequent taxa in the historic samples.
Taking into account all these factors we attribute the observed change in moss taxa mainly to climate differences since the 17th century and we assume that the historical climate was significantly wetter and cooler than the present one. Historic climatic data are not available for the investigated area, however our hypothesis is underpinned by overall climate change models, which report an average temperature during the 16th and 17th centuries in Europe which was 2.5 °C lower than today (von Storch et al. 2004; Glaser and Riemann 2009). This Little Ice Age period is documented not only by scientists but also in art and in social research (Fagan 2002; Roelofs 2010). Our data confirm a cooler and wetter climate in the early 17th century in the investigated area by using bryophytes and their ecological needs. The climatic conditions at that time were probably the main cause of a vigorous growth of large pleurocarp bryophytes on tree trunks in the village and on rocks in the surrounding hills, and in valleys such as Straßertal.
We postulate that the historic samples represent the local bryophyte flora from that time very well. Although the samples were dominated by larger mosses, small acrocarps (with upright growth, mostly unbranched) could be found regularly. It is unlikely that these tiny bryophytes were collected for a specific purpose due to their small size. They were obviously deposited by chance, as in sweepings. Furthermore, comparable indicator values for substrate (R values) suggest that the historic samples derive from the same area as the recent ones. That the historic mosses grew in the latrine itself is very unlikely. Considering their humidity (3.9 ± 1.1) and light requirements 6.3 (± 1.86), they probably grew on trees or stone surfaces.
The question about the use of the mosses however remains speculative. In the latrine, mainly larger sized mosses were found, which could be evidence that they were gathered selectively for special purposes. Large mosses have often been recovered from ancient latrines and some of the same taxa have been found in our investigation. Dickson (2000) and van den Bos et al. (2014) refer to the use of Neckera crispa und N. complanata as a filling for mattresses. Glime (2017) reports on the use of mosses for sanitary products as well as for preserving food, while Constantin and Kuijper (2002) suggest use as cleaning materials. In the Hadersdorf latrine, mosses were distributed regularly through all strata, which indicates either regular use or at least regular deposition of moss. The total biomass of mosses in the latrine was small (1–2 g per 10 l). Therefore intensive or regular, intended uses, such as toilet paper, are not obvious so far. Given the size and volume of the Hadersdorf latrine, mosses were probably used every now and then, probably for reasons reported in literature, as a sanitary product, for instance.
References
Ahrens M (2007) Wachstum und Reproduktion des epiphytischen Laubmooses Leucodon sciuroides in der nördlichen Oberrheinebene (Südwestdeutschland). Carolinea 65:69–103
Ares A, Aboal JR, Carballeira A, Giordano S, Adamo P, Fernández JA (2012) Moss bag biomonitoring: a methodological review. Sci Total Environ 432:143–158
Blundell A, Holden J (2015) Using palaeoecology to support blanket peatland management. Ecol Indic 49:110–120
Constantin C, Kuijper WJ (2002) Utilisation de mousse comme dégraissant dans des céramiques néolithiques de France et de Belgique. Bull Soc Préhis Franc 99:775–783
Dickson JH (1986) Bryophyte analysis. In: Berglund BE (ed) Handbook of Holocene palaeoecology and palaeohydrology. Wiley, Chichester, pp 627–643
Dickson JH (2000) Bryology and the Iceman: Chorology, ecology, and ethnobotany of the mosses Neckera complanata Hedw. and N. crispa Hedw. In: Bortenschlager S, Oeggl K (eds) The Iceman and his natural environment: palaeobotanical results. Springer, Wien, pp 77–87
Dittrich S, Leuschner C, Hauck M (2016) Change in the bryophyte diversity and species composition of Central European temperate broad-leaved forests since the late 19th century. Biodivers Conserv 25:2,071–2,091
Ellenberg H, Weber HE, Düll R, Wirth V, Werner W, Paulißen D (1992) Zeigerwerte von Pflanzen in Mitteleuropa. Scr Geobot 18:9–166
Fagan BM (2002) The little ice age: how climate made history. Basic Books, New York, pp 1,300–1,850
Frahm J-P (1998) Moose als Bioindikatoren. Quelle & Mayer, Wiesbaden
Frahm J-P (2011) Moose aus einer frühneuzeitlichen Latrine in Münster (Westfalen). Arch Bryol 82:1–6
Frahm J-P, Wiethold J (2004) Die Moosflora des Mittelalters und der Frühen Neuzeit in Mitteleuropa, nach archäologischen Funden zusammengestellt. Herzogia 17:303–324
Franzen I (2001) Epiphytische Moose und Flechten als Bioindikatoren der Luftqualität am Westrand des Ruhrgebietes. Limprichtia 16:1–85
Geoland Basemap (2016) basemap.at. https://www.basemap.at/. Accessed 20 Nov 2018
Glaser R, Riemann D (2009) A thousand-year record of temperature variations for Germany and Central Europe based on documentary data. J Quat Sci 24:437–449
Glime JM (2017) Bryophyte ecology, Chap. 1–1: Household and personal uses. Ebook sponsored by Michigan Technological University and the International Association of Bryologists. http://digitalcommons.mtu.edu/bryophyte-ecology. Accessed 5 October 2017
Jacomet S, Kreuz A (1999) Archäobotanik. Aufgaben, Methoden und Ergebnisse vegetations- und agrargeschichtlicher Forschung. Ulmer, Stuttgart
Köckinger H, Schröck C, Krisai R, Zechmeister HG (2018) Checkliste der Moose Österreichs. http://cvl.univie.ac.at/projekte/moose. Accessed 25 Nov 2018
Nickel S, Schröder W, Wosniok W et al (2017) Modelling and mapping heavy metal and nitrogen concentrations in moss in 2010 throughout Europe by applying Random Forests models. Atmos Environ 156:146–159
Pharo EJ, Zartman CE (2007) Bryophytes in a changing landscape: The hierarchical effects of habitat fragmentation on ecological and evolutionary processes. Biol Conserv 135:315–325
Richardson DHS (1981) The biology of mosses. Blackwell, Oxford
Roelofs P (2010) Hendrick Avercamp: master of the ice scene. Rijksmuseum, Amsterdam
Sauer F (1992) KG Hadersdorf am Kamp. Fundberichte aus Österreich 31:14
Sauer M (2000) Moose als Bioindikatoren. In: Nebel M, Philippi G (eds) Die Moose Baden-Württembergs, vol 1. Ulmer, Stuttgart, pp 28–31
Scharrer-Liška G, Popovtschak M (2017) Alltag und Entsorgungspraktiken im ländlichen Niederösterreich der Frühzeit. Projektbericht, Universität Wien
Sérgio C, Carvalho P, Garcia CA et al (2016) Floristic changes of epiphytic flora in the metropolitan Lisbon area between 1980–1981 and 2010–2011 related to urban air quality. Ecol Indic 67:839–852
Tuba Z, Slack NG, Stark LR (2011) Bryophyte ecology and climate change. Cambridge University Press, Cambridge
Umweltbundesamt (2018) Schwefeldioxid. http://www.umweltbundesamt.at/umweltsituation/luft/luftschadstoffe/so2/. Accessed 26 Oct 2018
Van den Bos V, Brinkkemper O, Bull ID et al (2014) Roman impact on the landscape near castellum Fectio, The Netherlands. Veget Hist Archaeobot 23:277–298
Vanderpoorten A, Engels P (2003) Patterns of bryophyte diversity and rarity at a regional scale. Biodivers Conserv 12:545–553
Vanderpoorten A, Goffinet B (2009) Introduction to bryophytes. Cambridge University Press, Cambridge
Von Storch H, Zorita E, Jones JM, Dimitriev Y, González-Rouco F, Tett SFB (2004) Reconstructing past climate from noisy data. Science 306:679–682
Zechmeister HG, Grodzińska K, Szarek-Łukaszewska G (2003) Bryophytes. In: Markert BA, Breure AM, Zechmeister HG (eds) Bioindicators and biomonitors: principles, concepts and applications. Elsevier, Amsterdam, pp 329–375
Zechmeister HG, Dirnböck T, Hülber K, Mirtl M (2007) Assessing airborne pollution effects on bryophytes – Lessons learned through long-term integrated monitoring in Austria. Environ Pollut 147:696–705
Zechmeister HG, Hagel H, Gendo A et al (2013) Die Rote Liste der Moose Niederösterreichs. Wiss Mitt Niederösterr Landesmuseum 24:7–126
Zvereva E, Kozlov M (2011) Impacts of industrial polluters on bryophytes: a meta-analysis of observational studies. Water Air Soil Pollut 218:573–586
Acknowledgements
Open access funding provided by University of Vienna. The project was financed by Niederösterreichische Landesregierung (Lower Austrian Government) (K1-WF-1152/002-2017) and the deanery of the Faculty of Historical and Cultural Studies at the University of Vienna.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by K.-E. Behre.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Zechmeister, H.G., Kropik, M., Popovtschak, M. et al. Bryophytes in a latrine as indicators of climate change in the 17th century. Veget Hist Archaeobot 28, 575–581 (2019). https://doi.org/10.1007/s00334-019-00717-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-019-00717-6