
Abstract
Locomotion accounts for a significant proportion of the energy budget in birds, and selection is likely to act on its economy, particularly where energy conservation is essential for survival. Birds are capable of different forms of locomotion, such as walking/running, swimming, diving and flying, and adaptations for these affect the energetic cost [cost of locomotion (CoL)] and kinematics of terrestrial locomotion. Furthermore, seasonal changes in climate and photoperiod elicit physiological and behavioural adaptations for survival and reproduction, which also influence energy budget. However, little is understood about how this might affect the CoL. Birds are also known to exhibit sex differences in size, behaviour and physiology; however, sex differences in terrestrial locomotion have only been studied in two cursorially adapted galliform species in which males achieved higher maximum speeds, and in one case had a lower mass-specific CoL than females. Here, using respirometry and high-speed video recordings, we sought to determine whether season and sex would affect the CoL and kinematics of a principally aquatic diving bird: the circumpolar common eider (Somateria mollissima). We demonstrate that eiders are only capable of a walking gait and exhibit no seasonal or sex differences in mass-specific CoL or maximum speed. Despite sharing identical limb morphometrics, the birds exhibited subtle sex differences in kinematic parameters linked to the greater body mass of the males. We suggest that their principally aquatic lifestyle accounts for the observed patterns in their locomotor performance. Furthermore, sex differences in the CoL may only be found in birds in which terrestrial locomotion directly influences male reproductive success.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Balancing daily energy expenditure with intake is essential to the evolutionary fitness of animals (Goldstein 1988). Locomotor activity consumes a significant proportion of an animal’s daily energy budget (Goldstein and Nagy 1985; Elliott et al. 2013), meaning the energetic cost of locomotion (CoL) is likely to be under selection pressure. Balancing these costs is paramount, particularly in variable climates where energy conservation is essential to survival or reproduction (Tolkamp et al. 2002; Lees et al. 2010). For many species of bird, terrestrial locomotion is of critical importance to fitness at some point in their life history, for example during mating, feeding, incubation or wing moult when these animals may be restricted to moving on the ground (Nudds et al. 2010; Lees et al. 2011; Portugal and Guillemette 2011).
Terrestrial gaits are classified by both duty factor (DF, the proportion of a stride during which each foot is in contact with the ground) and the phasing of the mechanical energy fluctuations of the centre of mass (CoM). In walking gaits (DF >0.5), each stride has a phase with both feet on the ground, and the kinetic and potential energies of the CoM cycle 180° out of phase. In aerial running gaits (DF ≤0.5), each stride has a phase with both feet off the ground, and these energies cycle in phase. An intermediate gait, grounded-running, is also utilised by some birds, which has no aerial phase but has in phase energy cycling characteristic of aerial running (Gatesy and Biewener 1991; Rubenson et al. 2004; Hancock et al. 2007; Usherwood et al. 2008; Nudds et al. 2011). During terrestrial locomotion, a bird’s rate of energy metabolism, the cost of locomotion, increases as a function of speed (U) until a morphological (mechanical) or physiological (energetic) constraint is met (Brackenbury and Avery 1980; Roberts et al. 1998; Nudds et al. 2010). This increase is often linear (Pinshow et al. 1977; Bamford and Maloiy 1980; Taylor et al. 1982; Brackenbury and Elsayed 1985; Roberts et al. 1998; White et al. 2008); however, nonlinearity within and between gaits is also common (Rubenson et al. 2004, 2007; Watson et al. 2011; Nudds et al. 2011). Elevated energy metabolism at faster U correlates with shorter periods of foot–ground contact, which require higher rates of force production by muscle fibres (Kram and Taylor 1990; Roberts et al. 1998). The minimum energy required to move a unit body mass (M b) over a unit distance, the minimum cost of transport (CoTmin, J kg−1 m−1), is inversely proportional to M b (roughly, CoTmin = 10.8 M −0.32b ) across numerous species of different taxa (Taylor et al. 1982; Kram and Taylor 1990; Full and Tu 1991; Roberts et al. 1998; Nudds et al. 2009). Longer foot–ground contact durations that result from the longer legs of larger species and their more upright limb postures are thought to allow the recruitment of slower more efficient muscle fibres and improve the mechanical advantage of the limbs, respectively (Biewener 1989; Kram and Taylor 1990). Whereas more cursorially specialised birds, such as galliformes and ratites, tend to have CoTmin equal to, or less than, allometric predictions (Taylor et al. 1982; Roberts et al. 1998; Watson et al. 2011), studies on birds with aquatic specialisation have arrived at differing conclusions as to whether these birds have CoTmin equal to (White et al. 2008), or greater than allometric predictions (Fedak et al. 1974; Griffin and Kram 2000; Nudds et al. 2010). Furthermore, some (Griffin and Kram 2000; White et al. 2008; Nudds et al. 2010) but not all (Usherwood et al. 2008) dabbling and diving species are incapable of aerial running gaits and have a limited maximum attainable U. Such limitations are thought to be linked to the derived morphologies of these birds, including short and caudad hindlimbs relative to cursorial birds (Griffin and Kram 2000; Nudds et al. 2010). However, given the observed variation in locomotor ability between birds with aquatic specialisation, it is clear that a more complex combination of factors determines CoTmin and gait selection in birds.
Understanding how seasonal changes in environmental conditions may influence the CoL is complicated. At higher latitudes, seasonal variations in photoperiod elicit changes in avian physiology associated with energetically costly behaviours such as migration, egg-laying, incubation, wing moult and feather production (Piersma et al. 2003; Guillemette et al. 2007; Portugal and Guillemette 2011; Guillemette and Butler 2012; Guillemette et al. 2012). Furthermore, changes in climate may influence predation pressures, resource acquisition and thermoregulation, which may affect energy expenditure (Weathers and Sullivan 1993). Birds are known to experience seasonal variation in daily energy budget, but only one study (Lees et al. 2010) has considered the effect of changing season upon the CoL. The Svalbard rock ptarmigan (Lagopus muta hyperborea) almost doubles in M b prior to winter, acquiring significant fat stores that serve as thermal insulation and as an energy source (Stokkan et al. 1995). Interestingly, despite the additional weight in winter, ptarmigan have a reduced mass-specific CoL compared to summer, perhaps as an adaptation for energy conservation in cold and resource-limited Arctic winters (Lees et al. 2010). As this represents the only study to date, the ubiquity of seasonal influences on the CoL in birds is unclear.
Sex may also influence the CoL in birds. In species in which paternal care is minimal and males compete for females, sex differences in morphology, physiology and locomotor behaviours are common (Portugal and Guillemette 2011). For example, males often possess ornamental and colourful plumage and may be larger, compared to less conspicuous females (Dunn et al. 2001; Lislevand et al. 2009). Energetically costly courtship, territorial or lekking displays are exclusive to male birds giving rise to sex differences in energy budget (Unander and Steen 1985; Vehrencamp et al. 1989; Barske et al. 2011). Furthermore, egg formation, incubation and parental care require significant metabolic investment from only females (Parker and Holm 1990). It is therefore surprising that of the many studies on the CoL in such birds, only two have considered sex differences (Brackenbury and Elsayed 1985; Lees et al. 2011). Male domestic chickens (Gallus gallus domesticus) sustained higher maximum U than females (Brackenbury and Avery 1980; Brackenbury and Elsayed 1985), and both sexes shared an identical mass-specific CoL, despite the males being 56 % heavier (Brackenbury and Elsayed 1985). Similarly, male Svalbard rock ptarmigan, only 5 % heavier than females, sustained higher maximum U than females but, surprisingly, had a significantly lower mass-specific CoL, which could not be explained by differences in hindlimb kinematics (Lees et al. 2011). Lees et al. (2011) suggested that the lower mass-specific CoL in male ptarmigan might be an advantage for territorial behaviour, which may influence male reproductive success if females select males based upon their ability to defend territories.
Since seasonal and sex differences in the CoL have only been investigated in primarily cursorial birds, we consider the CoL, hindlimb kinematics and morphometrics of a principally aquatic diving species. The common eider (Somateria mollissima) is a large, wing- and foot-propelled diving duck (Heath et al. 2006) inhabiting coastal marine areas of temperate, boreal and Arctic climatic zones (Blix 2005). Terrestrial locomotion is only utilised by these birds during spring breeding and the 24–26 days of summer nesting, but is essential for their survival and reproduction (Portugal and Guillemette 2011). Daily energy expenditure varies seasonally in eiders with high energetic costs associated with synchronous wing moult and feather production during summer and thermoregulation on cold waters during winter (Guillemette and Butler 2012). However, neither sex gains M b as an adaptation to winter survival. Eiders are sexually dimorphic, with a small male bias in size [M b ~ 16 % heavier (Dunning 2008)]. Courtship displays by males are performed only on the water (Johnsgard 1964; Spurr and Milne 1976). Females are sole providers of parental care, and as an adaptation to this investment, they gain around 26 % M b prior to nesting (Rigou and Guillemette 2010), before losing 30–40 % pre-laying M b while fasting throughout laying and incubation (Parker and Holm 1990; Criscuolo et al. 2002; Portugal and Guillemette 2011). We hypothesise that the adaptive diving morphology of the eiders will limit their terrestrial locomotion compared to more cursorially specialised birds. Furthermore, given the limited amount of time these birds spend on the land, we predict that selection is not likely to drive any seasonal or sex differences in mass-specific CoL. Evidence to support this would suggest that locomotor specialisation has a strong influence over seasonal and sex-specific adaptations in these birds.
Materials and methods
Study species
Common eiders were hatched from the eggs of wild birds collected in Tromsø, Norway and reared outdoors at the Department of Arctic and Marine Biology, University of Tromsø, Norway. Sexually mature (3 years and above) males (n = 5: summer M b = 1.95 ± 0.05 kg; winter M b = 1.86 ± 0.05 kg, mean ± SE) and females (n = 4: summer M b = 1.79 ± 0.06 kg; winter M b = 1.62 ± 0.05 kg, mean ± SE) were housed in a large indoor enclosure (6 m2) with access to a pond and unlimited food and water. Artificial light conditions matched those in Tromsø (69°46′N) throughout the year (ranging from continuous light in May–August to 4.5 h of light per day in December) to ensure the natural seasonal physiological changes of the birds. Housing and experimental temperatures were kept within the species-specific thermoneutral zones (Gabrielsen et al. 1991). Birds were trained and acclimatised to all experimental conditions for 1 week prior to any data collection. Training consisted of the birds exercising for 5–10 min at four different speeds up to the maximum they could sustain (1.25 m s−1) each day. The same individual birds were used for both summer and winter experiments. The birds were not fasted prior to experiments.
Ethics statement
All experimental procedures were covered by a UK Home Office project licence (40/3001) held by Dr Codd and performed under ethical approval of the National Animal Research Authority of Norway (NARA, permit number 4884) and the University of Manchester.
Respirometry
Open-flow respirometry was used to measure the rates of O2 consumption (\(\dot{V}_{{{\text{O}}_{2} }}\); ml min−1) and CO2 production (\(\dot{V}_{{{\text{CO}}_{2} }}\); ml min−1) of eiders standing at rest and exercising within a Perspex® respirometry chamber (0.53 m tall, 0.45 m wide, 0.79 m long; volume = 188.40 L). The chamber was positioned over a motorised treadmill (Bremshey Trail Sport, Finland). Air was drawn from the front of the chamber at a fixed flow rate of 157 L min−1 using a vacuum pump. The main flow was then sub-sampled at a flow rate of 6 L min−1 into a carboy, from which a smaller sub-sample was drawn at a rate of 0.11 L min−1 for gas analysis. Water vapour pressure of the sub-sample was measured downstream using an RH300 humidity meter (Sable Systems International, Las Vegas, USA) before it was scrubbed of H2O using calcium chloride with indicator (Merck, Darmstadt, Germany) and passed on to a FoxBox-C field gas analysis system (Sable Systems international, Las Vegas, USA) for CO2 analysis. Air was subsequently scrubbed of CO2 using soda lime with indicator (Sigma-Aldrich, Steinheim, Germany) and pumped back to the gas analysis system for O2 analysis. The equipment set-up was identical for both summer and winter experiments. Room temperature during trials was 18.63 ± 0.47 °C in summer and 14.00 ± 1.00 °C in winter. The accuracy of the respirometry set-up (±2 % across all treadmill speeds) was validated using a N2 injection test (Fedak et al. 1981).
The primary flow rate (FR) was adjusted to a dry-corrected flow rate (FRc) to account for the loss of water from the sample prior to O2 and CO2 analysis using Eq. 1 (Lighton 2008):
where BP is barometric pressure and WVP is water vapour pressure. \(\dot{V}_{{{\text{O}}_{2} }}\) was calculated using Eq. 2 (Lighton 2008):
where ΔO2 is the difference between incurrent and excurrent O2 concentrations. \(\dot{V}_{{{\text{CO}}_{2} }}\) was then calculated using Eq. 3 (Lighton 2008):
where ΔCO2 is the difference between incurrent and excurrent CO2 concentrations.
Respiratory exchange ratios (RERs) were calculated as the ratio \(\dot{V}_{{{\text{CO}}_{2} }}\):\(\dot{V}_{{{\text{O}}_{2} }}\), and rates of oxygen consumption were then converted to rates of energy metabolism (J s−1) using RERs and thermal equivalents taken from Table 12.1 of (Brody 1945). Rates of energy metabolism were divided by individual M b to give mass-specific metabolic power consumption (P met, W kg−1). Net mass-specific metabolic power consumption (net-P met) is the rate of energy metabolism required for locomotion above that required for standing quietly and was calculated by subtracting resting metabolic rate (RMR, W kg−1) from locomotor P met. Total mass-specific cost of transport (CoTtot, J kg−1 m−1) was calculated at each speed class by dividing P met by U and net mass-specific cost of transport (CoTnet) by dividing net-P met by U. The minimum cost of transport (CoTmin) was determined using two methods: (1) by taking the slope of the relationship between P met (or net-P met) and U (Taylor et al. 1982); and (2) by reporting the minimum recorded value of CoTtot (or CoTnet) (Rubenson et al. 2007; Langman et al. 2012).
Birds were exercised on a treadmill at seven U increments up to their maximum sustainable U (0.22, 0.36, 0.50, 0.64, 0.80, 1.00 and 1.25 m s−1). Data were collected for three speeds during each trial, and speed order was randomised. Trials began by allowing birds to settle within the chamber until O2 concentration traces plateaued for at least 2 min, indicating a steady resting \(\dot{V}_{{{\text{O}}_{2} }}\). Steady \(\dot{V}_{{{\text{O}}_{2} }}\) was also used to determine the duration of exercise at each U, which was between 5 and 10 min. Each U was followed by a period of rest for a minimum of 5 min until the trace was again stable, indicating the birds had fully recovered. RMR values were taken from the final rest period of each trial and were used to calculate a mean RMR for each bird.
Kinematics
High-speed video recordings (100 frames s−1) were taken of all trials from a lateral view using a SONY Handycam HDR-XR520 (Japan). Quantitative analyses of kinematic parameters were conducted by tracking movements of the left foot (closest to the camera) using Tracker software v. 4.05 (Open Source Physics). Measured parameters were duty factor (DF), the durations of both the stance (t stance, s) and swing (t swing, s) phases of the limb during a stride, stride frequency (f stride, Hz) and stride length (l stride, m). A minimum of seven strides was analysed for each bird under each U condition when a constant U and position on the treadmill belt was maintained.
Morphometric measurements
The length of the right tarsus was measured from each of the study birds using a measuring rule (±1 mm). Femur, tibiotarsus and tarsometatarsus lengths and pelvis widths (shortest distance between the left and right acetabulums) were also measured from 5 male and 5 female adult common eider skeletal specimens from the National Museum of Scotland’s Collections Centre (Museum refs: 1995.190.17, 29, 33, 35, 39, 41, 45, 49, 51 and 53) using digital vernier callipers (±0.01 mm).
Statistical analyses
Results are represented as mean ± SE for each of the four groups: summer males (S M), summer females (S F), winter males (W M) and winter females (W F). The relationships between dependent variables (metabolic and kinematic parameters) and U were tested for differences between seasons and sexes using general linear models (GLMs). Season and sex were included in the models as fixed factors and U as a covariate. The slopes of the relationships between dependent variables and U were firstly tested for differences between seasons and between sexes (GLM1): if any interaction terms (sex × U/season × U) were non-significant (indicating similar slopes), they were removed from the GLM in a subsequent rerun (GLM2), assuming identical slopes, in order to test for any differences in the intercepts. Linear regressions were taken from the second GLM analyses for which the adjusted r 2 values are reported.
Male and female morphometric measurements were compared using independent-samples t tests (assuming equal variances), and GLMs were used to investigate differences in M b and RMR between seasons, between sexes and between interactions (season × sex). All statistical analyses were performed using SPSS Statistics (Version 19).
Results
Energetics
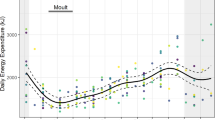
All birds were significantly heavier in summer than in winter, and males were significantly heavier than females regardless of season (See “Methods”, Table 1). Therefore, only mass-specific metabolic rates are reported here. Both male and female birds shared a maximum sustainable speed of 1.25 m s−1, which did not change with season. P met increased linearly with U (Fig. 1a). Neither season nor sex accounted for any variation in P met and the incremental increase in P met with U was not significantly different between seasons or sexes (Table 1). Accounting for RMR by calculating net-P met did not give rise to any seasonal or sex differences in mass-specific metabolic rate during locomotion (Fig. 1b; Table 1). CoTtot decreased curvilinearly with U across the speed range of the eiders (Fig. 1c), similar to that previously described for the barnacle goose (Nudds et al. 2010). Calculating CoTmin as the slope of the relationship between P met and U gives 7.23 J kg−1 m−1 (Fig. 1a), and using the slope of net-P met versus U gives 6.95 J kg−1 m−1 (Fig. 1b). The difference between these two slopes is the result of calculating net-P met using the mean RMR for each of S M, S F, W M and W F (9.75, 8.01, 9.11 and 8.22 W kg−1, respectively), which were not significantly different between groups (Table 1). The minimum measured values of CoTtot and CoTnet (19.32 ± 0.89 and 12.29 ± 0.91 J kg−1 m−1, respectively) were those at the maximum U of the birds (Fig. 1c, d). It should be noted, however, that a similar value is likely to be obtained across a range of their higher U. Under the assumption that the birds used only walking gait across their range of U, the allometric equation of Rubenson et al. (2007) (Walking CoTmin = 17.25 M −0.449b ) predicts a CoTmin of 12.78–13.77 J kg−1 m−1 for animals ranging 1.65–1.95 kg, which is higher than the values calculated using the slope method, but very close to the observed minimum CoTnet. If the birds were running at U where their cost of transport was minimised, allometry predicts a CoTmin of 10.23–10.85 J kg−1 m−1 for animals of this size, which falls between the values derived using the two different methods [Running CoTmin = 12.91 M −0.346b (Rubenson et al. 2007)].

Metabolic variables versus speed (U) for summer males (S M, closed triangles), summer females (S F, open triangles), winter males (W M, closed circles) and winter females (W F, open circles). a Mass-specific metabolic power consumption (P met) increased linearly with U in all groups (S M: P met = 16.02 + 7.23U; S F: P met = 15.43 + 7.23U; W M: P met = 15.15 + 7.23U; W F: P met = 14.56 + 7.23U). b Net-P met (P met during locomotion − P met whilst standing quietly) increased linearly with U in all groups (S M: net-P met = 6.61 + 6.95U; S F: net-P met = 7.36 + 6.95U; W M: net-P met = 6.39 + 6.95U; W F: net-P met = 7.14 + 6.95U). c Total mass-specific cost of transport (CoTtot) and d net mass-specific cost of transport (CoTnet) decreased curvilinearly with U. None of the relationships between energetic parameters and U were significantly different between groups (Table 1). At 1.25 m s−1, n = 2 and n = 1 for S F and W F, respectively
Mean exercising RER (±SE) for all speeds were 0.76 ± 0.03, 0.85 ± 0.03, 0.75 ± 0.04 and 0.79 ± 0.03 in S M, S F, W M and W F, respectively, in accordance with that previously reported for walking eiders (0.77 ± 0.12) (Hawkins et al. 2000). RER increased predictably with U in all groups with no significant difference in this relationship between groups (Table 1).
Kinematics
Minimum DF (0.57 ± 0.01) did not reach 0.5 indicating that eiders did not use aerial running gait (Fig. 2a). DF decreased linearly with U in all groups. The rate of decrease in DF (−0.20) was common between groups, but absolute DF was 0.02 higher in males, compared to females, across all U (Table 1). t stance (Fig. 2b) decreased curvilinearly with U in all groups and the intercepts of these relationships were higher in males, compared to females, and higher in summer, compared to winter (Table 1). The rate of decrease in t stance was identical between the sexes, but was greater in summer (−0.65) compared to winter (−0.58) (Table 1). Absolute values of t stance became more similar between seasons and sexes towards the higher range of U. t swing (Fig. 2b) decreased linearly with U in all groups with no significant differences in these relationships between groups (Table 1). In contrast, f stride (Fig. 2c) increased linearly in all groups with identical slopes (1.40) but was 0.07 Hz faster in females than in males across all U (Table 1). Similarly l stride (Fig. 2d) increased linearly with U with identical slopes (0.28) in all groups, but was 0.02 m longer in males than in females across all U (Table 1).

Kinematic variables versus speed (U) for summer males (S M, closed triangles, thick black line), summer females (S F, open triangles, dotted and dashed line), winter males (W M, closed circles, thick dashed line) and winter females (W F, open circles, dotted line). Best-fit lines represent significant differences between groups. Where sex was the only factor causing differences male and female best-fit lines are represented by solid lines and dashed lines, respectively, a duty factor (DF) decreased linearly with U in all groups (S M: DF = 0.82 − 0.20U; S F: DF = 0.80 − 0.20U; W M: DF = 0.82 − 0.20U; W F: DF = 0.80 − 0.20U) and was higher in males compared to females. b Stance phase duration (t stance) decreased curvilinearly with U in all groups (S M: t stance = 0.26U −0.65; S F: t stance = 0.24U −0.65; W M: t stance = 0.28U −0.58; W F: t stance = 0.25U −0.58), and the intercepts of these relationships were significantly higher in males compared to females and significantly higher in summer compared to winter. The rate of decrease in t stance was identical between the sexes but was greater in summer than winter. Swing phase duration (t swing) decreased linearly with U in all groups (S M: t swing = 0.18 − 0.02U; S F: t swing = 0.19 − 0.02U; W M: t swing = 0.18 − 0.02U; W F: t swing = 0.19 − 0.02U), and these relationships were not significantly different between groups. c Stride frequency (f stride) increased linearly with U (S M: f stride = 0.96 + 1.40U; S F: f stride = 1.03 + 1.40U; W M: f stride = 0.98 + 1.40U; W F: f stride = 1.06 + 1.40U) and was higher in females compared to males. d Stride length (l stride) increased linearly with U (S M: l stride = 0.15 + 0.28U; S F: l stride = 0.13 + 0.28U; W M: l stride = 0.14 + 0.28U; W F: l stride = 0.13 + 0.28U) and was longer in males compared to females
Morphometric measurements
External measurements of the tarsus lengths were not significantly different between male (67.6 ± 1.1 mm, n = 5) and female (63.0 ± 1.6 mm, n = 4) birds (independent-samples t test assuming unequal variances: t = 2.19, p = 0.078). Furthermore, there were no sex differences in the lengths of the femuri (male: 60.07 ± 1.16 mm, n = 5; female: 57.56 ± 1.33 mm, n = 4; t = 1.43, p = 0.401), tibiotarsi (male: 99.47 ± 1.64 mm, n = 5; female: 96.67 ± 1.65 mm, n = 4; t = 1.20, p = 0.947), tarsometatarsi (male: 51.11 ± 1.00 mm, n = 5; female: 48.76 ± 0.63 mm, n = 4; t = 1.98, p = 0.468) or total leg length (sum of three skeletal element lengths) (male: 210.64 ± 3.74 mm, n = 5; female: 202.99 ± 3.50 mm, n = 4; t = 1.49, p = 0.956) of the museum specimens. Neither were there any sex differences in pelvis width (male: 29.41 ± 0.70 mm, n = 5; female: 27.33 ± 0.45, n = 5; t = 2.49, p = 0.657).
Discussion
Eiders are incapable of an aerial running gait, in keeping with findings for another diving bird, the great cormorant (White et al. 2008). The slope of the relationship between f stride and U decreased by 52 % on transition to grounded-running in the great cormorant (White et al. 2008). However, in the present study, f stride increased linearly and continuously up to the maximum U of the birds with no inflections in gradient to suggest a transition to grounded-running. Furthermore, there was no evidence for a transition to grounded-running by energy savings (i.e. a step reduction in CoTtot). The maximum sustainable U of the eiders (1.25 m s−1) was 2.5-fold greater than that recorded for great-cormorants (0.5 m s−1), despite the cormorants from the study of White et al. (2008) being of similar M b (2.3 ± 0.1 kg) to our eiders. As maximum U is positively correlated to hindlimb length (Garland and Janis 1993), we expect that eiders have longer legs than cormorants, allowing the higher U (White et al. 2008). The kinematics of terrestrial locomotion in the common eider are in fact more comparable with those of the generalist barnacle goose (M b also ~ 2 kg) with which it shared identical maximum U, minimum DF and similar relationships between kinematic parameters and U (Nudds et al. 2010).
Maximum rate of oxygen consumption (\(\dot{V}_{{{\text{O}}_{2} \hbox{max} }}\)) was not reached by the ducks during trials, indicating their maximum U is likely limited by a mechanical constraint of their adaptive diving morphology or muscle physiology, rather than a limit on energy metabolism. In concurrence with findings for the great cormorant (White et al. 2008), eiders incur a CoTmin similar to cursorial birds of the same size. Why these divers have energetically efficient terrestrial locomotion while non-specialists (which also use aquatic locomotion), such as barnacle and greylag geese (Anser anser), have relatively energetically inefficient locomotion is unknown (Fedak et al. 1974; Nudds et al. 2010). However, the CoTmin of barnacle geese was compared to predictions from Taylor et al.’s (1982) allometric equation for terrestrial animals including both walking and running gaits, and these geese are incapable of running. When comparing the CoTmin of barnacle geese from Nudds et al.’s (2010) study to that predicted using Rubenson et al.’s (2007) allometric equation for CoTmin during walking gait, their CoTmin is almost equal to the prediction (M b = 1.5–2.1 kg; prediction = 12.36–14.38 J kg−1 m−1; slope = 12.26 J kg−1 m−1). It appears that we have a limited understanding of the constraints acting upon terrestrial locomotion in avian species adapted to different locomotor modes. A larger sample size of dabbling and diving species and consistency in the methodology for calculating and predicting CoTmin is required. Furthermore, attention to hindlimb tendon elastic energy savings (Biewener and Corning 2001) and the energy recovery from the phasing of the kinetic and potential energies of the CoM during waddling (Griffin and Kram 2000) could provide insight into the complexity of the constraints on both gait selection and energy metabolism in birds.
Despite the fact that daily energy expenditure is greater in summer (during feather synthesis) compared to winter in eiders (Guillemette and Butler 2012), and M b was greater in summer compared to winter, the birds in this study exhibited no seasonal differences in mass-specific CoL. Seasonal differences in the CoL have been investigated in the Svalbard rock ptarmigan, which was found to have a reduced mass-specific CoL in winter compared to summer (Lees et al. 2010). Ptarmigan are primarily cursorial, however, and selection would therefore be expected to act on their terrestrial locomotor economy in a changing environment. Given that eiders spend the majority of their lives at sea, selection is likely to favour adaptations towards aquatic locomotion rather than terrestrial locomotion. Unlike the ptarmigan which adjust M b dramatically in order to withstand unpredictable winters, eiders do not undergo pronounced changes in M b; relying on large surface area: volume ratios, the exceptional insulating ability of their feathers, huddling in dense colonies (Lovvorn et al. 2009) and foraging less (Heath et al. 2010) to conserve energy. Interestingly, unlike in the ptarmigan, RMR did not vary seasonally in the eiders, and calculating net-P met in order to account for any seasonal differences in non-locomotor energy expenditure associated with RMR did not change the outcome of our results.
Despite showing sex differences in plumage, M b and energy investment in reproduction and parental care, male and female eiders shared an identical mass-specific CoL and maximum sustainable U, which was expected given their identical tarsus and hindlimb skeletal element lengths. Interestingly, despite sharing identical limb morphometrics, their kinematic parameters differed subtly; with males having duty factors 0.02 greater, t stance 0.04 s longer, f stride 0.07 Hz slower and l stride 0.02 m longer across all U. It seems likely that this is linked to the greater M b of the males, which would be expected to require longer periods of foot–ground contact to generate the force required to support body weight. Sex differences in terrestrial locomotion have only been reported previously in cursorial Galliformes, with male domestic chickens and Svalbard rock ptarmigan achieving higher maximum sustainable U than females (Brackenbury and Elsayed 1985; Lees et al. 2011). Although male and female domestic chickens were shown to exhibit an identical mass-specific CoL, the male and female birds in this study were of different sized breeds (White Leghorn and Warren, respectively), making the study less than a true comparison of the sexes without controlling for breed (Brackenbury and Elsayed 1985). The male bias in M b in the Brackenbury and Elsayed (1985) study was 56 %; however, within any single domestic breed of chicken, the bias is usually near to 21 % (Remes and Szekely 2010). Male Svalbard rock ptarmigan exhibited a lower mass-specific CoL than females, which could not be accounted for by the shorter hindlimbs and faster f stride of the females (Lees et al. 2011). The difference in energy budget between ptarmigan sexes may be unique, however, as the extended photoperiod during the summer breeding season in Svalbard results in males defending territories for 24 h a day (Unander and Steen 1985). Significant metabolic investment has been recorded in males carrying out behaviours involved with attaining mates (Vehrencamp et al. 1989; Barske et al. 2011). Allocating such a large proportion of daily energy budget towards displaying or defending territories and exposure to the danger of predation may provide a way that females can assess male quality where paternal investment is little or none (i.e. locomotor performance in males may be sexually selected) (Husak and Fox 2008; Byers et al. 2010; Barske et al. 2011; Lees et al. 2011). Although sexual selection is important in the common eider [with a male bias in the sex ratio further making females the limited resource (Swennen et al. 1979)], male courtship displays occur in the aquatic and not the terrestrial environment; involving wing-flapping, head-turning and chin-lifting on the surface of the water (Johnsgard 1964). Therefore, our findings do not conflict with the hypothesis of Lees et al. (2011) that terrestrial locomotion should influence male reproductive success in order to drive a sex difference in the CoL.
It should be noted that sex differences in locomotor performance could arise from selection driving physiological constraints upon females. Male red jungle fowl (Gallus gallus) have larger aerobic scopes (maximum-resting oxygen consumption) than females (Chappell et al. 1996). This may be due to males being better physiologically adapted to locomotion, having larger hearts, lungs and muscles as a relative proportion of M b (Hammond et al. 2000). Female jungle fowl, however, are adapted to egg production, having relatively larger digestive systems and livers for energy acquisition (Hammond et al. 2000). Supporting and transporting relatively more organ mass with relatively less muscle potentially requires relatively more energy (Taylor et al. 1980; Tickle et al. 2010); however, no studies to date have linked male and female energetic and kinematic data with such morphological measurements. Sexual dimorphisms like these are associated with male–male (intra-specific) aggressive competition for females in some vertebrate species (Cullum 1998; Hammond et al. 2000; Bonnet et al. 2005). Since inter- (rather than intra-) specific sexual selection is important in the common eider, and since male and female eiders shared identical top attainable U, it may be that the sexes also share similar visceral and muscle proportions. However, it is not known which anatomical components make the male eiders heavier than females. Equally, it may be that sexual dimorphisms in anatomical proportions are more pronounced in more cursorial species under similar breeding and parental care strategies to the eiders. In fact, of avian species exhibiting lekking behaviour, male-biased size dimorphism is most prominent in species with ground displays (Hoglund 1989). Conversely, it has been suggested that female birds may be constrained mechanically by wider pelves (associated with reproduction), as found in humans (Smith et al. 2002; Cho et al. 2004; Lees et al. 2011). This theory is not challenged by our results as no sex differences in pelvic morphology or energetics were found. Furthermore, the reduction in bone fracture resistance by the provision of calcium for egg production has also been suggested as a potential limitation on female terrestrial locomotion (Whitehead 2004; Lees et al. 2011). It may be that female hindlimbs are weakened during, and just after the egg-laying period; however, this is likely to be of more importance to birds that spend more time locomoting terrestrially than the eiders.
In conclusion, the common eider shares a similar CoTmin with cursorial birds of a similar size. However, eiders are incapable of aerial running gait and neither season nor sex influences their mass-specific CoL. We suggest that selection is unlikely to act on their CoL given their principally aquatic lifestyle: although there is clear evidence for sexual selection by female choice in this species, any female choice on male locomotor performance would be expected to occur in the aquatic, and not the terrestrial environment. Sex differences in the CoL and maximum U may therefore be exclusive to birds for which terrestrial locomotion facilitates male reproductive success. Furthermore, locomotor specialisation appears to have a strong influence over seasonal and sex-specific adaptations for survival and reproduction in these birds.
References
Bamford OS, Maloiy GMO (1980) Energy metabolism and heart rate during treadmill exercise in the Marabou stork. J Appl Physiol 49:491–496
Barske J, Schlinger BA, Wikelski M, Fusani L (2011) Female choice for male motor skills. Proc R Soc B 278:3523–3528
Biewener AA (1989) Scaling body support in mammals—limb posture and muscle mechanics. Science 245:45–48
Biewener AA, Corning WR (2001) Dynamics of mallard (Anas platyrhynchos) gastrocnemius function during swimming versus terrestrial locomotion. J Exp Biol 204:1745–1756
Blix AS (2005) Arctic animals and their adaptations to life on the edge. Tapir Academic Press, Trondheim
Bonnet X, Ineich I, Shine R (2005) Terrestrial locomotion in sea snakes: the effects of sex and species on cliff-climbing ability in sea kraits (Serpentes, Elapidae, Laticauda). Biol J Linn Soc 85:433–441
Brackenbury JH, Avery P (1980) Energy consumption and ventilatory mechanisms in the exercising fowl. Comp Biochem Physiol A 66:439–445
Brackenbury JH, Elsayed MS (1985) Comparison of running energetics in male and female domestic fowl. J Exp Biol 117:349–355
Brody S (1945) Bioenergetics and growth, with special reference to the efficiency complex in domestic animals. Reinhold, New York
Byers J, Hebets E, Podos J (2010) Female mate choice based upon male motor performance. Anim Behav 79:771–778
Chappell MA, Zuk M, Johnsen TS (1996) Repeatability of aerobic performance in red junglefowl: effects of ontogeny and nematode infection. Funct Ecol 10:578–585
Cho SH, Park JM, Kwon OY (2004) Gender differences in three dimensional gait analysis data from 98 healthy Korean adults. Clin Biomech 19:145–152
Criscuolo F, Gabrielsen GW, Gendner JP, Le Maho Y (2002) Body mass regulation during incubation in female common eiders Somateria mollissima. J Avian Biol 33:83–88
Cullum AJ (1998) Sexual dimorphism in physiological performance of whiptail lizards (genus Cnemidophorus). Physiol Biochem Zool 71:541–552
Dunn PO, Whittingham LA, Pitcher TE (2001) Mating systems, sperm competition, and the evolution of sexual dimorphism in birds. Evolution 55:161–175
Dunning JB (2008) CRC handbook of avian body masses, 2nd edn. CRC Press, Boca Raton
Elliott KH, Le Vaillant M, Kato A, Speakman JR, Ropert-Coudert Y (2013) Accelerometry predicts daily energy expenditure in a bird with high activity levels. Biol Lett 9:20120919
Fedak MA, Pinshow B, Schmidt-Nielsen K (1974) Energy cost of bipedal running. Am J Physiol 227:1038–1044
Fedak MA, Rome L, Seeherman HJ (1981) One-step N2-dilution technique for calibrating open-circuit VO2 measuring systems. J Appl Physiol 51:772–776
Full RJ, Tu MS (1991) Mechanics of a rapid running insect: two-, four- and six-legged locomotion. J Exp Biol 156:215–231
Gabrielsen GW, Mehlum F, Karlsen HE, Andersen O, Parker H (1991) Energy cost during incubation and thermoregulation in the female common eider Somateria mollissima. Nor Polarinst Skr 195:51–62
Garland T, Janis CM (1993) Does metatarsal femur ratio predict maximal running speed in cursorial mammals. J Zool 229:133–151
Gatesy SM, Biewener AA (1991) Bipedal locomotion—effects of speed, size and limb posture in birds and humans. J Zool 224:127–147
Goldstein DL (1988) Estimates of daily energy expenditure in birds: the time-energy budget as an integrator of laboratory and field studies. Am Zool 28:829–844
Goldstein DL, Nagy KA (1985) Resource utilization by desert quail—time and energy, food and water. Ecology 66:378–387
Griffin TM, Kram R (2000) Biomechanics penguin waddling is not wasteful. Nature 408:929
Guillemette M, Butler PJ (2012) Seasonal variation in energy expenditure is not related to activity level or water temperature in a large diving bird. J Exp Biol 215:3161–3168. doi:10.1242/jeb.061119
Guillemette M, Pelletier D, Grandbois JM, Butler PJ (2007) Flightlessness and the energetic cost of wing molt in a large sea duck. Ecology 88:2936–2945
Guillemette M, Richman SE, Portugal SJ, Butler PJ (2012) Behavioural compensation reduces energy expenditure during migration hyperphagia in a large bird. Funct Ecol 26:876–883
Hammond KA, Chappell MA, Cardullo RA, Lin RS, Johnsen TS (2000) The mechanistic basis of aerobic performance variation in red junglefowl. J Exp Biol 203:2053–2064
Hancock JA, Stevens NJ, Biknevicius AR (2007) Whole-body mechanics and kinematics of terrestrial locomotion in the Elegant-crested Tinamou Eudromia elegans. Ibis 149:605–614
Hawkins PAJ, Butler PJ, Woakes AJ, Speakman JR (2000) Estimation of the rate of oxygen consumption of the common eider duck (Somateria mollissima), with some measurements of heart rate during voluntary dives. J Exp Biol 203:2819–2832
Heath JP, Gilchrist HG, Ydenberg RC (2006) Regulation of stroke pattern and swim speed across a range of current velocities: diving by common eiders wintering in polynyas in the Canadian Arctic. J Exp Biol 209:3974–3983
Heath JP, Gilchrist HG, Ydenberg RC (2010) Interactions between rate processes with different timescales explain counterintuitive foraging patterns of arctic wintering eiders. Proc R Soc B 277:3179–3186. doi:10.1098/rspb.2010.0812
Hoglund J (1989) Size and plumage dimorphism in lek-breeding birds—a comparative-analysis. Am Nat 134:72–87
Husak JF, Fox SF (2008) Sexual selection on locomotor performance. Evol Ecol Res 10:213–228
Johnsgard PA (1964) Comparative behaviour and relationships of the eiders. Condor 66:113–129
Kram R, Taylor CR (1990) Energetics of running—a new perspective. Nature 346:265–267
Langman VA, Rowe MF, Roberts TJ, Langman NV, Taylor CR (2012) Minimum cost of transport in Asian elephants: do we really need a bigger elephant? J Exp Biol 215:1509–1514
Lees J, Nudds R, Stokkan KA, Folkow L, Codd J (2010) Reduced metabolic cost of locomotion in Svalbard rock ptarmigan (Lagopus muta hyperborea) during winter. PLoS ONE 5:e15490
Lees JJ, Nudds RL, Folkow LP, Stokkan KA, Codd JR (2011) Understanding sex differences in the cost of terrestrial locomotion. Proc R Soc B. doi:10.1098/rspb.2011.1334
Lighton JRB (2008) Measuring metabolic rates: a manual for scientists. Oxford University Press, Oxford
Lislevand T, Figuerola J, Szekely T (2009) Evolution of sexual size dimorphism in grouse and allies (Aves: Phasianidae) in relation to mating competition, fecundity demands and resource division. J Evol Biol 22:1895–1905
Lovvorn JR, Grebmeier JM, Cooper LW, Bump JK, Richman SE (2009) Modeling marine protected areas for threatened eiders in a climatically changing Bering Sea. Ecol Appl 19:1596–1613
Nudds RL, Codd JR, Sellers WI (2009) Evidence for a mass dependent step-change in the scaling of efficiency in terrestrial locomotion. Plos One 4. doi:10.1371/journal.pone.0006927
Nudds RL, Gardiner JD, Tickle PG, Codd JR (2010) Energetics and kinematics of walking in the barnacle goose (Branta leucopsis). Comp Biochem Physiol A Mol Integr Physiol 156:318–324. doi:10.1016/j.cbpa.2010.01.023
Nudds RL, Folkow LP, Lees JJ, Tickle PG, Stokkan KA, Codd JR (2011) Evidence for energy savings from aerial running in the Svalbard rock ptarmigan (Lagopus muta hyperborea). Proc R Soc B 278:2654–2661
Parker H, Holm H (1990) Patterns of nutrient and energy expenditure in female common eiders nesting in the high Arctic. Auk 107:660–668
Piersma T, Lindstrom A, Drent RH, Tulp I, Jukema J, Morrison RIG, Reneerkens J, Schekkerman H, Visser GH (2003) High daily energy expenditure of incubating shorebirds on High Arctic tundra: a circumpolar study. Funct Ecol 17:356–362
Pinshow B, Fedak MA, Schmidt-Nielsen K (1977) Terrestrial locomotion in penguins: it costs more to waddle. Science 195:592–594
Portugal SJ, Guillemette M (2011) The use of body mass loss to estimate metabolic rate in birds. Comp Biochem Physiol A Mol Integr Physiol 158:329–336
Remes V, Szekely T (2010) Domestic chickens defy Rensch’s rule: sexual size dimorphism in chicken breeds. J Evol Biol 23:2754–2759
Rigou Y, Guillemette M (2010) Foraging effort and pre-laying strategy in breeding common eiders. Waterbirds 33:314–322
Roberts TJ, Kram R, Weyand PG, Taylor CR (1998) Energetics of bipedal running I. Metabolic cost of generating force. J Exp Biol 201:2745–2751
Rubenson J, Heliams DB, Lloyd DG, Fournier PA (2004) Gait selection in the ostrich: mechanical and metabolic characteristics of walking and running with and without an aerial phase. Proc R Soc B 271:1091–1099
Rubenson J, Heliams DB, Maloney SK, Withers PC, Lloyd DG, Fournier PA (2007) Reappraisal of the comparative cost of human locomotion using gait-specific allometric analyses. J Exp Biol 210:3513–3524
Smith LK, Lelas JL, Kerrigan DC (2002) Gender differences in pelvic motions and center of mass displacement during walking: stereotypes quantified. J Women Health Gend Based Med 11:453–458
Spurr E, Milne H (1976) Adaptive significance of autumn pair formation in the common eider Somateria mollissima (L.). Ornis Scand 7:85–89
Stokkan KA, Lindgard K, Reierth E (1995) Photoperiodic and ambient temperature control of the annual body mass cycle in Svalbard ptarmigan. J Comp Physiol B 165:359–365
Swennen C, Duiven P, Reyrink LA (1979) Notes on the sex ratio in the common eider Somateria mollissima (L.). Ardea 67:54–61
Taylor CR, Heglund NC, Mcmahon TA, Looney TR (1980) Energetic cost of generating muscular force during running—a comparison of large and small animals. J Exp Biol 86:9–18
Taylor CR, Heglund NC, Maloiy GMO (1982) Energetics and mechanics of terrestrial locomotion. 1. Metabolic energy-consumption as a function of speed and body size in birds and mammals. J Exp Biol 97:1–21
Tickle PG, Richardson MF, Codd JR (2010) Load carrying during locomotion in the barnacle goose (Branta leucopsis): the effect of load placement and size. Comp Biochem Physiol A Mol Integr Physiol 156:309–317. doi:10.1016/J.Cbpa.2010.01.022
Tolkamp BJ, Emmans GC, Yearsley J, Kyriazakis I (2002) Optimization of short-term animal behaviour and the currency of time. Anim Behav 64:945–953. doi:10.1006/Anbe.2002.2008
Unander S, Steen JB (1985) Behavior and social structure in Svalbard rock ptarmigan Lagopus mutus hyperboreus. Ornis Scand 16:198–204
Usherwood JR, Szymanek KL, Daley MA (2008) Compass gait mechanics account for top walking speeds in ducks and humans. J Exp Biol 211:3744–3749. doi:10.1242/jeb.023416
Vehrencamp SL, Bradbury JW, Gibson RM (1989) The energetic cost of display in male sage grouse. Anim Behav 38:885–896
Watson RR, Rubenson J, Coder L, Hoyt DF, Propert MW, Marsh RL (2011) Gait-specific energetics contributes to economical walking and running in emus and ostriches. Proc R Soc B 278:2040–2046. doi:10.1098/rspb.2010.2022
Weathers WW, Sullivan KA (1993) Seasonal patterns of time and energy allocation by birds. Physiol Zool 66:511–536
White CR, Martin GR, Butler PJ (2008) Pedestrian locomotion energetics and gait characteristics of a diving bird, the great cormorant, Phalacrocorax carbo. J Comp Physiol B 178:745–754. doi:10.1007/S00360-008-0265-9
Whitehead CC (2004) Overview of bone biology in the egg-laying hen. Poult Sci 83:193–199
Acknowledgments
We would like to thank Magnus Folkow, Hans Lian, Hans-Arne Solvang and John Ness for technical assistance and animal husbandry. Thanks are also due to Anne-Marit Vik for assistance with data collection, to Robert Nudds for advice on statistical analyses and to Bob McGowan and Zena Floody from the National Museum of Scotland for their assistance with locating skeletal specimens. This research was funded by the Biotechnology and Biological Sciences Research Council (BBSRC) (G01138/1) Grant to JC. KR and JL were supported by a National Environmental Research Council (NERC) PhD doctoral training awards and KR by a CASE partnership with the Manchester Museum (University of Manchester).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Rose, K.A., Tickle, P.G., Lees, J.J. et al. Neither season nor sex affects the cost of terrestrial locomotion in a circumpolar diving duck: the common eider (Somateria mollissima). Polar Biol 37, 879–889 (2014). https://doi.org/10.1007/s00300-014-1488-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-014-1488-8