
Abstract
While constrained by endogenous rhythms, morphology and ecology, animals may still exhibit flexible activity patterns in response to risk. Temporal avoidance of interspecific aggression can enable access to resources without spatial exclusion. Apex predators, including humans, can affect mesopredator activity patterns. Human context might also modify temporal interactions between predators. We explored activity patterns, nocturnality and the effects of human activity upon a guild of carnivores (grey wolf, Canis lupus; Eurasian lynx, Lynx lynx; red fox, Vulpes vulpes) using travel routes in Plitvice Lakes National Park, Croatia. Humans were diurnal, foxes nocturnal and large carnivores active during the night, immediately after sunrise and before sunset. Carnivore activity patterns overlapped greatly and to a similar extent for all pairings. Activity curves followed expectations based on interspecific killing, with activity peaks coinciding where body size differences were small (wolf and lynx) but not when they were intermediate (foxes to large carnivores). Carnivore activity, particularly fox, overlapped much less with that of diurnal humans. Foxes responded to higher large carnivore activity by being more nocturnal. Low light levels likely provide safer conditions by reducing the visual detectability of mesopredators. The nocturnal effect of large carnivores was however moderated and reduced by human activity. This could perhaps be due to temporal shielding or interference with risk cues. Subtle temporal avoidance and nocturnality may enable mesopredators to cope with interspecific aggression at shared spatial resources. Higher human activity moderated the effects of top-down temporal suppression which could consequently affect the trophic interactions of mesopredators.
Significance statement
Temporal partitioning can provide an important mechanism for spatial resource access and species coexistence. Our findings show that carnivores partition the use of shared travel routes in time, using the cover of darkness to travel safely where their suppressors (large carnivores) are more active. We however observed fox nocturnality to be flexible, with responses depending on the activity levels but also the composition of apex predators. High human activity modified the top-down temporal suppression of mesopredators by large carnivores. The use of time by predators can have demographic and trophic consequences. Prey accessibility and susceptibility can be temporally variable. As such, the ecosystem services and the ecological roles of predators may be affected by human time use as well as that of intraguild competitors. Temporal interactions should not be overlooked when evaluating human use and conservation priorities in protected areas.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Top-down regulation and the suppression of mesopredators by large carnivores can be elicited via direct killing, harassment and the risk associated with such encounters (Crooks and Soulé 1999; Palomares and Caro 1999; Ritchie and Johnson 2009). Ecological differentiation along a niche axis is deemed necessary for coexistence between competitors (Hardin 1960). Carnivores may spatially avoid competitors or differentiate dietary niche (Azevedo et al. 2006; Bassi et al. 2012; Newsome and Ripple 2014). Aggressive encounters between species are however not solely dependent upon niche overlap but can also be affected by body size differences, resource availability, physical characteristics, behavioural strategies and similarity in stimuli (appearance, behaviour, scent etc.) that trigger agonistic behaviour due to common ancestry (Donadio and Buskirk 2006; Grether et al. 2013; Haswell et al. 2018). Regardless of diet, intraguild aggression can still present risk for mesopredators and some spatial resources such as linear travel routes may exacerbate this risk because of their frequent use by apex predators (Shannon et al. 2014; Haswell et al. 2018). Accordingly, an animal’s circadian activity pattern may provide an important dimension for minimising the likelihood of aggressive encounters.
Circadian clocks help maintain optimal activity and likely provide restrictions to activity patterns because divergence from endogenous rhythms can carry ecological and physiological costs (Kronfeld-Schor and Dayan 2003; Relógio et al. 2011). Intrinsic characteristics, such as eye morphology and visual acuity, may also restrict temporal niche (Veilleux and Kirk 2014; Banks et al. 2015). Animal behaviour and decision-making are however contextual (Haswell et al. 2017; Owen et al. 2017). Most mammals (excepting anthropoid primates) retain a scotopic (low-light) eye design consistent with nocturnal origins, yet mammals exhibit diurnal, cathemeral and nocturnal activity patterns (Heesy and Hall 2010; Hall et al. 2012; Borges et al. 2018). Carnivore activity patterns may be particularly affected by prey accessibility or susceptibility (Cozzi et al. 2012; Heurich et al. 2014). Time use can be influenced by abiotic conditions, resource acquisition and foraging success, but may also be affected by competition and risk (Reimchen 1998; Hayward and Slotow 2009; Theuerkauf 2009).
Temporal partitioning of activity may be a mechanism allowing mesopredators to avoid costly interspecific interactions (Monterroso et al. 2014; Diaz-Ruiz et al. 2016). Complete spatial avoidance of suitable habitat prevents access to resources, whereas temporal avoidance of competitors or aggressors can enable coexistence (Holt and Polis 1997; Swanson et al. 2016). Recent methodological advancements now permit the more detailed study of fine-scale activity patterns and temporal interactions between species (Ridout and Linkie 2009; Frey et al. 2017; Gaynor et al. 2018).
Humans can act as super predators, exerting top-down pressure on carnivores (Darimont et al. 2015; Smith et al. 2017). Human disturbance can make predators more nocturnal (Gaynor et al. 2018). The presence of humans however also makes a predator guild more complex. Human modification of risk landscapes can consequently affect interactions between carnivores (Haswell et al. 2017). An important question is how, or if, the human context modifies temporal interactions within predator guilds. Humans may affect the activity patterns of mesopredators directly, but also in a cascading manor, via temporal effects on large carnivores or interference with the mechanisms by which they affect mesopredator behaviour.
The forest roads and trails of Plitvice Lakes National Park, Croatia, provided an ideal opportunity to study temporal interactions. The park is home to large carnivores and mesopredators but is also used by humans in non-consumptive capacities. We hypothesised that the activity patterns of a mesopredator (red fox, Vulpes vulpes) would not coincide with that of sympatric apex carnivores (wolves, Canis lupus, and lynx, Lynx lynx). We also hypothesised that foxes might show spatial flexibility in their propensity for nocturnal activity depending upon the level of activity exhibited by large carnivores or humans at a given locality. We hypothesised that human trail use would present temporal restrictions to all carnivores and would interfere with intraguild interactions between large carnivores and foxes.
Methods
Study site
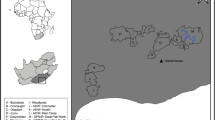
Plitvice Lakes National Park (Plitvice) is situated between 44° 44′ 34″ and 44° 57′ 48″ N and 15° 27′ 32″ and 15° 42′ 23″ E, in the Dinaric Mountains, Croatia (Šikić 2007) (Fig. 1). The mountainous karst (limestone and dolomite) landscape ranges from 367 to 1279 m above sea level and, excepting the iconic lakes and waterfalls, is characterised by scarce surface water (~ 1%), underground drainage systems, sink holes and caves (Šikić 2007; Romanić et al. 2016). Annual precipitation is 1550 mm with temperatures fluctuating between winter lows of − 3 °C and summer highs of 36 °C (Šikić 2007). One camera station contained planted stands of Scots and black pine (Pinus sylvestris and Pinus nigra), but elsewhere forest cover was predominantly Dinaric beech and fir trees (Fagus sylvatica and Abies alba). Tourism and recreation are permissible within the 297-km2 park where approximately 1770 people live within 19 settlements (Firšt et al. 2005; Romanić et al. 2016). The number of people visiting Plitvice has grown from 928,000 visitors in 2007 to over 1.72 million in 2017 (Smith 2018).

Map of study location, Plitvice Lakes National Park, Croatia. Black triangles detail the camera stations (October 2015–October 2016). Solid pale grey areas detail the boundaries of the national park. Roads are represented by solid grey lines, country boundaries by black lines and water bodies by dark grey areas
Data collection
We utilised records from 20 passive infrared motion sensor cameras placed on unpaved forest roads and trails in Plitvice between October 2015 and October 2016 (Fig. 1). Behavioural data collection was blind as activity record times were labelled by camera traps. Similarly to Santulli et al. (2014), we utilised data that was initially collected for other purposes. Camera station placement was ad hoc, with locations targeted according to large carnivore field signs (scats, tracks and markings). The national park’s desire to capture images of large carnivores may mean areas of lower large carnivore use are underrepresented in the dataset, but we do not believe this to be problematic for the questions being posed. Camera locations and periods included in the analysis were selected a priori to data examination. Fox density in Croatia is estimated at 0.7 per km2 with a territory size of 1.43 km2 (Slavica et al. 2010; Galov et al. 2014). Like Robinson et al. (2014), we assumed a circular territory size and utilised the radius (675 m) as the minimum acceptable distance between camera stations. Occasionally, an extra camera was deployed to capture both sides of a lynx spot pattern at promising stations. When two cameras were present at the same time (N = 3), we only used data from a single camera selected at random. Camera stations received almost year round coverage (range, 320–366 days). Considering the year as three 122-day periods based on fox reproductive behaviour (dispersal, October–January 30; denning, January 31–May; and weaning, June–September; see Haswell (2019)), each station received at least 89 observation days during any seasonal period.
Cameras were placed between 1 and 1.5 m high on trees or rocks adjacent to unbaited trails. A mixture of Acorn 5210A covert infrared, Uovision UV565HD, Uovision UM565, Reconyx HC500 Hyperfire and Bolyguard MG882K-8M cameras were utilised as logistics permitted. Cameras were checked monthly in summer but only at the start and end of winter due to accessibility restrictions. Data were collated in Camera Base 1.7 (www.atrium-biodiversity.org/tools/camerabase).
Data analysis
Like Rowcliffe et al. (2014), we defined activity records as the times of day that cameras were triggered by a given species. Only independent triggers (> 30 min apart) were utilised (Ridout and Linkie 2009; Linkie and Ridout 2011; Torretta et al. 2016). Individual animals could not be recognised, leading to some pseudoreplication.
Interspecific time use
In longer term studies of behavioural timings, it is important to ensure that actual timings, as given by the position of the sun, are used instead of clock time to prevent the generation of false activity patterns (Nouvellet et al. 2012). Clock time does not have any biological or environmental meaning, whereas the sun’s position in the sky does (Nouvellet et al. 2012). We adjusted the clock time of activity records to sun time using the “overlap” package in R version 3.5.1 (Meredith and Ridout 2018b). Activity patterns were then estimated as probability density functions using kernel density estimation (Ridout and Linkie 2009; Linkie and Ridout 2011; Meredith and Ridout 2018a).
We explored overlap in species activity patterns non-parametrically. Under the presumption that animals were equally likely to be photographed at any time they were active on trails, we fitted kernel density curves and estimated the coefficient of overlapping, Δ, which is the area lying under both curves (Ridout and Linkie 2009; Linkie and Ridout 2011; Meredith and Ridout 2018a). The coefficient of overlapping ranges from 0, indicating no overlap, to 1, indicating complete overlap (Ridout and Linkie 2009; Linkie and Ridout 2011). Sample sizes for each species were > 75 so, as recommended when estimating overlap, we used the non-parametric estimator \( \hat{\Delta} \)4 (Meredith and Ridout 2018a). We note that human data reflects pooled observations of motorised and non-motorised activity.
Within the “overlap” package in R, we generated 10,000 smoothed bootstrap samples to estimate a mean coefficient of overlap and 95% confidence intervals for each species pairing (Meredith and Ridout 2018a, b). The 2.5% and 97.5% percentiles of the bootstrap samples were adjusted to account for bootstrap bias (approach “basic0”) (Meredith and Ridout 2018a). We performed interval corrections on a logistic scale and back-transformed them to correct for any confidence interval estimates falling outside the possible range of 0–1 (Meredith and Ridout 2018a).
Nocturnality
We created a dichotomous dependent variable, labelling daytime activity records (between sunrise and sunset) as zero and night time records (before sunrise and after sunset) as one, using sunrise and sunset times from the United States Naval Observatory (http://aa.usno.navy.mil/data/index.php). We calculated photo capture rate indices (PCRI) for humans at each station, with PCRI being the number of independent (> 30 min apart) photo captures per 100 days (Rayan and Linkie 2016). Additionally, we calculated the PCRI for both large carnivores combined. Using generalized linear models (binomial distribution and logit link function), we examined if human PCRI affected whether each carnivore’s activity records occurred at night. The events variable was the number of nocturnal records and the trials variable was the total number of records for each camera station. In the fox model, we also examined the effect of large carnivore PCRI and the interaction between large carnivore PCRI and human PCRI. Null (intercept only) mixed models suggested no significant random effect of camera station for any species, so we did not develop multi-level models. Robust standard error estimation was however used to provide more conservative tests of model parameter significance, taking potential clustering effects into consideration.
Overlap between species activity patterns was conducted in R version 3.5.1 (R Core Development Team 2008). Generalized linear models examining nocturnality were conducted in IBM SPSS statistics 25 (IBM Corp 2017).
Results
During 6833 camera trapping days, 1197 activity records were obtained for fox, 80 for wolves, 156 for lynx and 3715 for humans. Foxes had the highest proportion of records occurring at night (88%), with wolves and lynx each having 71%. Humans were highly diurnal with only 4% of their records occurring at night. Foxes and humans were observed at all camera stations, wolves at 15 (75% of stations) and lynx at 16 (80%) stations.
Interspecific time use
Confidence intervals suggested all pairs of carnivores showed similar activity pattern overlap. Mean overlap of foxes with wolves was \( \hat{\Delta} \)4 0.73 (95% CI, 0.65 to 0.82) and with lynx was \( \hat{\Delta} \)4 0.75 (0.65 to 0.79). Wolf and lynx overlap was \( \hat{\Delta} \)4 0.79 (0.72 to 0.89). Carnivore activity overlap with humans was lower than with other carnivores. Fox overlap with human activity curves was the lowest, \( \hat{\Delta} \)4 0.17 (0.14 to 0.17). Wolves and lynx had similar overlap with human activity curves. Wolf and human overlap was \( \hat{\Delta} \)4 0.28 (0.19 to 0.34). Lynx overlap with humans was also \( \hat{\Delta} \)4 0.28 (0.22 to 0.32).
Foxes were very inactive during daylight hours with activity peaking after sunset and declining across the night, reaching low levels shortly after sunrise (Fig. 2a, b, d). The peak of fox activity did not coincide with activity peaks of larger species; however, all carnivores were highly active during the night (Fig. 2a, b). Lynx activity peaked in the later part of the night between midnight and sunrise but this period of higher activity was briefer than in wolves, with lynx utilising the early parts of the day at levels intermediate between foxes (Fig. 2b) and wolves (Fig. 2c). Lynx had a second period of higher activity around sunset—using the late afternoon more than foxes (Fig. 2b) and wolves (Fig. 2c). Wolves were more active than foxes (Fig. 2a) and lynx (Fig. 2c) during the early hours of the day, with activity lowest after noon, rising after sunset and peaking similar to lynx in the later part of the night (Fig. 2c). Humans dominated the daylight hours with activity peaking just before noon, which contrasted strongly to nocturnal carnivores (Fig. 2d–f).

Temporal niche overlap (grey area) between carnivores and humans in Plitvice Lakes National Park, October 2015–October 2016. Dotted lines represent kernel density estimates for red foxes, Vulpes vulpes; dot-dash lines for grey wolves, Canis lupus; dashed lines for Eurasian lynx, Lynx lynx; and solid lines for humans, Homo sapiens. Kernel density estimates are plotted as a function of sunrise time
Nocturnality
Generalized linear modelling revealed that large carnivore trail use (PCRI) had a significant effect on whether fox activity records occurred at night (Wald χ2 = 9.68, df = 1, P = 0.002). Increases in large carnivore PCRI increased the log odds that fox activity would be nocturnal (β = 0.142; 95% CI, 0.053 to 0.232). The effect of large carnivores was however moderated by this covariates’ interaction with human trail use (Wald χ2 = 5.03, df = 1, P = 0.025). Unit increases in human PCRI reduced the nocturnal effect large carnivores had upon foxes (β = − 0.002, − 0.003 to − 0.0002). Human PCRI had no direct effect on fox nocturnality (Wald χ2 = 2.19, df = 1, P = 0.139). The fox model had utility in predicting whether fox activity records occurred at night, providing a significant improvement in fit over the null model (likelihood ratio χ2 = 15.09, df = 3, P = 0.002). Human PCRI did not have a significant effect on whether lynx (Wald χ2 = 1.80, df = 1, P = 0.179) or wolf records were nocturnal (Wald χ2 = 2.51, df = 1, P = 0.113).
Discussion
We observed temporal partitioning among carnivores and humans on trails within Plitvice. Fox nocturnality was also contextual, being dependant on the intensity of human and large carnivore activity. Our findings support the notion of a level of flexibility in activity patterns, with animals avoiding activity during high-risk periods (Lima and Bednekoff 1999; Kronfeld-Schor and Dayan 2003). Higher large carnivore activity made foxes more nocturnal. The extent to which mesopredators utilise nocturnal safety may however depend on the composition and activity level of local predator communities. Humans reduced the nocturnal effect large carnivores had on foxes in Plitvice, supporting the notion that humans can dampen the top-down ecological effects of large carnivores (Hebblewhite et al. 2005).
Subordinate mesopredators may need to move their activity around the foraging bouts of larger carnivores (Hayward and Slotow 2009). In response to their nocturnal intraguild competitors, American mink, Neovison vison, have been observed to become diurnal; we did not however observe this in foxes (Harrington et al. 2009). In Plitvice, fox activity was predominantly nocturnal and overlapped highly, although not completely, with that of large carnivores. Activity curves show large carnivores made more use of parts of the day when humans were less active, seemingly restricting daylight activity by foxes. Activity peaks coincided in time where body size differences were small (wolf and lynx) but not when they were intermediate (foxes to large carnivores). This follows the patterns of interspecific killing associated with body size differences (Donadio and Buskirk 2006), but not interspecific competition avoidance, which would be greater between similarly sized species (Schoener 1974a, b). Confidence intervals however suggested no difference in activity overlap between any carnivore pairing. Predators that evolved under similar ecological conditions and share ecological traits may have similar activity patterns and co-occur often, limiting the potential for substantial temporal avoidance (Kronfeld-Schor and Dayan 2003; Davis et al. 2018).
Foxes were more nocturnal where large carnivore activity was higher. Low light levels and visual obstacles can increase spatial tolerance and reduce agonistic interactions between intraspecific competitors (Reimchen 1998). Presumably the same might be true of interspecific competitors. Animals may be less conspicuous in lower light levels; predation and harassment risk might therefore be lower at night, providing a time period where habitat and resources can be accessed more safely (Beauchamp 2007). Foxes avoided the risk of daytime trail encounters where large carnivores were more active but made greater use of a broader temporal niche in less risky contexts. Mesopredators can expand their niche axes in the absence of top-down pressure (Gese and Grothe 1995; Prugh et al. 2009; Kamler et al. 2013). Monterroso et al. (2013) observed foxes to be the dominant daytime user in a Mediterranean national park devoid of larger carnivores (Cabañeros, Spain). Our findings suggest that foxes can readily adjust their activity patterns as required in response to localised variation in top-down pressure. Risk perception may play a pivotal role in informing such flexible behaviour (Leo et al. 2015; Haswell et al. 2018; Kohl et al. 2018).
Behavioural responses to risk can result in significant demographic consequences (Preisser et al. 2005; Creel and Christianson 2008). Demographic consequences may however be negligible when avoidance is on a very fine, moment-to-moment scale (Swanson et al. 2014, 2016). Avoiding interspecific aggression along the temporal niche axis could however carry costs to individual body condition (Harrington et al. 2009). Temporal obstruction by larger carnivores could also inhibit mesopredator foraging, having indirect trophic consequences by offering respite to certain prey species. For example, when diurnal prey (Orthoptera) provides an important dietary component for red foxes, this can drive increased diurnal activity (Cavallini and Lovari 1991). Small mammal prey may also be capable of altering their own activity patterns, becoming more diurnal to avoid encounters with red foxes (Fenn and Macdonald 1995). Foxes, like other mesopredators, fulfil key trophic functions (Smedshaug et al. 1999; Roemer et al. 2009). Whether temporal avoidance of large carnivores occurs broadly across the landscape or at other microhabitat sites, resulting in demographic and trophic consequences, requires further investigation. The scope of our study was also limited to 1 year; temporal variation in factors such as mesopredator food availability might too result in alternative dynamics, and the consistency of interactions across years requires attention.
Humans can provide additional predation risk and function as super predators (Walther 1969; Smith et al. 2017). The general activity patterns we observed on trails in Plitvice suggest humans functioned as dominant super predators with regard to the temporal niche. Humans were highly active during the day, and carnivore activity, particularly that of foxes, overlapped much less with humans than other carnivores. Nocturnality in carnivores could suggest avoidance, particularly when humans present high risk (Kusak et al. 2005; Diaz-Ruiz et al. 2016). Limited temporal overlap might however be expected given species adaptations and evolutionary history (Heesy and Hall 2010; Hall et al. 2012). A lack of carnivore activity during the central parts of the day could reflect avoidance of heat, but human activity during twilight can still affect carnivore hunting success (Hayward and Slotow 2009; Theuerkauf 2009).
Unlike Gaynor et al. (2018), we did not find evidence to support increased mammal nocturnality in response to higher human activity. Given that carnivores were already highly nocturnal in Plitvice, we might not have detected variation in response to human activity, but it also might not have existed. We did however find that human activity moderated top-down effects in Plitvice. The nocturnal effect large carnivores had on foxes was dampened by human activity. Benitez-Lopez (2018) suggest that the human disturbance of apex predators from daylight activity might affect ecological interactions. An undetected effect of humans on large carnivore daytime activity, with humans shielding foxes from large carnivore daytime use, might explain our observations. That said, human activity may have lessened the effect of large carnivores on foxes via an alternative mechanism. High human activity might disrupt scent pictures and make the detection of risk cues from large carnivores more difficult, resulting in foxes modifying their behaviour less even though large carnivores were more active at a given station. The exact mechanism remains unknown but we can conclude that humans disrupted ecological interactions in Plitvice.
Interference with predator to predator interactions and consequent changes to mesopredator foraging activity could alter the pressure these efficient predators place upon prey communities (Vance-Chalcraft et al. 2007; Ritchie and Johnson 2009). Such interactions are however unlikely to be solely modern phenomena. Hominins have potentially been a part of European predator communities for 1.2 million years, with modern humans present at least 43,000 years (Carbonell et al. 2008; Benazzi et al. 2011). The spatial extent and numbers of humans in modern Europe is now, of course, dramatically more substantial. A key issue for protected areas thus lies in understanding the tipping points at which human activity becomes detrimental to biodiversity, ecosystem function and conservation efforts.
Observing changes in behaviour, such as activity patterns, can improve our understanding of ecological processes but can also provide early warning signals; e.g. temporal avoidance of humans might be a precursor to spatial exclusion, population decline or regional extinction following growing anthropogenic pressure (Berger-Tal et al. 2011; Caravaggi et al. 2017). Intense human pressure is prevalent in almost a third of global protected lands and undermines biodiversity preservation (Jones et al. 2018). Furthermore, interference risks altering baselines, not only negating the function of reserves in detecting ecological change, but also distorting public understanding of intact ecological processes (Sarmento and Berger 2017). Increasing intensity, temporal or spatial coverage of human activities beyond species tolerance could also conflict with conservation goals (Firšt et al. 2005; Štrbenac et al. 2005). Human activities can negatively affect foraging success, territorial defence, mate acquisition and reproductive output, as well as causing spatial displacement, stress and reduced energy intake, which have the potential to ultimately affect body condition, survival, fitness and demography (Frid and Dill 2002; Strasser and Heath 2013; Pauli et al. 2017). Given the lack of true wilderness areas in Europe however, many believe the most probable scenario of saving wildlife will require the dynamic interspersion of both wildlife and humans (Chapron et al. 2014).
Our findings show that mesopredators apply temporal strategy to enable the use of shared travel routes. How they use trails is affected by the level of use of other predators, as well as the interactions between multiple trail users. The effect of large carnivore activity on mesopredator trail use and the resultant trophic consequences may be dependent on the activity of humans. The significance and costs of such interference to conservation goals requires further exploration. Given temporal displacement may serve as an early warning sign to further ecological degradation, we urge parks to carefully consider the spatial and temporal extent of recreation and to monitor its impacts.
Data availability
The datasets generated and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Azevedo FCC, Lester V, Gorsuch W, Lariviere S, Wirsing AJ, Murray DL (2006) Dietary breadth and overlap among five sympatric prairie carnivores. J Zool 269:127–135
Banks MS, Sprague WW, Schmoll J, Parnell JAQ, Love GD (2015) Why do animal eyes have pupils of different shapes? Sci Adv 1:e1500391
Bassi E, Donaggio E, Marcon A, Scandura M, Apollonio M (2012) Trophic niche overlap and wild ungulate consumption by red fox and wolf in a mountain area in Italy. Mamm Biol 77:369–376
Beauchamp G (2007) Exploring the role of vision in social foraging: what happens to group size, vigilance, spacing, aggression and habitat use in birds and mammals that forage at night? Biol Rev 82:511–525
Benazzi S, Douka K, Fornai C et al (2011) Early dispersal of modern humans in Europe and implications for Neanderthal behaviour. Nature 479:525–529
Benitez-Lopez A (2018) Animals feel safer from humans in the dark. Science 360:1185–1186
Berger-Tal O, Polak T, Oron A, Lubin Y, Kotler BP, Saltz D (2011) Integrating animal behavior and conservation biology: a conceptual framework. Behav Ecol 22:236–239
Borges R, Johnson WE, O'Brien SJ, Gomes C, Heesy CP, Antunes A (2018) Adaptive genomic evolution of opsins reveals that early mammals flourished in nocturnal environments. BMC Genomics 19:121
Caravaggi A, Banks PB, Burton AC, Finlay CMV, Haswell PM, Hayward MW, Rowcliffe MJ, Wood MD (2017) A review of camera trapping for conservation behaviour research. Remote Sens Ecol Conserv 3:109–122
Carbonell E, de Castro JMB, Pares JM et al (2008) The first hominin of Europe. Nature 452:465–470
Cavallini P, Lovari S (1991) Environmental factors influencing the use of habitat in the red fox, Vulpes vulpes. J Zool 223:323–339
Chapron G, Kaczensky P, Linnell JD, von Arx M, Huber D, Andrén H, López-Bao JV, Adamec M, Álvares F, Anders O, Balčiauskas L, Balys V, Bedő P, Bego F, Blanco JC, Breitenmoser U, Brøseth H, Bufka L, Bunikyte R, Ciucci P, Dutsov A, Engleder T, Fuxjäger C, Groff C, Holmala K, Hoxha B, Iliopoulos Y, Ionescu O, Jeremić J, Jerina K, Kluth G, Knauer F, Kojola I, Kos I, Krofel M, Kubala J, Kunovac S, Kusak J, Kutal M, Liberg O, Majić A, Männil P, Manz R, Marboutin E, Marucco F, Melovski D, Mersini K, Mertzanis Y, Mysłajek RW, Nowak S, Odden J, Ozolins J, Palomero G, Paunović M, Persson J, Potočnik H, Quenette PY, Rauer G, Reinhardt I, Rigg R, Ryser A, Salvatori V, Skrbinšek T, Stojanov A, Swenson JE, Szemethy L, Trajçe A, Tsingarska-Sedefcheva E, Váňa M, Veeroja R, Wabakken P, Wölfl M, Wölfl S, Zimmermann F, Zlatanova D, Boitani L (2014) Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 346:1517–1519
Cozzi G, Broekhuis F, McNutt JW, Turnbull LA, Macdonald DW, Schmid B (2012) Fear of the dark or dinner by moonlight? Reduced temporal partitioning among Africa’s large carnivores. Ecology 93:2590–2599
Creel S, Christianson D (2008) Relationships between direct predation and risk effects. Trends Ecol Evol 23:194–201
Crooks KR, Soulé ME (1999) Mesopredator release and avifaunal extinctions in a fragmented system. Nature 400:563–566
Darimont CT, Fox CH, Bryan HM, Reimchen TE (2015) The unique ecology of human predators. Science 349:858–860
Davis CL, Rich LN, Farris ZJ, Kelly MJ, di Bitetti MS, Blanco YD, Albanesi S, Farhadinia MS, Gholikhani N, Hamel S, Harmsen BJ, Wultsch C, Kane MD, Martins Q, Murphy AJ, Steenweg R, Sunarto S, Taktehrani A, Thapa K, Tucker JM, Whittington J, Widodo FA, Yoccoz NG, Miller DAW (2018) Ecological correlates of the spatial co-occurrence of sympatric mammalian carnivores worldwide. Ecol Lett 21:1401–1412
Diaz-Ruiz F, Caro J, Delibes-Mateos M, Arroyo B, Ferreras P (2016) Drivers of red fox (Vulpes vulpes) daily activity: prey availability, human disturbance or habitat structure? J Zool 298:128–138
Donadio E, Buskirk SW (2006) Diet, morphology, and interspecific killing in carnivora. Am Nat 167:524–536
Fenn MGP, Macdonald DW (1995) Use of middens by red foxes - risk reverses rhythms of rats. J Mammal 76:130–136
Firšt B, Frković A, Gomerčić T et al (2005) Lynx management plan for Croatia. State Institute for Nature Protection, Zagreb
Frey S, Fisher JT, Burton AC, Volpe JP (2017) Investigating animal activity patterns and temporal niche partitioning using camera-trap data: challenges and opportunities. Remote Sens Ecol Conserv 3:123–132
Frid A, Dill L (2002) Human-caused disturbance stimuli as a form of predation risk. Conserv Ecol 6:11
Galov A, Sindičić M, Andreanszky T, Čurković S, Dežđek D, Slavica A, Hartl GB, Krueger B (2014) High genetic diversity and low population structure in red foxes (Vulpes vulpes) from Croatia. Mamm Biol 79:77–80
Gaynor KM, Hojnowski CE, Carter NH, Brashares JS (2018) The influence of human disturbance on wildlife nocturnality. Science 360:1232–1235
Gese EM, Grothe S (1995) Analysis of coyote predation on deer and elk during winter in Yellowstone National Park, Wyoming. Am Midl Nat 133:36–43
Grether GF, Anderson CN, Drury JP, Kirschel ANG, Losin N, Okamoto K, Peiman KS (2013) The evolutionary consequences of interspecific aggression. Ann N Y Acad Sci 1289:48–68
Hall MI, Kamilar JM, Kirk EC (2012) Eye shape and the nocturnal bottleneck of mammals. Proc R Soc Lond B 279:4962–4968
Hardin G (1960) The competitive exclusion principle. Science 131:1292–1297
Harrington LA, Harrington AL, Yamaguchi N, Thom MD, Ferreras P, Windham TR, Macdonald DW (2009) The impact of native competitors on an alien invasive: temporal niche shifts to avoid interspecific aggression? Ecology 90:1207–1216
Haswell PM (2019) Behind enemy lines: investigating suppression & coexistence between sympatric carnivores in Plitvice Lakes, Croatia. PhD Thesis, School of Natural Sciences, Bangor University, UK
Haswell PM, Kusak J, Hayward MW (2017) Large carnivore impacts are context-dependent. Food Webs 12:3–13
Haswell PM, Jones KA, Kusak J, Hayward MW (2018) Fear, foraging and olfaction: how mesopredators avoid costly interactions with apex predators. Oecologia 187:573–583
Hayward M, Slotow R (2009) Temporal partitioning of activity in large African carnivores: tests of multiple hypotheses. S Afr J Wildl Res 39:109–125
Hebblewhite M, White CA, Nietvelt CG, Mckenzie JA, Hurd TE, Fryxell JM, Bayley SE, Paquet PC (2005) Human activity mediates a trophic cascade caused by wolves. Ecology 86:2135–2144
Heesy CP, Hall MI (2010) The nocturnal bottleneck and the evolution of mammalian vision. Brain Behav Evol 75:195–203
Heurich M, Hilger A, Kuchenhoff H et al (2014) Activity patterns of Eurasian lynx are modulated by light regime and individual traits over a wide latitudinal range. PLoS One 9:e114143
Holt RD, Polis GA (1997) A theoretical framework for intraguild predation. Am Nat 149:745–764
IBM Corp (2017) IBM SPSS statistics for windows, Version 25.0. IBM Corp, Armonk, NY
Jones KR, Venter O, Fuller RA, Allan JR, Maxwell SL, Negret PJ, Watson JEM (2018) One-third of global protected land is under intense human pressure. Science 360:788–791
Kamler JF, Stenkewitz U, Macdonald DW (2013) Lethal and sublethal effects of black-backed jackals on cape foxes and bat-eared foxes. J Mammal 94:295–306
Kohl MT, Stahler DR, Metz MC, Forester JD, Kauffman MJ, Varley N, White PJ, Smith DW, MacNulty DR (2018) Diel predator activity drives a dynamic landscape of fear. Ecol Monogr 88:638–652
Kronfeld-Schor N, Dayan T (2003) Partitioning of time as an ecological resource. Annu Rev Ecol Evol Syst 34:153–181
Kusak J, Skrbinšek A, Huber D (2005) Home ranges, movements, and activity of wolves (Canis lupus) in the Dalmatian part of Dinarids, Croatia. Eur J Wildlife Res 51:254–262
Leo V, Reading RP, Letnic M (2015) Interference competition: odours of an apex predator and conspecifics influence resource acquisition by red foxes. Oecologia 179:1033–1040
Lima SL, Bednekoff PA (1999) Temporal variation in danger drives antipredator behavior: the predation risk allocation hypothesis. Am Nat 153:649–659
Linkie M, Ridout MS (2011) Assessing tiger-prey interactions in Sumatran rainforests. J Zool 284:224–229
Meredith M, Ridout M (2018a) Overview of the overlap package, https://cran.r-project.org/web/packages/overlap/vignettes/overlap.pdf
Meredith M, Ridout M (2018b) R package ‘overlap’: estimates of coefficient of overlapping for animal activity patterns, version 0.3.2, https://cran.r-project.org/web/packages/overlap/index.html
Monterroso P, Alves PC, Ferreras P (2013) Catch me if you can: diel activity patterns of mammalian prey and predators. Ethology 119:1044–1056
Monterroso P, Alves PC, Ferreras P (2014) Plasticity in circadian activity patterns of mesocarnivores in southwestern Europe: implications for species coexistence. Behav Ecol Sociobiol 68:1403–1417
Newsome TM, Ripple WJ (2014) A continental scale trophic cascade from wolves through coyotes to foxes. J Anim Ecol 84:49–59
Nouvellet P, Rasmussen GSA, Macdonald DW, Courchamp F (2012) Noisy clocks and silent sunrises: measurement methods of daily activity pattern. J Zool 286:179–184
Owen MA, Swaisgood RR, Blumstein DT (2017) Contextual influences on animal decision-making: significance for behavior-based wildlife conservation and management. Integr Zool 12:32–48
Palomares F, Caro T (1999) Interspecific killing among mammalian carnivores. Am Nat 153:492–508
Pauli BP, Spaul RJ, Heath JA (2017) Forecasting disturbance effects on wildlife: tolerance does not mitigate effects of increased recreation on wildlands. Anim Conserv 20:251–260
Preisser EL, Bolnick DI, Benard MF (2005) Scared to death? The effects of intimidation and consumption in predator-prey interactions. Ecology 86:501–509
Prugh LR, Stoner CJ, Epps CW, Bean WT, Ripple WJ, Laliberte AS, Brashares JS (2009) The rise of the mesopredator. Bioscience 59:779–791
R Core Development Team (2008) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria, http://www.R-project.org
Rayan DM, Linkie M (2016) Managing conservation flagship species in competition: tiger, leopard and dhole in Malaysia. Biol Conserv 204:360–366
Reimchen TE (1998) Nocturnal foraging behaviour of black bears, Ursus americanus, on Moresby Island, British Columbia. Can Field-Nat 112:446–450
Relógio A, Westermark PO, Wallach T, Schellenberg K, Kramer A, Herzel H (2011) Tuning the mammalian circadian clock: robust synergy of two loops. PLoS Comput Biol 7:e1002309
Ridout MS, Linkie M (2009) Estimating overlap of daily activity patterns from camera trap data. J Agric Biol Environ Stat 14:322–337
Ritchie EG, Johnson CN (2009) Predator interactions, mesopredator release and biodiversity conservation. Ecol Lett 12:982–998
Robinson QH, Bustos D, Roemer GW (2014) The application of occupancy modeling to evaluate intraguild predation in a model carnivore system. Ecology 95:3112–3123
Roemer GW, Gompper ME, Valkengurgh BV (2009) The ecological role of the mammalian mesocarnivore. Bioscience 59:165–173
Romanić SH, Kljaković-Gašpić Z, Bituh T et al (2016) The impact of multiple anthropogenic contaminants on the terrestrial environment of the Plitvice Lakes National Park, Croatia. Environ Monit Assess 188:27
Rowcliffe JM, Kays R, Kranstauber B, Carbone C, Jansen PA (2014) Quantifying levels of animal activity using camera trap data. Methods Ecol Evol 5:1170–1179
Santulli G, Palazon S, Melero Y, Gosalbez J, Lambin X (2014) Multi-season occupancy analysis reveals large scale competitive exclusion of the critically endangered European mink by the invasive non-native American mink in Spain. Biol Conserv 176:21–29
Sarmento WM, Berger J (2017) Human visitation limits the utility of protected areas as ecological baselines. Biol Conserv 212:316–326
Schoener TW (1974a) Resource partitioning in ecological communities. Science 185:27–39
Schoener TW (1974b) The compression hypothesis and temporal resource partitioning. Proc Natl Acad Sci USA 71:4169–4172
Shannon G, Lewis JS, Gerber BD (2014) Recommended survey designs for occupancy modelling using motion-activated cameras: insights from empirical wildlife data. PeerJ 2:e532
Šikić Z (2007) Plitvice Lakes National Park management plan. Ministry of Culture of the Republic of Croatia, Zagreb
Slavica A, Severin K, Čač Ž, Cvetnić Ž, Lojkić M, Dež-dek D, Konjević D, Pavlak M, Budinšćak Z (2010) Model širenja silvatične bjesnoće na teritoriju Republike Hrvatske tijekom perioda od trideset godina. Vet Stanica 41:199–210
Smedshaug CA, Selas V, Lund SE, Sonerud GA (1999) The effect of a natural reduction of red fox Vulpes vulpes on small game hunting bags in Norway. Wildlife Biol 5:157–166
Smith B (2018) Life stage and season affect temporal responses of brown bears (Ursus arctos) to human disturbance. MSc Thesis, Department of Life Sciences, Imperial College London, UK
Smith JA, Suraci JP, Clinchy M, Crawford A, Roberts D, Zanette LY, Wilmers CC (2017) Fear of the human ‘super predator’ reduces feeding time in large carnivores. Proc R Soc B 284:20170433
Strasser EH, Heath JA (2013) Reproductive failure of a human-tolerant species, the American kestrel, is associated with stress and human disturbance. J Appl Ecol 50:912–919
Štrbenac A, Huber D, Kusak J, Majić-Skrbinšek A, Frković A, Štahan Ž, Jeremić-Martinko J, Desnica S, Štrbenac P (2005) Wolf management plan for Croatia. State Institute for Nature Protection, Zagreb
Swanson A, Caro T, Davies-Mostert H, Mills MGL, Macdonald DW, Borner M, Masenga E, Packer C (2014) Cheetahs and wild dogs show contrasting patterns of suppression by lions. J Anim Ecol 83:1418–1427
Swanson A, Arnold T, Kosmala M, Forester J, Packer C (2016) In the absence of a “landscape of fear”: how lions, hyenas, and cheetahs coexist. Ecol Evol 6:8534–8545
Theuerkauf J (2009) What drives wolves: fear or hunger? Humans, diet, climate and wolf activity patterns. Ethology 115:649–657
Torretta E, Serafini M, Puopolo F, Schenone L (2016) Spatial and temporal adjustments allowing the coexistence among carnivores in Liguria (N-W Italy). Acta Ethol 19:123–132
Vance-Chalcraft HD, Rosenheim JA, Vonesh JR, Osenberg CW, Sih A (2007) The influence of intraguild predation on prey suppression and prey release: a meta-analysis. Ecology 88:2689–2696
Veilleux CC, Kirk EC (2014) Visual acuity in mammals: effects of eye size and ecology. Brain Behav Evol 83:43–53
Walther FR (1969) Flight behaviour and avoidance of predators in Thomson's gazelle (Gazella thomsoni Guenther 1884). Behaviour 34:184–220
Acknowledgements
We thank Nacionalni Park Plitvička Jezera who provided accommodation and logistical support, particularly those staff members (Ivica Matovina, Željko Rendulić and Dalibor Vuković) who helped maintain cameras. Many thanks go to J.P. Kamp for assistance with data entry and Dr. M. Rowcliffe for advice on data handling. We thank Dr. T.E. Reimchen and B.R. Smith for discussions on the subject that have resultantly assisted this work. We thank Dr. P.J. Baker and Dr. G. Shannon for comments on a prior version of the script. We also kindly thank the anonymous reviewers for helping improve the script.
Funding
This work received funding from Nacionalni Park Plitvička Jezera (Plitvice Lakes National Park), The Bernd Thies Foundation, EuroNatur, Bangor University, The UK Wolf Conservation Trust, The Coalbourn Charitable Trust, Ann Vernon Memorial Travel Fund and Sir Ian McKellen.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Ethical approval
All applicable institutional and/or national guidelines for the care and use of animals were followed. The study was carried out with the permission of Bangor University and the University of Zagreb. In Croatia, no permits were required for the use of motion activated cameras but field work was conducted with the permission of Nacionalni Park Plitvička Jezera.
Additional information
Communicated by T. Stankowich
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Haswell, P.M., Kusak, J., Jones, K.A. et al. Fear of the dark? A mesopredator mitigates large carnivore risk through nocturnality, but humans moderate the interaction. Behav Ecol Sociobiol 74, 62 (2020). https://doi.org/10.1007/s00265-020-02831-2
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-020-02831-2