
Abstract
Purpose
This study investigated which of the reciprocal stimuli between articular chondrocytes (ACs) and mesenchymal stem cells (MSCs) played the more important role in enhancing cartilage matrix formation, and examined the relative importance of physical contact and soluble factors in the co-culture system.
Methods
Rat ACs and bone marrow MSCs with green fluorescent protein (GFP-BMSCs) were co-cultured in vitro with or without direct cell–cell contact at the ratio of 2:1. After co-culturing in direct cell–cell contact, ACs and GFP-BMSCs were separated by flow cytometry. The effects of different co-culture methods were analysed by quantitative real-time reverse transcription-polymerase chain reaction (RT-PCR) and western blotting.
Results
SOX-9, COL2 and aggrecan mRNA levels and protein expression in ACs co-cultured with direct cell–cell contact were significantly higher than in ACs co-cultured without direct cell–cell contact; and similar results were found in GFP-BMSCs. After co-culture either with or without direct cell–cell contact, mRNA levels and protein expression of SOX-9, COL2 and aggrecan in GFP-BMSCs were significantly lower than in ACs in the equivalent co-culture systems. Though the expression of chondrocyte-specific proteins in GFP-BMSCs was enhanced, the protein expression was still much lower than in ACs cultured alone.
Conclusions
Reciprocal interactions exist between ACs and BMSCs in co-culture. The stimulating and supporting effects of BMSCs on ACs were more important in enhancing cartilage-matrix formation than the reciprocal effect of ACs on BMSCs. Both soluble factors and direct physical contact occur in AC/BMSC co-cultures, with physical contact playing a predominant, or at least very important role.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Articular chondrocytes (ACs) and mesenchymal stem cells (MSCs) are common sources of seeding cells for cartilage-tissue engineering. However, each is associated with disadvantages when applied individually to the construction of tissue-engineered cartilage. Thus, MSCs have been co-cultured with ACs to reduce the number of required chondrocytes, promote chondrogenic differentiation of MSCs and enhance cartilage-matrix formation [1–6]. However, the mechanisms behind the interactions between ACs and bone marrow mesenchymal stem cells (BMSCs) in co-culture have not been fully clarified.
Most previous studies have focused mainly on the induction and promotion of MSC chondrogenesis via co-culture with chondrocytes. Chondrocytes can secrete numerous differentiation and growth factors to induce and promote the chondrogenic differentiation of MSCs [7–9]. The trophic function of MSCs in stimulating and supporting chondrocytes has also recently been investigated [10]. However, the relative importance of the reciprocal stimuli between ACs and MSCs in enhancing cartilage-matrix formation remains unknown.
Previous studies have demonstrated effects of both soluble factors [8, 11–13] and direct physical contact [5, 14, 15] in co-culture systems. Both these represent important aspects of the interactions between ACs and MSCs. However, the relative roles of these methods of interaction in co-cultures are still unclear.
The aim of this study was to investigate which of the reciprocal stimuli between ACs and MSCs played the more important role in enhancing cartilage matrix formation, and to examine the relative importances of physical contact and soluble factors in the co-culture system.
Materials and methods
All experimental procedures involving animals conformed with the National Institutes of Health (NIH) Guidelines for the Care and Use of Laboratory Animals and were approved by the Administration Committee of Experimental Animals, Jiangsu Province, China.
-
1.
Isolation and culture of ACs
Articular cartilage was harvested in a sterile environment from the weight-bearing section of the distal femur of Sprague-Dawley rats. ACs were separated, digested and cultured as described previously [16].
-
2.
Isolation, culture and characterisation of BMSCs
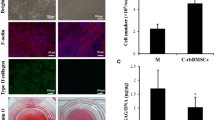
BMSCs from four-week-old green fluorescent protein (GFP)-transgenic Sprague-Dawley rats (supplied by Jiangsu Key Laboratory of Neuroregeneration, Nantong University) were harvested and cultured as described previously [17]. BMSCs at passage three were induced into adipogenic [18], osteogenic [18] and chondrogenic lineages [19], respectively. (Fig. 1)

Cell culture and characterisation of bone marrow mesenchymal stem cells (BMSCs). a ACs. (b & c) GFP-BMSCs. (d–f) Characterisation of green fluorescent protein (GFP)-BMSCs. d Oil Red O staining. e Alizarin red staining. f Alcian blue staining
-
3.
Co-culture and study design
Primary ACs and passage-3 GFP-BMSCs were used in the co-culture experiments at a ratio of 2:1 [16]. For co-culture without direct cell–cell contact, GFP-BMSCs were seeded onto Transwell inserts (six-well plates, BD Biosciences) with a 0.4-μm porous membrane and lowered into the wells with ACs. For co-culture with direct cell–cell contact, ACs and GFP-BMSCs were mixed directly and then seeded onto six-well plates. ACs alone seeded onto six-well plates and GFP-BMSCs alone seeded onto inserts were used as controls. The cell numbers seeded onto the wells and inserts are listed in Table 1. Cells were cultured in Dulbecco’s modified Eagle’s medium: Nutrient Mixture F-12 (DMEM/F12; Gibco) containing 2 % foetal bovine serum (FBS; Gibco), 100 U/mL penicillin and 100 U/mL streptomycin. Co-cultures were maintained for three weeks and the medium was changed every three days.
Both ACs and GFP-BMSCs were separated by flow cytometry on day 21 of co-culture with direct cell–cell contact. The following cell samples were harvested: (1) ACs cultured alone; (2) ACs co-cultured without direct cell–cell contact; (3) ACs co-cultured with direct cell–cell contact; (4) GFP-BMSCs cultured alone; (5) GFP-BMSCs co-cultured without direct cell–cell contact; (6) GFP-BMSCs co-cultured with direct cell–cell contact.
-
4.
Quantitative real-time RT-PCR
Total RNA was extracted from cell samples using Trizol (Invitrogen). cDNA was synthesised from total RNA using an Omniscript RT kit (Qiagen). The mRNA levels for SRY (sex determining region Y)-box 9 (SOX-9), type II collagen (COL2), aggrecan, type I collagen (COL1) and type X collagen (COL10) were measured by quantitative real-time RT-PCR (Stepone real-time PCR Applied Biosystems) as described previously [16]. PCR was performed using specific primers designed from the published sequence of each cDNA, as listed in Table 2.
-
5.
Western blotting
The expression levels of SOX-9, COL2, aggrecan, COL10 and COL1 proteins from cell samples were analysed as described previously [16]. All the antibodies used for western blotting were from Abcam.
-
6.
Statistical analysis
The statistical significance of differences was determined by one-way analysis of variance (ANOVA). The significance level was set at α= 0.05. IBM SPSS Statistics 19.0 was used for data analyses.
Results
mRNA levels and protein expression of SOX-9, COL2 and aggrecan were significantly upregulated in both BMSCs and ACs co-cultured with or without direct cell–cell contact, compared with ACs and BMSCs cultured alone (p< 0.05, Figs. 2a–c and 3a–c). SOX-9, COL2 and aggrecan mRNA levels and protein expression in ACs co-cultured with direct cell–cell contact were significantly higher than in ACs co-cultured without direct cell–cell contact (p< 0.05, Figs. 2a–c and 3a–c). Similarly, the mRNA and protein expression of SOX-9, COL2 and aggrecan in GFP-BMSCs co-cultured with direct cell–cell contact were also significantly higher than in GFP-BMSCs co-cultured without direct cell–cell contact (p< 0.05, Figs. 2a–c and 3a–c). After co-culture either with or without direct cell–cell contact, mRNA levels and protein expression of SOX-9, COL2 and aggrecan in GFP-BMSCs were significantly lower than in ACs in the equivalent co-culture systems (p< 0.05, Figs. 2a–c and 3a–c). Though the expression of chondrocyte-specific proteins in GFP-BMSCs was enhanced, the protein expression was still much lower than in ACs cultured alone (Fig. 3a–c). No significant mRNA and protein expression of COL10 was observed in ACs in any of the six samples (Figs. 2d & 3d), but slight COL1 protein expression was observed in ACs (Fig. 3e). COL10 mRNA and protein expression by GFP-BMSCs was significantly downregulated after co-culturing (p< 0.05, Figs. 2d and 3d). COL1 mRNA level by BMSCs was upregulated (p< 0.05, Fig. 2e), and slight COL1 protein expression was detected in GFP-BMSCs co-cultured with direct cell–cell contact (Fig. 3e).

Quantitative analysis of SOX-9 (a), COL2 (b), Aggrecan (c), COL10 (d) and COL1 (e) gene expression. Statistically significant differences were found by one-way ANOVA in all five genes. *p < 0.05, with a statistically significant difference between the two groups

Protein expression of SOX-9 (a), COL2 (b), Aggrecan (c), COL10 (d) and COL1 (e). (1) ACs cultured alone. (2) ACs co-cultured without direct cell–cell contact. (3) ACs co-cultured with direct cellcell contact. (4) GFP-BMSCs cultured alone. (5) GFP-BMSCs co-cultured without direct cell–cell contact. (6) GFP-BMSCs co-cultured with direct cell–cell contact
Discussion
The ratio of chondrocytes to MSCs is an important factor in co-culture systems, and establishing the optimal ratio is key to successful co-culturing and construction of high-quality tissue-engineered cartilage [16]. Different experimental conditions of co-culture may lead to different optimal ratios. Richardson et al. [5] co-cultured human nucleus pulposus (NP) cells with human BMSCs at ratios of 3:1, 1:1 and 1:3, and found that co-culturing at a ratio of 3:1 increased mRNA expression of chondrocyte-specific markers. Fischer et al. [8] co-cultured human ACs with human BMSCs at ratios of 2:1 and 1:1, and found that co-culturing at a ratio of 2:1 increased the gene expression ratio of COL2 to COL10. Yang et al. [6] found that a chondrocyte/MSC-ratio of 63:1 was identified as the appropriate proportion that chondrocytes drive MSCs in chondrogenic development. In our previous study, ACs and BMSCs from rabbits were co-cultured at ratios of 4:1, 2:1, 1:1, 1:2 and 1:4, and 2:1 was identified as the optimal ratio of ACs to BMSCs [16]. The culture conditions in our latest experiment were similar to those in our previous study [16], and an AC:BMSC ratio of 2:1 was therefore adopted in this study.
The idea of using MSCs for cartilage-tissue engineering originated because of their extensive proliferation capacity and chondrogenic differentiation potential. However, the benefits of using MSCs in tissue repair cannot be explained fully by their differentiation into tissue-specific cells. Our study demonstrated the existence of reciprocal interactions between ACs and BMSCs during co-culture; ACs could not only induce and promote the chondrogenic differentiation of BMSCs, but BMSCs could also stimulate and support ACs to express more chondrocyte-specific proteins.
However, the relative importance of the reciprocal stimuli between ACs and MSCs in enhancing cartilage matrix formation remains unresolved. Wu et al. [10] considered that cartilage-matrix deposition originated mainly from bovine ACs, and that the primary function of BMSCs was to provide nutrition in this xenogeneic co-culture system. However, their results may have been affected by the use of heterogeneic, rather than allogeneic cells, which might have hampered the response between different types of cells in co-culture. In this study, rat ACs and rat GFP-BMSCs were co-cultured. Our results suggest that ACs may contribute more to cartilage-matrix formation in co-culture systems; i.e., compared with the inducing effect of ACs on BMSCs, the stimulating and supporting effects of BMSCs on ACs may play a more important role in enhancing cartilage-matrix formation.
In most co-culture experiments without direct cell–cell contact, the effect of soluble factors alone was sufficient to enhance cartilage-matrix formation and promote chondrogenic differentiation of MSCs. However, Richardson et al. [5] co-cultured human nucleus pulposus (NP) cells with human BMSCs for seven days and found that co-culturing without direct cell–cell contact resulted in no significant changes in matrix-gene expression, while co-culture with direct cell–cell contact significantly upregulated matrix-gene expression by both NP cells and MSCs. However, the co-culture duration was only seven days, which may partly explain why co-culture without direct cell–cell contact had no significant effect in their experiments. Our results demonstrated that the effects of both soluble factors and direct physical contact play roles in the co-culture system.
However, the relative importance of soluble factors and direct physical contact in enhancing cartilage-matrix formation remains unclear. Yamamoto et al. [14] co-cultured human NP cells with human BMSCs with or without direct cell–cell contact and reported increased expression of numerous soluble factors in co-cultures with direct cell–cell contact, compared to co-cultures without direct cell–cell contact. Concentrations of these soluble factors in conditional media have also been reported to be much lower than those used for conventional chondrogenic induction, and too low to induce efficient chondrogenic differentiation of BMSCs [20]. These results imply that the effects of soluble factors may be limited in co-cultures. The results of our study showed that mRNA expression levels of chondrocyte-specific markers in both ACs and GFP-BMSCs co-cultured with direct cell–cell contact were significantly upregulated (2–3-fold higher), compared with co-culturing without direct cell–cell contact. Co-culture with direct cell–cell contact also increased the expression of chondrocyte-specific proteins compared to co-cultures without direct cell–cell contact. These results suggest that direct physical contact plays a predominant, or at least very important role, in enhancing cartilage-matrix formation and promoting BMSC chondrogenesis in co-cultures.
Conclusion
In AC/BMSC co-cultures, ACs can induce and promote the chondrogenic differentiation of BMSCs, while BMSCs can stimulate and support ACs to express more chondrocyte-specific proteins. Compared to the inducing effect of ACs on BMSCs, the stimulating and supporting effects of BMSCs on ACs may play a more important role in enhancing cartilage-matrix formation. Soluble factors and direct physical contact both exert effects in co-culture, and both have a beneficial influence on chondrogenesis of BMSCs and cartilage-matrix formation. Direct physical contact appears to play a predominant, or at least very important role in AC/BMSC co-culture systems.
References
Mo XT, Guo SC, Xie HQ et al (2009) Variations in the ratios of co-cultured mesenchymal stem cells and chondrocytes regulate the expression of cartilaginous and osseous phenotype in alginate constructs. Bone 45(1):42–51
Gruber HE, Deepe R, Hoelscher GL et al (2010) Human adipose-derived mesenchymal stem cells: direction to a phenotype sharing similarities with the disc, gene expression profiling, and coculture with human annulus cells. Tissue Eng Part A 16(9):2843–2860
Chen WH, Lai MT, Wu AT et al (2009) In vitro stage-specific chondrogenesis of mesenchymal stem cells committed to chondrocytes. Arthritis Rheum 60(2):450–459
Bigdeli N, Karlsson C, Strehl R et al (2009) Coculture of human embryonic stem cells and human articular chondrocytes results in significantly altered phenotype and improved chondrogenic differentiation. Stem Cells 27(8):1812–1821
Richardson SM, Walker RV, Parker S et al (2006) Intervertebral disc cell-mediated mesenchymal stem cell differentiation. Stem Cells 24(3):707–716
Yang YH, Lee AJ, Barabino GA (2012) Coculture-driven mesenchymal stem cell-differentiated articular chondrocyte-like cells support neocartilage development. Stem Cells Transl Med 1(11):843–854
Kim JS, Ryoo ZY, Chun JS (2007) Cytokine-like 1 (Cytl1) regulates the chondrogenesis of mesenchymal cells. J Biol Chem 282(40):29359–29367
Fischer J, Dickhut A, Rickert M et al (2010) Human articular chondrocytes secrete parathyroid hormone-related protein and inhibit hypertrophy of mesenchymal stem cells in coculture during chondrogenesis. Arthritis Rheum 62(9):2696–2706
Keller B, Yang T, Chen Y et al (2011) Interaction of TGFbeta and BMP signaling pathways during chondrogenesis. PLoS One 6(1):e16421
Wu L, Leijten JC, Georgi N et al (2011) Trophic effects of mesenchymal stem cells increase chondrocyte proliferation and matrix formation. Tissue Eng Part A 17(9–10):1425–1436
Hwang NS, Varghese S, Puleo C et al (2007) Morphogenetic signals from chondrocytes promote chondrogenic and osteogenic differentiation of mesenchymal stem cells. J Cell Physiol 212(2):281–284
Hwang NS, Im SG, Wu PB et al (2011) Chondrogenic priming adipose-mesenchymal stem cells for cartilage tissue regeneration. Pharm Res 28(6):1395–1405
Aung A, Gupta G, Majid G et al (2011) Osteoarthritic chondrocyte-secreted morphogens induce chondrogenic differentiation of human mesenchymal stem cells. Arthritis Rheum 63(1):148–158
Yamamoto Y, Mochida J, Sakai D et al (2004) Upregulation of the viability of nucleus pulposus cells by bone marrow-derived stromal cells: significance of direct cell-to-cell contact in coculture system. Spine (Phila Pa 1976) 29(14):1508–1514
Heng BC, Cao T, Lee EH (2004) Directing stem cell differentiation into the chondrogenic lineage in vitro. Stem Cells 22(7):1152–1167
Qing C, Wei-ding C, Wei-min F (2011) Co-culture of chondrocytes and bone marrow mesenchymal stem cells in vitro enhances the expression of cartilaginous extracellular matrix components. Braz J Med Biol Res 44(4):303–310
Wei A, Chung SA, Tao H et al (2009) Differentiation of rodent bone marrow mesenchymal stem cells into intervertebral disc-like cells following coculture with rat disc tissue. Tissue Eng Part A 15(9):2581–2595
Schilling T, Noth U, Klein-Hitpass L et al (2007) Plasticity in adipogenesis and osteogenesis of human mesenchymal stem cells. Mol Cell Endocrinol 271(1–2):1–17
Johnstone B, Hering TM, Caplan AI et al (1998) In vitro chondrogenesis of bone marrow-derived mesenchymal progenitor cells. Exp Cell Res 238(1):265–272
Liu X, Sun H, Yan D et al (2010) In vivo ectopic chondrogenesis of BMSCs directed by mature chondrocytes. Biomaterials 31(36):9406–9414
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 81000800) and the Natural Science Foundation of Jiangsu Province (No.BK2011843).
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Qiang Zuo and Weiding Cui contributed equally to this work.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Zuo, Q., Cui, W., Liu, F. et al. Co-cultivated mesenchymal stem cells support chondrocytic differentiation of articular chondrocytes. International Orthopaedics (SICOT) 37, 747–752 (2013). https://doi.org/10.1007/s00264-013-1782-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00264-013-1782-z