
Abstract
Therapeutic vaccination with dendritic cells (DC) is an emerging investigational therapy for eradication of minimal residual disease in acute myeloid leukemia. Various strategies are being explored in manufacturing DC vaccines ex vivo, e.g., monocyte-derived DC (MoDC) loaded with leukemia-associated antigens (LAA). However, the optimal source of LAA and the choice of DC-activating stimuli are still not well defined. Here, loading with leukemic cell preparations (harboring both unknown and known LAA) was explored in combination with a DC maturation-inducing cytokine cocktail (CC; IL-1β, IL-6, TNF-α, and PGE2) and Toll-like receptor ligands (TLR-L) to optimize uptake. Since heat shock induced apoptotic blasts were more efficiently taken up than lysates, we focused on uptake of apoptotic leukemic cells. Uptake of apoptotic blast was further enhanced by the TLR7/8-L R848 (20–30%); in contrast, CC-induced maturation inhibited uptake. CC, and to a lesser extent R848, enhanced the ability of MoDC to migrate and stimulate T cells. Furthermore, class II-associated invariant chain peptide expression was down-modulated after R848- or CC-induced maturation, indicating enhanced processing and presentation of antigenic peptides. To improve both uptake and maturation, leukemic cells and MoDC were co-incubated with R848 for 24 h followed by addition of CC. However, this approach interfered with CC-mediated MoDC maturation as indicated by diminished migratory and T cell stimulatory capacity, and the absence of IL-12 production. Taken together, our data demonstrate that even though R848 improved uptake of apoptotic leukemic cells, the sequential use of R848 and CC is counter-indicated due to its adverse effects on MoDC maturation.
Similar content being viewed by others

Introduction
Acute myeloid leukemia (AML) is characterized by the proliferation of clonal neoplastic myeloid hematopoietic precursor cells and impaired normal hematopoiesis. Although 70–80% of patients (<60 years) achieve complete remission after intensive chemotherapy, AML frequently relapses due to the persistence of minimal residual disease (MRD) [1]. Escape of leukemic cells from immune surveillance has been associated with the poor clinical outcome. For instance, a high occupancy of HLA-DR molecules with the class II-associated invariant chain peptide (CLIP) instead of an antigenic peptide is correlated with a shortened disease-free and overall survival [2, 3]. In contrast, immune control of leukemia, as shown for instance by the graft-versus-leukemia effect induced by allogeneic stem cell transplantation (SCT) or the reinduction of complete remission after donor lymphocyte infusion following allogeneic SCT, demonstrates the potential of exploiting the immune system in aid of anti-AML therapy [1].
The induction, regulation, and maintenance of primary immune responses, including specific anti-tumor T cell responses are coordinated by dendritic cells (DC). Vaccination with DC has been recognized as a promising investigational therapy due to the uniquely powerful antigen (Ag)-presenting capacity of DC and its potential to circumvent immunosuppressive features of leukemia [4]. The first steps down the road to DC vaccination in AML have been taken and results from small clinical trials have been reported [5]. The general lack of clinical responses evoked important questions concerning the optimal methodologies for DC vaccine preparation as well as the design of clinical vaccination protocols. Many strategies are explored in the preparation of DC vaccines ex vivo; among these, autologous monocyte-derived DC (MoDC) loaded with leukemia-associated Ag (LAA) are promising [6]. Various sources of LAA and different methods of loading LAA onto DC have been explored in an attempt to optimize anti-tumor responses [7]. For AML, several relevant LAA have been identified including PRAME, RHAMM, WT1, and PR1. Unfortunately, overexpression of these LAA is common, but not uniform in leukemia [8]. Moreover, HLA restriction of LAA-derived peptides limits application of such vaccines to patients with certain HLA profiles. These restrictions inherent to the use of defined LAA or LAA-derived peptides may be overcome using the whole AML cells as a source of LAA, for instance by generation of AML lysates or apoptotic leukemic cells. Among other whole AML cell-derived antigen loading strategies that have been explored is electroporation of DC with AML-derived RNA [9]. Also vaccination with modified AML cells, such as AML-derived DC or fusions between AML cells and DC, has been investigated; further modification of DC with 4-1BB-L or CD40 might enhance the efficacy of such vaccines [10–13]. It has long been assumed that the apoptotic cell death is poorly immunogenic or even tolerogenic, whereas necrotic cell death is considered to be immunogenic. However, stress-induced heat shock protein (HSP)-peptide complexes (commonly induced during apoptosis) are more efficiently taken up via scavenger receptors and Toll-like receptors (TLR) on the DC surface and induce efficient cross-priming and skewing of the immune response towards a Th1-type profile [14, 15], whereas necrosis has been associated with the local immune suppression in solid tumors [16]. Furthermore, apoptosis induction after irradiation with UV light or by treatment with chemotherapeutic drugs results in upregulation of calreticulin, a scavenger receptor class-A ligand associated with immunogenic apoptosis as demonstrated for colon carcinoma cells [17, 18]. For AML, it is not yet clear whether cell lysates or apoptotic cells are preferable for Ag loading onto MoDC [9, 19–25]. Next to enhancing immunogenicity of tumor Ag sources by, for instance, heat shock, addition of DC-maturing stimuli, such as Toll-like receptor ligands (TLR-L), is explored; e.g., electroporation of TLR3-L Poly(I:C) into AML cells results in enhanced uptake of leukemic cells by DC and improves their subsequent maturation and cytokine production [26]. Furthermore, intracellular binding of TLR8 by its ligand R848 has been reported to result in enhanced cross-priming of exogenous Ag by MoDC [27]. Various whole AML cell preparations loaded onto MoDC in combination with DC maturation-inducing cocktails have been explored [9, 21]; however, the quantitative effects of TLR-L and cytokines on uptake of leukemic cells are still unclear.
In this study, we compared the uptake of allogeneic apoptotic leukemic cells with lysates derived from leukemic cells. We investigated the uptake of heat shock-induced apoptotic leukemic cells by MoDC and DC maturation by combining a standard cytokine cocktail (CC) with the clinically applicable TLR7/8-L R848.
Materials and methods
Patient samples
Peripheral blood or bone marrow mononuclear cells from AML patients were isolated by density centrifugation using Ficoll-Paque (Amersham Pharmacia Biotech, Uppsala, Sweden). All samples were drawn after obtaining the patients’ informed consent at the time of collection. Cells were cultured immediately or cryopreserved at a controlled rate in liquid nitrogen using RPMI-1640 medium (Gibco, Paisley, UK) supplemented with 20% heat-inactivated FCS (Greiner, Alphen a/d Rijn, The Netherlands) and 10% dimethylsulphoxide (Merck, Darmstadt, Germany). Before application of stored AML cells, cryopreserved material was rapidly thawed and washed twice in RPMI-1640 supplemented with 40% FCS. The cells were then resuspended in culture medium as described below.
Culture of monocyte-derived dendritic cells
Buffy coats were obtained from healthy volunteers according to the institutional guidelines. Peripheral blood mononuclear cells (PBMC) were isolated by density centrifugation using Ficoll-Paque. Subsequently, monocytes were obtained by magnetic bead isolation, using CD14-labeled microbeads (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany). MoDC were cultured from CD14+ monocytes in CellGro serum-free medium (CG) (CellGenix, Freiburg, Germany) supplemented with 100 IU/ml penicillin and 100 μg/ml streptomycin, GM-CSF (800 IU/ml, specific activity 1 × 105 IU/mg protein; Pepro Tech, Rocky Hill, NJ), and IL-4 (500 IU/ml, specific activity 1 × 107 IU/mg protein, Pepro Tech). After 5 days, immature MoDC were harvested. Purity and phenotype of immature DC were evaluated by flow cytometry for the expression of CD14, CD40, CD34, CD80, CD86, HLA-DR, CD54, and CD83. For maturation induction, the following TLR-L were used: R848 (3 μg/ml; Alexis Biochemicals, Axxora, Grünberg, Germany), LPS (100 ng/ml; Sigma-Aldrich, Saint Louis, MO), polyinosinic–polycyticylic acid potassium salt (Poly(I:C); 25 μg/ml; Sigma-Aldrich), flagellin (5 μg/ml; Invivogen, San Diego, CA), and peptidoglycan (PGN; 5 μg/ml; Sigma-Aldrich). Maturation with CC was induced using the following combination of cytokines and reagents: recombinant TNF-α (200 IU/ml; Strathmann Biotec, Hannover, Germany), IL-1β (10 ng/ml; Strathmann Biotec), PGE2 (1 μg/ml; Sigma-Aldrich), and IL-6 (10 ng/ml; R&D Systems, Abingdon, UK).
Loading of MoDC with allogeneic leukemic cell lysates and apoptotic leukemic cells
Leukemic cells were labeled with 5- (and 6-) carboxyfluorescein diacetate, succinimidyl ester (CFSE; Molecular Probes, Eugene, Oregon). Cells were suspended in buffered saline (PBS) at 10 × 106−1 ml and labeled for 10 min with 1 M CFSE at 37°C, washed three times with RPMI-1640, 10% FCS and incubated overnight in RPMI-1640, supplemented with 100 IU/ml penicillin and 100 μg/ml streptomycin and 10% FCS. Leukemic cells were harvested and taken up in PBS and apoptosis was induced by heat shock (2 h at 42°C) or by incubation for 2 h at 37°C with ARA-C (10 μg/ml; Mayne Pharma, Warwickshire, UK). The percentage of necrotic, apoptotic, and viable cells was determined before and after apoptosis induction by incubation with Syto-62 (5 nM; Molecular Probes, Eugene, OR), PSC833 (2 μM; Novartis, Basel, Switzerland) and 7-aminoactinomycin D (7-AAD; ViaProbe, Pharmingen, San Diego, CA) for 45 min at 37°C, and by flow cytometric analysis as described previously [28]. Syto-62 is a fluorescent nucleic acid stain that exhibits bright fluorescence upon binding to nucleic acids and is retained in viable cells. Together with the dead-cell dye 7-aminoactinomycin D (7-AAD), discrimination between viable (Syto+/7AAD-) and early (Syto-/7AAD-) and late (Syto-/7AAD+) stages of apoptosis can be made. MoDC were labeled with CellVue Plum (Polysciences, Washington, CO) or with PKH26 (Sigma, Missouri, USA) according to the manufacturers’ guidelines. After labeling, DC were washed three times and put in CG medium. MoDC were cultured in a 1:1 or 1:3 ratio with pretreated leukemic cells in CG medium combined with 100 IU/ml penicillin and 100 μg/ml streptomycin and 800 IU/ml GM-CSF and 500 IU/ml IL-4 at 37°C, 5% CO2. Using different time schedules, cultures were supplemented with the indicated maturation stimuli.
Flow cytometric analysis of MoDC
Four-color flow cytometry was performed on a FACSCalibur flow cytometer (Becton Dickinson, San Jose, CA). Results were analyzed using CellQuest software (Becton Dickinson). Immature DC and DC after maturation were stained by appropriate dilutions or mouse isotype-matched control mAb of fluorescein isothiocyanate (FITC)-labeled CD14 (Becton Dickinson, San Jose, CA), CD1a (Sanquin), CD54 (DAKO, Glostrup, Denmark). Phyco-erythrin (PE)-labeled monoclonal antibodies (mAb) were used against CD40 (Coulter Immunotech, Marseilles, France), CD80, CD86, CCR-7 (CD197, Becton Dickinson, San Jose, CA), CLIP (cerCLIP.1; Santa Cruz Biotechnology, Santa Cruz CA), CD83 (Coulter Immunotech) and peridinin chlorophyll protein (PerCP)-labeled anti-HLA-DR and CD34 (Becton Dickinson). Isotype controls used were FITC-labeled IgG1 (DAKO) and IgG2b (Sanquin), PE-labeled IgG1 (Becton Dickinson) and IgG2b (DAKO), and PerCP-labeled IgG1 and IgG2a (Becton Dickinson). For HLA-DO and HLA-DM, the following mouse mAb were used: HLA-DM (Becton Dickinson) and IgG1 isotype control (Becton Dickinson); FITC-labeled HLA-DO (Becton Dickinson) and IgG2b (Becton Dickinson). Cells were stained for DO and DM after fixation with paraformaldehyde followed by permeabilization with PBS–0.1% Saponine (Sigma-Aldrich). Results are presented as either the percentage of positive cells compared to the appropriate isotype control or the mean fluorescence index (MFI). MFI represents the mean fluorescence obtained through binding of the mAb of interest divided by the mean fluorescence of the appropriate isotype control. Relative CLIP expression was determined by calculating the ratio between CLIP and HLA-DR expression based on both the percentage and MFI of positive cells, as described previously [2].
DC migration assay
The migratory capacity of DC was evaluated in a transwell system (5 μm poresize, Corning Costar, Corning, NY, USA) under serum-free conditions. Cells were allowed to migrate 16 h towards medium with or without chemoattractant CCL19 (MIP3β, 300 ng/ml; R&D Systems, Abingdon, UK). The percentage of migrated cells was quantified by flow cytometry using a fixed amount of fluorescent beads as a reference (FlowCount Fluorospheres, Coulter, Miami, FL, USA).
CD40 ligation and cytokine release
DC were harvested, washed, and seeded in a 96-well round-bottomed plate at 40 × 103 cells in 200 μl of medium containing 10% FCS per well. CD40L-expressing cells (i.e., muCD154-transfected J558 cells) were used as stimulator cells at 40 × 103 per well. After 24 h, supernatants were harvested and analyzed for IL-12 by ELISA, as described previously [29].
Mixed leukocyte reaction
The ability of the various AML-loaded DC to stimulate T cells was tested in an allogeneic mixed leukocyte reaction (MLR). After isolation of PBMC from a buffy-coat, CD8+ cells were obtained by magnetic cell sorting with anti-CD8-labeled magnetic microbeads (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany) and used as responder cells. DC were irradiated at 30 Gy and then added to round-bottomed 96-well tissue-culture plates (Costar, Corning, NY) at various stimulator to responder ratios. All ratios were tested in triplicate; cells were cultured in RPMI containing 10% FCS. After 5 days, thymidine was added (0.4 μCi per well; Amersham Pharmacia Biotech, Buckinghamshire, UK) for 16 h, after which the cells were harvested onto fiberglass filters and 3H-thymidine incorporation was determined using a flatbed scintillation counter (Wallac, Turku, Finland).
Statistical analysis
Statistical significance of differences between the various loading and maturation procedures was determined by the paired sample Student’s t test (two-tailed). P values of <0.05 were regarded as significant.
Results
Uptake of apoptotic leukemic cells or lysates by MoDC is a dose-dependent and active process
To investigate the use of whole leukemic cells as a source of LAA for loading onto DC, we co-incubated MoDC from healthy donors (HD) with either apoptotic leukemic cells or lysates, each labeled with different fluorochromes. Apoptotic samples contained less than 25% (range 25–0%) viable cells; the ratio of early apoptotic/viable cells was five times higher after 2 h incubation at 42°C compared with the control situation (2 h at 37°C, data not shown). The percentages of apoptotic cells did not differ between the methods [incubation with Cytarabine (ARA-C) or heat shock (HS) at 42°C] used to induce apoptosis. Uptake of apoptotic cells or apoptotic cell fragments and lysates by MoDC was dose-dependent (Fig. 1a). Moreover, uptake of HS-induced apoptotic AML cells was more efficient than AML lysates (p = 0.03; paired t test, n = 6; tested in MoDC from three different HD). Fluorescence microscopic observations revealed actual uptake of AML preparations and not mere binding to DC cell surface (Fig. 1b). Co-incubation of CFSE-labeled leukemic cells with MoDC for 2 h at 37°C resulted in high levels of CFSE+ MoDC (35%), whereas co-incubation for 2 h at 0°C resulted in only low percentages of CFSE+ MoDC (5%), indicating that the observed uptake is an active endocytic process (Fig. 1c, d). The efficiency of uptake differed per patient; no correlation was observed regarding AML subtype or percentage of apoptotic cells. Since HS-induced apoptotic leukemic cells were more efficiently taken up than lysates, further analysis was focused on uptake of HS-induced apoptotic leukemic cells.

Uptake of leukemic cells by MoDC is a dose-dependent and active process. Leukemic cells were labeled with CFSE and subsequently apoptosis was induced by heat shock at 42°C or incubation with ARA-C for 2 h; lysates were generated by three freeze-thawing cycles. Immature MoDC were labeled with CellVue Plum (emitting in the FL4 channel) or with PKH-26 (emitting in the FL2 channel) and co-incubated with leukemic cells. a Uptake of leukemic cells by MoDC is depicted after 24 h of co-incubation. Results of 1:3 leukemic cell:DC ratio (filled bars) are presented as relative uptake compared to uptake at leukemic cell:DC ratio of 1:1 (open bars) (n = 14 ± SEM). b CFSE-positive MoDC after co-incubation of CFSE labeled lysate (×400). Dot plots representing uptake of CFSE-labeled leukemic cells after 2 h incubation at 0°C (c) or 37°C (d); insets represent percentage of PKH26-labeled MoDC positive for CFSE
Uptake of heat shock-induced apoptotic AML cells by MoDC is enhanced by R848 and diminished by a DC maturation-inducing cytokine cocktail
As TLR-mediated activation of DC has previously been associated with enhanced endocytosis [30], we tested the influence of the clinically applicable TLR-L Poly(I:C), and R848 and, as well as PGN, flagellin and LPS on AML cell uptake by MoDC. Administration of a maturation-inducing CC 2 h after co-incubation of MoDC with lysed or apoptotic leukemic cells resulted in a significantly lower uptake by MoDC (Fig. 2). Of all the TLR-L tested (data of Poly(I:C), R848, LPS, PGN, and flagellin not shown), only R848 was able to enhance the uptake of HS-induced apoptotic leukemic cells significantly. Of note, addition of R848 enhanced uptake of apoptotic but not of lysed AML cells by MoDC (Fig. 2). Remarkably, combined administration of R848 and CC abolished this positive effect. We hypothesized that the prolonged incubation with R848 combined with delayed administration of CC might result in higher uptake. Therefore, we pre-incubated HS-induced apoptotic leukemic cells and MoDC with R848 for 24 h, after which CC was added for 48 h (Fig. 3). In this time frame, we found more efficient uptake of HS-induced apoptotic leukemic cells by R848-treated MoDC as compared to control and CC-treated MoDC. However, this enhanced uptake was counteracted by sequential incubation with CC.

Differential effects of R848 and a cytokine cocktail on uptake of apoptotic AML cells or AML lysates. Apoptosis of AML cells was induced by heat shock at 42°C or incubation in ARA-C for 2 h. Leukemic cells and MoDC were incubated for 2 h, R848 was added for another 2 h for the R848 and R848 and cytokine cocktail (CC) conditions. Four hours after the start of co-culture, CC was added for the CC and R848 and CC condition (as depicted in the schematic illustration). Uptake was measured by flow cytometry and indicated as the percentage of CFSE-positive MoDC, 48 h after the start of co-culture. Relative uptake of leukemic cell products by MoDC is depicted, i.e., relative increase as compared to no addition of cytokines or R848. Mean percentage of CFSE-positive DC in the control group is listed in the lower left corner of each graph; horizontal lines represent means, *p < 0.05; n = 6

Differential effects of R848 and a cytokine cocktail on uptake of apoptotic AML. Uptake of leukemic cell preparations by MoDC is depicted. Apoptosis of AML cells was induced by heat shock for 2 h at 42°C. After 24 h of co-incubation of apoptotic leukemic cells and MoDC, either in presence or absence of R848, a cytokine cocktail (CC) was added. Uptake was measured by flow cytometry and indicated as the percentage of CFSE-positive MoDC, 72 h after the start of co-culture. Relative increase to no addition of cytokines or R848 is depicted. Mean percentage of CFSE-positive MoDC from the control group is listed in the lower left corner of the graph; horizontal lines represent means, *p < 0.05; **p < 0.005, n = 5
Maturation induction of AML-loaded MoDC
Next, we investigated if incubation of MoDC from HD with AML cells with or without R848 and/or CC affected their maturation state. Uptake of HS-induced apoptotic cells by immature MoDC resulted in significant decrease of CD86 and CD40 expression (Fig. 4). Addition of CC 24 h after loading resulted in MoDC maturation as shown by de novo expression of CD83 and chemokine receptor CCR7 (Fig. 4). Only partial maturation was induced after addition of R848 alone, as demonstrated by low-level CD83 expression and the absence of CCR7. Of note, pre-incubation with R848 blocked full maturation by CC as shown by significantly lower levels of CD83, CCR7, CD40, and CD86 (Fig. 4).

Immunophenotype of MoDC after co-incubation with apoptotic leukemic cells. Marker expression levels of a CD83, b CCR7, c CD86, d CD40, and e relative CLIP are shown after uptake of heat shock-induced apoptotic leukemic cells by MoDC, either in presence or absence of R848 or CC; the time intervals used are depicted in the schematic illustration in f. For CD83 and CCR7, the percentage of positive cells is shown (a, b, respectively). For CD86 and CD40, mean fluorescence indices (MFI, as calculated in relation to the isotype control) are shown (c, d, respectively) (n = 10). Fold change compared with immature MoDC (immat) in relative CLIP expression is depicted in e by calculating the ratio of CLIP and HLA-DR expression as indicated in “Material and methods” (n = 13). Horizontal lines represent mean, *p < 0.05, **p < 0.005
MHC class II antigen presentation requires the exchange of CLIP for an antigenic peptide in the peptide-binding groove, a process that is catalyzed by HLA-DM. HLA-DO, another nonclassical MHC class II molecule, is able to down-regulate the catalytic function of HLA-DM [2]. The intensity of HLA-DR varied between the different maturation stimuli (data not shown). Relative to HLA-DR, moderate CLIP down-regulation was observed upon loading of immature DC with HS-induced apoptotic AML cells, but a more profound decrease in CLIP expression was seen upon maturation induction by R848 and/or CC (Fig. 4e). These data indicate efficient HLA-DR-mediated presentation of endocytosed antigens after maturation. Both HLA-DM and -DO levels were in accordance with the observed relative CLIP levels (data not shown).
Function of heat shock-induced apoptotic AML-loaded versus unloaded MoDC
To further investigate the effects of the applied loading strategies on HD-derived MoDC, we examined their migratory and T cell stimulating capacity. Upon CC-induced maturation, the lymph node homing receptor CCR7 was upregulated (Fig. 4b). In line with this, maturation with CC resulted in a high migration rate towards the CCR7 ligand CCL19; combined maturation with R848 resulted in lower migration efficiencies (Fig. 5a).

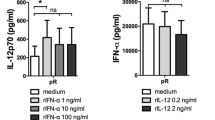
Function of unloaded and heat shock-induced apoptotic leukemic cell loaded MoDC after incubation with or without R848 and/or cytokine cocktail. a Migration of DC towards CCL19 was tested in a 0.5-μm pore transwell system; percentages of MoDC that migrated towards CCL19 (gray bars) and medium (white bars) are depicted; *p < 0.05. b IL-12p70 production was measured after overnight CD40 ligation; relative IL-12p70 release compared with immature unloaded MoDC is depicted, mean IL-12p70 release in the control group is listed in the lower left corner of the graph; *p < 0.05; **p < 0.005. c The ability of DC to induce proliferation in a mixed leukocyte reaction is shown after 5 days of co-incubation of MoDC and allogeneic CD8+ T cells in various MoDC:T cell ratios. [3H]thymidine was added and incorporation was measured after 16 h. Each ratio was tested in triplicate and means ± SD are shown. One representative experiment out of six is shown
In response to overnight CD40 ligation, HS-induced apoptotic AML-loaded MoDC stimulated with R848 or CC released reduced levels of the Th1-skewing and CTL-activating cytokine IL-12p70 in comparison to non-stimulated unloaded MoDC [mean relative decrease (±SD): 0.65 (±0.27) and 0.52 (±0.16), respectively; median IL12 production unloaded non-stimulated MoDC: 520 (286–5199) pg/ml] (Fig. 5b). Of note, IL-12p70 release was almost completely abolished by the combined sequential use of R848 and CC [mean relative decrease 0.97 (±0.06)]. However, incubation with R848 and CC increased the ability of MoDC to prime naïve T cells in an allogeneic MLR in accordance with the observations for phenotypic maturation: addition of CC after 24 h pre-incubation with R848 enhanced T cell stimulating capacity, but not to the same level as seen for CC alone (Fig. 5c).
Discussion
This study was undertaken to investigate the various MoDC vaccine preparation strategies in order to create an optimized MoDC vaccine for the treatment of MRD in AML. The ability of HD-derived MoDC to take up whole cell-derived material in combination with TLR-L was investigated in order to enhance the uptake of LAA. Furthermore, we explored the capability of MoDC to mature under the influence of various TLR-L and a standard CC as positive control.
Immature DC are capable of taking up Ag, e.g., from infected cells or tumor cells, in the form of apoptotic bodies or necrotic cell particles [31, 32]. Next to the Ag source, agents such as TLR-L or cytokines can affect the outcome of this uptake [33]. Upon encounter with for instance microbial agents or other inflammatory stimuli, DC mature from cells specialized in Ag uptake into cells specialized in T cell stimulation. Inflammatory cytokines are considered as strong maturation inducers but, as we demonstrated, CC inhibits further Ag uptake by DC. In contrast, TLR-L are known to enhance the uptake of antigenic material and subsequently induce maturation [30]. In accordance with this, we found the enhanced uptake after co-incubation with TLR- L R848, whereas addition of CC after 24 h of co-incubation with R848 halted further uptake.
The method by which leukemic cell death was induced influenced the amount of cellular material taken up by MoDC: less uptake was achieved in DC after co-incubation with lysate as compared to apoptotic cells. In the process of dying, apoptotic cells are known to shed immunogenic particles, i.e., so-called blebs. Murine studies have demonstrated that these blebs are taken up more efficiently by DC than the remaining, larger, apoptotic bodies [34]. This may offer an explanation for our observation that the apoptotic AML cells are more efficiently taken up by MoDC than AML lysates. Additional knowledge about the ability of DC to prime LAA-specific T cells is clearly warranted to further differentiate between both whole leukemic cell-derived antigen sources.
One of the key players in the MHC class II presentation pathway is CLIP and its regulators HLA-DM and HLA-DO. TLR-L regulate processing of the invariant chain (Ii) and influence the exchange of CLIP with antigenic peptides. TLR signaling is necessary to direct the internalized antigens via the classical pathway for presentation by MHC II, whereas tolerogenic DC can upregulate CLIP expression and hamper T cell signaling [35–37]. Consistent with this, CLIP expression was decreased when DC and apoptotic AML cells were co-incubated in the presence of R848 and/or CC, indicative of a putative increase in LAA presentation by the loaded MoDC.
Maturation of DC is necessary to enable migration to lymph nodes and activation of T cells. Consistent with literature, CC induced upregulation of CD86, expression of CD83, CCR7, and hence enhanced migratory capacity, the latter most likely due to the presence of prostaglandin-E2 (PGE2) in the cocktail. R848 is able to induce phenotypic maturation, but far less prominently than CC [38, 39]. In this setting, our results indicate that R848 is rather an enhancer of antigen uptake than a straightforward maturation inducer, whereas CC only promotes maturation. We hypothesized that both high uptake and maturation might be achieved by combining prolonged R848 incubation (24 h) followed by the addition of CC. However, maturation was only slightly increased compared to R848 alone. These data indicate that TLR-L antagonize the maturation efficacy of CC. Recent studies showed the inhibitory effects of TLR-L LPS and Poly(I:C) on DC by the activation of so-called suppressors of cytokine signaling (SOCS). It has been described that SOCS1, 2, and 3 are required for appropriate TLR signaling in maturing human DC via both the MyD88-dependent and -independent signaling pathway [40–43]. By first enhancing uptake with TLR-L, the DC might become resistant to cytokines due to the activation of SOCS1/2, resulting in disturbed maturation. Moreover, TLR-mediated induction of maturation-inhibitory cytokines (e.g., IL-10) might be responsible for this maturation inhibitory effect [44, 45]. We did the attempt inhibition of putatively involved down-stream signaling elements (a.o. p38 MAPK, PI3 K, STAT3) to abolish this R848-mediated block in CC-induced DC maturation, but to no avail.
PGE2, part of the cytokine cocktail, is known to impair production of IL-12p70 [46, 47]. As a consequence of reduced IL-12p70 levels, immunosuppressive cells, such as T regulatory cells might be recruited [48, 49]. Various combinations of cytokines and other DC-maturing factors, such as TLR-L, direct MoDC maturation towards a migratory and/or Th1-activating cytokine release profile [38, 50]. Recently, it was shown that the simultaneous addition of PGE2 and TLR 3/7/8 ligands resulted in higher levels of co-stimulatory molecules on DC as well as IL-12p70 production and migration capacity [38, 51]. We hypothesized that by pre-incubating MoDC with R848, followed by maturation with CC (containing PGE2), both effective IL-12p70 production, migratory capacity as well as improved AML cell uptake might be established. Addition of CC after prolonged incubation with R848 enhanced the migratory capacity, but not to the same levels as CC alone. Furthermore, IL-12p70 production was abolished as compared to CC stimulation alone. This might be explained by the longer incubation time and, thereby exhaustion of MoDC before measuring the overnight IL-12p70 production. However, in a mixed leukocyte reaction, we demonstrated that the CD8+ T cell stimulatory capacity of R848 and/or CC matured MoDC was significantly higher than that of controls, indicating that irrespective of IL-12p70 production, R848 and/or CC matured DC is able to stimulate CD8+ T cells. Since IL-12p70 is an important cytokine in Th1 and CTL activation, careful analysis of the ability to prime functional leukemia-specific T cells is needed to design optimal MoDC vaccine preparation strategies.
In conclusion, our results demonstrate that the uptake of leukemic cells by MoDC and their maturation state is differentially affected by TLR-L R848 and CC. As compared to CC alone, the sequential use of R848 and CC interfered with the effective MoDC maturation as indicated by diminished migration and T cell stimulatory capacity, and abolished IL-12p70 production. These findings do not justify incorporation of TLR7/8-L in DC-maturing cocktails for clinical development of whole-cell AML-loaded MoDC vaccines.
References
Provan D, Singer CRJ, Baglin T, Lilleyman J (2004) Acute myeloblastic leukaemia (AML). In: Provan D, Singer CRJ, Baglin T, Lilleyman J (eds) Oxford handbook of clinical haematology, 2nd edn. Oxford University Press, Oxford, UK, pp 149–157
Chamuleau ME, Souwer Y, Van Ham SM et al (2004) Class II-associated invariant chain peptide expression on myeloid leukemic blasts predicts poor clinical outcome. Cancer Res 64:5546–5550
van Luijn MM, Chamuleau ME, Thompson JA et al (2010) Class II-associated invariant chain peptide down-modulation enhances the immunogenicity of myeloid leukemic blasts resulting in increased CD4+ T-cell responses. Haematologica 95:485–493
Banchereau J, Palucka AK (2005) Dendritic cells as therapeutic vaccines against cancer. Nat Rev Immunol 5:296–306
Ridgway D (2003) The first 1000 dendritic cell vaccines. Cancer Invest 21:873–886
Rosenberg SA, Yang JC, Restifo NP (2004) Cancer immunotherapy: moving beyond current vaccines. Nat Med 10:909–915
van den Ancker W, Westers TM, Ossenkoppele GJ, van de Loosdrecht AA (2008) Back to basics: in search of the optimal dendritic cell for vaccination in AML. Leuk Res 32:1641–1643
Greiner J, Ringhoffer M, Taniguchi M et al (2004) mRNA expression of leukemia-associated antigens in patients with acute myeloid leukemia for the development of specific immunotherapies. Int J Cancer 108:704–711
Galea-Lauri J, Darling D, Mufti G, Harrison P, Farzaneh F (2002) Eliciting cytotoxic T lymphocytes against acute myeloid leukemia-derived antigens: evaluation of dendritic cell-leukemia cell hybrids and other antigen-loading strategies for dendritic cell-based vaccination. Cancer Immunol Immunother 51:299–310
Chan L, Hardwick NR, Guinn BA et al (2006) An immune edited tumour versus a tumour edited immune system: prospects for immune therapy of acute myeloid leukaemia. Cancer Immunol Immunother 55:1017–1024
Houtenbos I, Westers TM, Ossenkoppele GJ, van de Loosdrecht AA (2006) Leukemia-derived dendritic cells: towards clinical vaccination protocols in acute myeloid leukemia. Haematologica 91:348–355
de Gruijl TD, Luykx-de Bakker SA, Tillman BW et al (2002) Prolonged maturation and enhanced transduction of dendritic cells migrated from human skin explants after in situ delivery of CD40-targeted adenoviral vectors. J Immunol 169:5322–5331
Melero I, Shuford WW, Newby SA et al (1997) Monoclonal antibodies against the 4-1BB T-cell activation molecule eradicate established tumors. Nat Med 3:682–685
Castellino F, Boucher PE, Eichelberg K, Mayhew M, Rothman JE, Houghton AN, Germain RN (2000) Receptor-mediated uptake of antigen/heat shock protein complexes results in major histocompatibility complex class I antigen presentation via two distinct processing pathways. J Exp Med 191:1957–1964
Shi H, Cao T, Connolly JE et al (2006) Hyperthermia enhances CTL cross-priming. J Immunol 176:2134–2141
Vakkila J, Lotze MT (2004) Inflammation and necrosis promote tumour growth. Nat Rev Immunol 4:641–648
Obeid M, Tesniere A, Ghiringhelli F et al (2007) Calreticulin exposure dictates the immunogenicity of cancer cell death. Nat Med 13:54–61
Berwin B, Hart JP, Rice S, Gass C, Pizzo SV, Post SR, Nicchitta CV (2003) Scavenger receptor-A mediates gp96/GRP94 and calreticulin internalization by antigen-presenting cells. EMBO J 22:6127–6136
Lee JJ, Choi BH, Kang HK et al (2008) Monocyte-derived dendritic cells from HLA-matched allogeneic donors showed a greater ability to induce leukemic cell-specific T cells in comparison to leukemic cell-derived dendritic cells or monocyte-derived dendritic cells from AML patients. Leuk Res 32:1653–1660
Lee JJ, Park MS, Park JS et al (2006) Induction of leukemic-cell-specific cytotoxic T lymphocytes by autologous monocyte-derived dendritic cells presenting leukemic cell antigens. J Clin Apher 21:188–194
Decker WK, Xing D, Li S et al (2006) Double loading of dendritic cell MHC class I and MHC class II with an AML antigen repertoire enhances correlates of T-cell immunity in vitro via amplification of T-cell help. Vaccine 24:3203–3216
Gong J, Koido S, Kato Y et al (2004) Induction of anti-leukemic cytotoxic T lymphocytes by fusion of patient-derived dendritic cells with autologous myeloblasts. Leuk Res 28:1303–1312
Lee JJ, Nam CE, Nam JH et al (2004) Generation of cytotoxic donor CD8+ T cells against relapsing leukemic cells following allogeneic transplantation by stimulation with leukemic cell- or leukemic lysate pulsed donor cell-derived dendritic cells. Leuk Res 28:517–524
Spisek R, Chevallier P, Morineau N et al (2002) Induction of leukemia-specific cytotoxic response by cross-presentation of late-apoptotic leukemic blasts by autologous dendritic cells of nonleukemic origin. Cancer Res 62:2861–2868
Klammer M, Waterfall M, Samuel K, Turner ML, Roddie PH (2005) Fusion hybrids of dendritic cells and autologous myeloid blasts as a potential cellular vaccine for acute myeloid leukaemia. Br J Haematol 129:340–349
Smits EL, Ponsaerts P, Van de Velde AL et al (2007) Proinflammatory response of human leukemic cells to dsRNA transfection linked to activation of dendritic cells. Leukemia 21:1691–1699
Weck MM, Grunebach F, Werth D, Sinzger C, Bringmann A, Brossart P (2007) TLR ligands differentially affect uptake and presentation of cellular antigens. Blood 109:3890–3894
Westers TM, Houtenbos I, Schuurhuis GJ, Ossenkoppele GJ, van de Loosdrecht AA (2005) Quantification of T-cell-mediated apoptosis in heterogeneous leukemia populations using four-color multiparameter flow cytometry. Cytometry A 66:71–77
Snijders A, Hilkens CM, van der Pouw Kraan TC, Engel M, Aarden LA, Kapsenberg ML (1996) Regulation of bioactive IL-12 production in lipopolysaccharide-stimulated human monocytes is determined by the expression of the p35 subunit. J Immunol 156:1207–1212
West MA, Wallin RP, Matthews SP et al (2004) Enhanced dendritic cell antigen capture via toll-like receptor-induced actin remodeling. Science 305:1153–1157
Cella M, Scheidegger D, Palmer-Lehmann K, Lane P, Lanzavecchia A, Alber G (1996) Ligation of CD40 on dendritic cells triggers production of high levels of interleukin-12 and enhances T cell stimulatory capacity: T-T help via APC activation. J Exp Med 184:747–752
Caux C, Massacrier C, Vanbervliet B et al (1997) CD34+ hematopoietic progenitors from human cord blood differentiate along two independent dendritic cell pathways in response to GM-CSF+TNF alpha. Adv Exp Med Biol 417:21–25
Niedergang F, Chavrier P (2004) Signaling and membrane dynamics during phagocytosis: many roads lead to the phagos(R)ome. Curr Opin Cell Biol 16:422–428
Fransen JH, Hilbrands LB, Ruben J, Stoffels M, Adema GJ, Van der Vlag J, Berden JH (2009) Mouse dendritic cells matured by ingestion of apoptotic blebs induce T cells to produce interleukin-17. Arthritis Rheum 60:2304–2313
Blander JM (2007) Coupling Toll-like receptor signaling with phagocytosis: potentiation of antigen presentation. Trends Immunol 28:19–25
Blander JM, Medzhitov R (2006) On regulation of phagosome maturation and antigen presentation. Nat Immunol 7:1029–1035
Torres-Aguilar H, Guilar-Ruiz SR, Gonzalez-Perez G, Munguia R, Bajana S, Meraz-Rios MA, Sanchez-Torres C (2010) Tolerogenic dendritic cells generated with different immunosuppressive cytokines induce antigen-specific anergy and regulatory properties in memory CD4+ T cells. J Immunol 184:1765–1775
Boullart AC, Aarntzen EH, Verdijk P et al (2008) Maturation of monocyte-derived dendritic cells with Toll-like receptor 3 and 7/8 ligands combined with prostaglandin E(2) results in high interleukin-12 production and cell migration. Cancer Immunol Immunother 57:1589–1597
Assier E, Marin-Esteban V, Haziot A, Maggi E, Charron D, Mooney N (2007) TLR7/8 agonists impair monocyte-derived dendritic cell differentiation and maturation. J Leukoc Biol 81:221–228
Hu J, Winqvist O, Flores-Morales A, Wikstrom AC, Norstedt G (2009) SOCS2 influences LPS induced human monocyte-derived dendritic cell maturation. PLoS One 4:e7178
Yoshimura A, Naka T, Kubo M (2007) SOCS proteins, cytokine signalling and immune regulation. Nat Rev Immunol 7:454–465
Bartz H, Avalos NM, Baetz A, Heeg K, Dalpke AH (2006) Involvement of suppressors of cytokine signaling in toll-like receptor-mediated block of dendritic cell differentiation. Blood 108:4102–4108
Melillo JA, Song L, Bhagat G et al (2010) Dendritic cell (DC)-specific targeting reveals Stat3 as a negative regulator of DC function. J Immunol 184:2638–2645
Frick JS, Grunebach F, Autenrieth IB (2010) Immunomodulation by semi-mature dendritic cells: a novel role of Toll-like receptors and interleukin-6. Int J Med Microbiol 300:19–24
Oliveira CJ, Carvalho WA, Garcia GR, Gutierrez FR, de Miranda santos IK, Silva JS, Ferreira BR (2010) Tick saliva induces regulatory dendritic cells: MAP-kinases and Toll-like receptor-2 expression as potential targets. Vet Parasitol 167:288–297
van Helden SF, Krooshoop DJ, Broers KC, Raymakers RA, Figdor CG, van Leeuwen FN (2006) A critical role for prostaglandin E2 in podosome dissolution and induction of high-speed migration during dendritic cell maturation. J Immunol 177:1567–1574
Lebre MC, Burwell T, Vieira PL et al (2005) Differential expression of inflammatory chemokines by Th1- and Th2-cell promoting dendritic cells: a role for different mature dendritic cell populations in attracting appropriate effector cells to peripheral sites of inflammation. Immunol Cell Biol 83:525–535
Kalinski P, Vieira PL, Schuitemaker JH, de Jong EC, Kapsenberg ML (2001) Prostaglandin E(2) is a selective inducer of interleukin-12 p40 (IL-12p40) production and an inhibitor of bioactive IL-12p70 heterodimer. Blood 97:3466–3469
Luft T, Jefford M, Luetjens P et al (2002) Functionally distinct dendritic cell (DC) populations induced by physiologic stimuli: prostaglandin E(2) regulates the migratory capacity of specific DC subsets. Blood 100:1362–1372
Dowling D, Hamilton CM, O’Neill SM (2008) A comparative analysis of cytokine responses, cell surface marker expression and MAPKs in DCs matured with LPS compared with a panel of TLR ligands. Cytokine 41:254–262
Lehner M, Morhart P, Stilper A, Petermann D, Weller P, Stachel D, Holter W (2007) Efficient chemokine-dependent migration and primary and secondary IL-12 secretion by human dendritic cells stimulated through Toll-like receptors. J Immunother 30:312–322
Acknowledgments
We would like to thank Dinja Oosterhoff for technical advice and assistance. This work was supported by grant 125 from Stichting Vanderes (MM van Luijn) and Avanti-STR (JM Ruben).
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
van den Ancker, W., van Luijn, M.M., Ruben, J.M. et al. Targeting Toll-like receptor 7/8 enhances uptake of apoptotic leukemic cells by monocyte-derived dendritic cells but interferes with subsequent cytokine-induced maturation. Cancer Immunol Immunother 60, 37–47 (2011). https://doi.org/10.1007/s00262-010-0917-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00262-010-0917-y