
Abstract
The oleaginous yeast Yarrowia lipolytica has become a recognized system for expression/secretion of heterologous proteins. This non-conventional yeast is currently being developed as a workhorse for biotechnology by several research groups throughout the world, especially for single-cell oil production, whole cell bioconversion and upgrading of industrial wastes. This mini-review presents established tools for protein expression in Y. lipolytica and highlights novel developments in the areas of promoter design, surface display, and host strain or metabolic pathway engineering. An overview of the industrial and commercial biotechnological applications of Y. lipolytica is also presented.
Similar content being viewed by others
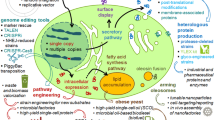

Introduction
Yarrowia lipolytica is a dimorphic oleaginous yeast, belonging to the Ascomycota phylum (subphylum: Saccharomycotina, previously called Hemiascomycetes), that exhibits remarkable lipolytic and proteolytic activities. Its natural occurrence appears rather ubiquitous, including soils, marine waters, mycorrhizae, and a variety of foods (particularly dairy products, including numerous cheeses, and meat), as recently reviewed by Groenewald et al. (2014) and Zinjarde (2014). Since more than 50 years, this non-conventional yeast has promoted interest for industrial applications, firstly oriented toward the production of metabolites with commercial value and secondly aimed at producing heterologous proteins. The fact that Y. lipolytica has been frequently isolated from environments heavily polluted by hydrocarbons has also prompted interest for its use in bioremediation of oil-contaminated soils and waters or in upgrading oily wastes (Bankar et al. 2009). On account of its peculiar characteristics, Y. lipolytica has also become a model for basic research in the study of dimorphism, secretion, salt tolerance, mitochondrial complex, peroxisomes, or lipid accumulation (as reviewed by Nicaud 2012). More recently, the combination of increased knowledge of its metabolism, sequencing of its genome, and development of more efficient genetic tools offered new perspectives in metabolic pathway engineering, allowing using this yeast as a cell factory. In the rapidly developing domain of Y. lipolytica research and applications, publications abound but the most extensive reviews are published in books (such as the two recent volumes from Springer’s Microbiology Monographs series— Barth 2013a, b), which limits their impact. This mini-review, besides providing a concise guide for the beginner, will attempt to be more synthetic than exhaustive and will focus on the most recent achievements in the domain of heterologous protein expression in Y. lipolytica, especially concerning the engineering of its metabolic pathways for various applications.
A brief history of Y. lipolytica use and safety
The industrial use of Y. lipolytica was pioneered in the 1950s by British Petroleum (UK), during the search for microorganisms growing on cheap substrates (especially on crude oil) and able to produce, on a large-scale, high-quality protein products for nutritional supply. Wild-type strains have been used to produce single-cell proteins (SCP) for livestock feeding until the 1970 oil crisis. Since then, Y. lipolytica has also been applied to the production of different organic acids, especially citric acid (Pfizer Inc. and ADM, USA), and of erythritol (Baolingbao Biology Co., China). More recently, the Polish company Skotan SA started to commercialize Y. lipolytica biomass, for use as fodder yeast in Europe, and to develop prebiotic and probiotic applications of this yeast for food industry. In the environmental domain, Artechno (Belgium) produces a starter for bioremediation of oily wastewaters, composed of freeze-dried wild-type Y. lipolytica cells and extracellular lipase. A lot of wild-type Y. lipolytica strains have been isolated from hydrocarbon-contaminated soils or waters, which makes them interesting candidates for bioremediation applications. Such studies are abundantly represented in the scientific literature, at the laboratory scale, but, to our knowledge, only one in situ field study has been conducted, for the bioremediation of a petroleum-contaminated soil (Bankar et al. 2009). Although the use of genetically modified microorganisms (GMM) could raise safety concerns, Y. lipolytica appears as one of the non-conventional yeasts that presents the best guarantees in this regard. With a longstanding history of safe use, Y. lipolytica is considered as a biosafety class 1 microorganism, namely a microorganism which is not known to cause disease in healthy adult humans (Groenewald et al. 2014). The industrial applications cited above have obtained a generally regarded as safe (GRAS) status from the Food and Drug Administration (FDA, USA). Consequently, Y. lipolytica is listed by many countries as a microorganism acceptable for constructing GMM as long as its large-scale production facilities comply with the good industrial large-scale practice (GILSP) level of physical containment (Groenewald et al. 2014). Since the 2000s, the use of genetically modified Y. lipolytica strains has developed and several products have been (or are on the edge to be) marketed. DuPont (USA) has extensively engineered Y. lipolytica metabolic pathways (Xue et al. 2013) for the production of eicosapentaenoic acid (EPA)-rich products: EPA-rich Y. lipolytica biomass is currently used as an ω-3 supplement in fish feed (harmoniously raised salmon VerlassoTM), and an EPA-rich single-cell oil (SCO), for use as dietary supplement, has been marketed in 2010 under the name of New HarvestTM. However, its commercialization has been recently discontinued (personal communication from DuPont) despite the obtention of a GRAS notice. DuPont and Microbia (USA—now a subsidiary from the Dutch-based multinational company DSM) have independently developed Y. lipolytica as a production host for carotenoids. Extensive data on the safety of β-carotene produced from GM Y. lipolytica has been recently disclosed by Microbia (Grenfell-Lee et al. 2014). Engineered Y. lipolytica strains are also used by two European companies for the production of recombinant enzymes, for enzyme replacement therapies (ERTs): human lysosomal enzymes, for treatment of lysosomal storage diseases (Oxyrane, Belgium—Tiels et al. 2012) and an overexpressed homologous lipase (YlLip2p) (Pignède et al. 2000) for treatment of exocrine pancreatic insufficiency (Mayoly Spindler, France). These ERTs are currently evaluated in clinical trials. In addition, engineered Y. lipolytica strains are employed in protein manufacturing platforms by Protéus (France) and Oxyrane (UK), both making use of technologies developed at Institut National de la Recherche Agronomique (INRA, France). More details on Y. lipolytica industrial use and safety assessment could be found in recent reviews by Groenewald et al. (2014) and Sibirny et al. (2014); the subject of bioremediation has been extensively treated by Bankar et al. (2009), and food-related applications have been recently reviewed by Zinjarde (2014).
Tools for heterologous protein expression, secretion, and surface display
The use of Y. lipolytica as a host for heterologous expression was initiated, 30 years ago, by the almost simultaneous development of transformation methods by Pfizer Inc. (USA) and a French INRA team (Davidow and DeZeeuw 1984; Gaillardin et al. 1984). Since then, more than 130 proteins, from more than 80 species, have been successfully produced in this yeast (Madzak and Beckerich 2013 and more recent survey of scientific literature). The use of synthetic genes, although relatively recent, represents 22 % of these cases. When the native genes were used, their phylogenic origins were diverse, with Ascomycota representing 27 % of the cases, mammals 20 % (human genes alone: 15 %), and Proteobacteria 14 %. The remaining 39 % of cases include viruses, Firmicutes, Actinobacteria, Basidiomycota, Zygomycota, Heterokonta, protists, plants (monocotyledons or dicotyledons), insects, and a mollusc. An overview of the possibilities offered by the Y. lipolytica expression system is schematized in the Fig. 1.

Overview of possibilities for heterologous gene expression in Y. lipolytica expression vectors harboring one (or more) expression cassette(s) composed of a promoter (P), the open reading frame (ORF) of the gene of interest and a terminator (T), together with a selection marker (M), and bordered by sequences for integration, are linearized prior to transformation. Alternatively, a yeast expression cassette can be separated from the bacterial vector backbone and purified for transformation. Sequences for integration generally consist in 5′ and 3′ homologous regions (5′H and 3′H). Zeta sequences from Ylt1 retrotransposon can be used for that purpose in Ylt1-bearing strains or can promote random ectopic integration by non-homologous recombination in strains devoid of this retrotransposon. The additional use of proper signal sequence(s) will direct the recombinant protein to the secretory pathway (secretion signal) and, possibly, to surface display via fusion to anchoring domains (open triangle), notably homologous or heterologous glycosylphosphatidylinositol (GPI) anchor but also protein internal repeat (Pir) or chitin binding module (CBM). Other possibilities include targeting to cellular microbodies such as peroxysomes (peroxisomal targeting signal (PTS)) or oléosomes (fusion to heterologous oleosin domain: black triangle). Some glyco-engineered strains allow producing recombinant glycoproteins with human-compatible N-linked oligosaccharides for therapeutic applications. Others are able to produce heterologous lysosomal enzymes with high levels of mannose-6-phosphate which will target them to lysosomes in patient cells. Details and references are given in the text
Strains
A few wild-type Y. lipolytica isolates have been selected, on the basis on noticeable properties, and engineered for peculiar applications. Thus, several marine Y. lipolytica strains from the Ocean University of China (OUC, Qindao) library have been recently engineered for use as citric acid, SCO, or SCP producers (respectively: Liu et al. 2010; Zhao et al. 2010; Cui et al. 2011). Similarly, the wild-type WSH-Z06 strain, a natural α-ketoglutaric acid (KGA) overproducer, has been engineered in order to optimize KGA production (Yin et al. 2012; Zhou et al. 2012; Guo et al. 2014) and the German wild-type isolate, H222 (MatA), or a KGA-overproducing derivative, were engineered for various organic acid production (Holz et al. 2009, 2011; Otto et al. 2012; Yovkova et al. 2014; Jost et al. 2015). Besides these examples, most heterologous expression works have used only a limited number of inbred lines. An extensive list of these Y. lipolytica host strains can be found in Madzak et al. (2004), but the most used are E129 and the Po1 series of strains (Po1d, f, g and h), as more recently reviewed by Madzak and Beckerich (2013). Like the E150 strain (MatB, his1, leu2-270, ura3-302, xpr2-322), whose genome has been fully sequenced (Dujon et al. 2004), the E129 strain (MatA, lys11-23, leu2-270, ura3-302, xpr2-322) is issued from a mating followed by multiple back-crosses between derivatives of a French wild-type isolate, W29 (MatA), and an American one, YB423-12 (MatB), as reviewed by Barth and Gaillardin (1996). The Po1 series of strains are derived only from W29 and, consequently, do not bear the Ylt1 retrotransposon, which was absent from this French wild-type isolate. Po1d (MatA, leu2-270, ura3-302, xpr2-322) presents interesting features for heterologous protein production: high secretion levels, deletion of alkaline extracellular protease (AEP, encoded by XPR2 gene), non-reverting Leu− and Ura− auxotrophies, and production of recombinant invertase (ura3-302 allele corresponds to the disruption of URA3 by the SUC2 gene from Saccharomyces cerevisiae under the control of XPR2 promoter) allowing to use sucrose or molasses as carbon source (Nicaud et al. 1989). A series of derivatives have been further adapted for heterologous protein production (Madzak et al. 2000 and 2004): Po1f, Po1g, and Po1h were further deleted for the acid extracellular protease (axp1-2 allele). Po1g was also fitted with an integrated pBR322 docking platform (ura3-302::URA3:pBR322), for further integration of pBR322-based vectors, and retains only Leu− auxotrophy. Po1h was modified, using genic LEU2 conversion, to retain only Ura− auxotrophy. These strains can be ordered from INRA’s CIRM-Levures Yeasts Library (http://www7.inra.fr/cirmlevures/page.php?page=home&lang=en). A draft genome sequence of Po1f has recently been published (Liu and Alper 2014). Another derivative of Po1d strain has been engineered for high-throughput applications, especially in the domain of lipid metabolism: JMY1212 (MatA, ura3-302, leu2-270::LEU2:zeta, xpr2-322, Δlip2, Δlip7, Δlip8), has the main lipases deleted, and was fitted with an integrated zeta (Ylt1 retrotransposon long terminal repeat (LTR) docking platform, for further integration of zeta-based vectors; it retains only Ura− auxotrophy. JMY1212 is used as recipient strain in the high-throughput system developed by INSA, CNRS, and INRA (France), for screening new biocatalysts through expression cloning and for improving selected enzymes through directed enzyme evolution (Bordes et al. 2007). At last, the ScSUC2 expression cassette has been recently modified in a new Po1d derivative, JMY2593, in order to provide a Y. lipolytica strain with improved growth on sucrose for industrial applications (Lazar et al. 2013).
Vectors
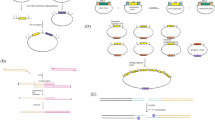
Two types of shuttle vectors can be used for transforming yeast cells: episomal (replicative) plasmids and vectors designed for integration into the yeast genome, for which linearization is required to direct targeting at a precise locus. No Y. lipolytica strain bears any natural episome, but replicative plasmids using chromosomal autonomously replicating sequence/centromer (ARS/CEN) replication origins have been designed (Fournier et al. 1993; Matsuoka et al. 1993). Such vectors are useful when transient expression is required, e.g., for the Cre-lox gene disruption method (Fickers et al. 2003), but their use for heterologous expression is limited by their low copy number (average of one to three plasmids per cell under selective conditions) and their high loss frequency (Vernis et al. 1997). Liu et al. (2014) have recently designed an improved Y. lipolytica ARS/CEN plasmid by fusing a promoter upstream of the centromeric region, thus increasing the relative copy number by 80 % and providing a starting point for more potent replicative plasmids in Y. lipolytica.
Nevertheless, the use of integrative vectors remains the preferred strategy for heterologous expression and/or genetic engineering in Y. lipolytica. As schematized in the Fig. 1, the integration of such vectors can be targeted to a genomic locus or docking platform, by linearization into a corresponding homologous region. However, in contrast to S. cerevisiae (but like other yeasts), Y. lipolytica uses mainly the non-homologous end-joining (NHEJ) recombination pathway and not the homologous recombination (HR) pathway for DNA double-strand break (DSB) repair (Richard et al. 2005). In consequence, homologous integration of exogenous DNA by a single crossing-over into Y. lipolytica genome can occur at acceptable rates (over 80 %) only in presence of large enough homologous 5′- and 3′-flanking regions, in the range of 0.5 to 1 kb (Barth and Gaillardin 1996; Fickers et al. 2003). In order to palliate this problem, a Y. lipolytica strain deleted for KU70 gene, responsible for DSB repair in the NHEJ recombination pathway, was recently developed at INRA (France) (Verbeke et al. 2013). The ku70 deletion significantly reduced transformation efficiency but promoted efficient HR with short length flanking regions. Targeted gene conversion, by double crossing-over, was obtained at a frequency of 43 % with 5′- and 3′-flanking homologous regions as short as 50 bp (Verbeke et al. 2013).
The requirement of large homologous flanking regions for efficient HR in Y. lipolytica has however not impaired the development of effective integrative expression vectors. The use of an integrated docking platform, usually a bacterial plasmid backbone (such as pBR322), allows targeting a unique copy at a precisely known genomic site (Madzak et al. 2000 and 2004; Bordes et al. 2007). This system is particularly adapted to the genetic engineering of proteins, since the effect of mutations on recombinant enzyme activity can be compared directly on the transformants. The transformation efficiency can be very high, in the range of 105 colonies per microgram of vector DNA for pBR322-based vectors transforming Po1g recipient strain (Madzak and Beckerich 2013). This efficient and easy-to-use expression/secretion system is commercialized under an INRA license: YLEX™ Expression Kit can be purchased from Yeastern Biotech Co. (Taiwan) or Gentaur (Belgium). In order to avoid the presence of bacterial DNA (especially antibiotic resistance genes) into recombinant yeast strains devoted to industrial applications, some autocloning vectors have been developed: The bacterial backbone can be excised, and only a yeast cassette (composed of the expression cassette and selection marker, bordered by sequences for integration) is used for transformation (Pignède et al. 2000; Nicaud et al. 2002).
Vectors carrying two or three expression cassettes, for co-expression of several heterologous genes, have been employed by different research groups (Chuang et al. 2010; Celińska and Grajek 2013). The possibility to increase and/or tune gene expression is crucial for both heterologous protein production and metabolic engineering. To this end, several research teams have developed various strategies, notably endogenous promoter characterization (Müller et al. 1998; Juretzek et al. 2000), hybrid promoter engineering (Madzak et al. 1995 and 2000; Blazeck et al. 2011 and 2013a), and multicopy integration (Le Dall et al. 1994; Juretzek et al. 2001).
Promoters
The characteristics of the major promoters used for heterologous expression in Y. lipolytica have been extensively described elsewhere (Madzak and Beckerich 2013) and are resumed in the Table 1. If the XPR2 promoter (pXPR2) has been historically important during the development of Y. lipolytica expression system, its complex regulation and the cost of using peptones for its induction urged the search for new promoters, more fit for industrial constraints. Other major strong endogenous promoters are the constitutive pTEF1 and some inducible promoters from the fatty acids pathway: pPOX2 and pICL1. The functional dissection of pXPR2 allowed to use one of its upstream activating sequences (UAS), poorly affected by cultivation conditions (Madzak et al. 1999), to design the recombinant promoter hp4d (Madzak et al. 1995 and 2000). This recombinant promoter, able to drive a strong expression in various culture media, exhibits a growth phase-dependent expression pattern. Its expression increases at the beginning of the stationary phase, a characteristic particularly interesting for heterologous production since it allows a partial dissociation of the growth and expression phases. Due to these interesting characteristics, the use of hp4d has rapidly developed, especially since the YLEX kit for heterologous expression using this patented promoter has been commercialized by Yeastern Biotech Co. (Taiwan). This recombinant promoter has been employed up to now in more than half of the published examples of heterologous protein production in Y. lipolytica (Madzak and Beckerich 2013 and more recent survey of scientific literature). More recently, an important improvement in the design of strong and tunable Y. lipolytica promoters has been carried out at UT Austin (USA), based on an extension of the concept of the recombinant hp4d promoter. Blazeck et al. (2011) constructed a large set of hybrid promoters, carrying various copy numbers (from 1 to 32) of an UAS inserted upstream of a minimal or reduced endogenous promoter (Table 1). The strongest of these engineered promoters exhibited an unprecedented efficiency, 8-fold higher than that of any known endogenous Y. lipolytica promoter (Blazeck et al. 2011). In addition to providing a series of new hybrid promoters with tunable strength, this work proposed a general approach for improving selected endogenous promoters. Moreover, it also suggested that endogenous Y. lipolytica promoters were enhancer-limited and that this limitation could be alleviated through UAS addition, whereas transcription factor availability was not a limiting factor.
Selection markers
Either dominant or auxotrophic markers are available for selection in Y. lipolytica. Although naturally resistant to most antibiotics, Y. lipolytica is sensitive to hygromycin B and to antibiotics of the bleomycin/phleomycin group. Heterologous genes conferring resistance to these antibiotics can be used as dominant selection markers in Y. lipolytica (Gaillardin and Ribet 1987; Fickers et al. 2003). In contrast, heterologous expression of S. cerevisiae SUC2 gene, which was developed at first as a dominant marker (Nicaud et al. 1989), is impaired by residual growth on sucrose plate impurities (Barth and Gaillardin 1996) and is used preferably for recipient strain improvement (in E129 and Po1d derivatives) than for selection. A number of auxotrophic marker genes have been used for selection in Y. lipolytica, but the most widely employed are LEU2 and URA3, for which non-leaky and non-reverting mutations are available: leu2-270 and ura3-302 alleles, present in E129 and in some Po1d derivatives, contain large deletions into their coding sequences (Nicaud et al. 1989; Barth and Gaillardin 1996). In order to increase heterologous protein production, some promoter-defective versions of URA3 marker have been designed (Le Dall et al. 1994). For example, vectors carrying the ura3d4 allele are unable to correct ura3 auxotrophy when in single copy but can restore growth on selective medium after an amplification process leading to multiple integrations (Le Dall et al. 1994). Similar promoter-defective versions of LEU2 marker are currently tested at INRA (Madzak, unpublished results).
Multiple integrations
Integrating multiple copies of a vector (or of an expression cassette) into the genome remains the preferred strategy for increasing heterologous gene copy number in Y. lipolytica. Selection of such events requires defective markers, especially the ura3d4 allele which is employed in most multicopy vectors (Juretzek et al. 2001; Nicaud et al. 2002). This defective marker allows obtaining several tens of copies integrated at unique or multiple sites, especially when targeting the ribosomal DNA (rDNA) G unit or the zeta sequences (LTRs from Ylt1). However, such high copy number integrants were shown to be unstable under cultivation conditions inducing heterologous expression, their number of copies eventually stabilizing around ten (Juretzek et al. 2001). This low stability seemed to be linked to the fact that multiple integrations occurred almost always in tandem copies, at only one or two sites (Le Dall et al. 1994).
Due to Y. lipolytica efficient NHEJ recombination pathway, the zeta-based autocloning vectors are also able to transform effectively strains devoid of Ylt1 retrotransposon, such as W29 and its derivatives (like the Po1 series of strains), by random ectopic integration. When using a promoter-defective selection marker, this non-homologous integration process generates multiple copies which are more dispersed within the genome than those targeted by homology, ensuring a better stability of high copy number integrants (Nicaud et al. 1998). Such non-homologously transformed multicopy strains have allowed producing high yields of numerous heterologous proteins, as reviewed by Madzak and Beckerich (2013). However, they remain difficult to construct, due to a very low efficiency of the transformation step. Moreover, as the transformants can present alterations and/or large differences in production levels, both linked to random integration loci, a careful checking is required. High copy number strains were also reported to suffer from a productivity loss linked to metabolic burden (Roth et al. 2009), and their long-term stability appeared unable to comply with the high standard of GMP guidelines (Nicaud et al. 2008).
In order to overcome these limitations, Nicaud et al. (2008) have developed a more controlled amplification strategy: A genetically engineered strain, deleted for genes of major secreted homologous proteins (thus alleviating metabolic burden), presents several loci selected for the targeting of integrative vectors carrying different auxotrophic or dominant markers. Up to ten stable integrations can be performed sequentially, which can be checked easily since each step confers a new phenotype, detectable on plate, to the transformed strain. Although patented as a method for the obtention of stable multicopy transformants (Nicaud et al. 2008), this strategy appears however laborious compared to the recently developed alternative strategy of promoter tuning from Blazeck et al. (2011), if one considers only the purpose of increasing expression. In contrast, it seems much more interesting in the domain of genetic engineering of complex metabolic pathways, for which the expression of several different genes is required.
However, sequential gene integration methods remain laborious and time-consuming. The DNA assembler method, previously used only on a few other yeast species, was very recently proposed by Gao et al. (2014) as an effective alternative strategy for Y. lipolytica genetic engineering. This method, which allows assembling multiple fragments in a single step via in vivo homologous recombination, was successfully used to integrate an entire β-carotene biosynthesis pathway (a selection marker and three expression cassettes, with a total size near 11 kb) at the rDNA locus of Y. lipolytica. Despite the fact that efficient homologous recombination in Y. lipolytica requires large 5′- and 3′-flanking regions (0.5 to 1 kb), the total efficiency of in vivo one-step assembly of the four DNA fragments was around 20 % with overlaps between two successive cassettes of only 65 bp (Gao et al. 2014). The choice of the β-carotene biosynthesis pathway for making a proof of concept of the DNA assembler method in Y. lipolytica allowed an easy visual screening of the transformants, since the colonies bearing a functional set of genes turned orange/red. Gao et al. (2014) suggested that the overall efficiency of this method for one-step multiple gene transformation could be higher in strains deleted for KU70 gene, impaired in NHEJ mechanism (Verbeke et al. 2013). This time-saving DNA assembler method is expected to greatly facilitate metabolic engineering in Y. lipolytica.
Signals for secretion, targeting, or display
In order to direct a heterologous protein to the secretory pathway of Y. lipolytica, with the purpose to obtain its release into the cultivation medium, its targeting to cellular microbodies, or its display on the cell surface, a translational fusion with a proper signal is required. The characteristics of the major signals used are resumed in the Table 2. Efficient secretion of heterologous proteins, involving mainly the co-translational pathway (Boisramé et al. 1998), is a key strength of the Y. lipolytica expression system: A signal sequence is fused upstream from the mature sequence of the protein of interest, in order to target the nascent polypeptide to the secretion pathway. Most secretion signals used are derived from Y. lipolytica XPR2 or LIP2 genes, but a few heterologous secreted proteins from fungi or plants have also been produced successfully using their own secretion signal (reviewed by Madzak and Beckerich 2013).
Although of more restricted use, the possibility to target heterologous proteins to cellular microbodies, such as peroxisomes or oleosomes, constitutes an interesting tool for peculiar high value applications. Targeting of heterologous proteins to Y. lipolytica peroxisomes can be performed via fusion of a peroxisomal targeting signal (PTS) domain to their C terminus. The tripeptide AKI, considered as a consensus PTS for the related Yarrowia and Candida genera, has been used for directing a bacterial polyhydroxyalkanoate synthase to Y. lipolytica peroxisomes (Haddouche et al. 2010), and the tripeptide SKL, closer to the S. cerevisiae consensus PTS, has been used for targeting green fluorescent protein (GFP) (Xue et al. 2013). Recently, Y. lipolytica has been developed as a platform for producing tunable arming oleosomes (Han et al. 2013) that could serve multiple purposes, like cell targeting and reporting activities (targeted drug delivery, pathogen detection) or self-assembly of functionalized nanostructures (nanofactories). Several heterologous proteins have been targeted to Y. lipolytica oleosomes and anchored on their surface via their fusion to the C-terminal domain of a plant oleosin (Han et al. 2013), a structural protein embedded in the monolayer phospholipidic membrane of plant oleosomes. The engineered oleosomes were purified, using floating centrifugation, as highly stable nanoparticles with a mean diameter of 200 to 300 nm (Han et al. 2013). The feasibility of co-displaying multiple proteins on Y. lipolytica oleosomes was demonstrated by using the high-affinity binding properties of the cellulosome scaffolding system from Clostridium cellulolyticum: Oleosomes displaying a cohesin domain were able to interact with different fusion proteins bearing a dockerin domain, thus forming multi-functional nanoparticles able to target specific cells and to provide new catalytic functions at their surface (Han et al. 2013).
The relatively recent development of several surface display systems now allows using Y. lipolytica as arming yeast, which increases its possibilities for various applications such as immobilized biocatalysis or bioconversion, biosensor and live vaccine development, or high-throughput screening of new enzymatic activities. The first example of surface displaying a heterologous protein in Y. lipolytica used its fusion to the glycosylphosphatidylinositol (GPI) anchor domain of a cell wall protein (Yue et al. 2008). In this GPI-anchoring system, the C-terminal domain of YlCWP1p served as a signal for creating a covalent bond between the fusion protein and cell wall β-1,6 glucans, via a GPI anchor (post-translational modification including a phosphoethanolamine linker, a glycan core, and a phospholipid tail). Several heterologous proteins have been displayed successfully using this system (reviewed by Madzak and Beckerich 2013), and GPI anchor domains from other Y. lipolytica cell wall proteins have also been used for the same purpose (Table 2) (Yuzbasheva et al. 2010; Moon et al. 2013). Besides GPI anchor domains, three alternative systems have been used for surface displaying heterologous proteins in Y. lipolytica (Table 2): (i) fusion with homologous (Yuzbasheva et al. 2010) or heterologous (Yang et al. 2009) flocculation domains which bind non-covalently to cell surface mannan chains, (ii) fusion with a protein internal repeat (Pir) domain which binds covalently to cell wall β-1,3 glucans and, via disulfide bonds, to structural proteins, and (iii) fusion with a chitin binding module (CBM) which binds non-covalently to cell surface via interactions with chitin from cell wall (Duquesne et al. 2014).
Heterologous protein production and pathway engineering
The use of Y. lipolytica for obtaining industrially valuable compounds has developed in two interconnected directions: as a host for production of heterologous proteins and as an engineered whole cell biocatalyst. Using Y. lipolytica as a host sometimes requires modifying its metabolism, notably developing humanized glyco-engineered strains. Using Y. lipolytica as a microbial factory often requires introducing new enzymatic activities by expressing foreign proteins. A genome-scale metabolic model for Y. lipolytica has been recently produced by a consortium of French research teams (Loira et al. 2012), using a scaffold derived from the well-studied but phylogenetically distant yeast S. cerevisiae. This metabolic model constitutes a powerful tool that will be helpful for metabolic engineering and high-throughput data analysis. As production of heterologous proteins in Y. lipolytica has been extensively reviewed recently (Madzak and Beckerich 2013), this mini-review will focus on the production of enzymes of economic significance (notably lipases, proteases, amylases, mannanases, and laccases) and on recent advances in genetic engineering of metabolic pathways. Among paramount data, a few selected examples of recombinant proteins or metabolic products production in Y. lipolytica are shown in Table 3, especially those highlighting successful effects of expression amplification, gene design, optimized cultivation, or metabolic engineering.
Heterologous production of industrially relevant enzymes
Lipases are a subclass of esterases that catalyzes the hydrolysis of lipids. They have various applications in food and detergent industries, in bioremediation (wastewater treatment), or as biocatalysts for organic chemistry and biodiesel production. They can also be used in exocrine pancreatic insufficiency treatments, like the ERT developed by Mayoly Spindler (France) with a recombinant strain overexpressing homologous YlLip2p (Pignède et al. 2000). An arming Y. lipolytica strain displaying YlLip2p has also been designed for use as whole-cell biocatalyst (Yuzbasheva et al. 2011). Extracellular lipases from Rhizopus genus are noticeable industrial enzymes, in which properties vary depending on post-translational modifications. One of these fungal lipases has been displayed on Y. lipolytica cells, for bioremediation (Song et al. 2011). Another was secreted by Y. lipolytica as a post-translationally modified form exhibiting increased thermostability (Yuzbashev et al. 2012). The Y. lipolytica high-throughput expression platform (INRA-CNRS-INSA, France—Bordes et al. 2007) has been applied to the engineering of Candida antarctica lipase B, one of the main enzymes for industrial biocatalysis, and allowed identifying new mutants with higher catalytic efficiencies (Emond et al. 2010).
Proteases include different classes of enzymes able to hydrolyze peptide bonds by various catalytic mechanisms. They have numerous applications, mainly in food and detergent industries, but also in wastewater treatment and in the medical domain. Two fungal alkaline proteases, with potential applications in the production of bioactive peptides, have been either secreted by Y. lipolytica (Ni et al. 2008) or displayed on its cell surface (Ni et al. 2009). A fungal acid protease has also been either secreted by Y. lipolytica (Yu et al. 2010a) or displayed on its cell surface (Yu et al. 2010b), for potential use as rennet substitute in cheese industry.
Alpha-amylases are glycoside hydrolases of major importance for starch processing applications. Y. lipolytica has been used as production host for two industrially relevant α-amylases, from a mesothermophilic bacteria (Yang et al. 2010) and from an insect pest of rice (Celińska et al. 2014).
Beta-mannanases are hemicellulases able to hydrolyze 1,4-β-D-mannosidic linkages from the main chains of mannan and derivatives. As such, they play a significant role in the bioconversion of lignocellulosic biomass and can be applied to pulp bleaching. They are also employed in food and feed industries, in detergent industry and in oil drilling (Yang et al. 2009). A fungal β-mannanase has been secreted in Y. lipolytica and its production optimized from a multicopy strain (Roth et al. 2009). An arming Y. lipolytica strain displaying a bacterial β-mannanase has been constructed and proposed for use as whole-cell biocatalyst in industrial processes (Yang et al. 2009).
Laccases are multi-copper oxidoreductases able to oxidize a large range of phenols and arylamines. Fungal laccases are involved in lignin catabolism and can be applied to the degradation of pesticides, dyes, and polycyclic aromatic hydrocarbons (PAHs), which makes them environmentally relevant enzymes (Mougin et al. 2003). Three laccases from Basidiomycota have been secreted in Y. lipolytica (Madzak et al. 2005; Jolivalt et al. 2005; Theerachat et al. 2012) and, for two of them, improved by genetic engineering (Madzak et al. 2006; Theerachat et al. 2012). These enzymes could find applications in the biodegradation of xenobiotics (Galli et al. 2011) and/or in the development of oxygen cathodes in microbial biofuel cells (Sané et al. 2014).
Design of glyco-engineered strains for therapeutic applications
Using yeasts as host for producing recombinant therapeutic proteins has been at first impaired by the differences existing between their N-glycosylation pathway and the mammalian one. Namely, yeasts modify glycoproteins with high mannose-type N-glycans, which often reduces in vivo protein half-life and can also be immunogenic in humans and other mammals. This prompted the development of glyco-engineered “humanized” strains in the few yeast species commonly used for heterologous production, as recently reviewed by Kim et al. (2014). Exploration of the genetics of N-glycosylation in Y. lipolytica has already allowed some research groups from South Korea and Belgium to design and patent strains producing more human-compatible glycoproteins, which will serve as starting hosts for further glyco-engineering (Kang et al. 2007; Callewaert et al. 2007). A Y. lipolytica strain deleted for α-1,6-mannosyltransferase YlOCH1p was shown to synthesize only the core oligosaccharide Man8GlcNAc2 (Song et al. 2007), when wild-type strains synthesize oligosaccharides with heterogeneous sizes (up to Man12GlcNAc2). After demonstrating that YlMPO1p was necessary for the mannosylphosphorylation of N-linked oligosaccharides, the same consortium of South Korean research groups constructed a Δoch1 Δmpo1 double mutant strain lacking yeast-specific hypermannosylation and mannosyl phosphorylation (Park et al. 2011). In order to develop a glyco-engineered Y. lipolytica strain with mannose trimming activity, this Δoch1 Δmpo1 strain was recently engineered further by surface display of a fungal α-1,2-mannosidase. This arming strain was able to convert Man8GlcNAc2 to Man5GlcNAc2, not only in activity tests, but also on a large portion of its own cell wall mannoproteins (Moon et al. 2013), demonstrating its efficiency not only for in vitro trimming but also for in vivo trimming of N-glycans during the secretion process.
A consortium of Belgian research groups, including Oxyrane (Belgium), also designed Y. lipolytica engineered strains producing glycoproteins homogenously carrying either Man8GlcNAc2 or Man5GlcNAc2. They constructed a strain lacking both yeast-specific Golgi α-1,6-mannosyltransferases YlOch1p and YlMnn9p, which produced homogeneous Man8GlcNAc2 N-glycans. This strain was modified further by heterologous expression of an ER-targeted fungal α-1,2-mannosidase, in order to be able to produce homogeneous Man5GlcNAc2 residues (De Pourcq et al. 2012a).
The same Belgian consortium has also engineered N-glycan biosynthesis in Y. lipolytica for production of N-glycoproteins homogenously carrying a Man3GlcNAc2 core. The involved steps were the following (De Pourcq et al. 2012b): (i) disruption of YlAlg3 α-1,3-mannosyltransferase, (ii) overexpression of YlAlg6 glucosyltransferase, (iii) heterologous overexpression of a fungal glucosidase, and (iv) overexpression of the same ER-targeted fungal α-1,2-mannosidase. Interestingly, the resulting Man3GlcNAc2 core produced is common to all mammalian N-glycan structures and can be modified further in vitro to yield any of the complex type N-glycans, providing that adequate glycosyltranferases and sugar-nucleotide donors were used (De Pourcq et al. 2012b). These new expression platforms will promote Y. lipolytica as a competitive heterologous host able to produce humanized N-linked oligosaccharides compatible with therapeutic applications.
The same consortium of Belgian research groups, including Oxyrane (Belgium), has also developed a glyco-engineered Y. lipolytica strain expressing a bacterial glycosidase, in which the level of mannose-6-phosphate modification of glycoproteins was increased. This strain was designed for producing recombinant human lysosomal enzymes for ERTs of lysosomal storage diseases. It was applied to the production of a recombinant Pompe disease enzyme, an α-glucosidase enriched in mannose-6-phosphate, which enabled its efficient targeting to the lysosomes of diseased cells via interaction with specific receptors (Tiels et al. 2012). This technology has high potential for the production of cost effective and more efficient enzymes for replacement therapy of lysosomal diseases.
Use of recombinant Y. lipolytica strains as whole-cell biocatalysts
Epoxide hydrolase-expressing Y. lipolytica strains
Epoxide hydrolases (EHs) are able to catalyze the hydrolysis of epoxides into their corresponding diols. Fungal EHs are particularly enantioselective in this process, which makes them interesting as catalysts in the kinetic resolution of various epoxides for applications in fine chemicals and pharmaceutical industries (Labuschagné 2005). Several heterologous EH genes of various phylogenetic origins (from a bacterium, fungi, a plant, an insect, or a mammal) have been expressed intracellularly in Y. lipolytica by a consortium of South African research groups (Labuschagné 2005; Labuschagné and Albertyn 2007; Maharajh et al. 2008a, b). For recombinant strains producing fungal EHs from Rhodotorula genus, both activities and enantiomeric selectivities in the whole-cell biotransformation of a racemic substrate were higher than those observed with the native fungi (Labuschagné and Albertyn 2007; Maharajh et al. 2008a). The process was optimized using a multicopy strain and an exponential feed model of fermentation (Maharajh et al. 2008a, b). The spin-off company Oxyrane (UK) has constructed a portfolio of EH-engineered Y. lipolytica strains that constitutes a biotransformation platform for whole-cell enantiomeric bioconversion of racemic epoxides (Pienaar et al. 2008).
Cytochrome P450-expressing Y. lipolytica strains
Eukaryotic cytochromes P450 (CYP450s) are a superfamily of membrane-bound, haem-containing mono-oxygenases involved in the metabolism of drugs, steroid hormones, and fatty acids. Their heterologous expression is both of fundamental (investigation of their role in xenobiotics and drug degradation pathways) and of economical interest (use in bioconversion for synthesis of hydroxylated biomolecules in biotechnological or therapeutic applications), as reviewed by Novikova et al. (2009). Several heterologous CYP450 genes of various phylogenetic origins (from a fungus, a plant, or mammals) have been expressed intracellularly in Y. lipolytica, from fatty acid inducible promoters (pPOX2, pICL1), for use in whole-cell biocatalysis (Shiningavamwe et al. 2006; Bourel et al. 2004; Nthangeni et al. 2004; Braun et al. 2012). As, in contrast to plant CYP450, fungal and mammalian CYP450s require a cofactor, the concomitant expression of the corresponding heterologous cytochrome P450 reductase (CPR) (Braun et al. 2012) or overexpression of the homologous YlCPR (Shiningavamwe et al. 2006; Nthangeni et al. 2004; Braun et al. 2012) was necessary. These studies demonstrated the potential of Y. lipolytica for expression of fungal or mammalian CYP450s and the in vivo efficiency of YlCPR in shuttling electrons to these CYP450s, although human CPR was found to be a more efficient redox partner for human CYP450 (Braun et al. 2012). The Y. lipolytica strain expressing a plant CYP450 (a fatty acid hydroperoxide lyase from Capsicum annuum) was proposed as a whole-cell biocatalyst for the synthesis of volatile aldehydes, for use in the aroma industry (Bourel et al. 2004). A research group from the Graz University of Technology (Austria) demonstrated the potential of two recombinant Y. lipolytica strains expressing human liver CYP450s (CYP2D6 or CYP3A4 genes) and human CPR as whole-cell biocatalysts for the oxidation of hydrophobic steroids in a two-liquid biphasic culture (Braun et al. 2012). In this biphasic system, cells and water-soluble nutrients were maintained in an aqueous phase, when substrates and most of the products were in a second water-immiscible organic solvent phase, which enabled more efficient bioconversion and simplified continuous process (Braun et al. 2012). This work took advantage of the lipophilic properties of Y. lipolytica to propose it as a model for the hydroxylation of other poorly soluble substrates to molecules of biotechnological or therapeutic interest.
Engineering Y. lipolytica metabolic pathways for organic acid production
Although wild-type Y. lipolytica strains have for decades attracted interest as producers of several organic acids, such as pyruvic (PA), citric (CA), isocitric (ICA), α-ketoglutaric (KGA), and succinic acids (SA), engineering strains for such applications is much more recent. Selected examples and yields are given in Table 3 (part C/). A German research group from Dresden University reported that overexpressing aconitase gene (YlACO1) in a multicopy system, in the wild-type H222 strain, resulted in a shift toward ICA of the CA/ICA ratio in organic acids produced from sunflower oil: ICA proportion increased from approximately 40 % in the parent strain to around 70 % in the engineered strain (Holz et al. 2009). The same research group used an engineered H222 strain, harboring an invertase cassette (pXPR2:ScSUC2) and multiple (15) YlICL1 copies to enhance CA production (Förster et al. 2007). A Chinese research group from OUC has displayed a fungal exo-inulinase on the surface of marine SWJ-1b Y. lipolytica strain in order to produce CA and ICA from inulin-containing low-cost raw materials (Liu et al. 2010). This recombinant strain was engineered further, by deleting ATP-citrate lyase gene (YlACL1) and overexpressing YlICL1, in order to increase CA production from inulin (Liu et al. 2013). A consortium of research groups from Jiangnan University (China) explored several engineering strategies on WSH-Z06 strain, a natural KGA overproducer, in order to enhance KGA production while reducing by-product (mainly PA) accumulation (Yin et al. 2012; Zhou et al. 2012; Guo et al. 2014). The most successful approaches were the expression of a fungal pyruvate carboxylase gene (Yin et al. 2012), the heterologous expression of a mammalian ATP-citrate lyase gene (Zhou et al. 2012), or the overexpression of the α subunit of the E1 component from the pyruvate dehydrogenase complex (YlPDA1) (Guo et al. 2014), with increases of KGA yields in the range of one third compared to the wild-type strain. Similarly, a consortium of German research groups (centered on Dresden University and UBZ from Leipzig) engineered H355 strain (a derivative of the wild-type H222 strain with high KGA productivity from alkanes) for improved KGA production using raw glycerol as a renewable carbon source. The effects of overexpressing the following genes in a multicopy system were investigated: three genes encoding the KGA dehydrogenase complex (YlKGD1, YlKGD2, YlLPD1), the fumarase (YlFUM1), and the pyruvate carboxylase (YlPYC1) genes. These studies showed that modifying the expression level of these genes affected both organic acid yields and product spectrum, in a production condition-dependent manner (Holz et al. 2011; Otto et al. 2012). The overexpression of YlFUM1 was shown to drive a substantial increase in KGA yield under adapted production conditions (Otto et al. 2012). More recently, the same Dresden and Leipzig research groups established that overexpression of YlIDP1 gene (encoding NADP+-dependent isocitrate dehydrogenase) or preferably simultaneous overexpression of YlIDP1 and YlPYC1 had the strongest positive effects on KGA production from raw glycerol (Yovkova et al. 2014). Under adapted production conditions, the engineered strain co-overexpressing YlIDP1 and YlPYC1 produced nearly 20 % more KGA than the parent H355 strain (Yovkova et al. 2014). Very recently, the same German research groups showed that reducing the expression of Y. lipolytica succinate dehydrogenase gene by a promoter exchange, in wild-type H222 strain, increased SA production by a more than 12-fold factor (Jost et al. 2015).
Engineering Y. lipolytica lipid metabolism pathways
As attested by the presence in its genome of several multigene families involved in lipid metabolism pathways, Y. lipolytica is an oleaginous yeast able to use a wide range of hydrophobic substrates (notably n-alkanes, fatty acids, and triacylglycerols (TAGs)). It can accumulate lipids up to more than 40 % of its dry cell weight (DCW) and is the only yeast known in which the proportion of linoleic acid is so high (more than 50 % of fatty acids). It is also the only one for which efficient genetic tools are available. Y. lipolytica constitutes a model organism for hydrophobic substrate degradation and lipid accumulation, which have been studied notably at INRA (France) (Beopoulos et al. 2008, 2009a, b) and at Tokyo University (Japan) (Fukuda 2013; Iwama et al. 2014). Depending on C/N ratio, different metabolisms predominate in Y. lipolytica: cell growth, organic acid production, or lipid accumulation (mainly TAGs and steryl esters), which occurs notably under nitrogen limitation via de novo synthesis or ex novo accumulation (Beopoulos et al. 2009a). Lipid accumulation has been shown to depend on the peroxisomal β-oxidation flux, regulated by six acyl-CoA oxidases with different substrate specificities (genes POX1 to POX6). The size and fatty acid composition of intracellular lipid bodies is determined by the POX genotype. For example, the overexpression of POX2 in a quadruple mutant Δpox2-5 resulted in an “obese yeast” phenotype (Beopoulos et al. 2009a). The same INRA research group obtained, by combining mutations in the glycerol-3-phosphate shuttle pathway (Δgut2 and overexpression of YlGPD1) with inactivation of the β-oxidation pathway (POX1-6 or MFE1 genes), strains that accumulated lipids up to 75 % of DCW (Dulermo and Nicaud 2011). Several research groups from Jiangnan University (China) enhanced lipid accumulation in Y. lipolytica by increasing conversion of citrate to acetyl-CoA via expression of a mammalian ATP-citrate lyase gene (Zhang et al. 2014). In consequence of the oleaginous status of Y. lipolytica, most proposed applications involve engineering its lipid metabolism pathways. They took advantage of the very effective uptake of lipids, characteristic of this yeast, which can be attributed to several factors: production of bio-surfactants (e.g., liposan), action of extracellular lipases, modification of cell surface facilitating adhesion of hydrophobic droplets (formation of protrusions), and complex transport mechanisms (Beopoulos et al. 2009a). Some major examples of the engineering of Y. lipolytica lipid metabolism pathways for bioconversion will be examined further.
Single-cell oil production for health applications
Microorganisms can be a valuable source of edible SCOs enriched in essential fatty acids, namely polyunsaturated fatty acids (PUFAs) not synthesized by mammals but essential for health. PUFAs must be supplied by diet, and SCOs can play a role in their supply, as a cheaper and environment-friendly alternative to plant or fish oils. There is a strong economic interest in engineering Y. lipolytica ω-3/ω-6 biosynthetic pathway for producing SCO enriched in ω-3 or ω-6 fatty acids for use as dietary supplements in feed or food. The main targets are the ω-3 eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA) and α-linolenic acid (ALA), and the ω-6 arachidonic acid (AA) and γ-linolenic acid (GLA). Selected examples and yields are given in Table 3 (part C/). Research groups from DuPont Company (USA) have engineered a Y. lipolytica strain for ALA production by expressing a bifunctional fungal Δ12/ω3 desaturase (Damude et al. 2006). DuPont Company has also patented a process for optimizing EPA production in Y. lipolytica (Picataggio et al. 2003) and has marketed EPA-rich Y. lipolytica biomass for use in fish feed (VerlassoTM) and EPA-rich SCO for use as dietary supplement (New HarvestTM). The corresponding Y. lipolytica production strain constitutes the first recombinant microorganisms able to achieve the productivity necessary for commercial EPA production (Xue et al. 2013). It was obtained through extensive remodeling of its metabolic pathways: In order to produce EPA from endogenous LA, while avoiding GLA buildup, the Δ-9 pathway was selected by overexpressing several (more than 20) heterologous enzymes (Δ-9 elongases, Δ-8 desaturases, Δ-5 desaturases, Δ-17 desaturases, Δ12 desaturases, and C16/18 elongases) and by knocking out three homologous genes encoding a lipase, a sterol carrier, and a protein involved in peroxisomal import (Xue et al. 2013). More recently, a consortium of Taiwanese (including Yeastern Biotech Co.) and French (INRA) research groups obtained efficient GLA production in Y. lipolytica by co-expressing two Δ6 and Δ12 fungal desaturases (Chuang et al. 2010).
Single-cell oils for biofuel production
The search for renewable fuels as petroleum alternatives has prompted study of SCOs as a potentially viable source of fatty acids for biodiesel synthesis. As an oleaginous yeast able to grow on various agro-industrial by-products or wastes, Y. lipolytica constitutes an interesting target for biofuel production. The yeast TAGs can be transesterified into fatty acid methyl esters (FAMEs), which constitute the main component of biodiesel. Transesterification can occur as a distinct in vitro process or can be integrated as a direct in vivo step, in an engineered cell factory. In order to make the use of SCOs for biofuel production economically feasible, low-cost feedstock needs to be employed as the carbon source for both biomass production and neutral lipid synthesis. Various waste feedstocks have been proposed for that purpose, such as biodiesel-derived waste glycerol, lignocellulosic hydrolysates, used oils, sewage, and agricultural or food-processing wastes (Cheirsilp and Louhasakul 2013; Sestric et al. 2014 and references therein). A Chinese research group from OUC has expressed a fungal exo-inulinase into a marine Y. lipolytica strain in order to obtain high SCO yields (more than 50 % of DCW) for biofuel production from inulin-containing low-cost raw materials (Zhao et al. 2010). Like other oleaginous yeasts, Y. lipolytica lacks the array of cellulolytic enzymes required to break down cellulose and cannot therefore use vegetal biomass directly as a carbon source. In order to develop the production of SCOs from lignocellulosic biomass, composed of carbohydrate polymers (cellulose, hemicellulose) and lignin (aromatic polymer), research groups from the National Renewable Energy Laboratory (USA) have engineered Y. lipolytica strains for expression of fungal cellulases and xylanases (Wei et al. 2014; Wang et al. 2014). A chimeric cellobiohydrolase I, secreted by Y. lipolytica, exhibited a synergistic effect on the degradation of cellulosic substrates when associated with an endoglucanase II and a cellobiohydrolase II in a consortia system approach (Wei et al. 2014). Similarly, a synergistic effect on the conversion of xylan to xylose was observed when two fungal xylanases (an endo-xylanase and an exo-xylosidase) secreted by Y. lipolytica strains were used (Wang et al. 2014).
In order to establish Y. lipolytica as a platform for biofuel production, its metabolic flux needs to be redirected toward TAG synthesis. Besides the obtaining of obese yeast phenotypes at INRA (France), as cited above (Beopoulos et al. 2009a; Dulermo and Nicaud 2011), a research group from Massachusetts Institute of Technology (MIT, USA) co-overexpressed diacylglycerol acyltransferase (final step of the TAG synthesis) and acetyl-CoA carboxylase (first step of fatty acid synthesis), thus obtaining a synergistic effect on lipid production which reached 62 % of DCW (Tai and Stephanopoulos 2013).
Very recently, the department of Chemical engineering from the University of Texas at Austin (USA) brought unprecedented improvement in harnessing Y. lipolytica for lipid and biofuel production. They rewired Y. lipolytica’s metabolism for maximum de novo lipogenesis by using a combinatorial multiplexing approach (Blazeck et al. 2014). The overexpression of five lipogenesis enzymes was multiplexed with deletions that reduced fatty-acid catabolism by impairing β-oxidation (mfe1 deletion), peroxisome biogenesis (pex10 deletion) or both, and coupled with phenotypic induction. This combinatorial multiplexing produced 57 genotypes that were analyzed for their lipogenesis capacity, in comparison with the wild-type strain. The Δpex10, Δmfe1, leucine+, uracil+, and YlDGA1 (acyl-CoA:diacylglycerol acyltransferase isozyme I) overexpression genotype was selected as the strain exhibiting the highest lipogenesis potential: It allowed increasing the lipid content up to 90 % of DCW, which represents an enhancement of de novo lipid accumulation of 60-fold compared to the parental Po1f strain (Blazeck et al. 2014). This study also advanced the fundamental understanding of lipogenesis by presenting evidence that two of its central tenets, the necessity for nitrogen starvation and citric acid cycling, are not universal. It reported the highest lipid titer obtained for Y. lipolytica (25 g/l) and constitutes a stepping stone in creating a robust and efficient platform for industrial production of value-added oleochemicals and biofuels. Previously, the same research groups had also engineered Y. lipolytica for pentane production from linoleic acid, by overexpressing a soybean lipoxygenase, revealing the potential of this yeast for short chain n-alkane synthesis (Blazeck et al. 2013b).
Engineering Y. lipolytica for carotenoid production
Carotenoids are lipophilic organic pigments widely used as natural coloring and antioxidant agents for food and feed and also as dietary supplements. DuPont (USA) and Microbia (DSM, USA) (safety data of recombinant β-carotene disclosed in Grenfell-Lee et al. 2014) are independently using GM Y. lipolytica for carotenoid production (Groenewald et al. 2014). Selected examples and yields are given in Table 3 (part C/). DuPont Company has engineered Y. lipolytica for the production of lycopene, a carotene that is also an intermediate in the biosynthesis of other carotenoids, by expressing three bacterial genes from the CRT cluster (Ye et al. 2012). More recently, the production of unprecedented lycopene levels for a eukaryotic host was reported by a research group from Dresden University (Germany) by using an “obese” strain, developed at INRA (France), in which two bacterial genes from the CRT cluster were expressed and two Y. lipolytica rate-limiting genes for isoprenoid biosynthesis were overexpressed (Matthäus et al. 2014). At last, Gao et al. (2014) recently applied a DNA assembler method to engineer Y. lipolytica for carotenoid production: Two distinct β-carotene biosynthesis pathways were targeted at the rDNA locus of a Y. lipolytica strain, using an in vivo multiple fragment assembly method. The best results, in terms of both pathway construction efficiency and β-carotene production level, were obtained by integrating a selection marker and three expression cassettes, composed of different promoters and terminators, driving expression of two fungal genes from Mucor circinelloides (carB encoding phytoene dehydrogenase and carRP encoding a bifunctional enzyme with lycopene cyclase and phytoene synthase activities) and overexpression of YlGGS1 (rate-limiting gene for isoprenoid biosynthesis). As high as 20 % of the transformants were found to produce β-carotene, and the best production levels were in the range of 2 mg per g DCW (Gao et al. 2014).
Y. lipolytica as a source of valuable lipid-derived compounds for the future?
Besides its potential as provider for PUFA-enriched SCOs, biofuels, and carotenoids, Y. lipolytica is also expected to become a source for other high-value lipid-derived products like polyhydroxyalkanoates (PHAs), wax esters (industrial lubricants), and free hydroxylated fatty acids (HFAs, used as chiral building blocks in fine chemical synthesis). A consortium of research groups, including notably laboratories from Ghent University (Belgium) and INRA (France), is currently using specific enzymatic pathways from hydrocarbonoclastic bacteria and from other hydrocarbon or lipid-degrading bacteria isolated from contaminated environments, to redirect Y. lipolytica lipid metabolism toward the production of valuable lipid-derived compounds (Sabirova et al. 2011). Up to now, the most advanced projects concern PHAs, biodegradable linear polyesters naturally produced by some bacteria from excess supply of sugar or lipids, which can be used for producing bioplastics. Recombinant Y. lipolytica strains expressing a peroxisome-targeted bacterial PHA synthase gene were shown to produce various PHA yields, depending on their POX genotype (Haddouche et al. 2010). The same research groups were then able to increase PHA production up to more than 7 % of DCW by deleting several genes from Y. lipolytica neutral lipid synthesis pathway (LRO1, DGA1, DGA2, and ARE1), thus redirecting the fatty acid flux toward β-oxidation (Haddouche et al. 2011). The composition of the PHAs produced was also shown to be dependent on the Y. lipolytica β-oxidation multifunctional enzyme, since the use of different variants (deletions or inactivations of the R-3-hydroxyacyl-CoA dehydrogenase domain) influenced the PHA structure (hetero- or homo-polymers) (Haddouche et al. 2011). These studies constitute a stepping stone in establishing Y. lipolytica as a potential platform for converting cheap and vastly available lipid substrates into high-value biotechnological, nutritional, or pharmacological products.
Future perspectives
Since several decades, Y. lipolytica has promoted interest for multiple uses such as organic acids, SCP, and SCO production or bioremediation and upgrading of industrial wastes. Y. lipolytica has also become a model for basic research, and the development of advanced genetic and molecular tools has established it as an efficient platform for heterologous protein expression/secretion. The relatively recent development of surface display and oleosome targeting methods has opened new possibilities of using arming Y. lipolytica cells or derived nanoparticles for applications in molecular recognition or bioconversion. Further improvements of the Y. lipolytica expression system will probably be realized in the near future, through strain engineering. Complete genome, transcriptome, and proteome data will allow to identify bottlenecks in protein secretion and quality control pathways and to study the response of cells challenged with the stress of overproducing heterologous proteins. The development of high-throughput screening platforms will favor the use of Y. lipolytica for protein evolution, production, and structure analysis. The development of -omic approaches and of metabolic modeling will offer new perspectives in metabolic pathway engineering, allowing a better use of Y. lipolytica as cell factory. The recent proof of concept of the possible use of DNA assembler methods in Y. lipolytica will also facilitate the engineering of this yeast. Some new tools would however be desirable, such as ORFome, knockout or overexpression libraries, a two-hybrid system or fully repressible strong inducible promoters. It remains also critical to know if, despite its GRAS status, Y. lipolytica could be socially acceptable for use as a GMM, especially for whole-cell applications in the environmental domain or in food industry.
References
Bankar AV, Kumar AR, Zinjarde SS (2009) Environmental and industrial applications of Yarrowia lipolytica. Appl Microbiol Biotechnol 84:847–865. doi:10.1007/s00253-009-2156-8
Barth G (2013a) Yarrowia lipolytica—genetics, genomics, and Physiology. Microbiology Monographs Vol. 24, Springer, Heidelberg, Germany. doi: 10.1007/978-3-642-38320-5
Barth G (2013b) Yarrowia lipolytica—biotechnological applications. Microbiology Monographs Vol. 25, Springer, Heidelberg, Germany. doi:10.1007/978-3-642-38583-4
Barth G, Gaillardin C (1996) Yarrowia lipolytica. In: Wolf K (ed) Nonconventional yeasts in biotechnology: a handbook. Springer-Verlag, Heidelberg, Germany, pp 313–388
Beopoulos A, Mrozova Z, Thevenieau F, Le Dall MT, Hapala I, Papanikolaou S, Chardot T, Nicaud JM (2008) Control of lipid accumulation in the yeast Yarrowia lipolytica. Appl Environ Microbiol 74:7779–7789. doi:10.1128/AEM.01412-08
Beopoulos A, Chardot T, Nicaud JM (2009a) Yarrowia lipolytica: a model and a tool to understand the mechanisms implicated in lipid accumulation. Biochimie 91:692–696. doi:10.1016/j.biochi.2009.02.004
Beopoulos A, Cescut J, Haddouche R, Uribelarrea JL, Molina-Jouve C, Nicaud JM (2009b) Yarrowia lipolytica as a model for bio-oil production. Prog Lipid Res 48:375–387. doi:10.1016/j.plipres.2009.08.005
Blazeck J, Liu L, Redden H, Alper H (2011) Tuning gene expression in Yarrowia lipolytica by a hybrid promoter approach. Appl Environ Microbiol 77:7905–7914. doi:10.1128/AEM.05763-11
Blazeck J, Reed B, Garg R, Gerstner R, Pan A, Agarwala V, Alper H (2013a) Generalizing a hybrid synthetic promoter approach in Yarrowia lipolytica. Appl Microbiol Biotechnol 97:3037–3052. doi:10.1007/s00253-012-4421-5
Blazeck J, Liu L, Knight R, Alper HS (2013b) Heterologous production of pentane in the oleaginous yeast Yarrowia lipolytica. J Biotechnol 165:184–194. doi:10.1016/j.jbiotec.2013.04.003
Blazeck J, Hill A, Liu L, Knight R, Miller J, Pan A, Otoupal P, Alper HS (2014) Harnessing Yarrowia lipolytica lipogenesis to create a platform for lipid and biofuel production. Nat Commun 5:3131. doi:10.1038/ncomms4131
Boisramé A, Kabani M, Beckerich JM, Hartmann E, Gaillardin C (1998) Interaction of Kar2p and Sls1p is required for efficient co-translational translocation of secreted proteins in the yeast Yarrowia lipolytica. J Biol Chem 273:30903–30908. doi:10.1074/jbc.273.47.30903
Bordes F, Fudalej F, Dossat V, Nicaud JM, Marty A (2007) A new recombinant protein expression system for high-throughput screening in the yeast Yarrowia lipolytica. J Microbiol Methods 70:493–502. doi:10.1016/j.mimet.2007.06.008
Bourel G, Nicaud JM, Nthangeni B, Santiago-Gomez P, Belin JM, Husson F (2004) Fatty acid hydroperoxide lyase of the green bell pepper: cloning in Yarrowia lipolytica and biogenesis of volatile aldehydes. Enzyme Microb Technol 35:293–299. doi:10.1016/j.enzmictec.2003.12.014
Braun A, Geier M, Bühler B, Schmid A, Mauersberger S, Glieder A (2012) Steroid biotransformations in biphasic systems with Yarrowia lipolytica expressing human liver cytochrome P450 genes. Microb Cell Fact 11:106. doi:10.1186/1475-2859-11-106
Callewaert NLM, Vervecken W, Marcel de Pourcq KJ, Geysens SCJ, Guerfal M (2007) Glycosylation of molecules. International Patent Application WO2008120107.
Celińska E, Grajek W (2013) A novel multigene expression construct for modification of glycerol metabolism in Yarrowia lipolytica. Microb Cell Fact 12:102. doi:10.1186/1475-2859-12-102
Celińska E, Białas W, Borkowska M, Grajek W (2014) Cloning, expression, and purification of insect (Sitophilus oryzae) alpha-amylase, able to digest granular starch, in Yarrowia lipolytica host. Appl Microbiol Biotechnol 99:2727–2739. doi:10.1007/s00253-014-6314-2
Chang CC, Ryu DD, Park CS, Kim JY (1998) Improvement of heterologous protein productivity using recombinant Yarrowia lipolytica and cyclic fed-batch process strategy. Biotechnol Bioeng 59:379–385. doi:10.1002/(SICI)1097-0290(19980805)59:3<379::AID-BIT15>3.0.CO;2-9
Cheirsilp B, Louhasakul Y (2013) Industrial wastes as a promising renewable source for production of microbial lipid and direct transesterification of the lipid into biodiesel. Bioresour Technol 142:329–337. doi:10.1016/j.biortech.2013.05.012
Chuang LT, Chen DC, Nicaud JM, Madzak C, Chen YH, Huang YS (2010) Co-expression of heterologous desaturase genes in Yarrowia lipolytica. N Biotechnol 27:277–282. doi:10.1016/j.nbt.2010.02.006
Cui W, Wang Q, Zhang F, Zhang SC, Chi ZM, Madzak C (2011) Direct conversion of inulin into single cell protein by the engineered Yarrowia lipolytica carrying inulinase gene. Process Biochem 46:1442–1448. doi:10.1016/j.procbio.2011.03.017
Damude HG, Zhang H, Farrall L, Ripp KG, Tomb JF, Hollerbach D, Yadav NS (2006) Identification of bifunctional delta12/omega3 fatty acid desaturases for improving the ratio of omega3 to omega6 fatty acids in microbes and plants. Proc Natl Acad Sci USA 103:9446–9451. doi:10.1073/pnas.0511079103
Davidow LS, DeZeeuw JR (1984) Process for transformation of Yarrowia lipolytica. US Patent Application US4880741
De Pourcq K, Vervecken W, Dewerte I, Valevska A, Van Hecke A, Callewaert N (2012a) Engineering the yeast Yarrowia lipolytica for the production of therapeutic proteins homogeneously glycosylated with Man8GlcNAc2 and Man5GlcNAc2. Microb Cell Fact 11:53. doi:10.1186/1475-2859-11-53
De Pourcq K, Tiels P, Van Hecke A, Geysens S, Vervecken W, Callewaert N (2012b) Engineering Yarrowia lipolytica to produce glycoproteins homogeneously modified with the universal Man3GlcNAc2 N-glycan core. PLoS One 7:e39976. doi:10.1371/journal.pone.0039976
Dominguez A, Ferminan E, Sanchez M, Gonzalez FJ, Perez-Campo FM, Garcia S, Herrero AB, San Vicente A, Cabello J, Prado M, Iglesias FJ, Choupina A, Burguillo FJ, Fernandez-Lago L, Lopez MC (1998) Non-conventional yeasts as hosts for heterologous protein production. Internatl Microbiol 1:131–142
Dujon B, Sherman D, Fischer G, Durrens P, Casaregola S, Lafontaine I, De Montigny J, Marck C, Neuveglise C, Talla E, Goffard N, Frangeul L, Aigle M, Anthouard V, Babour A, Barbe V, Barnay S, Blanchin S, Beckerich JM, Beyne E, Bleykasten C, Boisrame A, Boyer J, Cattolico L, Confanioleri F, De Daruvar A, Despons L, Fabre E, Fairhead C, Ferry-Dumazet H, Groppi A, Hantraye F, Hennequin C, Jauniaux N, Joyet P, Kachouri R, Kerrest A, Koszul R, Lemaire M, Lesur I, Ma L, Muller H, Nicaud JM, Nikolski M, Oztas S, Ozier-Kalogeropoulos O, Pellenz S, Potier S, Richard GF, Straub ML, Suleau A, Swennen D, Tekaia F, Wesolowski-Louvel M, Westhof E, Wirth B, Zeniou-Meyer M, Zivanovic I, Bolotin-Fukuhara M, Thierry A, Bouchier C, Caudron B, Scarpelli C, Gaillardin C, Weissenbach J, Wincker P, Souciet JL (2004) Genome evolution in yeasts. Nature 430:35–44. doi:10.1038/nature02579
Dulermo T, Nicaud JM (2011) Involvement of the G3P shuttle and β-oxidation pathway in the control of TAG synthesis and lipid accumulation in Yarrowia lipolytica. Metab Eng 13:482–491. doi:10.1016/j.ymben.2011.05.002
Duquesne S, Bozonnet S, Bordes F, Dumon C, Nicaud J-M, Marty A (2014) Construction of a highly active xylanase displaying oleaginous yeast: comparison of anchoring systems. PLoS ONE 9:e95128. doi:10.1371/journal.pone.0095128
Emond S, Montanier C, Nicaud JM, Marty A, Monsan P, André I, Remaud-Siméon M (2010) New efficient recombinant expression system to engineer Candida antarctica lipase B. Appl Environ Microbiol 76:2684–2687. doi:10.1128/AEM.03057-09
Fickers P, Le Dall MT, Gaillardin C, Thonart P, Nicaud JM (2003) New disruption cassettes for rapid gene disruption and marker rescue in the yeast Yarrowia lipolytica. J Microbiol Methods 55:727–37. doi:10.1016/j.mimet.2003.07.003
Förster A, Aurich A, Mauersberger S, Barth G (2007) Citric acid production from sucrose using a recombinant strain of the yeast Yarrowia lipolytica. Appl Microbiol Biotechnol 75:1409–1417. doi:10.1007/s00253-007-0958-0
Fournier P, Abbas A, Chasles M, Kudla B, Ogrydziak DM, Yaver D, Xuan JW, Peito A, Ribet AM, Feynerol C, He F, Gaillardin C (1993) Colocalization of centromeric and replicative functions on autonomously replicating sequences isolated from the yeast Yarrowia lipolytica. Proc Natl Acad Sci 90:4912–4916
Franke AE, Kaczmarek FS, Eisenhard ME, Geoghegan KF, Danley DE, DeZeeuw JR, O'Donnell MM, Gollaher MG, Davidow LS (1988) Expression and secretion of bovine prochymosin in Yarrowia lipolytica. Devel Ind Microbiol 29:43–57
Fukuda R (2013) Metabolism of hydrophobic carbon sources and regulation of it in n-alkane-assimilating yeast Yarrowia lipolytica. Biosci Biotechnol Biochem 77:1149–1154. doi:10.1271/bbb.130164
Gaillardin C, Ribet AM (1987) LEU2 directed expression of β-galactosidase activity and phleomycin resistance in Yarrowia lipolytica. Curr Genet 11:369–375
Gaillardin C, Heslot H, Ribet AM (1984) Vecteurs de transformation de la levure Yarrowia lipolytica, procédé de transformation et levure transformée. French Patent Application FR2566424.
Galli C, Gentili P, Jolivalt C, Madzak C, Vadalà R (2011) How is the reactivity of laccase affected by single-point mutations? Engineering laccase for improved activity towards sterically demanding substrates. Appl Microbiol Biotechnol 91:123–131. doi:10.1007/s00253-011-3240-4
Gao S, Han L, Zhu L, Ge M, Yang S, Jiang Y, Chen D (2014) One-step integration of multiple genes into the oleaginous yeast Yarrowia lipolytica. Biotechnol Lett 36:2523–2528. doi:10.1007/s10529-014-1634-y
Gasmi N, Fudalej F, Kallel H, Nicaud JM (2011a) A molecular approach to optimize hIFN α2b expression and secretion in Yarrowia lipolytica. Appl Microbiol Biotechnol 89:109–119. doi:10.1007/s00253-010-2803-0
Gasmi N, Ayed A, Ammar BB, Zrigui R, Nicaud JM, Kallel H (2011b) Development of a cultivation process for the enhancement of human interferon alpha 2b production in the oleaginous yeast, Yarrowia lipolytica. Microb Cell Fact 10:90–100. doi:10.1186/1475-2859-10-90
Grenfell-Lee D, Zeller S, Cardoso R, Pucaj K (2014) The safety of β-carotene from Yarrowia lipolytica. Food Chem Toxicol 65:1–11. doi:10.1016/j.fct.2013.12.010
Groenewald M, Boekhout T, Neuvéglise C, Gaillardin C, van Dijck PW, Wyss M (2014) Yarrowia lipolytica: safety assessment of an oleaginous yeast with a great industrial potential. Crit Rev Microbiol 40:187–206. doi:10.3109/1040841X.2013.770386
Guo H, Madzak C, Du G, Zhou J, Chen J (2014) Effects of pyruvate dehydrogenase subunits overexpression on the α-ketoglutarate production in Yarrowia lipolytica WSH-Z06. Appl Microbiol Biotechnol 98:7003–12. doi:10.1007/s00253-014-5745-0
Haddouche R, Delessert S, Sabirova J, Neuvéglise C, Poirier Y, Nicaud JM (2010) Roles of multiple acyl-CoA oxidases in the routing of carbon flow towards β-oxidation and polyhydroxyalkanoate biosynthesis in Yarrowia lipolytica. FEMS Yeast Res 10:917–927. doi:10.1111/j.1567-1364.2010.00670.x
Haddouche R, Poirier Y, Delessert S, Sabirova J, Pagot Y, Neuvéglise C, Nicaud JM (2011) Engineering polyhydroxyalkanoate content and monomer composition in the oleaginous yeast Yarrowia lipolytica by modifying the ß-oxidation multifunctional protein. Appl Microbiol Biotechnol 91:1327–1340. doi:10.1007/s00253-011-3331-2
Han Z, Madzak C, Su WW (2013) Tunable nano-oleosomes derived from engineered Yarrowia lipolytica. Biotechnol Bioeng 110:702–710. doi:10.1002/bit.24761
Holz M, Förster A, Mauersberger S, Barth G (2009) Aconitase overexpression changes the product ratio of citric acid production by Yarrowia lipolytica. Appl Microbiol Biotechnol 81:1087–1096. doi:10.1007/s00253-008-1725-6
Holz M, Otto C, Kretzschmar A, Yovkova V, Aurich A, Pötter M, Marx A, Barth G (2011) Overexpression of alpha-ketoglutarate dehydrogenase in Yarrowia lipolytica and its effect on production of organic acids. Appl Microbiol Biotechnol 89:1519–1526. doi:10.1007/s00253-010-2957-9
Hong SP, Seip J, Walters-Pollak D, Rupert R, Jackson R, Xue Z, Zhu Q (2012) Engineering Yarrowia lipolytica to express secretory invertase with strong FBA1(IN) promoter. Yeast 29:59–72. doi:10.1002/yea.1917
Iwama R, Kobayashi S, Ohta A, Horiuchi H, Fukuda R (2014) Fatty aldehyde dehydrogenase multigene family involved in the assimilation of n-alkanes in Yarrowia lipolytica. J Biol Chem 289:33275–33286. doi:10.1074/jbc.M114.596890
Jolivalt C, Madzak C, Brault A, Caminade E, Malosse C, Mougin C (2005) Expression of laccase IIIb from the white-rot fungus Trametes versicolor in the yeast Yarrowia lipolytica for environmental applications. Appl Microbiol Biotechnol 66:450–6. doi:10.1007/s00253-004-1717-0
Jost B, Holz M, Aurich A, Barth G, Bley T, Müller RA (2015) The influence of oxygen limitation for the production of succinic acid with recombinant strains of Yarrowia lipolytica. Appl Microbiol Biotechnol 99:1675–1686. doi:10.1007/s00253-014-6252-z
Juretzek T, Wang HJ, Nicaud JM, Mauersberger S, Barth G (2000) Comparison of promoters suitable for regulated overexpression of β-galactosidase in the alkane-utilizing yeast Yarrowia lipolytica. Biotechnol Bioprocess Eng 5:320–326. doi:10.1007/BF02942206
Juretzek T, Le Dall M, Mauersberger S, Gaillardin C, Barth G, Nicaud JM (2001) Vectors for gene expression and amplification in the yeast Yarrowia lipolytica. Yeast 18:97–113. doi:10.1002/1097-0061(20010130)18:2<97::AID-YEA652>3.0.CO;2-U
Kang HA, Kim JY, Oh DB, Park JN, Song Y (2007) A novel ylmpo1 gene derived from Yarrowia lipolytica and a process for preparing a glycoprotein not being mannosylphosphorylated by using a mutated Yarrowia lipolytica in which ylmpo1 gene is disrupted. International Patent Application WO2008136564
Kim JW, Park TJ, Ryu DD, Kim JY (2000) High cell density culture of Yarrowia lipolytica using a one-step feeding process. Biotechnol Prog 16:657–60. doi:10.1021/bp000037n
Kim H, Yoo SJ, Kang HA (2014) Yeast synthetic biology for the production of recombinant therapeutic proteins. FEMS Yeast Res. doi:10.1111/1567-1364.12195
Labuschagné M (2005) Cloning and expression of epoxide hydrolases: Yarrowia lipolytica as an attractive heterologous expression host. PhD Thesis, University of the Free State, South Africa
Labuschagné M, Albertyn J (2007) Cloning of an epoxide hydrolase-encoding gene from Rhodotorula mucilaginosa and functional expression in Yarrowia lipolytica. Yeast 24:69–78. doi:10.1002/yea.1437
Lazar Z, Rossignol T, Verbeke J, Crutz-Le Coq AM, Nicaud JM, Robak M (2013) Optimized invertase expression and secretion cassette for improving Yarrowia lipolytica growth on sucrose for industrial applications. J Ind Microbiol Biotechnol 40:1273–83. doi:10.1007/s10295-013-1323-1
Le Dall MT, Nicaud JM, Gaillardin C (1994) Multiple-copy integration in the yeast Yarrowia lipolytica. Curr Genet 26:38–44. doi:10.1007/BF00326302
Liu L, Alper HS (2014) Draft genome sequence of the oleaginous yeast Yarrowia lipolytica PO1f, a commonly used metabolic engineering host. Genome Announc 2:e00652–14. doi:10.1128/genomeA.00652-14
Liu XY, Chi Z, Liu GL, Wang F, Madzak C, Chi ZM (2010) Inulin hydrolysis and citric acid production from inulin using the surface-engineered Yarrowia lipolytica displaying inulinase. Metab Eng 12:469–476. doi:10.1016/j.ymben.2010.04.004
Liu XY, Chi Z, Liu GL, Madzak C, Chi ZM (2013) Both decrease in ACL1 gene expression and increase in ICL1 gene expression in marine-derived yeast Yarrowia lipolytica expressing INU1 gene enhance citric acid production from inulin. Mar Biotechnol (NY) 15:26–36. doi:10.1007/s10126-012-9452-5
Liu L, Otoupal P, Pan A, Alper HS (2014) Increasing expression level and copy number of a Yarrowia lipolytica plasmid through regulated centromere function. FEMS Yeast Res 14:1124–7. doi:10.1111/1567-1364.12201
Loira N, Dulermo T, Nicaud JM, Sherman DJ (2012) A genome-scale metabolic model of the lipid-accumulating yeast Yarrowia lipolytica. BMC Syst Biol 6:35. doi:10.1186/1752-0509-6-35
Madzak C and Beckerich JM (2013) Heterologous protein expression and secretion in Yarrowia lipolytica. In: Barth G (ed) Yarrowia lipolytica: Biotechnological Applications, Microbiology Monographs vol. 25, Springer, Heidelberg, Germany, pp 1–76. doi:10.1007/978-3-642-38583-4_1
Madzak C, Blanchin-Roland S, Gaillardin C (1995) Upstream activating sequences and recombinant promoter sequences functional in Yarrowia and vectors containing them. European Patent Application EP0747484
Madzak C, Blanchin-Roland S, Cordero Otero R, Gaillardin C (1999) Functional analysis of upstream regulating regions from the Yarrowia lipolytica XPR2 promoter. Microbiology 145:75–87. doi:10.1099/13500872-145-1-75
Madzak C, Tréton B, Blanchin-Roland S (2000) Strong hybrid promoters and integrative expression/secretion vectors for quasi-constitutive expression of heterologous proteins in the yeast Yarrowia lipolytica. J Mol Microbiol Biotechnol 2:207–216. doi:10.1159/issn.1464-1801
Madzak C, Gaillardin C, Beckerich JM (2004) Heterologous protein expression and secretion in the non-conventional yeast Yarrowia lipolytica: a review. J Biotechnol 109:63–81. doi:10.1016/j.jbiotec.2003.10.027
Madzak C, Otterbein L, Chamkha M, Moukha S, Asther M, Gaillardin C, Beckerich JM (2005) Heterologous production of a laccase from the basidiomycete Pycnoporus cinnabarinus in the dimorphic yeast Yarrowia lipolytica. FEMS Yeast Res 5:635–646. doi:10.1016/j.femsyr.2004.10.009
Madzak C, Mimmi MC, Caminade E, Brault A, Baumberger S, Briozzo P, Mougin C, Jolivalt C (2006) Shifting the optimal pH of activity for a laccase from the fungus Trametes versicolor by structure-based mutagenesis. Protein Eng Des Sel 19:77–84. doi:10.1093/protein/gzj004
Maharajh D, Roth R, Lalloo R, Simpson C, Mitra R, Görgens J, Ramchuran S (2008a) Multi-copy expression and fed-batch production of Rhodotorula araucariae epoxide hydrolase in Yarrowia lipolytica. Appl Microbiol Biotechnol 79:235–44. doi:10.1007/s00253-008-1420-7
Maharajh D, Lalloo R, Görgens J (2008b) Effect of an exponential feeding regime on the production of Rhodotorula araucariae epoxide hydrolase in Yarrowia lipolytica. Lett Appl Microbiol 47:520–5. doi:10.1111/j.1472-765X.2008.02425.x
Matsuoka M, Matsubara M, Daidoh H, Imanaka T, Uchida K, Aiba S (1993) Analysis of regions essential for the function of chromosomal replicator sequences from Yarrowia lipolytica. Mol Gen Genet 237:327–333. doi:10.1007/BF00279435
Matthäus F, Ketelhot M, Gatter M, Barth G (2014) Production of lycopene in the non-carotenoid-producing yeast Yarrowia lipolytica. Appl Environ Microbiol 80:1660–1669. doi:10.1128/AEM.03167-13
Moon HY, Van TL, Cheon SA, Choo J, Kim JY, Kang HA (2013) Cell-surface expression of Aspergillus saitoi-derived functional α-1,2-mannosidase on Yarrowia lipolytica for glycan remodeling. J Microbiol 51:506–14. doi:10.1007/s12275-013-3344-x
Mougin C, Jolivalt C, Briozzo P, Madzak C (2003) Fungal laccases: from structure-activity studies to environmental applications. Environ Chem Lett 1:145–148. doi:10.1007/s10311-003-0024-9
Müller S, Sandal T, Kamp-Hansen P, Dalboge H (1998) Comparison of expression systems in the yeasts Saccharomyces cerevisiae, Hansenula polymorpha, Kluyveromyces lactis, Schizosaccharomyces pombe and Yarrowia lipolytica. Cloning of two novel promoters from Yarrowia lipolytica. Yeast 14:1267–1283. doi:10.1002/(SICI)1097-0061(1998100)14:14<1267::AID-YEA327>3.0.CO;2-2
Ni X, Chi Z, Ma C, Madzak C (2008) Cloning, characterization, and expression of the gene encoding alkaline protease in the marine yeast Aureobasidium pullulans 10. Mar Biotechnol (NY) 10:319–327. doi:10.1007/s10126-007-9067-4
Ni X, Yue L, Chi Z, Li J, Wang X, Madzak C (2009) Alkaline protease gene cloning from the marine yeast Aureobasidium pullulans HN2-3 and the protease surface display on Yarrowia lipolytica for bioactive peptide production. Mar Biotechnol (NY) 11(1):81–89. doi:10.1007/s10126-008-9122-9
Nicaud JM (2012) Yarrowia lipolytica. Yeast 29:409–418. doi:10.1002/yea.2921
Nicaud JM, Fabre E, Gaillardin C (1989) Expression of invertase activity in Yarrowia lipolytica and its use as a selective marker. Curr Genet 16:253–260. doi:10.1007/BF00422111
Nicaud JM, Gaillardin C, Seman M, Pignède G (1998) Procédé de transformation non-homologue de Yarrowia lipolytica. French Patent Application FR2782733
Nicaud JM, Madzak C, van den Broek P, Gysler C, Duboc P, Niederberger P, Gaillardin C (2002) Protein expression and secretion in the yeast Yarrowia lipolytica. FEMS Yeast Res 2:371–379. doi:10.1016/S1567-1356(02)00082-X
Nicaud JM, Fudalej F, Neuvéglise C, Beckerich JM (2008) Procédé d'intégration ciblée de multicopies d'un gène d'intérêt dans une souche de Yarrowia. French Patent Application FR2927089
Novikova LA, Faletrov YV, Kovaleva IE, Mauersberger S, Luzikov VN, Shkumatov VM (2009) From structure and functions of steroidogenic enzymes to new technologies of gene engineering. Biochemistry (Mosc) 74:1482–1504
Nthangeni MB, Urban P, Pompon D, Smit MS, Nicaud JM (2004) The use of Yarrowia lipolytica for the expression of human cytochrome P450 CYP1A1. Yeast 21:583–92. doi:10.1002/yea.1127
Otto C, Yovkova V, Aurich A, Mauersberger S, Barth G (2012) Variation of the by-product spectrum during α-ketoglutaric acid production from raw glycerol by overexpression of fumarase and pyruvate carboxylase genes in Yarrowia lipolytica. Appl Microbiol Biotechnol 95:905–917. doi:10.1007/s00253-012-4085-1
Park JN, Song Y, Cheon SA, Kwon O, Oh DB, Jigami Y, Kim JY, Kang HA (2011) Essential role of YlMPO1, a novel Yarrowia lipolytica homologue of Saccharomyces cerevisiae MNN4, in mannosylphosphorylation of N- and O-linked glycans. Appl Environ Microbiol 77:1187–1195. doi:10.1128/AEM.02323-10
Picataggio SK, Yadav NS, Zhu QQ (2003) Production of polyunsaturated fatty acids in oleaginous yeasts. European Patent Application EP2402448
Pienaar DP, Mitra RK, Deventer TIV, Botes AL (2008) Microbiological bioconversions by Yarrowia lipolytica: synthesis of a variety of optically active hydroxylated heterocyclic compounds using epoxide hydrolase technology. Tetrahed Lett 49:6752–6755. doi:10.1016/j.tetlet.2008.08.097
Pignède G, Wang H, Fudalej F, Seman M, Gaillardin C, Nicaud JM (2000) Autocloning and amplification of LIP2 in Yarrowia lipolytica. Appl Environ Microbiol 66:3283–3289. doi:10.1128/AEM.66.8.3283-3289.2000
Richard GF, Kerrest A, Lafontaine I, Dujon B (2005) Comparative genomics of hemiascomycete yeasts: genes involved in DNA replication, repair, and recombination. Mol Biol Evol 22:1011–1023. doi:10.1093/molbev/msi083
Roth R, Moodley V, van Zyl P (2009) Heterologous expression and optimized production of an Aspergillus aculeatus endo-1,4-beta-mannanase in Yarrowia lipolytica. Mol Biotechnol 43:112–120. doi:10.1007/s12033-009-9187-3
Sabirova JS, Haddouche R, Van Bogaert I, Mulaa F, Verstraete W, Timmis K, Schmidt-Dannert C, Nicaud J, Soetaert W (2011) The 'LipoYeasts' project: using the oleaginous yeast Yarrowia lipolytica in combination with specific bacterial genes for the bioconversion of lipids, fats and oils into high-value products. Microb Biotechnol 4:47–54. doi:10.1111/j.1751-7915.2010.00187
Sané S, Richter K, Rubenwolf S, Matschke NJ, Jolivalt C, Madzak C, Zengerle R, Gescher J, Kerzenmacher S (2014) Using planktonic microorganisms to supply the unpurified multi-copper oxidases laccase and copper efflux oxidases at a biofuel cell cathode. Bioresour Technol 158:231–238. doi:10.1016/j.biortech.2014.02.038
Sestric R, Munch G, Cicek N, Sparling R, Levin DB (2014) Growth and neutral lipid synthesis by Yarrowia lipolytica on various carbon substrates under nutrient-sufficient and nutrient-limited conditions. Bioresour Technol 164:41–46. doi:10.1016/j.biortech.2014.04.016
Shiningavamwe A, Obiero G, Albertyn J, Nicaud JM, Smit M (2006) Heterologous expression of the benzoate para-hydroxylase encoding gene (CYP53B1) from Rhodotorula minuta by Yarrowia lipolytica. Appl Microbiol Biotechnol 72:323–329. doi:10.1007/s00253-005-0264-7
Sibirny A, Madzak C, Fickers P (2014) Genetic engineering of non-conventional yeast for the production of valuable compounds. In: Darvishi Harzevili F, Chen H (eds) Microbial biotechnology: progress and trends. CRC Press, USA, pp 63–111. doi:10.1201/b17587-5
Song Y, Choi MH, Park JN, Kim MW, Kim EJ, Kang HA, Kim JY (2007) Engineering of the yeast Yarrowia lipolytica for the production of glycoproteins lacking the outer-chain mannose residues of N-glycans. Appl Environ Microbiol 73:4446–4454. doi:10.1128/AEM.02058-06
Song H, Zhou L, Zhang L, Gao B, Wei D, Shen Y, Wang R, Madzak C, Jiang Z (2011) Construction of a whole-cell catalyst displaying a fungal lipase for effective treatment of oily wastewaters. J Mol Catal B Enzym 71:166–170. doi:10.1016/j.molcatb.2011.04.015
Swennen D, Paul MF, Vernis L, Beckerich JM, Fournier A, Gaillardin C (2002) Secretion of active anti-Ras single-chain Fv antibody by the yeasts Yarrowia lipolytica and Kluyveromyces lactis. Microbiology 148:41–50
Tai M, Stephanopoulos G (2013) Engineering the push and pull of lipid biosynthesis in oleaginous yeast Yarrowia lipolytica for biofuel production. Metab Eng 15:1–9. doi:10.1016/j.ymben.2012.08.007
Tharaud C, Ribet AM, Costes C, Gaillardin C (1992) Secretion of human blood coagulation factor XIIIa by the yeast Yarrowia lipolytica. Gene 121:111–119. doi:10.1016/0378-1119(92)90168-O
Theerachat M, Emond S, Cambon E, Bordes F, Marty A, Nicaud JM, Chulalaksananukul W, Guieysse D, Remaud-Siméon M, Morel S (2012) Engineering and production of laccase from Trametes versicolor in the yeast Yarrowia lipolytica. Bioresour Technol 125:267–274. doi:10.1016/j.biortech.2012.07.117
Tiels P, Baranova E, Piens K, De Visscher C, Pynaert G, Nerinckx W, Stout J, Fudalej F, Hulpiau P, Tännler S, Geysens S, Van Hecke A, Valevska A, Vervecken W, Remaut H, Callewaert N (2012) A bacterial glycosidase enables mannose-6-phosphate modification and improved cellular uptake of yeast-produced recombinant human lysosomal enzymes. Nat Biotechnol 30:1225–1231. doi:10.1038/nbt.2427
van Zyl PJ (2010) A method for producing a polypeptide in Yarrowia lipolytica. World Patent Application WO2011148339
Verbeke J, Beopoulos A, Nicaud JM (2013) Efficient homologous recombination with short length flanking fragments in Ku70 deficient Yarrowia lipolytica strains. Biotechnol Lett 35:571–576. doi:10.1007/s10529-012-1107-0
Vernis L, Abbas A, Chasles M, Gaillardin C, Brun C, Huberman JA, Fournier P (1997) An origin of replication and a centromere are both needed to establish a replicative plasmid in the yeast Yarrowia lipolytica. Mol Cell Biol 17:1995–2004
Wang W, Wei H, Alahuhta M, Chen X, Hyman D, Johnson DK, Zhang M, Himmel ME (2014) Heterologous expression of xylanase enzymes in lipogenic yeast Yarrowia lipolytica. PLoS One 9:e111443. doi:10.1371/journal.pone.0111443
Wei H, Wang W, Alahuhta M, Vander Wall T, Baker JO, Taylor LE 2nd, Decker SR, Himmel ME, Zhang M (2014) Engineering towards a complete heterologous cellulase secretome in Yarrowia lipolytica reveals its potential for consolidated bioprocessing. Biotechnol Biofuels 7:148. doi:10.1186/s13068-014-0148-0
Xue Z, Sharpe P, Hong SP, Yadav N, Xie D, Short D, Damude HG, Rupert RA, Seip JE, Wang J, Pollak DW, Bostick MW, Bosak MD, Macool DJ, Hollerbach DH, Zhang H, Arcilla DM, Bledsoe SA, Croker K, McCord EF, Tyreus BD, Jackson EN, Zhu Q (2013) Production of omega-3 eicosapentaenoic acid by metabolic engineering of Yarrowia lipolytica. Nat Biotechnol 31:734–740. doi:10.1038/nbt.2622
Yang XS, Jiang ZB, Song HT, Jiang SJ, Madzak C, Ma LX (2009) Cell-surface display of the active mannanase in Yarrowia lipolytica with a novel surface-display system. Biotechnol Appl Biochem 54:171–176. doi:10.1042/BA20090222
Yang CH, Huang YC, Chen CY, Wen CY (2010) Heterologous expression of Thermobifida fusca thermostable alpha-amylase in Yarrowia lipolytica and its application in boiling stable resistant sago starch preparation. J Ind Microbiol Biotechnol 37:953–960. doi:10.1007/s10295-010-0745-2
Ye RW, Sharpe PL, Zhu Q (2012) Bioengineering of oleaginous yeast Yarrowia lipolytica for lycopene production. Methods Mol Biol 898:153–159. doi:10.1007/978-1-61779-918-1_9
Yin X, Madzak C, Du G, Zhou J, Chen J (2012) Enhanced alpha-ketoglutaric acid production in Yarrowia lipolytica WSH-Z06 by regulation of the pyruvate carboxylation pathway. Appl Microbiol Biotechnol 96:1527–37. doi:10.1007/s00253-012-4192-z
Yovkova V, Otto C, Aurich A, Mauersberger S, Barth G (2014) Engineering the α-ketoglutarate overproduction from raw glycerol by overexpression of the genes encoding NADP+-dependent isocitrate dehydrogenase and pyruvate carboxylase in Yarrowia lipolytica. Appl Microbiol Biotechnol 98:2003–2013. doi:10.1007/s00253-013-5369-9
Yu XJ, Li HJ, Li J, Chi ZM (2010a) Overexpression of acid protease of Saccharomycopsis fibuligera in Yarrowia lipolytica and characterization of the recombinant acid protease for skimmed milk clotting. Biotechnol Bioprocess Eng 15:467–475. doi:10.1007/s12257-009-3081-5
Yu XJ, Madzak C, Li HJ, Chi ZM, Li J (2010b) Surface display of acid protease on the cells of Yarrowia lipolytica for milk clotting. Appl Microbiol Biotechnol 87:669–677. doi:10.1007/s00253-010-2549-8
Yue L, Chi Z, Wang L, Liu J, Madzak C, Li J, Wang X (2008) Construction of a new plasmid for surface display on cells of Yarrowia lipolytica. J Microbiol Met 72:116–123. doi:10.1016/j.mimet.2007.11.011
Yuzbashev TV, Yuzbasheva EY, Vibornaya TV, Sobolevskaya TI, Laptev IA, Gavrikov AV, Sineoky SP (2012) Production of recombinant Rhizopus oryzae lipase by the yeast Yarrowia lipolytica results in increased enzymatic thermostability. Protein Expr Purif 82:83–89. doi:10.1016/j.pep.2011.11.014
Yuzbasheva EY, Yuzbashev TV, Konstantinova TK, Laptev IA, Sineoky SP (2010) Capacity of the N- and C-domains of the cell wall protein Flo1p homologue in yeast Yarrowia lipolytica to display lipase Lip2 on the cell surface. Biotechnologiya 6:23–36, Article in Russian
Yuzbasheva E, Yuzbashev T, Laptev I, Konstantinova T, Sineoky S (2011) Efficient cell surface display of Lip2 lipase using C-domains of glycosylphosphatidylinositol-anchored cell wall proteins of Yarrowia lipolytica. Appl Microbiol Biotechnol 91:645–654. doi:10.1007/s00253-011-3265-8
Zhang H, Zhang L, Chen H, Chen YQ, Chen W, Song Y, Ratledge C (2014) Enhanced lipid accumulation in the yeast Yarrowia lipolytica by over-expression of ATP:citrate lyase from Mus musculus. J Biotechnol 192PA:78–84. doi:10.1016/j.jbiotec.2014.10.004
Zhao CH, Cui W, Liu XY, Chi ZM, Madzak C (2010) Expression of inulinase gene in the oleaginous yeast Yarrowia lipolytica and single cell oil production from inulin-containing materials. Metab Eng 12:510–517. doi:10.1016/j.ymben.2010.09.001
Zhou J, Yin X, Madzak C, Du G, Chen J (2012) Enhanced α-ketoglutarate production in Yarrowia lipolytica WSH-Z06 by alteration of the acetyl-CoA metabolism. J Biotechnol 161:257–64. doi:10.1016/j.jbiotec.2012.05.025
Zinjarde SS (2014) Food-related applications of Yarrowia lipolytica. Food Chem 152:1–10. doi:10.1016/j.foodchem.2013.11.117
Conflict of interest
The author declares no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Madzak, C. Yarrowia lipolytica: recent achievements in heterologous protein expression and pathway engineering. Appl Microbiol Biotechnol 99, 4559–4577 (2015). https://doi.org/10.1007/s00253-015-6624-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-015-6624-z