
Abstract
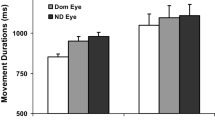
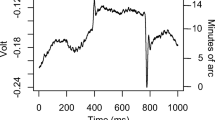
Previous research has suggested that binocular vision plays an important role in prehension. It has been shown that removing binocular vision affects (negatively) both the planning and on-line control of prehension. It has been suggested that the adverse impact of removing binocular vision is because monocular viewing results in an underestimation of target distance in visuomotor tasks. This suggestion is based on the observation that the kinematics of prehension are altered when viewing monocularly. We argue that it is not possible to draw unambiguous conclusions regarding the accuracy of distance perception from these data. In experiment 1, we found data that contradict the idea that a consistent visuomotor underestimation of target distance is an inevitable consequence of monocular viewing. Our data did show, however, that positional variance increases under monocular viewing. We provide an alternative explanation for the kinematic changes found when binocular vision is removed. Our account is based on the changes in movement kinematics that occur when end-point variance is altered following the removal of binocular vision. We suggest that the removal of binocular vision leads to greater perceptual uncertainty (e.g. less precise stimulus cues), resulting in changes in the kinematics of the movement (longer duration movements). Our alternative account reconciles some differences within the research literature. We conducted a series of experiments to explore further the issue of when binocular information is advantageous in prehension. Three subsequent experiments were employed which varied binocular/monocular viewing in selectively lit conditions. Experiment 2 explored the differences in prehension measured between monocular and binocular viewing in a full cue environment with a continuous view of the target object. Experiment 3 required participants to reach, under a monocular or binocular view, for a continuously visible self-illuminated target object in an otherwise dark room. In Experiment 3, the participant could neither see the target object nor the reaching hand following initiation of the prehension movement. Our results suggest that binocular vision contributes to prehension by providing additional information (cues) to the nervous system. These cues appear to be weighted differentially according to the particular constellation of stimulus cues available to the participants when reaching to grasp. One constant advantage of a binocular view appears to be the provision of on-line information regarding the position of the hand relative to the target. In reduced cue conditions (i.e. where a view of the target object is lost following initiation of the movement), binocular information regarding target location appears to be particularly useful in the initial programming of reach distance. Our results are a step towards establishing the specific contributions that binocular vision makes to the control of prehension.



Similar content being viewed by others

Notes
Terminal grip aperture refers to the grip aperture at the end of the transport component of prehension (as specified by velocity criterion, typically 2 cm/s). This is often larger than the width of the object. The final stage of prehension thus sometimes consists of a final ‘squeeze’ component at the end of which the fingers make contact with the object (the ‘final’ grip aperture is equal in magnitude to the object width).
References
Berberovic N, Mattingley JB (2003) Effects of prismatic adaptation on judgements of spatial extent in peripersonal and extrapersonal space. Neuropsychologia 41:493–503
Bingham GP, Pagano CC (1998) The necessity of a perception-action approach to definite distance perception: monocular distance perception to guide reaching. J Exp Psychol Hum Percept Perform 24:145–168
Dijkerman HC, Milner AD, Carey DP (1996) The perception and prehension of objects orientated in the depth plane. 1. Effects of visual form agnosia. Exp Brain Res 72:425–435
Fitts PM (1954) The information capacity of the human motor system in controlling the amplitude of movement. J Exp Psychol 47:381–391
Foley JM (1977) Effect of distance information and range on two indices of visually perceived distance. Perception 6:449–460
Harris CM, Wolpert DM (1998) Signal-dependent noise determines motor planning. Nature 394:780–784
Jackson SR, Husain M (1996) Visuomotor functions of the lateral pre-motor cortex. Curr Opin Neurobiol 6:788–795
Jackson SR, Jones CA, Newport R, Pritchard C (1997) A kinematic analysis of goal-directed prehension movements executed under binocular, monocular and memory-guided viewing conditions. Vis Cogn 4:113–142
Landy MS, Maloney LT, Johnston ET, Young M (1995) Measurement and modeling of depth cue combination: in defense of weak fusion. Vision Res. 35:389–412
Marotta JJ, Goodale MA (1998) The role of learned pictorial cues in the programming and control of grasping. Exp Brain Res 121:465–470
Marotta JJ, Behrmann M, Goodale MA (1997) The removal of binocular cues disrupts the calibration of grasping in patients with visual form agnosia. Exp Brain Res 116:113–121
Michel C, Rossetti Y, Rode G, Tilikete C (2002) After effects of visuo-manual adaptation to prisms on body posture in normal subjects. Exp Brain Res 148:219–226
Mon-Williams M, Dijkerman HC (1999) The use of vergence information in the programming of prehension. Exp Brain Res 128:578–582
Mon-Williams M, Tresilian J (1999) A review of some recent studies on the extra-retinal contribution to distance perception. Perception 28:167–181
Mon-Williams M, Tresilian JR, Plooy A, Wann JP, Broerse J (1997) Looking at the task in hand: vergence eye movements and perceived size. Exp Brain Res 117:501–506
Mon-Williams M, Tresilian JR, McIntosh RD, Milner D (2001) Monocular and binocular distance cues: insights from visual form agnosia I (of III). Exp Brain Res 139:127–136
Pagano CC, Bingham GP (1998) Comparing measures of monocular distance perception: verbal and reaching errors are not correlated. J Exp Psychol Hum Percept Perform 24:1037–1051
Redding GM, Wallace B (1992) Adaptive hand-eye coordination: implications of prism adaptation for perceptual motor organization. In: Proteau L, Elliot D (eds) Vision and motor control. Amsterdam, Elsevier, pp 105–127
Redding GM, Wallace B (1997) Prism adaptation during pointing from visible & non-visible starting locations. J Mot Behav 29:119–130
Servos P (2000) Distance estimation in the visual and visuomotor systems. Exp Brain Res 130:35–47
Servos P, Goodale MA (1994) Binocular vision and the on-line control of human prehension. Exp Brain Res 54:121–132
Servos P, Goodale MA, Jakobson LS (1992) The role of binocular vision in prehension: a kinematic analysis. Vision Res 32:1513–1521
Van Opstal AJ, van Gisbergen JAM (1987) Skewness of saccadic velocity profiles: a unifying parameter for normal and slow saccades. Vision Res 5:731–745
Wallace SA, Weeks DL (1988) Temporal constraints on the control of prehensile movement. J Mot Behav 20:81–105
Watt SJ, Bradshaw MF (2000) Binocular cues are important in controlling the grasp but not the reach in natural prehension movements. Neuropsychologia 28:1473–1481
Wing AM, Turton A, Fraser C (1986) Grasp size and accuracy of approach in reaching. J Mot Behav 3:245–260
Wolpert DM, Miall CR, Kawato M (1998) Internal models in the cerebellum. Trends in Cognitive Science 2:338–347
Acknowledgements
The research was funded in part by a grant from the Natural Sciences and Engineering Research Council of Canada to M.A. Goodale and P. Servos.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Loftus, A., Servos, P., Goodale, M.A. et al. When two eyes are better than one in prehension: monocular viewing and end-point variance. Exp Brain Res 158, 317–327 (2004). https://doi.org/10.1007/s00221-004-1905-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-004-1905-2