
Abstract
Aims/hypothesis
Evidence suggests that bacterial components in blood could play an early role in events leading to diabetes. To test this hypothesis, we studied the capacity of a broadly specific bacterial marker (16S rDNA) to predict the onset of diabetes and obesity in a general population.
Methods
Data from an Epidemiological Study on the Insulin Resistance Syndrome (D.E.S.I.R.) is a longitudinal study with the primary aim of describing the history of the metabolic syndrome. The 16S rDNA concentration was measured in blood at baseline and its relationship with incident diabetes and obesity over 9 years of follow-up was assessed. In addition, in a nested case–control study in which participants later developed diabetes, bacterial phylotypes present in blood were identified by pyrosequencing of the overall 16S rDNA gene content.
Results
We analysed 3,280 participants without diabetes or obesity at baseline. The 16S rDNA concentration was higher in those destined to have diabetes. No difference was observed regarding obesity. However, the 16S rDNA concentration was higher in those who had abdominal adiposity at the end of follow-up. The adjusted OR (95% CIs) for incident diabetes and for abdominal adiposity were 1.35 (1.11, 1.60), p = 0.002 and 1.18 (1.03, 1.34), p = 0.01, respectively. Moreover, pyrosequencing analyses showed that participants destined to have diabetes and the controls shared a core blood microbiota, mostly composed of the Proteobacteria phylum (85–90%).
Conclusions/interpretation
16S rDNA was shown to be an independent marker of the risk of diabetes. These findings are evidence for the concept that tissue bacteria are involved in the onset of diabetes in humans.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The incidence of diabetes, in developed and emerging countries, has reached epidemic proportions [1]. A body of evidence demonstrates that the intestinal microbiota, which correspond to the overall bacterial community present in the intestine, play a role in the onset of metabolic disease [2–4]. The causal role of the intestinal microbiota in weight gain has been demonstrated in animal models [2]. In humans, obesity is associated with changed phylum levels in the gut microbiota [3]. In addition, the role of bacterial components in blood has also been demonstrated: thus, mice chronically infused with a low dose of lipopolysaccharides developed inflammation and diabetes [4]. Importantly, the plasma lipopolysaccharides concentration has been found to predict the onset of diabetes [5]. From these data, we hypothesised that the presence of a blood microbiome could be one of the initial steps leading to diabetes and obesity. To test this hypothesis, we investigated whether the 16S rDNA concentration in blood could be a marker of the risk of diabetes and obesity in a large general population. We studied this gene as it is highly conserved between different species of bacteria and is hence considered to be a marker of the overall microbiota [6]. Moreover, by pyrosequencing the overall 16S rDNA gene population in blood, we determined the profile of bacteria present in individuals who were destined to develop diabetes versus those present in controls.
Methods
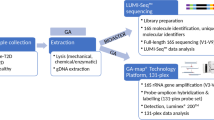
Study overview Data from an Epidemiological Study on the Insulin Resistance Syndrome (D.E.S.I.R.) is a longitudinal cohort study of 5,212 adults aged 30 to 65 years at baseline; the primary aim of the study was to describe the natural history of the metabolic syndrome [7]. Participants were recruited between 1994 and 1996 from volunteers insured by the French national social security system in ten social security health examination centres. All participants gave written informed consent and the study protocol was approved by the Comité Consultatif de Protection des Personnes pour la Recherche Biomédicale of the Hôpital Bicêtre (Paris, France). Participants were clinically and biologically evaluated at inclusion and at follow-up visits in years 3, 6 and 9. We measured the baseline 16S rDNA concentration in individuals without diabetes (defined by treatment for diabetes or fasting plasma glucose ≥ 7.0 mmol/l) or obesity (BMI ≥ 30 kg/m2) at baseline and in those who had a known diabetes status at the year 9 examination. We excluded those who were likely to have a current infection (C-reactive protein > 10 mg/l, abundant leucocyturia) or who were taking antiviral therapy.
Variables studied
Weight and height were measured in lightly clad participants and BMI was calculated. Waist circumference, i.e. the smallest circumference between the lower ribs and the iliac crests, was also measured. The examining physician noted the existence of a family history of diabetes, and treatments for diabetes and hypertension were recorded. Hypertension was defined as a systolic/diastolic blood pressure of at least 140/90 mmHg or taking antihypertensive medication. Smoking habits were documented in a questionnaire filled in by the participants. Central adiposity was defined as a waist circumference ≥102 cm in men and ≥88 cm in women, while high fasting glucose was defined as ≥ 6.1 mmol/l [8].
Biological analyses
Blood was drawn after a 12 h fast. The methods used for biological analyses, except for bacterial DNA preparation and analysis, have been described elsewhere [7].
Bacterial DNA preparation
DNA was extracted from peripheral blood leucocytes using a classical phenol/chloroform extraction method followed by alcohol precipitation (ice-cold 70% alcohol by vol.). Air-dried DNA was resuspended in TRIS EDTA and stored at −80°C before use at a concentration of 1 μg/μl [9, 10]. It is noteworthy that this method did not use glass microbeads to increase the efficiency of bacterial DNA extraction, as commonly done in current methods. The reason for this was that isolation of bacterial DNA was not the goal of the work at the time of the sample preparation. However, despite this important technical difference, we succeeded in amplifying 16S rDNA from blood. To validate the difference between both extraction procedures, we extracted total blood DNA in the presence or absence of microbeads. The efficacy of bacterial DNA extraction, as assessed by 16S rDNA quantitative PCR amplification, was ten times greater when using microbeads. Although the absolute value of the bacterial DNA concentration was higher, the proportion between samples remained the same.
Quantification of 16S rDNA
The total DNA concentration was determined using a broad-range assay kit (Quant-iT dsDNA; Life Technologies, Villebon-sur-Yvette, France). The mean concentration ± SD was 121.1 ± 208.3 ng/μl. Each sample was diluted tenfold in TRIS buffer EDTA. The DNA was amplified by real-time PCR (Life Technologies) in optical-grade 96-well plates. The PCR reaction was performed in a total volume of 25 μl using a master mix (Power SYBR Green PCR; Life Technologies) containing 300 nmol/l of each of the universal forward and reverse primers for Eubact: forward 5′-TCCTACGGGAGGCAGCAGT-3′ and reverse 5′-GGACTACCAGGGTATCTAATCCTGTT-3′. The reaction conditions for amplification of DNA were 95°C for 10 min, and 35 cycles at 95°C for 15 s and at 60°C for 1 min. The amplification step was followed by a melting curve step according to the manufacturer’s instructions (from 60°C to 90°C) to determine the specificity of the amplification product obtained. The amount of amplified DNA was determined using a 16S rDNA standard curve obtained by real-time PCR from dilutions ranging from 0.001 to 10 ng/μl of E. coli BL21 DNA.
Identification of bacterial sequences
This sub-study included 14 participants with incident diabetes at year 9, but who had been at low risk of diabetes at baseline, having been non-smokers with a waist perimeter of < 85 cm for men and < 75 cm for women. These 14 participants were matched for age (±3 years), sex, waist circumference (±3 cm) and fasting blood glucose (±0.3 mmol/l) with 28 control participants who remained free of diabetes over the entire follow-up period. The V1–V2 variable region of the 16S rDNA gene was amplified by PCR for each participant, and amplicons from case and control participants were pooled separately after size verification by 2% agarose gel electrophoresis. Each pool was assigned to specific multiplex identifiers (MIDs) that were used as tags. These two pools of amplicons were pyrosequenced using a genome sequencer (454 Life Sciences; Branford, CT, USA) on a multiplexed pyrosequencing run (GS FLX-Ti; Life Technology). Sequences were binned for removal of pyrosequencing MIDs and PCR primers (forward: AG-AGT-TTG-ATC-MTG-GCT-CAG; reverse: GC-TGC-CTC-CCG-TAG-GAG-T) [2, 3]. Meta_RNA software, designed for the identification of 16S rRNA sequences based on hidden Markov models, was further applied to select prokaryotic 16S rRNA sequences [11]. The resulting 16S rRNA sequences were assigned to taxonomies using mothur software [12] and the SILVA reference taxonomic outline [13].
Bioinformatic analyses
Of the 919,095 and 768,245 sequence reads recovered for case and control participants respectively, ∼30% were removed by initial quality filters. A de-noising step was performed to remove most of the common sequencing errors encountered with the 454 platform by clustering chimeric sequences as well as reads that were most likely to have been derived from the same sequence.
Statistical analyses
For statistical analysis, in order to carry out parametric tests, the 16S rDNA concentrations were log-transformed, as the distribution was skewed (as were levels of triacylglycerol and insulin). The characteristics of the participants who did and did not have incident diabetes over follow-up are shown and compared by t and χ² tests. Note that log-transformed 16S rDNA concentrations were compared statistically. For incident diabetes and abdominal adiposity, logistic regression was used to calculate the standardised odds ratios and 95% CIs for an increase of one SD of baseline 16S rDNA concentration as a continuous variable (logarithm). Adjustments were made for sex, baseline age, family history of diabetes, hypertension, waist circumference, BMI, smoking status and fasting plasma glucose. The relationship with 16S rDNA concentration was linear, as an additional squared term was not significant. ORs were also calculated over risk factor strata. The C statistic was used to quantify the discriminative ability of 16S rDNA and that of various risk factors for diabetes. SAS versions 9.1 and 9.2 (Cary, NC, USA) were used for statistical analysis.
Results
Characteristics of the population studied At baseline, of the 5,212 participants in the D.E.S.I.R. study, 126 had diabetes, 474 were obese, 65 had biological signs of infection or were taking antiviral therapy, and 333 did not undergo 16S rDNA concentration determination. For 1,146 participants, the diabetes status was not known at the end of the 9 years, as they did not attend the year 9 examination. These volunteers were excluded from the analysis. By comparison, the participants analysed (n = 3,280) were older and fewer were current smokers. There was no significant difference in baseline 16S rDNA gene concentrations, waist circumference or fasting plasma glucose. The characteristics of the study population are shown in Table 1; 131 incident cases of diabetes were recorded. The mean 16S rDNA concentration was higher in those with incident diabetes (Table 2) and the distribution of log-transformed 16S rDNA concentration was shifted to the right (Fig. 1). No difference in baseline blood 16S rDNA concentration was observed in participants destined to become obese. In contrast, mean baseline 16S rDNA concentration (log-transformed) tended to be higher (mean ± standard deviation −2.73 ± 1.04 vs −2.63 ± 1.04; n = 485; p = 0.05) in those who were to present with abdominal adiposity after 9 years of follow-up. A bivariate analysis between blood bacterial DNA and other biological variables showed a modest negative correlation with fasting blood glucose, while a positive correlation with fibrinogen and leucocyte count was observed (Table 3).

Distribution of participants destined to become diabetic (continuous line) and those who did not (dotted line) according to baseline blood 16S rDNA gene concentration
Prediction of diabetes and abdominal adiposity
The 16S rDNA concentration predicted the onset of diabetes after adjustment for confounding factors, with a standardised OR of 1.29 (95% CI 1.08, 1.55) after adjustment for age, sex and fasting blood glucose, and of 1.35 (95% CI 1.11, 1.64) in a fully adjusted model (Table 3). While higher 16S rDNA concentrations appeared to carry more risk in women and in those with lower fasting glucose, there was no significant difference between strata for the risk of incident diabetes between bacterial DNA and any of these strata (Table 4). We also analysed the risk of developing diabetes by follow-up time. The OR of developing diabetes was higher in participants who developed diabetes within the 6 to 9 year window (OR 1.61, 95% CI 1.23, 2.10; p = 0.0005) than in those who developed diabetes soon after the initial measurement, i.e. in the 0 to 3 year window (OR 1.10, 95% CI 0.87, 1.39; p = 0.45). On its own, 16S rDNA as a marker carried little discriminative ability between those who did and did not develop incident diabetes, with a C statistic of 0.564; however, when the level of 16S rDNA was added to a predictive score for diabetes derived in the D.E.S.I.R. population [14], the C statistic increased significantly from 0.862 to 0.871 (p = 0.02) and 16S rDNA was still predictive of diabetes after adjustment for this score (p = 0.004)
The 16S rDNA concentration also predicted the presence of abdominal adiposity at the end of 9 years, after adjustment for confounders, with a standardised OR of 1.18 (95% CI 1.03, 1.34; p = 0.01).
Identification of blood bacteria phyla in participants with incident diabetes
The 16S rDNA gene was sequenced in the pooled baseline DNA samples from participants destined to become diabetic and from controls. We first analysed the alpha diversity, which corresponds to groups of sequences characterised by less than 10% differences. Each group of similar sequences defines an operational taxonomic diversity unit (OTU). The number of OTUs (at 10%) was 745 and 800 in cases and controls, respectively (see electronic supplementary material [ESM] Fig. 1). A common core of 443 OTUs was identified. At the phylum level, proteobacteria represented 80% to 90% of all phyla in the blood, both in cases and controls (Fig. 2). Within the Proteobacteria phylum, at a genera level, the Ralstonia genus was the most prevalent (see ESM Fig. 2).

Abundance of bacterial phyla for those who developed diabetes (black bars) and controls without diabetes over the entire follow-up period (white bars)
Discussion
We show, for the first time, that the blood concentration of a bacterial gene predicts the onset of diabetes, particularly 6 to 9 years after baseline, and also predicts abdominal adiposity in a large sample of non-obese participants from a general population. We found that more than 90% of this bacterial DNA belonged to the Proteobacteria phylum.
The involvement of inflammation in the development of insulin resistance [14–16] and the pro-inflammatory effects of bacterial components present within tissues [4] suggest that the role played by tissue bacteria in relation to diabetes and its complications should be explored. Indeed, experimental data have already linked the metabolic syndrome with gut microbiota and innate immunity against infectious diseases [3, 4]. We [4, 17, 18] and others [3] have demonstrated the influence of gut microbiota on metabolic disease in animal models. Consistently, in humans, periodontitis, a chronic gram-negative infectious disease of the oral cavity, has been associated with the metabolic syndrome [19–23]. In line with these data, the longitudinal study reported here provides for the first time evidence of the involvement of tissue microbiota in the development of diabetes in humans.
These epidemiological data also provide some insight into the mechanisms of action of the microbiota. Indeed, although no difference was observed in blood microbiota composition in participants destined to develop diabetes and in controls, possibly because of a lack of statistical power, we did establish the predominance of proteobacteria, which represent 90% of overall microbiota. In this respect, the role of lipopolysaccharides, a major component of proteobacteria, in the onset of diabetes has already been demonstrated [3, 5]. Interestingly, in faeces, this phylum is in the minority, since Bacteroidetes and Firmicutes are the most predominant phyla [24], suggesting selectivity of the filtering mechanism, which allows only some bacteria to be present in the blood [17]. From these data, it seems possible that proteobacteria play a role in the onset of metabolic diseases. Our results show at baseline a negative correlation between bacterial DNA levels and fasting blood glucose. At this stage, we cannot rule out a transitory improvement in insulin sensitivity in the early phase of metabolic infection, as shown in an experimental model of gram-negative sepsis in healthy humans [25, 26]. This could be due to the release of nitric oxide during latent infection, which would in turn improve insulin sensitivity [27, 28]. On the other hand, the exclusion of patients with treated or non-treated diabetes at baseline, i.e. participants in whom bacteraemia would already have had a deleterious influence on glucose metabolism, is more likely to have contributed to this association. Obviously, the present study is observational in nature and thus cannot demonstrate the causative role played by blood microbiota in the onset of diabetes. Indeed, the blood bacterial DNA burden may only represent an innocent bystander in the disease process. Moreover, these results are preliminary and need to be replicated in other longitudinal data sets.
In conclusion, our results show for the first time that blood microbiota are a marker of the risk of diabetes and abdominal adiposity in a general population. These findings establish evidence for the concept of an involvement of tissue bacteria in the onset of diabetes in humans.
Abbreviations
- D.E.S.I.R:
-
Data from an Epidemiological Study on the Insulin Resistance Syndrome
- MID:
-
Multiplex identifier
- OTU:
-
Operational taxonomic diversity unit
References
Shaw JE, Sicree RA, Zimmet PZ (2010) Global estimates of the prevalence of diabetes for 2010 and 2030. Diabetes Res Clin Pract 87:4–14
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI (2006) An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444:1027–1031
Turnbaugh PJ, Hamady M, Yatsunenko T et al (2009) A core gut microbiome in obese and lean twins. Nature 457:480–484
Cani PD, Amar J, Iglesias MA et al (2007) Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 56:1761–1772
Pussinen PJ, Havulinna AS, Lehto M, Sundvall J, Salomaa V (2011) Endotoxemia is associated with an increased risk of incident diabetes. Diabetes Care 34:392–397
Coenye T, Vandamme P (2003) Intragenomic heterogeneity between multiple 16S ribosomal RNA operons in sequenced bacterial genomes. FEMS Microbiol Lett 228:45–49
Fumeron F, Aubert R, Siddiq A et al (2004) Adiponectin gene polymorphisms and adiponectin levels are independently associated with the development of hyperglycemia during a 3-year period: the epidemiologic data on the insulin resistance syndrome prospective study. Epidemiologic Data on the Insulin Resistance Syndrome (DESIR) Study Group. Diabetes 53:1150–1157
Grundy SM, Cleeman JI, Daniels SR et al (2005) American Heart Association; National Heart, Lung, and Blood Institute. Diagnosis and management of the metabolic syndrome: an American Heart Association/National Heart, Lung, and Blood Institute Scientific Statement. Circulation 112:2735–2752
Rigat B, Hubert C, Alhenc-Gelas F, Cambien F, Corvol P, Soubrier F (1990) An insertion/deletion polymorphism in the angiotensin I-converting enzyme gene accounting for half the variance of serum enzyme levels. J Clin Invest 86:1343–1346
Marcadet A, O’Connell P, Cohen D (1987) Standardized Southern blot workshop technique. In: Dupont B (ed) Histocompatibilty testing, vol 1. Springer, New York, pp 553–560
Huang Y, Gilna P, Li W (2009) Identification of ribosomal RNA genes in metagenomic fragments. Bioinformatics 25:1338–1340
Schloss PD, Westcott SL, Ryabin T et al (2009) Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol 75:7537–7541
Pruesse E, Quast C, Knittel K et al (2007) SILVA: a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res 35:7188–7196
Balkau B, Lange C, Fezeu L et al (2008) Predicting diabetes: clinical, biological, and genetic approaches: data from the Epidemiological Study on the Insulin Resistance Syndrome (DESIR). Diabetes Care 31:2056–2061
Festa A, D’Agostino R Jr, Howard G, Mykkänen L, Tracy RP, Haffner SM (2000) Chronic subclinical inflammation as part of the insulin resistance syndrome: the Insulin Resistance Atherosclerosis Study (IRAS). Circulation 102:42–47
Mauras N, Delgiorno C, Kollman C et al (2010) Obesity without established comorbidities of the metabolic syndrome is associated with a proinflammatory and prothrombotic state, even before the onset of puberty in children. J Clin Endocrinol Metab 95:1060–1068
Amar J, Chabo C, Waget A et al (2011) Intestinal mucosal adherence and translocation of commensal bacteria at the early onset of type 2 diabetes: molecular mechanisms and probiotic treatment. EMBO Mol Med 3:559–572
Cani PD, Bibiloni R, Knauf C et al (2008) Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 57:1470–1481
Xu H, Barnes GT, Yang Q et al (2003) Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Invest 112:1821–1830
D’Aiuto F, Sabbah W, Netuveli G et al (2008) Association of the metabolic syndrome with severe periodontitis in a large U.S. population-based survey. J Clin Endocrinol Metab 93:3989–3994
Benguigui C, Bongard V, Ruidavets JB et al (2010) Metabolic syndrome, insulin resistance, and periodontitis: a cross-sectional study in a middle-aged French population. J Clin Periodontol 37:601–608
Timonen P, Niskanen M, Suominen-Taipale L, Jula A, Knuuttila M, Ylöstalo P (2010) Metabolic syndrome, periodontal infection, and dental caries. J Dent Res 89:1068–1073
Morita T, Yamazaki Y, Mita A et al (2010) A cohort study on the association between periodontal disease and the development of metabolic syndrome. J Periodontol 81:512–519
Qin J, Li R, Raes J et al (2010) A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464:59–65
Agwunobi AO, Reid C, Maycock P, Little RA, Carlson GL (2000) Insulin resistance and substrate utilization in human endotoxemia. J Clin Endocrinol Metab 85:3770–3778
van der Crabben SN, Blümer RM, Stegenga ME et al (2009) Early endotoxemia increases peripheral and hepatic insulin sensitivity in healthy humans. J Clin Endocrinol Metab 94:463–468
Perreault M, Marette A (2001) Targeted disruption of inducible nitric oxide synthase protects against obesity-linked insulin resistance in muscle. Nat Med 10:1138–1143
Duplain H, Burcelin R, Sartori C et al (2001) Insulin resistance, hyperlipidemia and hypertension in mice lacking endothelial nitric oxide synthase. Circulation 104:342–345
Acknowledgements
This work was funded by the Agence Nationale de la Recherche (French national research agency) (to R. Burcelin) and by the Société Francophone de Diabète (to B. Balkau). The D.E.S.I.R. study has been supported by Inserm contracts with CNAMTS, Lilly, Novartis Pharma and sanofi-aventis; by Inserm (Réseaux en Santé Publique, Interactions entre les déterminants de la santé, Cohortes Santé TGIR 2008), the Association Diabète Risque Vasculaire, the Fédération Française de Cardiologie, La Fondation de France, ALFEDIAM, ONIVINS, Société Francophone du Diabète, Ardix Medical, Bayer Diagnostics, Becton Dickinson, Cardionics, Merck Santé, Novo Nordisk, Pierre Fabre, Roche, Topcon.
Contribution statement:
JA co-designed the study, contributed to the statistical analyses and interpretation of data, and wrote the first draft of the manuscript. RB co-designed the study, conducted the biological analyses, and contributed to interpretation of data and the writing of the manuscript. BB (principal investigator of the DESIR study) conducted statistical analyses, and contributed to the interpretation of data and to the writing of the manuscript. MS, LP and PK carried out biological analyses, and contributed to the interpretation of data and to the writing of the manuscript. CL carried out statistical analyses, and contributed to interpretation of data and to the writing of the manuscript. CC carried out biological analyses, and contributed to interpretation of data and to the writing of the manuscript. JI carried out bioinformatic analyses, and contributed to interpretation of data and to the writing of the manuscript. SM, PL, CK, JM, OB and JD contributed to pyrosequencing analyses, interpretation of data and the writing of the manuscript. MC and MAC contributed to the interpretation of data and the writing of the manuscript. MM created the DNA collection of the D.E.S.I.R. study and contributed to interpretation of data and to the writing of the manuscript. OL researched data, and contributed to interpretation of data and the writing of the manuscript. All authors approved the final version of the manuscript to be published.
Duality of interest
J. Amar, R. Burcelin and M. Courtney are major shareholders of VAIOMER, a biotechnology company in the field of metabolic diseases. The remaining authors declare that they have no duality of interest associated with this manuscript.
Author information
Authors and Affiliations
Consortia
Corresponding authors
Additional information
J. Amar and M. Serino contributed equally to this study.
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM Fig. 1
Venn diagram at 10% differences showing the numbers of common (shaded) and different operational taxonomic diversity units (OTUs), specific for those who developed diabetes 6 to 9 years after inclusion and controls without diabetes over the entire follow-up period (PDF 3,440 kb)
ESM Fig. 2
Abundance of bacterial genera within Proteobacteria phylum for those patients who developed diabetes 6 to 9 years after inclusion (black bars) and controls without diabetes over the entire follow-up period (white bars) (PDF 23 kb)
Rights and permissions
About this article
Cite this article
Amar, J., Serino, M., Lange, C. et al. Involvement of tissue bacteria in the onset of diabetes in humans: evidence for a concept. Diabetologia 54, 3055–3061 (2011). https://doi.org/10.1007/s00125-011-2329-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-011-2329-8