
Abstract
Conodonts and re-deposited shallow-water benthic foraminifera co-occur in the Upper Norian-Rhaetian Slatnik Formation of the Slovenian Basin (Southern Alps). The Mt. Kobla and Mt. Slatnik sections were investigated in order to give the first direct correlation between these two groups. The Norian-Rhaetian boundary (NRB) is placed at the First Appearance Datum of the conodont Misikella posthernsteini and is approached with the First Occurrence of the foraminifer Involutina turgida. An association of Trocholina turris with duostominids is a good marker for the Rhaetian. Careful revision of literature combined with our data culminated in reambulation of stratigraphical ranges for several foraminiferal species. The foraminiferal Triassic-Jurassic boundary (TJB) can be defined with the Last Occurrence of Galeanella tollmanni, “Sigmoilina” schaeferae or duostominids. The absence of the uppermost Rhaetian Misikella ultima conodont zone and the close coincidence of foraminiferal and conodont TJB could point to the presence of hiati in the upper part of the Slatnik Formation. Consequently, the exact pattern of disappearance of Late Triassic foraminifera at the boundary could not be observed. It is clear, however, that the foraminiferal assemblage was decimated for reef-dwelling foraminifera, duostominids and most of the platform-dwelling foraminifera. Biotic changes at the TJB are best explained as a result of a biocalcification crisis.
Similar content being viewed by others


1 Introduction
Fossils present the most common and also the most powerful tool for correlation of different stratigraphic sections. Because not every fossil group is present in each section, it is first necessary to establish the correlation between different fossil groups. This can be done only on the basis of the co-occurrence of two (or more) fossil groups in common sections. The problem arises when different organisms occupy completely different environments and the presence of one group often excludes the presence of the other. This is also the case for the Rhaetian: while correlations among the deep-water biota are well established (e.g. Kozur and Mostler 1973; Kozur and Mock 1991; Carter 1993; Gullo 1996; Sephton et al. 2002; Kozur 2003; Carter and Orchard 2004; Muttoni et al. 2004; Ward et al. 2004; Bertinelli et al. 2005; Krystyn and Kuerschner 2005; Rigo et al. 2005; Gallet et al. 2007; Krystyn et al. 2007a, b; Tanner et al. 2007; Williford et al. 2007; Giordano et al. 2008; McRoberts et al. 2008; Krystyn 2008; Rigo et al. 2008, 2009; Giordano et al. 2010; Muttoni et al. 2010; Lucas 2010a; Hüsing et al. 2011), it is mostly not possible to link shallow water biota with deeper living groups. Shallow water carbonate platforms were widespread during the Late Triassic (e.g. Haas 2004; Vlahović et al. 2005; Golonka 2007; Haas et al. 2007) and their stratigraphy is mostly based on algae and benthic foraminifera (Grgasović 1997; Vachard and Fontaine 1988; Barattolo and Romano 2005; Jadoul et al. 2005; Mancinelli et al. 2005; Velić 2007; Rigaud et al. 2010). To date, these two groups have been correlated only to palynomorphs (Peybernes et al. 1988; Haas et al. 1997) and rarely to groups living in deeper waters, i.e. conodonts or ammonoids (Gaździcki and Michalik 1980). The Norian-Rhaetian boundary (NRB) is presently based on conodonts (Kozur 1996; Orchard and Tozer 1997; Krystyn et al. 2007a, b; Krystyn 2008; McRoberts et al. 2008; Rožič et al. 2009; Giordano et al. 2010; Lucas 2010b) and the lack of correlation with benthic foraminifera or green algae results in limited stratigraphic resolution and dubious placements of the age/stage boundaries within shallow-water depositional systems.
We wish to further pursue the correlation of Rhaetian shallow-water benthic foraminifera from the reef and platform areas with the conodonts. We present data from the Upper Norian–lowermost Jurassic Slatnik Formation from the Mt. Kobla and Mt. Slatnik sections (eastern Southern Alps, western Slovenia). The formations consist of calciturbidites that contain rich, predominantly reef-derived foraminiferal assemblages and hemipelagic limestones with conodonts. Hemipelagites predominate in the Mt. Kobla section, which is the more distal of the two sections, and they are subordinate in the more proximal Mt. Slatnik section (Rožič et al. 2009). Consequently, a more precise conodont stratigraphy is established for the Mt. Kobla section, while the conodont data from the Mt. Slatnik section are more robust, but still provide vital information. Both sections contain rich shallow-water benthic foraminiferal assemblages, but the stratigraphically important species Triasina hantkeni and Involutina turgida were found only in the Mt. Slatnik section. Integrated biostratigraphy from both sections culminates in the strongly improved conodont resolution given by Rožič et al. (2009) and also introduces a precise biostratigraphical distribution of Foraminifera.
The aims of this paper are:
-
to provide the first account of the complete foraminiferal assemblage for the Sevatian to Rhaetian period from the Slovenian Basin;
-
to re-evaluate and supplement conodont data from the Mt. Kobla section given by Rožič et al. (2009);
-
to make the direct correlation between the conodont Sevatian to Rhaetian biozones with benthic foraminifera from the same sections;
-
to discuss a significant change in benthic foraminiferal assemblage at the Triassic-Jurassic boundary.
2 Previous research
Though several papers have shown the precise distribution of foraminiferal species in sections (e.g., Gaździcki 1974, 1983; Oravecz-Scheffer 1987), there are few correlations with other groups. Peybernes et al. (1988) and Haas et al. (1997) showed the distribution of palynomorphs parallel to foraminifera. The section by Haas et al. (1997) includes the NRB, but control with sporomorphs was not possible across this boundary. Gaździcki and Michalik (1980) found conodonts and ammonoids, as well as some shallow water biota, in the same section as foraminifera, but their sections comprise only the Rhaetian. Pálfy et al. (2007) and Haas et al. (2010) used the Last Occurrence (LO) of Triassic foraminifera as one of the proxies for the TJB but did not give detailed information on the distribution of species. Bertinelli et al. (2005) and Galli et al. (2005) instead provided a full list of taxa from near the TJB in the Lagonegro Basin, but foraminifera were grouped into a single assemblage and their bed-by-bed occurrence is not known. Michalík et al. (2007) used foraminifera to locate the TJB in the Zliechov Basin in the Western Carpathians. Although their assemblage has low diversity (only nine genera) due to the restricted environment, they made an important step towards the correlation with deep-water sections by drawing a carbon isotopic curve alongside the foraminiferal distribution. Hillebrandt (2008) traced foraminifera across the TJB in the Kuhjoch section with well-known ammonoid, palynomorph and isotopic data, but unfortunately, only the families Oberhauserellidae and Duostominidae are present in the section.
3 Geological setting
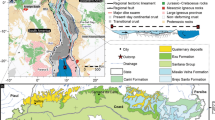
The Mt. Kobla and Mt. Slatnik sections are located in the eastern part of the Bohinj Range in the Julian Alps (NW Slovenia; Fig. 1a, b). Structurally, the Julian Alps represent an eastern continuation of the Southern Alps and consist of the Tolmin Nappe and the overlying Julian Nappe (Buser 1986; Placer 1999, 2008). The studied sections belong to the Kobla Nappe, a structural subunit of the Tolmin Nappe (Fig. 1c).

Geographic and palaeogeographic position of investigated area. a, b Position of the studied sections. c Structural map of the area in Fig. a (after Buser 1986; and M. Demšar personal communication, 2010) d, e Palaeogeographic position of the Slovenian Basin during the Late Triassic. d Simplified after Golonka (2007). e Simplified after Haas et al. (1995)
During the Triassic, the Southern Alps belonged to the southern passive continental margin of the Meliata branch of the Neotethys Ocean (Haas et al. 1995; Schmid et al. 2008) (Fig. 1d). The studied Late Norian-Rhaetian Slatnik Formation was deposited in the marginal part of the deep water Slovenian Basin, adjacent to the shallow water Julian Carbonate Platform (Rožič et al. 2009) (Fig. 1e). At Mt. Kobla, the formation is composed of hemipelagic limestones, intercalated with calciturbidites; i.e. graded calcarenites and limestone breccias. The succession was deposited in the basin plain to the lower slope environment. At Mt. Slatnik, the more proximal succession consists of resedimented limestones and only sporadic hemipelagites, characterizing deposition on the lower to upper slope sedimentary environment. Although the formation exhibits apparent differences between both sections, the main progradational/retrogradational sedimentary trends are well recognised and correlated (Rožič et al. 2009). Due to the absence of the uppermost Rhaetian Misikella ultima Zone in the upper part of the Slatnik Formation and facies characteristics (i.e. numerous channel structures, coarse-grained resediments), the upper part of the sections possibly contains hiata (H. J. Gawlick and L. Krystyn, 2011, pers. comm.; Goričan et al., 2012).
4 Materials and methods
The first detailed investigation of the Slatnik Formation by Rožič et al. (2009) focused on the facies analysis that was supported by conodont dating. For this study, the hemipelagic limestones of both sections were independently investigated and sampled. At Mt. Kobla, an additional quantity of rock was processed and supplemented with 30 new samples for the interval between 81.5 and 95 m. Fourteen additional samples were processed from the Mt. Slatnik section and eight yielded important supplementary datations; however, only one sample was taken from hemipelagic limestone. The average weight of the samples was 2 kg, with the largest sample weighing 4 kg. A standard technique to recover conodonts was applied, using diluted acetic acid followed by heavy liquid separation. The illustrated conodont elements presented herein were photographed with the JEOL JSM 6490LV Scanning Electron Microscope at the Geological Survey of Slovenia. The taxonomic terminology of the genus Misikella and Epigondolella of Kozur and Mock (1991) and Orchard (1991) was applied in the paper by Rožič et al. (2009) and is retained herein. The conodont biostratigraphy follows Kozur and Mock (1991) and Kozur (2003). All conodont samples are in the depository of the Geological Survey of Slovenia.
Foraminifera from the Mt. Kobla section were determined from 85 thin sections, 47 × 28 mm and 75 × 49 mm in size, sampling 43 beds. From the Mt. Slatnik section, 48 thin sections from 38 beds were made. The thin sections are stored at the Geological Survey of Slovenia.
Before discussing the stratigraphic value of foraminifera from the Slovenian Basin, we tried to make a revision of the literature data on stratigraphic ranges of the species in assemblages. The ranges in Fig. 2 are thus based on the author’s synonymy list (Gale 2012).

Stratigraphic ranges of foraminifera found in the Mt. Kobla section. Ranges are based on the author’s synonymy list. Stratigraphic resolution is kept at the stage level. In most cases it is not known which criteria for the Rhaetian has been used
5 Results
5.1 Conodont distribution
The distribution of the identified conodont taxa (Figs. 3, 4) is presented in Figs. 5, 6, 7 and 8. We focused on the identification of the Misikella representatives, mainly M. hernsteini and M. posthernsteini, which are the key taxa for the biostratigraphic definition of the Norian–Rhaetian boundary (Kozur and Mock 1974; Gazdzicki et al. 1979; Golebiowski 1986; Krystyn 1987; Kozur and Mock 1991; Kozur 1996; Krystyn et al. 2007a, b; Krystyn 2008; Moix et al. 2007; Giordano et al. 2010). Besides improved stratigraphy, a new result of this conodont study is the recognition of several new taxa: Misikella buseri, Misikella sp. A, Misikella sp. B, Parvigondolella lata and Zieglerioconus rhaeticus (see Kolar-Jurkovšek 2011).

Conodonts from the Mt. Kobla section. Scale bar 100 μm. A–D, Zieglerioconus rhaeticus Kozur and Mock 1991. Misikella posthernsteini A. Z. A–C, Sample K2/70-75 (GeoZS 4098). D, Sample K2/82.5 (GeoZS 4403). E, Transitional form between Misikella hernsteini (Mostler, 1967) and Misikella sp. B - highly evolved form. Misikella posthernsteini A. Z. Sample K2/90.4 (GeoZS 4420). F, J–M, Misikella posthernsteini Kozur and Mock 1974 . Misikella posthernsteini A. Z. F, Sample K2/88.7 (GeoZS 4416). J, K, Sample K2/85.5 (GeoZS 4409). L, M, Sample K2/90.1 (GeoZS 4419). G, Misikella buseri Kolar-Jurkovšek 2011 . Misikella posthernsteini A. Z. Sample K2/76.0 (GeoZS 4242). H, I, N, Misikella sp. B Mostler, 1978. H, I, Misikella posthernsteini A. Z. Sample K2/85.5 (GeoZS 4409). N, Un-named Misikella level. Sample K2/93.8 (GeoZS 4424). a lateral, b upper, c lower, d oblique lateral, e oblique lower, f lower posterior view

Conodonts from the Mt. Kobla section. Misikella posthernsteini A. Z. Scale bar 100 μm, in figure O 50 μm. A-G Ramiform elements: A, S3-4 element. B, S3-4 element. D, S1-2 element. C, ?S element. E, M element. F, G, M element. H–J, Oncodella paucidentata (Mostler, 1967). K–N, Norigondolella steinbergensis (Mosher, 1968). O, Parvigondolella andrusovi Kozur & Mock, 1972. A, H, I, Sample K2/88.0 (GeoZS 4415). B, C, F, G, Sample K2/82.5 (GeoZS 4403). D, Sample K2/87.6 (GeoZS 4414). E, Sample K2/86.0 (GeoZS 4410). J, Sample K2/89.1 (GeoZS 4417). K, N Sample K2/86.5 (GeoZS 4411). L, M Sample K2/87.0 (GeoZS 4412). O Sample K2/81.7 (GeoZS 4401). a upper, b lateral, c oblique lower, d oblique lateral, e lower view


An un-simplified part of the Mt. Kobla section (shaded area in Fig. 5) including the Norian-Rhaetian and the Triassic-Jurassic boundary interval. See Fig. 5 for legend. Option 2 (the First Appearance Datum of Misikella posthernsteini) of Krystyn et al. (2007a) is the favored level for the Norian-Rhaetian Boundary (Kozur 1996)


In determining the conodont elements, a taxonomy considered in Kolar-Jurkovšek (2011) is applied and criteria for some taxa are emphasised here:
The genus Parvigondolella includes elements regarded as the ultimate stage in the epigondolellid evolution. However, some authors regard the platformless gondolellid forms as morphological variants or ecostratigraphic morphotypes of E. bidentata (Gallet et al. 2007; Krystyn 2008). Discrimination of the platformless forms as a separate genus is therefore reflected in the introduction of different conodont biozones. The biozonation for the Norian-Rhaetian of Hallstatt is based on Epigondolella with separation of platformless element denoted as Epigondolella bidentata juv. (Krystyn 2008). On the other hand, Kozur and Mock (1991) proposed a biozonation based on recognition of Parvigondolella that was modified by Kozur (2003). Their view was later followed by some authors (Jadoul et al. 2007; Moix et al. 2007; Rožič et al. 2009; Muttoni et al. 2010; Kolar-Jurkovšek 2011) and is used in this study.
Misikella hernsteini and M. posthernsteini have significant stratigraphic values. Both of them were used as markers in proposals for the candidate GSSP for the base of the Rhaetian stage (e.g. Lucas 2007; Krystyn et al. 2007a, b; Lucas 2010b). In distinguishing M. posthernsteini from M. hernsteini, we follow Giordano et al. (2010). Some other species of Misikella (M. koessenensis, M. rhaetica), which were used in earlier stratigraphic schemes (Mostler et al. 1978; Krystyn 1987; Kozur and Mock 1991) and may have local stratigraphic values, are also present in investigated sections. The determination of both species renders some difficulties, as the first description of both taxa includes elements that might bear additional denticle(s) behind the cusp (Mostler et al. 1978). Therefore, M. koessenensis and M. rhaetica should be emended. In the study of the Mt. Kobla faunas, a more limited concept in recognition of the two species was used (Kolar-Jurkovšek 2011). The short and high elements that reveal no posterior denticle, composed of three to five denticles and an appropriate length: height ratio were attributed to M. koessenensis sensu lato. However, these elements are herein assigned to Misikella sp. B as a compromise suggested by Kozur (pers. comm.).
The elements of Misikella with one to two denticles of smaller size behind the cusp and with a widely opened basal area extending nearly full length were attributed to M. rhaetica sensu lato by Kolar-Jurkovšek (2011) and illustrated in Rožič et al. (2009, Fig. 9e). Similar specimens are regarded as elongated M. hernsteini by some authors (L. Krystyn and H. Kozur, both pers. comm.) and are assigned here to Misikella sp. A (sensu Muttoni et al. 2010).
5.1.1 Conodonts from the Mt. Kobla section
The conodont assemblage from the Mt. Kobla section consists of genera Epigondolella, Norigondolella, Oncodella, Parvigondolella, Misikella and Zieglerioconus.
Norigondolella steinbergensis and Epigondolella ex gr. E. abneptis were found below the Slatnik Formation, in the upper part of the Bača Dolomite. Epigondolella bidentata and N. steinbergensis were found in the lowermost part of the Slatnik Formation, where only composite samples were taken. The first Epigondolella ex gr. E. postera was found around 30 stratigraphic metres higher. Several new species enter the assemblage at the 55 m level (Figs. 5, 6) where precise point-sampling in one-meter-interval steps started. These are: Epigondolella ex gr. E. postera, Parvigondolella andrusovi, Misikella hernsteini, Zieglerioconus rhaeticus and Misikella sp. At the 58 m level, Misikella sp. A and Misikella sp. B first occur. Oncodella paucidentata was first found in the 71 m level, and stratigraphically the most important Misikella posthernsteini at the 75 m level. Misikella buseri, described as a new species by Kolar-Jurkovšek (2011), enters the assemblage at the 76 m level. The Last Appearance Datum (LAD) of Misikella hernsteini is at 81.5 m, and Misikella posthernsteini reaches up to the 91 m level. The last conodonts were retrieved just below the last channelised breccia in the Slatnik Formation, at the 94.5 m level.
5.1.2 Conodonts from the Mt. Slatnik section
The Mt. Slatnik section presents the more proximal development of the two, and very few micritic limestone beds are available for sampling (Figs. 7, 8). The complete assemblage consists of the following species (in the order of first occurrences): Epigondolella ex gr. E. abneptis, N. steinbergensis, P. andrusovi, Misikella sp. B, Misikella sp., O. paucidentata, Z. rhaeticus, M. hernsteini, transitional form between M. hernsteini and M. posthernsteini, M. posthernsteini.
Most samples were taken from very fine-grained packstones so at least a short-term resedimentation cannot be totally excluded. However, clearly re-sedimented older elements from the upper part of the Slatnik Formation (above the 67 m level) were easily recognised due to their fragmented nature and different colour. The stratigraphic occurrence of E. abneptis is confined in Slovenia to the Norian strata (Kolar-Jurkovšek 1991, Buser et al. 2007). It is important to note that Misikella posthernsteini was found in the residue of micritic, hemipelagic limestone. The Norian-Rhaetian boundary can thus be set between the 36 m level (with a transitional form between M. hernsteini and M. posthernsteini) and the 41 m level (with M. posthernsteini).
5.2 Foraminiferal distribution
Among foraminifera, two types of assemblages are distinguished. The first type is found in hemipelagites. It is characterised by low diversity and abundance. Lagenides predominate, presumably representing in situ associations. The second type of assemblage is limited to calcarenites (packstone, grainstone and fine-grained rudstone), which were interpreted as turbidity current deposits (Rožič et al. 2009; Gale 2010, 2011; Gale et al. 2011). These assemblages are low in abundance, but highly diversified, comprising 50 genera and 69 species. Reef-dwelling forms predominate, with a minor admixture of platform and fore-reef dwelling forms.
The lithological dependence of foraminifera is responsible for some apparent abrupt appearances of many species in the lower part of the Slatnik Formation, but these are a consequence of a rather small number of calciturbiditic beds in the lower, Sevatian part of the sections. Data become more consistent in the upper part of the formation where calcarenites and breccias predominate. The occurrences and local ranges of all determined foraminifera are shown in Figs. 4 and 5 and the foraminifera are shown in Figs. 9, 10, 11 and 12.

Foraminifera from the Mt. Kobla and Mt. Slatnik sections. Scale bar 200 μm, in figures n–p 100 μm. a ?Coronipora etrusca. Sample 70308. b ?Coronipora austriaca. Sample 70295. c Triasina oberhauseri. Sample 70308. Note rare pillars in the last whorl (arrowhead). d Trocholina umbo. Sample 70631. e Gandinella falsofriedli. Sample 70284. f “Duotaxis birmanica”. Sample 70295. g Galeanella tollmanni. Tangential section. Sample K2-94.20. h ?Variostoma helicta. Sample K2-70.40. i Variostoma catilliforme. Sample 70277. j “Sigmoilina” schaeferae. Sample 70324. k G. tollmanni. Sample 70295. l Ammobaculites corpulentus. Sample 70276. m ?Austrocolomia cordevolica. Sample 70087. n ?Cryptoseptida klebelsbergi. Sample 70080. o Nodosaria ordinata. Sample 70248

Foraminifera from the Mt. Kobla and Mt. Slatnik sections. Scale bar for figures b–d, e, g, h, m 100 μm, for figure r 250 μm, for figure t 500 μm, for others 200 μm. a Variostoma cochlea. Sample 70295. b Pseudonodosaria obconica. Sample K2-81.40. c Trocholina turris. Sample K2-94.20. d Siphovalvulina colomi. Sample K2-107.60. e Endotriada tyrrhenica. Sample 70294. f Kaeveria fluegeli. Sample K2-30.70. g “Trochammina” almtalensis. Sample 70284. h “Trochammina” jaunensis. Sample 70284. i ?”Tetrataxis” humilis. Sample 70276. j Reophax rudis. Sample 70327. k Reophax asperus. Sample K2-30.70. l Scherochorella eominutus. Sample 70304. m ?Gaudryinella clavuliniformis. n Alpinophragmium perforatum. o Earlandia tintinniformis. Arrowhead points to holothuria Theelia sp. Sample 70297. p Earlandia amplimuralis. Sample 70308. r ?Variostoma falcata. Sample 69859. s Endotebanella bicamerata. Sample 69858. t Acroliammina? sp. A. Sample 70286

Foraminifera from the Mt. Kobla and Mt. Slatnik sections. Scale bar 200 μm, in figure e 100 μm, in figure r 400 μm. Arrow in figures a–d shows the correct orientation of specimens. a ?Duostomina biconvexa. Sample K2-76.30. b Diplotremina subangulata. Sample 70322. c Diplotremina placklesiana. Sample K2-70.40. d ?Diplotremina astrofimbriata. Sample 70329. e Austrocolomia canaliculata. Sample 70294. f Trocholina crassa. Sample 70327. g Aulotortus tumidus. Sample 70326. h Aulotortus friedli. Sample 70326. i Aulotortus sinuosus. Sample 70627. j Angulodiscus impressus. Sample 70298. k Angulodiscus communis. Sample K2-91.30. l Auloconus permodiscoides. Sample 70295. m Ophthalmidium exiguum. Sample 70067. n Pilammina sulawesiana. Sample 70081. o Cucurbita cylindrica. Sample 70071. p ?Ophthalmidium walfordi. Sample 70072. r Triasina hantkeni. Sample 70247. s Involutina turgida (with hand-drawing). Sample 70256

Foraminifera from the Mt. Kobla section. Scale bar 200 μm, in figure d 50 μm, in figures b, h 100 μm. Arrows in figures f, g, p show the correct orientation of specimens. a, b Longitudinal section of G. tollmanni. Sample 70298. c Bispiranella salaji comb. nov. Sample 70327. d, e Agathammina austroalpina. Sample 70293. f Planiinvoluta carinata. Sample 70277. g Planiinvoluta deflexa. Arrowhead points to the surface of attachment (shell). Sample K2-51.60. h Ophthalmidium leischneri. Sample K2-51.60. i Ophthalmidium carinatum. Sample 70277. j Hydrania dulloi. Sample K2-30.70. k, m Miliolipora cuvillieri. Note coarse pores (arrowhead) in figure k. Sample 70308. l Miliolida. Sample 70276. n ?M. cuvillieri. Sample 70308. o Variostoma coniforme. Sample 70306. p ?Duostomina turboidea. Sample 70304
5.2.1 Foraminifera from the Mt. Kobla section
Foraminifera were first recorded 10 m above the base of the Slatnik Formation (Figs. 5 and 6). This sample is the only one yielding specimens of Endotriada tyrrhenica. The FO of “Sigmoilina” schaeferae is also recorded at this level, as well as the FO of “Trochammina” almtalensis, Agathammina austroalpina, Trocholina umbo, Aulotortus sinuosus, Planiinvoluta carinata, Austrocolomia canaliculata and ?Duostomina biconvexa. With the exception of the latter species, all reach the uppermost part of the Slatnik Formation. The succeeding 20 m of the section consist of hemipelagic limestones devoid of allochthonous material. With the renewed deposition of coarser material, several taxa enter the assemblage. Among them are single but undisputable specimens of Kaeveria fluegeli, Ammobaculites corpulentus, Aulotortus tumidus, Coronipora etrusca and Galeanella tollmanni. The last species, as well as “S.” schaeferae, are abundant, well preserved and easily recognisable throughout the upper part of the Slatnik Formation. Hydrania dulloi also appears at this level, but the assignment of this species is questionable, with only one higher level yielding more than one specimen. At 50 m, “Duotaxis birmanica” appears, and numerous specimens of this species were recovered up to the top of its range. Planiinvoluta deflexa, which is also first encountered at this level, differs from P. carinata in having up to four times less dense coiling. Several important species appear slightly above 70 m, among them Variostoma cochlea (with a very short range that could be extended if ?Variostoma helicta is reassigned as V. cochlea), “Tetrataxis” humilis, Aulotortus friedli, Angulodiscus communis, Gandinella falsofriedli and Variostoma coniforme. Variostoma coniforme is especially abundant and easily recognisable higher in the section. Single specimens of Coronipora austriaca and Auloconus permodiscoides were found in slightly younger beds. The appearance of Reophax rudis soon follows. An abundance of Bispiranella salaji (Orthotrinacria cf. O. expansa auct.) is prominent. Triasina oberhauseri appears at 88 m. Variostoma catilliforme, named a marker fossil for the Norian by Kristan-Tollmann (1976—cf. Haas et al. 1997), was found soon after, alongside Trocholina turris. One of the last positive samples contained Alpinophragmium perforatum embedded in an intraclast. The calcarenite bed directly overlying the last bed of hemipelagic limestone that yielded conodonts (at 94.5 m of the section) contains abundant foraminifera. Higher in the section, the assemblage is severely diminished. Though the following breccia layer was sampled several times, it yielded only lagenides (recognisable are genera Lenticulina, Pseudonodosaria and Austrocolomia) and is thus in strong contrast with lithologically similar lower breccia layers with many Triassic foraminifera. Lagenids are joined by agglutinated foraminifera, involutinids and small Ophthalmidium only above 11 m in the section. Siphovalvulina spp. appear soon after, 12 m above the last duostominids.
5.2.2 Foraminifera from the Mt. Slatnik section
The first foraminifera were found in the non-dolomitised parts of the underlying Bača Dolomite, almost 140 stratigraphic metres below the Slatnik Formation (Fig. 7). Allodapic limestones yielded several species. Of particular importance are M. cuvillieri, D. birmanica, A. perforatum and M. stellata. The last three species are only known from the Norian and Rhaetian. Palaeolituonella meridionalis, several species of duostominids, “S.” schaeferae and G. tollmanni were found at 115 m level. From the upper part of the Bača Dolomite, the findings of P. sulawesiana, C. cylindrica, H. dulloi, G. falsofriedli, T. oberhauseri and Tr. umbo are important. Questionable Ophthalmidium walfordi has been rarely cited in the literature and is not reported from strata older than Rhaetian until now (Gaździcki 1983). Calciturbidites of the succeeding Slatnik Formation are even richer in benthic foraminifera. Stratigraphically the most important is the finding of Involutina turgida (9 m above the fault separating the Bača Dolomite from the Slatnik Formation) from the immediate neighbourhood of the conodont NRB, although this form is difficult to recognise due to the susceptibility of its test to diagenetic changes. Duotaxis metula appears in the same level. The first occurrence of Tr. turris is recorded five metres higher. Twenty-two metres into the Slatnik Formation, C. deminuta, Tr. crassa and, soon after, K. fluegeli were found. The last “S.” schaeferae was determined five metres higher, and the last T. oberhauseri still five metres higher (the 66.5 m level in Fig. 8). Immediately above the Last Occurrence of T. oberhauseri, a single specimen of Triasina hantkeni was recovered. Duostominidae last until the 71 m level (on Fig. 8). Only lagenids were found in calcarenites above the LO of duostominids. These, last calcarenites of the Slatnik Formation, are followed by an interval of platy limestones of the topmost Slatnik Formation. The Slatnik Formation is separated from the Krikov Formation by a fault and the lower part of the latter is dolomitised. The first unfaulted beds of the Krikov Formation yielded Early Jurassic Siphovalvulina spp. and some of the more tolerant foraminiferal taxa.
6 Discussion
In the following text, we firstly set the stratigraphic frame on the basis of conodonts, to which the actual stratigraphic ranges of foraminifera are compared.
6.1 Conodont stratigraphic framework
Three Late Triassic conodont zones were recognised in the Mt. Kobla section (Fig. 6). The FAD of Misikella posthernsteini was taken as the marker for the base of the Rhaetian, as the currently most favoured option (Kozur 1996; Krystyn et al. 2007a, b; McRoberts et al. 2008; Rožič et al. 2009; Giordano et al. 2010; Lucas 2010b).
The lower part of the Slatnik Formation belongs to the E. bidentata Zone. Species E. bidentata, N. steinbergensis and Epigondolella ex gr. E. postera delineate the Sevatian age. The interval between the 55 m and the 75 m levels belongs to the P. andrusovi – M. hernsteini Zone. Up to the 92.5 m level, the M. posthernsteini Zone follows. According to H. J. Gawlick and L. Krystyn (2011, pers. comm.), the unusually long span of the M. posthernsteini Zone and the absence of the uppermost Rhaetian Misikella ultima Zone (see Krystyn 2008; Krystyn et al. 2009) points to the presence of hiati in this part of the section. This is in accordance with the development of the sediment by-pass zone during the forced regression (Gale et al. 2012).
In contrast, the low abundance of conodonts and the possibility of reworking in the Slatnik section prevent detailed subdivision into conodont zones. Fortunately, there is a low probability that the single sample yielding M. posthernsteini contains a reworked assemblage due to its micritic nature.
6.2 Stratigraphic value of foraminifera
From the foraminiferal assemblage of the Mt. Kobla and Mt. Slatnik sections, potentially stratigraphically the most useful species are chosen for discussion on the basis of their ranges in Fig. 2. Furthermore, we focus on the species, that are used in the existing biozonation schemes (e.g. Gaździcki 1974, 1983; Salaj 1977; Gaździcki et al. 1979; Salaj et al. 1983, 1988; Abate et al. 1984; Vachard and Fontaine 1988; Peybernes et al. 1998; He and Norling 1991; Chiocchini et al. 1994; Grgasović 1997; Peybernes et al. 1998; Mancinelli et al. 2005; Velić 2007).
6.2.1 Galeanella tollmanni
Genus Galeanella Kristan, 1958 is one of the most typical Late Triassic genera. Galeanella panticae Brönnimann et al., 1973 and Galeanella lucana Miconnet et al., 1983 are treated here as junior synonyms of Galeanella tollmanni (Kristan, 1957) (see also Kristan-Tollmann 1990).
The stratigraphic range of G. tollmanni is from the Norian to the Rhaetian (e.g. Kristan 1957; Kristan-Tollmann 1964, 1990; Brönnimann et al. 1973; Schäfer 1979; Wurm 1982; Zaninetti et al. 1982; Al-Shaibani et al. 1983; Miconnet et al. 1983; Senowbari-Daryan 1983; Matzner 1986; Martini et al. 1997), which is also in accordance with our data. Peybernes et al. (1998) determined Galeanella panticae, Galeanella laticarinata Zone from the base of the Norian to the end of the Rhaetian.
6.2.2 “Sigmoilina” schaeferae
The stratigraphic range of “S.” schaeferae is from the Norian to the Rhaetian (Zaninetti et al. 1982; Al-Shaibani et al. 1983; Salaj et al. 1983; Matzner 1986; Trifonova 1993; Senowbari-Daryan et al. 2010). In the Mt. Slatnik and Mt. Kobla sections, the combination of “S.” schaeferae with G. tollmanni provides a better consistency of data for the Norian-Rhaetian range. Both species are easily recognisable.
6.2.3 Involutina turgida
Involutina turgida is difficult to recognise due to neomorphic changes of its wall. Additionally, its abundance is low. Despite these deficiencies, it has never been reported from strata older than Rhaetian (e.g. Kristan 1957; Koehn-Zaninetti 1969; Matzner 1986; Kristan-Tollmann and Colwell 1992). It was found in the Mt. Slatnik section, between the transitional form from M. hernsteini to M. posthernsteini. Current data thus suggest its use as a proxy for the Norian-Rhaetian boundary, although further research is needed.
6.2.4 Trocholina turris
As in the case of I. turgida, Trocholina turris did not appear before the Rhaetian (Kristan-Tollmann 1986a, 1990; Kristan-Tollmann and Gramann 1992; Zaninetti et al. 1992; Senowbari-Daryan et al. 2010) and is better known from the Lower Jurassic (Ramovš and Kristan-Tollmann 1967; Brönnimann et al. 1970; Gušić 1975; Gaździcki 1983; Blau 1987b; Blau and Haas 1991; Ebli 1993; Böhm et al. 1999). It was found in both measured sections above the FAD of M. posthernsteini. Both species were also found in coexistence on Papua New Guinea by Kristan-Tollmann (1986b).
6.2.5 Duostominidae
The family Duostominidae comprises the genera Duostomina, Diplotremina, Variostoma, Krikoumbilica and Papillaria (di Bari and Laghi 1996). Duostominids first appeared in the Lower Triassic (Rettori et al. 1994; di Bari and Rettori 1996) and became extinct at the end of the Triassic (Kristan-Tollmann 1960; Chiocchini et al. 1994; di Bari and Rettori 1996; Hillebrandt and Urlichs 2008; Gale et al. 2011). They are among the most abundant foraminifera in both sections and are continuously present throughout the Slatnik Formation. As duostominids are easily recognisable as a group and cover a wide spectrum of environments, their LO makes a good marker for the Triassic-Jurassic boundary.
6.2.6 Triasina hantkeni
The type material of Triasina hantkeni was collected from Dachstein Limestone in Hungary by Majzon (1954). The age of the new species was determined in the absence of guide fossils as Norian and/or Rhaetian. Cros and Neumann (1964) determined species in the Rhaetian and/or Hettangian beds of the Dolomites. Again, the stratigraphic age was only robustly determined on the basis of superposition (above the Dolomia Principale and below Lower Jurassic beds). Oberhauser (1964) found T. hantkeni in Upper Norian (i.e.Rhaetian) beds of Austria. Bosellini and Broglio Loriga (1966) determined the species in the Rhaetian and/or Hettangian strata of the Dolomites and cited its age as undivided Norian-Hettangian. Pantić (1967) found the species in Serbia, Salaj et al. (1967), Gázdzicki and Zawidzka (1973), Gaździcki et al. (1979) in the Western Carpathians, and Kristan-Tollmann (1970) in Austria. All place it in the Rhaetian. Bassoullet and Guernet (1970) confirmed its presence in Greece, marking the age as Rhaetian or Hettangian. Gaździcki (1974) gave the species range from the Norian and/or Rhaetian to Hettangian. Gušić (1975) declined the Hettangian age, because the association of T. hantkeni with Early Jurassic fossils was not proven. His opinion was followed by Zaninetti (1976) and Piller (1978). Gaździcki and Michalik (1980) finally determined the overlapping ranges of T. hantkeni, the bivalve Rhaetavicula contorta and the conodont M. posthernsteini. According to Gaździcki (1983), the range of T. hantkeni (T. hantkeni and Glomospirella friedli Assemblage Zone) corresponds to “Choristoceras” haueri and “Choristoceras” marshi ammonoid zones. Salaj et al. (1983) assumed that the age of T. hantkeni corresponds to the “Choristoceras” marshi ammonoid zone, but with a possibility of its presence already in the upper part of the older “Rhabdoceras” suessi zone. According to Krystyn et al. (2007a, b), the Paracochloceras suessi zone starts with the FAD of M. posthernsteini. However, there is a dispute, as Krystyn et al. (2007a, b) place the FO of T. hantkeni (with question mark), as well as the FO of R. contorta, already in the lower part of their Sevatian 1. Ciarapica and Zaninetti (1984), Abate et al. (1984) and Dumont and Zaninetti (1985) placed T. hantkeni zone in the Rhaetian. Following the chronological order of publications, the species was later found in the Norian and/or Rhaetian Dachstein Limestone of the Transdanubian Range (Oravecz-Scheffer 1987), in Albania (Pirdeni 1988) and in Japan (Kristan-Tollmann 1990). At the same time, Peybernes et al. (1988) cited T. hantkeni zone from Rhaetavicula contorta beds in French Pyrenees. Zaninetti et al. (1992) and Chiocchini et al. (1994) attributed T. hantkeni zone (respectively T. hantkeni and Griphoporella curvata zone) to the Rhaetian. Martini et al. (1995) adopted a Late Norian to Rhaetian age of T. hantkeni; however, their specimens from Indonesia were found above the FAD of M. posthernsteini. Peybernes et al. (1998) count T. hantkeni as solely Rhaetian in age. Grgasović (2003, unpublished) declines the Late Norian age of T. hantkeni. According to him, the FO of T. hantkeni corresponds to the FO of R. contorta and the FO of M. posthernsteini. He correlates this event with the base of the Vandaites sturzenbaumi zone, which is, however, again not in accordance with data from the Steinbergkogel section (Krystyn et al. 2007a, 2007b). Martini et al. (2004) and Mancinelli et al. (2005) kept the Late Norian to Rhaetian age of the T. hantkeni zone. Velić (2007) names T. hantkeni the guide fossil for the Rhaetian, but does not refer to other fossil groups.
To summarise: (1) the total range of T. hantkeni corresponds to the range of the “Choristoceras” haueri and “Choristoceras” marshi ammonoid zones, sensu Gaździcki (1983); (2) the FO of T. hantkeni is co-temporal or younger than the FAD of M. posthernsteini (see Gaździcki and Michalik 1980; Martini et al. 1995); (3) the range of T. hantkeni overlaps with the range of R. contorta (see Gaździcki and Michalik 1980). When the FAD of M. posthernsteini is chosen as the base of the Rhaetian, T. hantkeni marks the Rhaetian age.
Triasina hanteni was found only in the Mt. Slatnik section in a single level, just above the last occurrence of T. oberhauseri and well above the Norian-Rhaetian boundary.
6.2.7 Triasina oberhauseri
Triasina oberhauseri is often included in foraminiferal biostratigraphy (Gaździcki et al. 1979; Salaj 1977; Gazdzicki 1983; Zaninetti et al. 1992). However, the stratigraphic range of the species is not clearly defined and is poorly correlated with other fossil groups (Röhl et al. 1991). Salaj (1977) placed its FO in the Middle Norian (Alaunian). Gaździcki et al. (1979) established the Semiinvoluta clari—Triasina oberhauseri Assemblage Zone, spanning the Alaunian—Sevatian interval. Gaździcki (1983) later cited the Lacinian–Lower Rhaetian age, from the Mojsisovicsites kerri to Cochloceras suessi amonoid zones. Specimens defined by Bystrický (1975) as Triasina hantkeni were revised as T. oberhauseri; thus, the co-existence of the latter species with Rhaetavicula contorta and Austrirhynchia cornigera (both considered as index species of the Rhaetian s.s.—cf. Gaździcki et al. 1979; Grgasović 2003, unpublished) was proven (Gaździcki 1983). Salaj et al. (1983) showed T. oberhauseri as spanning the Alaunian “Halorites horizon”, the “Rhabdoceras” suessi and the lower part of the “Choristoceras” marshi ammonoid zone, thus reaching the lower Rhaetian s.s. The Norian to basal Rhaetian (i.e., the lower part of the Triasina hantkeni Total Range Zone) range was also cited by Abate et al. (1984). In the zonation by Peybernes et al. (1988) the Triasina oberhauseri Total Range Zone spans only Lacinian and Alaunian time, but no explanation is given.
In summary, the upper limit of T. oberhauseri could be placed above the FAD of M. posthernsteini.
Triasina oberhauseri is rare in the Mt. Kobla section, with only one reliable and one unreliable determination. Both specimens are from the same interval, inside the range of M. posthernsteini. The species’ LO is slightly below the FO of Tr. turris. In the Mt. Slatnik section, T. oberhauseri first occurs about 13 m below the FAD of M. posthernsteini. It reaches high into the Rhaetian and its LO is just below the single occurrence of T. hantkeni.
6.3 Foraminifera with extended ranges
Based on the data from the Mt. Kobla and Mt. Slatnik sections, the stratigraphic ranges of some taxa are extended. Table 1 gives a shortened synonymy list for these species and their stratigraphic ranges according to the literature data, compared with the data from the Slovenian Basin. Besides the stratigraphic ranges of foraminifera according to the literature, we also give our own (L.G.) opinion on the stratigraphic ranges of the species. The range is considered extended when differing from the general information given in the literature (i.e. compared to the ranges without discarding questionable or wrong determinations). In contrast, Fig. 2 shows the ranges according to the critical evaluation of the literature.
6.4 Foraminiferal demise at the Triassic-Jurassic boundary
Foraminifera are often cited as a group that was not affected by environmental changes at the Triassic-Jurassic boundary (e.g., Hallam 2002; Tanner et al. 2004). In contrast, Kuznetzova et al. (2004) reported an extinction of about 32 % of the genera and two orders at the boundary. Galli et al. (2005) argued for a true extinction of the shallow-water benthic foraminifera at the TJB that was not simply the result of a habitat loss. Pálfy et al. (2007) marked significant changes in the foraminiferal assemblage, but these are partly masked by facies changes. The assemblage immediately after the TJB thus contains only long-ranging taxa, without typical Lower Jurassic forms (Pálfy et al. 2007). A prolonged period of recovery was observed also by Fugagnoli (2004) and Barattolo and Romano (2005). Kuerschner et al. (2007) and Hillebrandt and Urlichs (2008) showed that changes also happened among the planktonic foraminifera. Clémence et al. (2010) marked a substantial decline in diversity and abundance in the intra-platform basins of the Northern Calcareous Alps that took place simultaneously with the crysis of nannoplankton, and the post-traumatic foraminiferal assemblage is dominated by a disaster genus » Trochammina «. Kaminski et al. (2010) quantified changes in diversity among agglutinated foraminifera, but these were only mildly affected at the boundary.
The presence of possible hiati in the upper part of the Slatnik Formation does not allow detailed tracking of the changes in the foraminiferal assemblage, but a marked difference between the Late Triassic and the Lower Jurassic assemblages can nevertheless be observed. Empirical observations of the changes in the foraminiferal assemblage were quantified using the Shannon-Wiener index H’ (see Hammer and Harper 2006; Türkmen and Kazanci 2010). To exclude the differences in diversity due to (micro)facies differences, only coarse-grained packstones and fine-grained rudstones were considered. The diversity index thus reflects changes in reef and back-reef area, i.e. in the source area for turbidity currents.
Figure 13 shows changes in diversity for the Mt. Slatnik and Mt. Kobla sections. The average value of H’ prior to the TJB is 1.86 (Mt. Slatnik) and 2.23 (Mt. Kobla), and it drops after the TJB down to 0.82 and 0.35, respectively. The relatively high average values of H’ for the Slatnik Formation match the idea of a community from a mature and stable reef environment (see Türkmen and Kazanci 2010). The higher H’ values for Mt. Kobla section are attributed solely to the higher proportion of packstones relative to rudstones, due to the more distal position of the section.

Changes in Shannon-Wiener diversity index in Mt. Slatnik (a) and Mt. Kobla (b) sections. Numbers on horizontal axis indicate meters in Figs. 6 and 8. The dashed vertical line separates samples from the Slatnik Formation (left) from samples from the Krikov Formation (right) and roughly coincides with the Triassic-Jurassic boundary. The ragged appearance of the diversity line is due to differences in granulation, as the influence of lithology could not be totally reduced
The foraminiferal assemblage of the Slatnik Formation is markedly more diversified than the assemblage of the Krikov Formation, despite the same granulation of sampled beds. The diversity is lower, especially on account of foraminifera from the reef-area and duostominids. Both groups completely disappear at the Triassic-Jurassic boundary. The diversity is also lower among the back-reef foraminifera. As is evident from the grain composition, the platform morphology changed from reef-rimmed in Late Triassic to a carbonate ramp in Lower Jurassic. The disappearance of reef-dwelling foraminifera can thus be explained solely with the loss of their habitat, i.e. reefs. The disappearance of reefs in the Tethyan area is well known (Flügel 2002; Kiessling et al. 2007) and is attributed to the fall in relative sea-level (Borsato et al. 1994; McRoberts et al. 1997; Hallam and Wignall 1999; Hallam 2002; Hautman 2004; Lucas and Tanner 2004; Jadoul et al. 2005; Lindström and Erlström 2006; Ciarapica 2007; Khalifa 2007; but see Gómez et al. 2007), followed by transgression of oxygen-depleted water (Barras and Twitchett 2007; Ruhl et al. 2010), or to a biocalcification crisis (e.g. McRoberts and Newton 1995; McRoberts et al. 1997; Galli et al. 2005; Huynh and Poulsen 2005; Iannace et al. 2005; Ciarapica 2007; van de Schootbrugge et al. 2007; Ruhl et al. 2010; Črne et al. 2011). As yet, there is no evidence for a sea-level fall from this area.
The boundary was crossed by ecologically less demanding taxa, which could occupy a broader set of environments, especially small lagenids, simple agglutinates and Ophthalmidium. They are soon joined by few other taxa (Duotaxis spp., Siphovalvulina spp., Involutinidae), which lived on the platform. According to the literature (see Fig. 2) other species are expected to cross the boundary: D. birmanica, D. metula, Tr. umbo, Tr. turris, I. turgida, C. deminuta, C. etrusca, C. austriaca, P. carinata, O. carinatum, O. leischneri and ?O. walfordi. Of these, only P. carinata was found immediately above the boundary. Other species could be Lazarus taxa, therefore re-entering the community higher up in the sections.
Whether the disappearance of shallow-water foraminifera could be explained by changes in relative sea-level, this does not hold for duostominids. Representatives of this family can be found in a broad set of environments, from the platform (Hohenegger and Lobitzer 1971; Schäfer and Senowbari-Daryan 1978; Wurm 1982; Bérczi-Makk 1996a; Martini et al. 2004) to the basin (Kristan-Tollmann 1986a, 1988, 1990; di Bari and Baracca 1998). The disappearance of duostominids could point to a change in biochemical factors (oxygen level or ocean acidification, i.e. the biocalcification crisis), not merely to the habitat loss. A better understanding of the ecological demands of duostominids could thus provide further answers about environmental changes at the Triassic-Jurassic boundary.
7 Conclusions
The Slatnik Formation comprises interfingering hemipelagic limestones and calciturbidites deposited during the Late Norian-end-of-Rhaetian in the northern part of the Slovenian Basin (Rožič et al. 2009). Shallow water benthic foraminifera, derived from the adjacent Julian Carbonate Platform (cf. Buser 1986, 1989, 1996) are present, in addition to conodonts.
The NRB is placed at the FAD of M. posthernsteini, while foraminifera are not a good marker for this boundary. The exception could be the FO of Involutina turgida. The TJB is defined with the LO of duostominds and some other typical Late Triassic foraminifera, such as G. tollmanni and “S.” schaeferae.
An abrupt and significant change in foraminiferal assemblage from highly diversified to severely depleted is recorded across the TJB. While the disappearance of reef and back-reef dwelling forms could be explained solely by the change in relative sea-level, the extinction of duostominids cannot be related to the loss of their habitat. Thus, a biocalcification crisis seems a plausible explanation.
References
Abate, B., Ciarapica, G., & Zaninetti, L. (1984). Triasina oberhauseri Koehn-Zaninetti et Bronnimann, 1968, dans le Trias Superieur recifal (facies “back-reef”) de la plate-forme Panormide, Sicile. Revue de Paléobiologie, 3, 19–25.
Al-Shaibani, S. K., Carter, D. J., & Zaninetti, L. (1983). Geological and micropaleontological investigations in the Upper Triassic (Asinepe limestone) of Seram, outer Banda arc, Indonesia. Archives des Sciences Geneve, 36, 297–313.
Babić, L., Gušić, L., & Zupanič, J. (1979). Starost i fosili Lipovac-vapnenca (trijas) kod Samobora, zapadno od Zagreba. Geološki Vjesnik, 31, 21–35.
Barattolo, F., & Romano, R. (2005). Shallow carbonate platform bioevents during the Upper Triassic-Lower Jurassic: an evolutive interpretation. Bolletino della Societa Geologica Italiana, 124, 123–142.
Barras, C. G., & Twitchett, R. J. (2007). Response of the marine infauna to Triassic-Jurassic environmental change: ichnological data from southern England. Palaeogeography. Palaeoclimatology, Palaeoecology, 244, 223–241.
Bassoullet, J. P., & Guernet, C. (1970). Le Trias et le Jurassique de la région des Lacs de Thebes (Béotie et locride, Grece). Revue de Micropaléontologie, 12(4), 209–217.
Benjamini, C. (1988). Triassic foraminifera from Makhtesh Ramon, Central Negev, Southern Israel. Revue de Paléobiologie, Vol. Spec. No. 2, Benthos’86, 129–144.
Bérczi-Makk, A. (1996a). Foraminifera of the Triassic formations of Alsó Hill (Northern Hungary). Acta Geologica Hungarica, 39, 175–221.
Bérczi-Makk, A. (1996b). Foraminifera of the Triassic formations of Alsó Hill (Northern Hungary). Part 2: foraminifer assemblage of the Wetterstein Limestone Formation. Acta Geologica Hungarica, 39(3), 223–309.
Bérczi-Makk, A. (1996c). Foraminifera of the Triassic formations of Alsó Hill (Northern Hungary). Part 3: foraminifer assemblage of the basinal facies. Acta Geologica Hungarica, 39(4), 413–459.
Bertinelli, A., Ciarapica, G., & Passeri, L. (2005). Late Triassic-Jurassic basinal successions in Molise and northern Basilicata: the northernmost witness of the Ionian Ocean. Bolletino della Societa Geologica Italiana, 124, 177–188.
Blau, J. (1987a). Neue Foraminiferen aus dem Lias der Lienzer Dolomiten. Teil I: Die Foraminiferenfauna einer roten Spaltenfüllung in Oberrhätkalken. Jahrbuch der Geologischen Bundesanstalt, 129, 495–523.
Blau, J. (1987b). Neue Foraminiferen aus dem Lias der Lienzer Dolomiten. Teil II (Schluss): Foraminiferen (Involutinina, Spirillinina) aus der Lavanter Breccie (Lienzer Dolomiten) und den Nördlichen Kalkalpen. Jahrbuch der Geologischen Bundesanstalt, 130, 5–23.
Blau, J., & Haas, J. (1991). Lower Liassic involutinids (foraminifera) from the Transdanubian Central Range, Hungary. Paläontologische Zeitschrift, 65, 7–23.
Böhm, F., Ebli, O., Krystyn, L., Lobitzer, H., Rakús, M., & Siblík, M. (1999). Fauna, stratigraphy and depositional environment of the Hettangian-Sinemurian (Early Jurassic) of Adnet (Salzburg, Austria). Abhandlungen der geologischen Bundesanstalt, 56(2), 143–271.
Borsato, A., Frisia, S., Sartorio, D. (1994). Late Triassic–Early Liassic stratigraphic and diagenetic evolution of the margin between the Trento Platform and the Lombardy Basin in the Brenta Dolomites (Italy). Studi Trentini di Scienze Naturali. Acta Geologica, 69 (1992), 5–35.
Borza, K. (1975). Mikroproplematika aus der Oberen Trias der Westkarpaten. Geologický Zborník – Geologica Carpathica, 26(2), 199–236.
Bosellini, A., & Broglio Loriga, C. (1966). Presenza di unostrato a Foraminiferi nel Raibliano (Carnico seperiore) del Gruppo di Sella, Dolomiti occidentali. Studi Trentini di Scienze Naturali, Sez. A, 43 (1), 149–156.
Brönnimann, P., Cadet, J. P., Ricou, L. E., & Zaninetti, L. (1973). Révision morphologique et émendation du genre triasique Galeanella Kristan-Tollmann (Foraminifere) et description de Galeanella panticae, n.sp., (Dinarides yougoslaves et Zagros, Iran). Verhandlungen der Geologischen Bundesanstalt, 3, 411–435.
Brönnimann, P., Poisson, A., Zaninetti, L. (1970). L’unité du Domuz Dag (Taurus Lycien-Turquie). Microfacies et foraminiferes du Trias et du Lias. Rivista Italiana di Paleontologia, 76 (1), 1–36.
Buser, S. (1986). Explanatory book, Sheet Tolmin and Videm (Udine) L33–64, L33–63. Basic geological map of SFRJ 1:100,000. (103 p.). Beograd: Zvezni geološki zavod.
Buser, S. (1989). Development of the Dinaric and the Julian carbonate platforms and of the intermediate Slovenian Basin (NW Yugoslavia). Bolletino della Societa Geologica Italiana, 40, 313–320.
Buser, S. (1996). Geology of Western Slovenia and its paleogeographic evolution. In Drobne, K., Goričan, Š., Kotnik, B. (eds): The role of impact processes and biological evolution of planet Earth: International workshop, Postojna, 27. 9–2. 10. 1996, (pp. 111–123). Ljubljana: ZRC SAZU.
Buser, S., Kolar-Jurkovšek, T., & Jurkovšek, B. (2007). Triasni konodonti slovenskega bazena. Geologija, 50, 19–28.
Bystrický, J. (1975). The Kössen Beds in the West Carpathians and the problem of their stratigraphic division. Geolický Zbornik Geologica Carpathica, 26, 177–197.
Carter, E. S. (1993). Biochronology and Paleontology of uppermost Triassic (Rhaetian) radiolarians, Queen Charlotte Islands, British Columbia, Canada. Mémoires de Géologie (Lausanne), 11, 1–175.
Carter, E. S., & Orchard, M. J. (2004). A Norian-Rhaetian boudary at Kennecott Point (Queen Charlotte Islands, Canada) defined by radiolarians and conodonts. Albertiana, 31, 7.
Chiocchini, M., Farinacci, A., Mancinella, A., Molinari, V., Potetti, M. (1994). Biostratigrafia a foraminiferi, dasicladali e calpionelle delle successioni carbonatiche Mesozoiche dell’Appennino Centrale (Italia). Studi Geologici Camerti, “Biostratigrafia dell’Italia centrale”, 9–130.
Ciarapica, G. (2007). Regional and global changes around the Triassic-Jurassic boundary reflected in the late Norian-Hettangian history of the Apennine basins. Palaeogeography, Palaeoclimatology, Palaeoecology, 244, 34–51.
Ciarapica, G., Cirilli, S., Passeri, L., Trincianti, E., & Zaninetti, L. (1987). »Andriti di Burano « et » Formation du Monte Cetone « (nouvelle formation), biostratigraphie de deux series-types du Trias Superieur dans l’Apennin Septentrional. Revue de Paléobiologie, 6(2), 341–409.
Ciarapica, G., & Zaninetti, L. (1984). Foraminiferes et biostratigraphie dans le Trias superieur de la serie de la Spezia (Dolomies de Coregna et formation del la Spezia, Nouvellles formations) Apennin septentrional. Revue de Paléobiologie, 3(1), 117–134.
Clémence, M.-E., Gardin, S., Bartolini, A., Paris, G., Beaumont, V., & Guex, J. (2010). Bentho-planktonic evidence from the Austrian Alps for a decline in sea-surface carbonate production at the end of the Triassic. Swiss Journal of Geosciences, 103(2), 293–315.
Črne, A. E., Weissert, H. J., Goričan, Š., & Bernasconi, S. M. A. (2011). Biocalcification crisis at the Triassic-Jurassic boundary recorded in the Budva Basin (Dinarides, Montenegro). Geological Society of America Bulletin, 123, 40–50.
Cros, P., & Neumann, M. (1964). Contribution a l’étude des formations a Triasina Majzon des Dolomites Centrales. Revue de Micropaléontologie, 7(2), 125–137.
Di Bari, D. (1997). Lamellar wall of Cryptoseptida klebelsbergi (Oberhauser, 1960) (Foraminiferida) from San Cassiano Formation (Western Dolomites, Italy). Neues Jahrbuch für Geologie und Paläontologie Mithandlungen, 12, 319–328.
Di Bari, D., & Baracca, A. (1998). Late Triassic (Carnian) foraminifers of Northeastern Cortina d’Ampezzo (Tamarin, San Cassiano Fm., Dolomites, Italy). Annali Museo Civico di Rovereto: Sezione Archeologia, Storia, Scienze naturali, 12 (1996), 117–146.
Di Bari, D., & Laghi, G. F. (1996). Lamellar structure of Triassic trochospiral involutinids (Foraminiferida). Accad. Naz. Sci. Lett. Arti di Modena Collana di Studi, Miscellanea Geologica, 15, 331–340.
Di Bari, D., & Rettori, R. (1996). Morphological features of Triasina hantkeni Majzon, 1954 (Foraminiferida, Aulotortidae) and remarks on the test wall structure. Revue de Micropaléontologie, 39(4), 305–313.
Dumont, T., & Zaninetti, L. (1985). Découverte de Triasina hantkeni Majzon (Foraminifere) dans un facieau calcaire a polypiers (Rhabdophyllia) de la formation Rhético-Hettangienne Prépiémontaise (Nappe de Rochebrune, SE de Briancon, Alpes Occedentales). Archives des Sciences Geneve, 38(1), 63–70.
Ebli, O. (1993). Foraminiferen aus dem Unterlias der Nördlichen Kalkalpen. Zitteliana, 20, 155–164.
Flügel, E. (2002). Triassic reef patterns. In Kiessling, W., Flügel, E., Golonka, J. (eds): Phanerozoic Reef Patterns. (Vol. 72, pp. 391–463). SEPM Special Publication.
Fontaine, H., Khoo, H. P., & Vachard, D. (1988). Discovery of Triassic fossils at Bukit Chuping, in Gunung Sinyum area, and at Kota Jin, Peninsular Malaysia. Journal of Southeast Asian Earth Sciences, 2(3–4), 145–162.
Fugagnoli, A. (2004). Trophic regimes of benthic foraminiferal assemblages in Lower Jurassic shallow water carbonates from northeastern Italy (Calcari Grigi, Trento Platform, Venetia Prealps). Palaeogeography, Palaeoclimatology, Palaeoecology, 205, 111–130.
Gale, L. (2010). Microfacies analysis of the Upper Triassic (Norian) “Bača Dolomite”: early evolution of the western Slovenian Basin (eastern Southern Alps, western Slovenia). Geologica Carpathica, 61, 293–308.
Gale, L. (2012). Biostratigraphy and sedimentology of the Norian-Rhaetian beds of the western Slovenian Basin, Southern Alps, Slovenia. Ph.D. dissertation, University of Ljubljana, Ljubljana, Slovenia, 268 pp.
Gale, L., Rettori, R., Martini, R., Šmuc, A., Kolar-Jurkovšek, T., & Rožič, B. (2011). Duostominidae (Foraminiferea, Robertinida) from the Upper Triassic beds of the Slovenian Basin (Southern Alps, Slovenia). Rivista Italiana di Paleontologia e Stratigrafia, 117(3), 375–397.
Gale, L., Šmuc, A., Kolar-Jurkovšek, T., Skeberne, D., Celarc, B., Čar, J., Rožič, B. (2012). Norian-Rhaetian sedimentary evolution of the Slovenian Basin (eastern Southern Alps). Geophysical Research Abstracts, 14.
Gallet, Y., Krystyn, L., Marcoux, J., & Besse, J. (2007). New constraints on the End-Triassic (Upper Norian-Rhaetian) magnetostratigraphy. Earth and Planetary Science Letters, 255, 458–470.
Galli, M. T., Jadoul, F., Bernasconi, S. M., & Weissert, H. (2005). Anomalies in global carbon cycling and extinction at the Triassic/Jurassic boundary: evidence from a marine C-isotope record. Palaeogeography, Palaeoclimatology, Palaeoecology, 216, 203–214.
Gaździcki, A. (1974). Rhaetian microfacies, stratigraphy and facial development in the Tatra Mts. Acta Geologica Polonica, 24, 17–120.
Gaździcki, A. (1983). Foraminifers and biostratigraphy of Upper Triassic and Lower Jurassic of the Slovakian and Polish Carpathians. Palaeontologia Polonica, 44, 109–169.
Gaździcki, A., Kozur, H., & Mock, R. (1979). The Norian-Rhaetian boundary in the light of micropaleontological data. Geologija, 22, 71–112.
Gaździcki, A., Kozur, H., Mock, R., & Trammer, J. (1978). Triassic microfossils from the Korytnica Limestones at Lipovská Osada (Slovakia, ČSSR) and their stratigraphic significance. Acta Palaeontologica Polonica, 23(3), 351–373.
Gaździcki, A., & Michalik, J. (1980). Uppermost Triassic sequences of the Choč nappe (Hronic) in the West Carpathians of Slovakia and Poland. Acta Geologica Polonica, 30, 61–76.
Gázdzicki, A., & Smit, O. E. (1977). Triassic foraminifers from the Malay Peninsula. Acta Geologica Polonica, 27(3), 319–332.
Gaździcki, A., Trammer, J., & Zawidzka, K. (1975). Foraminifers from the Muschelkalk of Southern Poland. Acta Geologica Polonica, 25(2), 273–298.
Gázdzicki, A., & Zawidzka, K. (1973). Triassic foraminifer assemblages in the Choč nappe of the Tatra Mts. Acta Geologica Polonica, 23(3), 483–490.
Giordano, N., Rigo, M., Ciarapica, G., & Bertinelli, A. (2010). New biostratigraphical constraints for the Norian/Rhaetian boundary: data from Lagonegro Basin, Southern Apennines. Italy. Lethaia, 43(4), 573–586.
Giordano, N., Rigo, M., Ciarapica, G., Mietto, P., Passeri, L. (2008). The Norian/Rhaetian boundary in the Lagonegro Basin, Southern Apennines, Italy. In Krystyn, L., & Mandl, M. G. (eds): Upper Triassic subdivisions, zonations and events: meeting of the late IGCP 467 and STS, Abstracts and Excursion-Guide 24-27. Bad Goisern, 28. 9. – 2. 10. 2008. Berichte der Geologische Bundesanstalt 76, 24-27.
Golebiowski, R. (1986). Neue Misikellen-Funde (Conodonta) und ihre Bedeutung für die Abgrenzung des Rhät s. str. in den Kössener Schichten. Aus den Sitzungsberichten der Österreichischen Akademie der Wissenschaften Mathematisch-naturwissenschaftliche Klasse, 195(1–5), 53–65.
Golonka, J. (2007). Late Triassic and Early Jurassic palaeogeography of the world. Palaeogeography, Palaeoclimatology, Palaeoecology, 244, 297–307.
Gómez, J. J., Goy, A., & Barrón, E. (2007). Events around the Triassic-Jurassic boundary in northern and eastern Spain: a review. Palaeogeography, Palaeoclimatology, Palaeoecology, 244, 89–110.
Goričan, Š., Košir, A., Rožič, B., Šmuc, A., Gale, L., Kukoć, D., et al. (2012). Mesozoic deep-water basins of the eastern Southern Alps (NW Slovenia). Journal of Alpine Geology, 54, 101–143.
Grgasović, T. (1997). Upper Triassic biostratigraphy and algae from Žumberak (Croatia). Geologia Croatica, 50, 201–214.
Grgasović, T. (2003). Revizija rodova Physoporella Steinmann i Oligoporella Pia (Dasycladales). Ph.D. dissertation, University of Zagreb, Zagreb, Croatia, 484 pp.
Gullo, M. (1996). Conodont biostratigraphy of uppermost Triassic deep-water calcilutites from Pizzo Mondello (Sicani Mountains): evidence for Rhaetian pelagites in Sicily. Palaeogeography, Palaeoclimatology, Palaeoecology, 126, 309–323.
Gušić, I. (1975). Upper Triassic and Liassic Foraminiferida of Mt. Medvednica, Northern Croatia (families: Involutinidae, Nubeculariidae) [Gornjetrijaske i lijaske foraminifere Medvednice, sjeverna Hrvatska]. Palaeontologia Jugoslavica, 15, 7–45.
Haas, J. (2004). Characteristics of peritidal facies and evidences for subaerial exposures in Dachstein-type cyclic platform carbonates in the Transdanubian Range, Hungary. Facies, 50, 263–286.
Haas, J., Götz, A., & Pálfy, J. (2010). Late Triassic to Early Jurassic palaeogeography and eustatic history in the NW Tethyan realm: new insights from sedimentary and organic facies of the Csővár Basin (Hungary). Palaeogeography, Palaeoclimatology, Palaeoecology, 291, 456–468.
Haas, J., Kovácz, S., Krystyn, L., & Lein, R. (1995). Significance of Late Permian-Triassic facies zones in terrane reconstructions in the Alpine-North Pannonian domain. Tectonophysics, 242, 19–40.
Haas, J., Lobitzer, H., & Monostori, M. (2007). Characteristics of the Lofer cyclicity in the type locality of the Dachstein Limestone (Dachstein Plateau, Austria). Facies, 53, 113–126.
Haas, J., Tardi-Filácz, E., Oravecz-Scheffer, A., Góczán, F., & Dosztály, L. (1997). Stratigraphy and sedimentology of an Upper Triassic toe-of-slope and basin succession at Csővár, North Hungary. Acta Geologica Hungarica, 40, 111–177.
Hallam, A. (2002). How catastrophic was the end-Triassic mass extinction? Lethaia, 35, 147–157.
Hallam, A., & Wignall, P. B. (1999). Mass extinctions and sea-level changes. Earth-Science Reviews, 48, 217–250.
Hammer, Ø., & Harper, D. A. T. (2006). Paleontological data analysis (351 pp.). Oxford: Blackwell Publishing.
Hautman, M. (2004). Effect of end-Triassic CO2 maximum on carbonate sedimentation and marine mass extinction. Facies, 50, 257–261.
He, Y. (1984). Middle Triassic foraminifera from Central and Southern Guizhou, China. Acta Palaeontologica Sinica, 23(4), 420–431.
He, Y. (1999). Triassic foraminifera from northwestern Yunnan. Acta Micropalaeontologica Sinica, 16(1), 31–49.
He, Y., & Norling, E. (1991). Upper Triassic foraminifera and stratigraphy of Mianzhu, Sichuan province, China. Sveriges Geologiska Undersökning, Avhandlingar Och Uppsatser, 76, 1–47.
Hillebrandt, A. V. (2008). Aragonitische Foraminiferen (Robertinina) aus dem Trias/Jura-Grenzbereich der Nördlichen Kalkalpen und ihre stratigraphische Bedeutung. Erlanger Geologische Abhandlungen Sdb., 6, 34.
Hillebrandt, A. von & Urlichs, M. (2008). Foraminifera and ostracoda from the Northern Calcareous Alps and the end-Triassic biotic crisis. In Krystyn, L., & Mandl, W. (Eds): Upper Triassic subdivisions, zonations and events: meeting of the late IGCP 467 and STS, Abstracts and Excursion-Guide. Bad Goisern, 28. 9. – 2. 10. 2008. Berichte der Geologischen Bundesanstalt, 76, 28-35.
Hohenegger, J., & Lobitzer, H. (1971). Die Foraminiferen-Verteilung in einem obertriadischen Karbonatplattform-Becken-Komplex der östlichen Nördlichen Kalkalpen. Verhandlungen der geologischen Bundesanstalt, 3, 458–485.
Hüsing, S. K., Deenen, M. H. L., Koopmans, J. G., & Krijgsman, W. (2011). Magnetostratigraphic dating of the proposed Rhaetian GSSP at Steinbergkogel (Upper Triassic, Austria): implications for the Late Triassic time scale. Earth and Planetary Science Letters, 302, 203–216.
Huynh, T. T., & Poulsen, C. J. (2005). Rising atmospheric CO2 as a possible trigger for the end-Triassic mass extinction. Palaeogeography, Palaeoclimatology, Palaeoecology, 217, 223–242.
Iannace, A., Parente, M., & Zamparelli, V. (2005). The Upper Triassic platform margin facies of Southern Apennines and their Jurassic fate: state of the art. Bolletino della Societa Geologica Italiana, 124, 203–214.
Isintek, I., Altiner, D., & Koca, U. (2000). Middle Triassic foraminifera from the type section of the Laleköy Formation (Karaburun Peninsula, western Turkey): remarks on Paleolituonella meridionalis (Luperto, 1965). Revue de Paléobiologie, 19(1), 191–205.
Jadoul, F., Galli, M. T., Calabrese, L., & Gnaccolini, M. (2005). Stratigraphy of Rhaetian to Lower Sinemurian carbonate platfroms in Western Lombardy (Southern Alps, Italy): paleogeographic implications. Rivista Italiana di Paleontologia e Stratigrafia, 111, 285–303.
Jadoul, F., Galli, M. T., Muttoni, G., Rigo, M., Cirilli, S. (2007). The late Norian-Hettangian stratigraphic and paleogeographic evolution of the Bergamasc Alps. Geoitalia 2007, Pre-Congress Field Trip Guide Book – FW02, Rimini, 1–33.
Kaminski, M. A., Setoyama, E., & Cetean, C. G. (2010). The phanerozoic diversity of agglutinated foraminifera: orignation and extinction rates. Acta Palaeontologica Polonica, 55(3), 529–539.
Khalifa, M. A. (2007). Triassic-Jurassic boundary on the southern margin of Tethys: implications of facies, tectonics and volcanism. In Lucas, S. G., & Spielmann, J. A. (Eds): The Global Triassic, New Mexico Museum of Natural History Science Bulletin, 41, 110–119.
Kiessling, W., Aberhan, M., Brenneis, B., & Wagner, P. J. (2007). Extinction trajectories of benthic organisms across the Triassic-Jurassic boundary. Palaeogeography, Palaeoclimatology, Palaeoecology, 244, 201–222.
Kobayashi, F. (1996). Middle Triassic (Anisian) foraminifers from the Kaizawa Formation, southern Kanto Mountains, Japan. Trans. Proc. Palaeont. Soc. Japan, 183, 528–539.
Kobayashi, F., Martini, R., Rettori, R., Zaninetti, L., Ratanasthien, B., Saegusa, H., et al. (2006). Triassic foraminifers of the Lampang Group (Northern Thailand). Journal of Asian Earth Sciences, 27, 312–325.
Kobayashi, F., Martini, R., & Zaninetti, L. (2005). Anisian foraminifers from allochthonous limestones of the Tanoura formation (Kurosegawa Terrane, West Kyushu, Japan). Geobios, 38, 751–763.
Koehn-Zaninetti, L. (1969). Les Foraminiferes du Trias de la region de l’Almtal (Haute-Autriche). Jahrbuch der Geologischen Bundesanstalt, Sdb., 14, 1–155.
Kolar-Jurkovšek, T. (1991). Microfauna of Middle and Upper Triassic in Slovenia and its biostratigraphic significance. Geologija, 33, 21–170.
Kolar-Jurkovšek, T. (2011). Latest Triassic conodonts of the Slovenian Basin and some remarks on their evolution. Geologija, 54, 81–90.
Kozur, H. (1996). The position of the Norian-Rhaetian boundary. In Jost Wiedmann Symposium, Abstracts. Ber. Rep. Geol. Paläont. Univ. Kiel, 76, 27–35.
Kozur, H. (2003). Integrated ammonoid-, conodont and radiolarian zonation of the Triassic. Hallesches Jahrbuch Geowissenschaft, 25, 49–79.
Kozur, H., & Mock, R. (1974). Misikella posthernsteini n.sp., die jüngste Conodontenart der tethyalen Trias. Časopis pro Mineralogii a Geologii, 19, 245–250.
Kozur, H., & Mock, R. (1991). New Middle Carnian and Rhaetian conodonts from Hungary and the Alps. Stratigraphic importance and tectonic implications for the Buda Mountains and adjacent areas. Jahrbuch der geologischen Bundesanstalt, 134, 271–297.
Kozur, H., & Mostler, H. (1973). Mikrofauna und Stratigraphie im Raume Csövár Ungarn. Verhandlungen der Geologischen Bundesanstalt, 1973(2), 291–325.
Kristan, E. (1957). Ophthalmidiidae und Tetrataxinae (Foraminifera) aus dem Rhät der Hohen Wand in Nieder-Österreich. Jahrbuch der geologischen Bundesanstalt, 100, 269–298.
Kristan-Tollmann, E. (1960). Rotaliidea (Foraminifera) aus der Trias der Ostalpen. Jahrbuch der geologischen Bundesanstalt, Sdb., 5, 47–78.
Kristan-Tollmann, E. (1964). Die Foraminiferen aus den Rhätischen Zlambachmergeln der Fischerwiese bei Aussee im Salzkammergut. Jahrbuch der Geologischen Bundesanstalt, Sdb., 10, 1–189.
Kristan-Tollmann, E. (1970). Beiträge zur Mikrofauna des Rhät III. Foraminiferen aus dem Rhät des Königsbergzuges bei Göstling (Nieder-Österreich). Mitteilungen der Gesselschaft für Geologische Bergbaustudien, 19, 1–14.
Kristan-Tollmann, E. (1973). Neue sandschalige Foraminiferen aus der alpinen Obertrias [New agglutinated foraminifera of the Upper Trias from the Alps]. Neues Jahrbuch für Geologie und Paläontologie Mithandlungen, 7, 416–428.
Kristan-Tollmann, E. (1983). Foraminiferen aus dem Oberanis von Leidapo bei Guiyang in Südchina. Mitteilungen der Österreichischen Geologischen Gesellschaft, 76, 289–323.
Kristan-Tollmann, E. (1986a). Beobachtungen zur Trias am Südostende der Tethys – Papua/Neuguinea, Australien, Neuseeland [Observations on the Triassic of the southeastern margin of the Tethys – Papua/New Guinea, Australia and New Zealand]. Neues Jahrbuch für Geologie und Paläontologie Mithandlungen, 4, 201–222.
Kristan-Tollmann, E. (1986b). Foraminiferen aus dem rhätischen Kuta-Kalk von Papua/Neuguinea. Mitteilungen der Österreichischen Geologischen Gesellschaft, 78(1985), 291–317.
Kristan-Tollmann, E. (1988). A comparison of Late Triassic agglutinated foraminifera of Western and Eastern Tethys. Abhandlungen der geologischen Bundesanstalt, 41, 245–253.
Kristan-Tollmann, E. (1990). Rhät-Foraminiferen aus dem Kuta-Kalk des Gurumugl-Riffes in Zentral-Papua/Neuguinea. Mitteilungen der Österreichischen Geologischen Gesselschaft, 82, 211–289.
Kristan-Tollmann, E. (1991). Triassic Tethyan microfauna in Dachstein limestone blocks in Japan. In: Kotaka, T., Dickins, J. M., McKenzie, K. G., Mori, K., Ogasawara, K., Stanley, G. D. Jr. (editors): Shallow Tethys 3. Proceedings of the International Symposium on Shallow Tethys 3, Sendai, Japan, 20-23 September 1990. Special Publication, Saito Ho-on Kai, 3, 35-49.
Kristan-Tollmann, E., & Gramann, F. (1992). Paleontological evidence for the Triassi age of rocks dredged from the northern Exmouth Plateau (Tethyan foraminifers, echinoderms, and ostracodes). In von Rad, U., Haq, B. U., et al. (Eds): Proceedings of the Ocean Drilling Program, Scientific Results. College Station, Texas, 122, 463–471.
Krystan-Tollmann, E., & Colwell, J. (1992). Alpiner Enzesfelder Kalk (Unter-Lias) vom Exmouth-Plateau nordwestlich von Australien. Mitteilungen der Österreichischen Geologischen Gesellschaft, 84(1991), 301–308.
Krystyn, L. (1987). Zur Rhät-Stratigraphie in den Zlambach-Schichten (vorläufiger Bericht). Aus den Sitzungsberichten der Österreichischen Akademie der Wissenschaften Mathematisch-naturwissenschaftliche Klasse, 196(3), 21–36.
Krystyn, L. (2008). An ammonoid-calibrated Tethyan conodont time scale of the Late Upper Triassic. In Krystyn, L., & Mandl, W. (Eds): Upper Triassic subdivisions, zonations and events: meeting of the late IGCP 467 and STS, Abstracts and Excursion-Guide. Bad Goisern, 28. 9. – 2. 10. 2008. Berichte der Geologischen Bundesanstalt, 76, 9–11.
Krystyn, L., Bouquerel, H., Kuerschner, W., Richoz, S., Gallet, Y. (2007a). Proposal for a candidate GSSP for the base of the Rhaetian stage. In Lucas, S. G., & Spielmann, J. A. (Eds): The Global Triassic, New Mexico Museum of Natural History and Science Bulletin, 41, 189–199.
Krystyn, L., & Kuerschner, W. M. (2005) Biotic events around the Norian - Rhaetian boundary from a Tethyan perspective. Albertiana, 32, 17–20.
Krystyn, L., Mandl, G. W., & Schauer, M. (2009). Growth and termination of the Upper Triassic platform margin of the Dachstein area (Northern Calcareous Alps, Austria). Austrian Journal of Earth Sciences, 102, 23–33.
Krystyn, L., Richoz, S., Gallet, Y., Bouquerel, H., Kürschner, W. M., & Spötl, C. (2007b). Updated bio- and magnetostratigraphy from Steinbergkogel (Austria), candidate GSSP for the base of the Rhaetian stage. Albertiana, 36, 164–172.
Kuerschner, W. M., Bonis, N. R., & Krystyn, L. (2007). Carbon-isotope stratigraphy and palynostratigraphy of the Triassic-Jurassic transition in the Tiefengraben section – Northern Calcareous Alps (Austria). Palaeogeography, Palaeoclimatology, Palaeoecology, 244, 257–280.
Kuznetsova, K. I., Korchagin, O. A., Basov, V. A. (2004). Foraminifers and the Triassic-Jurassic biotic crisis. Proceedings of the 5th International Symposium on Eastern Mediterranean Geology, Thessaloniki, Greece, 14–20 April 2004, 5–12.
Lindström, S., & Erlström, M. (2006). The late Rhaetian transgression in southern Sweden: regional (and global) recognition and relation to the Triassic-Jurassic boundary. Palaeogeography, Palaeoclimatology, Palaeoecology, 241, 339–372.
Lucas, S. G. (2007). Proxies and Triassic GSSPs. Albertiana, 36, 174–175.
Lucas, S. G. (2010a). The Triassic timescale: an introduction. In Lucas, S. G. (Ed): The Triassic timescale, Geological Society Special Publication, 344, 1–16.
Lucas, S. G. (2010b). The Triassic chronostratigraphic scale: history and status, In Lucas, S. G. (Ed): The Triassic timescale, Geological Society Special Publication, 344, 17–39.
Lucas, S. G., & Tanner, L. H. (2004). Late Triassic extinction events. Albertiana, 31, 31–40.
Majzon, L. (1954). Contributions to the stratigraphy of the Dachstein limestone. Acta Geol. Acad. Sci. Hongrie, 2(3–4), 243–249.
Mancinelli, A., Chiocchini, M., Chiocchini, R. A., & Romano, A. (2005). Biostratigraphy of Upper Triassic-Lower Jurassic carbonate platform sediments of the central-southern Apennines (Italy). Rivista Italiana di Paleontologia e Stratigrafia, 111, 271–283.
Márquez, L., Calvet, F., Arnal, I., Trifonova, E. (1994). Asociación de foraminíferos en la Formación Isábena, Triásico superior Sudpirenaico (Espana). Bol. R. Soc. Esp. Hist. Nat. (Sec. Geol.), 89(1–4), 189–197.
Martini, R., Rettori, R., Urosevic, D., & Zaninetti, L. (1995). Le genre Piallina Rettori et Zaninetti (foraminifere) dans des calcaires a turriglomines du Trias (Carnien) de Serbie orientale (domaine Carpatho-Balkanique). Revue de Paléobiologie, 14(2), 411–415.
Martini, R., Vachard, D., Zaninetti, L., Cirilli, S., Cornée, J. J., Lathuiliere, B., et al. (1997). Sedimentology, stratigraphy, and micropalaeontology of the Upper Triassic reefal series in Eastern Sulawesi (Indonesia). Palaeogeography, Palaeoclimatology, Palaeoecology, 128, 157–174.
Martini, R., Zaninetti, L., Lathuilliere, B., Cirilli, S., Cornée, J.-J., & Villeneuve, M. (2004). Upper Triassic carbonate deposits of Seram (Indonesia): palaeogeographic and geodynamic implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 206, 75–102.
Matzner, C. (1986). Die Zlambach-Schichten (Rhät) in den Nördlichen Kalkalpen: Eine Platform-Hang-Beckenentwicklung mit allochthoner Karbonatsedimentation [The Zlambach Beds (Rhaetian) of the Northern Limestone Alps: platform-slope-basin environments with allochthonous carbonate sedimentation]. Facies, 14, 1–104.
McRoberts, C. A., Furrer, H., & Jones, D. S. (1997). Palaeoenvironmental interpretation of a Triassic-Jurassic boundary section from Western Austria based on palaeoecological and geochemical data. Palaeogeography, Palaeoclimatology, Palaeoecology, 136, 79–95.
McRoberts, C. A., Krystyn, L., & Shea, A. (2008). Rhaetian (Late Triassic) Monotis (Bivalvia: Pectinoida) from the eastern Northern Calcareous Alps (Austria) and the end-Norian Crisis in pelagic faunas. Palaeontology, 51, 721–735.
McRoberts, C. A., & Newton, C. R. (1995). Selective extinction among end-Triassic European bivalves. Geology, 23, 102–104.
Michalík, J., Lintnerová, O., Gaździcki, A., & Soták, J. (2007). Record of environmental changes in the Triassic-Jurassic boundary interval in the Zliechov Basin, Western Carpathians. Palaeogeography, Palaeoclimatology, Palaeoecology, 244, 71–88.
Miconnet, P., Ciarapica, G., & Zaninetti, L. (1983). Faune a Foraminiferes du Trias superieur d’affinite Sud-Tethysienne dans l’Apennin meridional (Bassin de Lagonegro, Province de Potenza, Italie); comparaison avec l’Apennin septentrional. Revue de Paléobiologie, 2(2), 131–147.
Moix, P., Kozur, H. W., Stampfli, G. M., Mostler, H. (2007). New paleontological, biostratigraphic and paleogeographic results from the Triassic of the Mersin Mélange, SE Turkey. In Lucas, S. G., & Spielmann, J. A. (Eds): The Global Triassic, New Mexico Museum of Natural History and Science Bulletin, 41, 282–311.
Mostler, H., Scheuring, B., & Ulrichs, M. (1978). Zur Mega- und Mikrofauna und Mikroflora der Kössenerschichten (alpine Obertrias) wom Weissloferbach in Tirol unter besondere Berücksichtigung der in der suessi- und marshi- Zone auftretenden Conodonten. Schrift. Erdwiss. Komm. Österr. Akad. Wiss., 4, 141–174.
Muttoni, G., Kent, D. V., Jadoul, F., Olsen, P. E., Rigo, M., Galli, M. T., et al. (2010). Rhaetian magneto-biostratigraphy from the Southern Alps (Italy): constraints on Triassic chronology. Palaeogeography, Palaeoclimatology, Palaeoecology, 285, 1–16.
Muttoni, G., Kent, D. V., Olsen, P. E., Di Stefano, P., Lowrie, W., Bernasconi, S. M., et al. (2004). Tethyan magnetostratigraphy from Pizzo Mondello (Sicily) and correlation to the Late Triassic Newark astrochronological polarity time scale. GSA Bulletin, 116, 1043–1058.
Nagy, E. (1964). Foraminiferen aus dem anizischen Kalkstein des Mecsek-Gebirges. Földtani Közlöny, 94 (2).
Oberhauser, R. (1964). Zur Kenntnis der Foraminiferengattungen Permodiscus, Trocholina und Triasina in der alpinen Trias und ihre Einordnung zu den Archaedisciden. Verhandlungen der geologischen Bundesanstalt, 2, 196–210.
Oberhauser, R. (1967). Zur Vorkommen der Foraminiferengattung Austrocolomia in der ostalpinen Trias. Verhandlungen der geologischen Bundesanstalt, 1–2, 193–199.
Oravecz-Scheffer, A. (1965). Carnian foraminifera from the Bakony Mts. M. Áll. Földtani Intézet évi Jelentése, 181-217.
Oravecz-Scheffer, A. (1978). Middle Triassic microbiofacies in the lithological log of borehole Szentantalfa-1 [in Hungarian]. Magyar állami földtani intézet Institutum geologicum publicum hungaricum, 1978, 205–231.
Oravecz-Scheffer, A. (1987). Triassic foraminifers of the Transdanubian Central Range. Geologica Hungarica, 50, 3–134.
Orchard, M. J. (1991). Upper Triassic conodont biochronology and a new index species from the Canadian Cordillera. In Orchard, M. J., McCracken, A. D. (Eds): Ordovician to Triassic conodont paleontology of the Canadian Cordillera, Geological Survey of Canada Bulletin, 417, 299-335.
Orchard, M. J., & Tozer, E. T. (1997). Triassic conodont biochronology, its calibration with the ammonoid standard, and a biostratigraphic summary for the western Canada sedimentary basin. Bulletin of Canadian Petroleum Geology, 45, 675–692.
Pálfy, J., Demény, A., Haas, J., Carter, E. S., Görög, Á., Halász, D., et al. (2007). Triassic-Jurassic boundary events inferred from integrated stratigraphy of the Csővár section, Hungary. Palaeogeography, Palaeoclimatology, Palaeoecology, 244, 11–33.
Pantić, S. (1967). Mikropaleontološke karakteristike srednjeg i gornjeg trijasa planine Tare (zapadna Srbija) [Les caracteristiques micropolentologiques du Trias moyen et superieur de la montage Tara (Serbie occidentale)]. Vesnik Ser A Geol, 24-25, 245-254.
Pantić, S. (1972). Aeolisaccus amplimuralis n. sp. i Aeolisaccus gracilis n. sp. iz srednjeg trijasa jugoistočnih Dinarida. Bulletin du Museum d’Histoire Naturelle, ser. A., 27, 207–221.
Pevný, J., & Salaj, J. (1997). The Anisian-Ladinian boundary at Zakázané (Slovak Karst, Western Carpathians). Zemní plyn a nafta, 42(2), 97–149.
Peybernes, B., Martini, R., Taugourdeau-Lantz, J., & Zaninetti, L. (1988). Caracterisation micropaleontologique du Rhetien dans les Pyrenees Francaises entre Geronne et Mediterranee. Revue de Paléobiologie, 7, 137–161.
Peybernes, B., Martini, R., Zaninetti, L., Fréchengues, M. (1998). Larger benthic foraminifera. In Hardenbol, J., Thierry, J., Farley, M. B., Jacquin, J., de Graciansky, P.-C., & Vail, P. R. (Eds): Mesozoic and Cenozoic sequence stratigraphic framework of European basins. Chart 8: Triassic sequence stratigraphy/biochronostratigraphy. In de Graciansky, P.-C., Hardenbol, J., Jacquin, J., Vail, P. R. (eds): Mesozoic and Cenozoic sequence stratigraphy of European basins, SEPM Special Publication, 60.
Piller, W. (1978). Involutinacea (Foraminifera) der Trias und des Lias. Beiträge zur Paläontologie der Österreich, 5, 1–164.
Pirdeni, A. (1988). The Triassic benthic foraminifera of Albania. Revue de Paléobiologie, Vol. Spec. No. 2, Benthos’86, 145–152.
Pirini, C. (1966). Alcuni Foraminiferi dei calcari liassici di Montemerano—Grosseto. Palaeontolographia Italica, 60, 89–98, Pisa.
Placer, L. (1999). Contribution to the macrotectonic subdivision of the border region between Southern Alps and External Dinarides. Geologija, 41, 223–255.
Placer, L. (2008). Principles of the tectonic subdivision of Slovenia. Geologija, 51, 205–217.
Premoli-Silva, I. (1971). Foraminiferi anisici della regione Giudicariense (Trento). Rivista Italiana di Paleontologia, 77(3), 303–374.
Ramovš, A., & Kristan-Tollmann, E. (1967). Die Lias-Schichten von Stol (Karawanken). Geološki Vjesnik, 20(1966), 57–62.
Resch, W. (1972). Statistische Untersuchung der Foraminiferen-Faunen aus dem Profil Köveskál/W-Ungarn (Fassan – tieferes Cordevol). Mitteilungen der Gesselschaft für Geologische Bergbaustudien, 21, 513–538.
Rettori, R., Angiolini, L., & Muttoni, G. (1994). Lower and Middle Triassic foraminifera from the Eros Limestone, Hydra Island, Greece. Journal of Micropalaeontology, 13, 25–46.
Rigaud, S., Martini, R., Rettori, R., & Stanley, G. D, Jr. (2010). Stratigraphic potential of the Upper Triassic benthic foraminifers. Albertiana, 38, 34–39.
Rigo, M., De Zanche, V., Gianolla, P., Mietto, P., Preto, N., & Roghi, G. (2005). Correlation of Upper Triassic sections throughout the Lagonegro Basin. Bollettino della Societa Geologica Italiana, 124, 293–300.
Rigo, M., Galli, M. T., Jadoul, F. (2008). Conodont biostratigraphy of the Late Triassic in the western Bergamasc Alps (Italy). In Hoffmann, T., & Krystyn, L. (Eds): Upper Triassic subdivisions, zonations and events: meeting of the late IGCP 467 and STS, Abstracts and Excursion-Guide. Bad Goisern, 28. 9. 2. 10. 2008. Berichte der Geologischen Bundesanstalt, 76, 59–60.
Rigo, M., Galli, M. T., & Jadoul, F. (2009). Late Triassic biostratigraphic constraints in the Imagna Valley (western Bergamasc Alps, Italy). Albertiana, 37, 39–42.
Röhl, U., Dumont, T., von Rad, U., Martini, R., & Zaninetti, L. (1991). Upper Triassic Tethyan carbonates off Northwest Australia (Wombat Plateau, ODP Leg 122). Facies, 25, 211–252.
Rožič, B., Kolar-Jurkovšek, T., & Šmuc, A. (2009). Late Triassic sedimentary evolution of Slovenian Basin (eastern Southern Alps): description and correlation of the Slatnik Formation. Facies, 55, 137–155.
Ruhl, M., Veld, H., & Kürschner, W. M. (2010). Sedimentary organic matter characterization of the Triassic-Jurassic boundary GSSP at Kuhjoch (Austria). Earth and Planetary Science Letters, 292, 17–26.
Salaj, J. (1977). Contribution a la microbiostratigraphie du Trias des Carpathes Occidentales Tchécoslovaques. Actes du VI° Colloque Africain de Micropaléontologie – Tunis 1974. Annales des Mines et de la Géologie, 28, 103–127.
Salaj, J., Biely, A., & Bystrický, J. (1967). Trias-foraminiferen in den Westkarpaten. Geologické Práce, 42, 119–136.
Salaj, J., Borza, K., Samuel, O. (1983). Triassic foraminifers of the West Carpathians (213 pp.). Bratislava: Geologický Ústav Dionýza Štúra.
Salaj, J., Trifonova, E., Gheorgian, D., & Coroneou, V. (1988). The Triassic foraminifera microbiostratigraphy of the Carpathian – Balkan and Hellenic realm. Mineralia Slovenica, 20, 387–415.
Samuel, O. (1991). Late Triassic foraminifers from profile Turík, well Závod-81 and locality Ráztoka (West Carpathians). Západné Karpaty, séria paleontológia, 15, 7–47.
Schäfer, P. (1979). Fazielle Entwicklung und palökologische Zonierung zweier obertriadischer Riffstrukturen in den Nördlichen Kalkalpen (» Oberrhät « -riff-kalke, Salzburg). Facies, 1, 3–245.
Schäfer, P., & Senowbari-Daryan, B. (1978). Die Häufigkeitsverteilung der Foraminiferen in drei oberrhätischen Riff-Komplexen der Nördlichen Kalkalpen (Salzburg, Österreich). Verhandlungen der geologischen Bundesanstalt, 2, 73–96.
Schmid, S. M., Bernoulli, D., Fügenschuh, B., Matenco, L., Schefer, S., Schuster, R., et al. (2008). The Alpine-Carpathian-Dinaric orogenic system: correlation and evolution of tectonic units. Swiss Journal of Geosciences, 101, 139–183.
Senowbari-Daryan, B. (1983). Zur Gattung Pseudocucurbita Borza & Samuel, 1978 (=pro Cucurbita Jablonsky, 1973) und Beschreibung vergleichbarer problematischer Organismen aus der Obertrias de salpin-mediterranen Raumes (Beiträge zur Paläontologie und Mikrofazies der obertriadischen Riffe de salpin-mediterranen Gebietes, 25) (1). Rivista Italiana di Paleontologia, 88(2), 181–250.
Senowbari-Daryan, B., Rashidi, K., & Torabi, H. (2010). Foraminifera and their associations of a possibly Rhaetian section of the Nayband Formation in central Iran, northeast of Esfahan. Facies, 56, 567–596.
Sephton, M. A., Amor, K., Franchi, I. A., Wignall, P. B., Newton, R., & Zonneveld, J. P. (2002). Carbon and nitrogen isotope disturbances and an end-Norian (Late Triassic) extinction event. Geology, 30(12), 1119–1122.
Tanner, L., Ciarapica, G., Reggiani, L., Atudorei, V. (2007). Upper Triassic carbon isotope stratigraphy of the Lagonegro succession, Southern Apennines, Italy. In: Lucas, S. G., & Spielmann, J. A. (Eds): The Global Triassic. New Mexico Museum of Natural History and Science Bulletin, 41, 406–410.
Tanner, L. H., Lucas, S. G., & Chapman, M. G. (2004). Assessing the record and causes of Late Triassic extinctions. Earth Science Reviews, 65, 103–139.
Trifonova, E. (1965). Nodosaria ordinata sp. nov. from the Upper Triassic in Bulgaria. Review of the Bulgarian Geological Society, 2, 213–216.
Trifonova, E. (1977). Foraminiferen aus der Trias des Ostbalkans. Palaeontology, Stratigraphy and Lithology, 6, 47–63.
Trifonova, E. (1993). Taxonomy of Bulgarian Triassic foraminifera. II. Families Endothyriidae to Ophthalmidiidae. Geologica Balcanica, 23(2), 19–66.
Trifonova, E. (1994). Taxonomy of Bulgarian Triassic foraminifera. III. Families Spiroloculiniidae to Oberhausereliidae. Geologica Balcanica, 24(2), 21–70.
Trifonova, E., & Čatalov, G. (1983). On the stratigraphy and lithology of the Upper Triassic in the eastern part of the Trojan Balkan Mountain. Paleontology, Stratigraphy and Lithology, 18, 25–33.
Trifonova, E. T., & Čatalov, G. A. (1988). Foraminifera data on the stratigraphy of the Strandža facial type of Triassic (Strandža Mt., SE Bulgaria). Geologica Balcanica, 18(5), 79–84.
Türkmen, G., & Kazanci, N. (2010). Applications of various diversity indices to benthic macroinvertebrate assemblages in streams of a natural park in Turkey. Review of Hidrobiology, 3–2, 111–125.
Vachard, D., & Fontaine, H. (1988). Biostratigraphic importance of Triassic foraminifera and algae from south-east Asia [Valeur biostratigraphique dea algues et des foraminiferes du Trias du sud-est Asiatique]. Revue de Paléobiologie, 7(1), 87–98.
Van de Schootbrugge, B., Tremolada, F., Rosenthal, Y., Bailey, T. R., Feist-Burkhardt, S., Brinkhuis, H., et al. (2007). End-Triassic calcification crisis and blooms of organic-walled ‘disaster species’. Palaeogeography, Palaeoclimatology, Palaeoecology, 244, 126–141.
Velić, I. (2007). Stratigraphy and palaeobiology of Mesozoic benthic foraminifera of the Karst Dinarides (SE Europe). Geologia Croatica, 60(1), 1–113.
Velledits, F., & Blau, J. (2003). The Büdöskút Olistolith, an exotic limestone block from the Bükk Mountains (NE-Hungary). Facies, 48, 23–48.
Vettorel, M. (1988). Studio micropaleontologico e distribuzione litostratigrafica di alcuni gruppi di foraminiferi della Formazione di S. Cassiano (Trias superiore, Dolomiti Orientali). Annali Museo Civico di Rovereto: Sezione Archeologia, Storia, Scienze naturali, 4, 159–204.
Vlahović, I., Tišljar, J., Velić, I., & Matičec, D. (2005). Evolution of the Adriatic Carbonate Platform: palaeogeography, main events and depositional dynamics. Palaeogeography, Palaeoclimatology, Palaeoecology, 220, 333–360.
Ward, P. D., Garrison, G. H., Haggart, J. W., Kring, D. A., & Beattie, M. J. (2004). Isotopic evidence bearing on Late Triassic extinction events, Queen Charlotte Islands, British Columbia, and implications for the duration and cause of the Triassic/Jurassic mass extinction. Earth and Planetary Science Letters, 244, 589–600.
Williford, K. H., Ward, P. D., Garrison, G. H., & Buick, R. (2007). An extended organic-isotope record across the Triassic-Jurassic boundary in the Queen Charlotte Islands, British Columbia, Canada. Palaeogeography, Palaeoclimatology, Palaeoecology, 244, 290–296.
Wurm, D. (1982). Mikrofazies, Paläontologie und Palökologie der Dachsteinriffkalke (Nor) des Gosaukammes, Österreich [Microfacies, paleontology and palecology of the Dachstein reef limestones (Norian) of the Gosaukamm Range, Austria]. Facies, 6, 203–296.
Zaninetti, L. (1976). Les Foraminiferes du Trais. Essai de synthese et corrélation entre les domaines Mésogéens Européen et Asiatique. Rivista Italiana di Paleontologia, 82(1), 1–258.
Zaninetti, L., Altiner, D., Dager, Z., Ducret, B. (1982). Les Milioliporidae (Foraminiferes) dans le Trias superieur a facies recifal du Taurus, Turquie. II: Microfaunes associees. Revue de Paléobiologie, 1(2), 105–139.
Zaninetti, L., Martini, R., Dumont, T. (1992). Triassic foraminifers from sites 761 and 764, Wombat Plateau, Northwest Australia. In Rad, U. von, Haq, B. U., et al. (Eds): Proceedings of the Ocean Drilling Program, Scientific Results, 122, 427–436.
Acknowledgments
This research was financially supported by the Slovenian Research Agency (program numbers P1-0011 and Z1-9759). The first version of the paper was reviewed by R. Martini, H.-J. Gawlick and L. Krystyn. Upon well justified remarks, the data was re-evaluated and the paper rewritten. The reviewers thus deserve high praise and our gratitude for their advice. Facilities and technical staff of the Geological Survey of Slovenia are gratefully acknowledged as well.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial handling: A. G. Milnes.
Appendices
Appendix A
Foraminifera recognized in Mt. Slatnik and Mt. Kobla sections (see Figs. 9–12).
Ammodiscus parapriscus Ho, 1959
Gandinella falsofriedli (Salaj, Borza & Samuel, 1983)
Pilammina sulawesiana Martini, Vachard & Zaninetti, 1995
Kaeveria fluegeli (Zaninetti, Altiner, Dager & Ducret, 1982)
Palaeolituonella meridionalis (Luperto, 1965)
Alpinophragmium perforatum Flügel, 1967
Duotaxis birmanica Zaninetti & Brönnimann, in Brönnimann et al., 1975
Gaudryina triadica Kristan-Tollmann, 1964
Gaudryinella clavuliniformis Trifonova, 1967
Reophax rudis Kristan-Tollmann, 1964
Reophax asperus Cushman & Waters, 1928
Scherochorella eominutus (Kristan-Tollmann, 1964)
Ammobaculites corpulentus Efimova, 1974
“Trochammina” almtalensis Koehn-Zaninetti, 1969
“Trochammina” jaunensis Brönnimann & Page, 1966
Earlandia tintinniformis (Mišik, 1971)
Earlandia amplimuralis (Pantić, 1972)
“Tetrataxis” humilis Kristan, 1957
Endotriada tyrrhenica Vachard, Martini, Rettori & Zaninetti, 1994
Endotebanella bicamerata (Salaj in Salaj et al., 1967)
Involutina turgida Kristan, 1957
Trocholina umbo Frentzen, 1941
Trocholina crassa Kristan, 1957
Trocholina turris Frentzen, 1941
Aulotortus tumidus (Kristan-Tollmann, 1964) emend. Piller, 1978
Aulotortus sinuosus Weynschenk, 1956
Aulotortus friedli (Kristan-Tollmann, 1962) sensu Piller (1978)
Angulodiscus impressus Kristan-Tollmann, 1964
Angulodiscus communis Kristan, 1957
Auloconus permodiscoides (Oberhauser, 1964)
Triasina oberhauseri Koehn-Zaninetti & Brönnimann, 1969
Triasina hantkeni Majzon, 1954
Coronipora deminuta Blau, 1987
Coronipora etrusca (Pirini, 1966)
Coronipora austriaca (Kristan, 1957)
Planiinvoluta carinata Leischner, 1961
Planiinvoluta deflexa Leischner, 1961
Ophthalmidium carinatum (Leischner, 1961)
Ophthalmidium leischneri (Kristan-Tollmann, 1962)
Ophthalmidium exiguum Koehn-Zaninetti, 1969
?Ophthalmidium walfordi Häusler, 1887
Hydrania dulloi Senowbari-Daryan, 1983
Galeanella tollmanni (Kristan, 1957)
Bispiranella salaji (Samuel & Borza, 1981) comb. nov.
Miliolipora cuvillieri Brönnimann & Zaninetti in Brönnimann et al., 1971
Miliolipora tamarae Gale, Rettori & Martini in Gale et al., 2011
Cucurbita cylindrica (Senowbari-Daryan, 1983)
Agathammina austroalpina Kristan-Tollmann & Tollmann, 1964
“Sigmoilina” schaeferae Zaninetti, Altiner, Dager & Ducret, 1982
Miliolechina stellata Zaninetti, Ciarapica, Cirilli & Cadet, 1985
Diplotremina subangulata Kristan-Tollmann, 1960
Diplotremina placklesiana Kristan-Tollmann, 1960
?Diplotremina astrofimbriata Kristan-Tollmann, 1960
Duostomina turboidea Kristan-Tollmann, 1960
?Duostomina biconvexa Kristan-Tollmann, 1960
Duostomina sp. A
Variostoma coniforme Kristan-Tollmann, 1960
Variostoma catilliforme Kristan-Tollmann, 1960
Variostoma cochlea Kristan-Tollmann, 1960
Variostoma falcata Kristan-Tollmann, 1973
?Variostoma helicta (Tappan, 1951)
Austrocolomia canaliculata (Kristan-Tollmann, 1964) sensu Oberhauser, 1967
?Austrocolomia cordevolica Oberhauser, 1967
?Cryptoseptida klebelsbergi (Oberhauser, 1960)
Nodosaria ordinata Trifonova, 1965
Pseudonodosaria obconica (Reuss, 1868)
Appendix B
Conodonts recognized in Mt. Slatnik and Mt. Kobla sections (see Figs. 3–4).
Epigondolella ex gr. E. abneptis (Huckriede, 1958)
Epigondolella bidentata Mosher, 1968
Epigondolella ex gr. E. postera (Kozur & Mostler, 1971)
Misikella buseri Kolar-Jurkovšek, 2011
Misikella hernsteini (Mostler, 1967)
Misikella posthernsteini Kozur & Mock, 1974
Misikella sp. A
Misikella sp. B
Misikella sp.
Norigondolella steinbergensis (Mosher, 1968)
Oncodella paucidentata (Mostler, 1967)
Parvigondolella andrusovi Kozur & Mock, 1972
Parvigondolella lata Kozur & Mock, 1974
Zieglerioconus rhaeticus Kozur and Mock, 1991
Rights and permissions
About this article
Cite this article
Gale, L., Kolar-Jurkovšek, T., Šmuc, A. et al. Integrated Rhaetian foraminiferal and conodont biostratigraphy from the Slovenian Basin, eastern Southern Alps. Swiss J Geosci 105, 435–462 (2012). https://doi.org/10.1007/s00015-012-0117-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00015-012-0117-1