
Abstract
An Early Triassic (Griesbachian) gastropod fauna is reported from South China (Shanggan, Guangxi) and consists of four species: Bellerophon abrekensis, Wannerispira shangganensis Kaim & Nützel sp. nov., Naticopsis sp., and Palaeonarica guangxinensis Kaim & Nützel sp. nov. The taxon Wannerispira Kaim & Nützel nom. nov. replaces Pagodina Wanner non Van Beneden. This is the first report of Bellerophon abrekensis from China. Previously, it was only known from its type locality in Far East Russia. Wannerispira shangganensis sp. is the first certain Triassic report of the Permian subfamily Neilsoniinae and represents a holdover taxon. The neritimorph Palaeonarica is reported for the first time from the Early Triassic and this is the oldest occurrence of this genus. Compared with other Griesbachian gastropods, the present material is relatively well preserved so that the taxonomy rests on rather firm ground. Very few nominal taxa have been reported from the Griesbachian and therefore the present report presents substantial additional information about gastropods from the aftermath of the end-Permian mass extinction event. The gastropod association from Shanggan shares one species with Primorye, Far East Russia (B. abrekensis). Two species, W. shangganensis and P. guangxinensis, closely resemble specimens reported from the Griesbachian of Oman. This could suggest that Griesbachian gastropod faunas of the Tethys were rather homogenous although the data are still scarce.
Similar content being viewed by others


Introduction
The fauna in the immediate aftermath of the end-Permian mass extinction event is of greatest interest because it is part of the extinction pattern itself, in that it defines which taxa actually went extinct and which survived, and how much “disastrous” early post-extinctions communities actually were. Although gastropods have been considered to be relatively extinction resistant (Erwin and Signor 1990), only about 80 nominate gastropod species representing roughly 40 genera (excluding Lazarus taxa) are known from the Early Triassic, which is a very small number when compared with gastropod diversity during the remaining Triassic (Nützel 2005). The Griesbachian diversity is even lower and the preservation is generally very poor in this interval of time in addition to the general scarcity of fossiliferous exposures of that age. Therefore, any additional information about reasonably well-preserved gastropods from this critical time interval is extremely useful. Although the present gastropod fauna from Shanggan (NW Guangxi, South China) comprises only four species, it provides important biostratigraphic data that have direct implications for the evolution of gastropods during the Griesbachian.
Early Triassic gastropods from China have been studied by Yü et al. (1963), Wang and Xi (1980), Pan (1982), Tong and Erwin (2001) and Pan et al. (2003). About 34 gastropod species have been reported from the Early Triassic of China, 11 of them, however, are in open nomenclature. The species are attributed to 27 genera but many attributions need critical evaluation or re-sampling because preservation of this material is usually poor.
Locality and stratigraphy
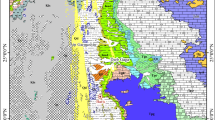
The new gastropod fauna was found near Shanggan, which is located a few kilometres north of Leye in Guangxi Province of South China (Fig. 1). In Early Triassic, this area was located South of the Yangtze carbonate platform in the Nanpanjiang Basin (Galfetti et al. 2007, 2008). The basin was widely open through present-day Guangxi and Yunnan provinces and extended into central Guizhou (Galfetti et al. 2007, 2008). Palaeogeographically it belongs to the South China Block, which occupied an equatorial position at the boundary between the Tethyan and Panthalassian domains in Early and Middle Triassic times (Gilder et al. 1995).

Location and stratigraphical occurrence of the gastropod fauna from Shanggan, Guangxi, South China. a Shanggan section. The fossiliferous lense SHA-3 occurs ca. 10 m above the Permian–Triassic boundary. Opb: ‘Ophiceras beds’; Pcb: ‘Proptychites candidus beds’. Modified from Brühwiler et al. (2008). b Palaeogeographical world map of Early Triassic times with South China (star) and other gastropod-bearing localities (shaded circles) indicated. Modified after Brayard et al. (2006). c Geographical location of Shanggan village in Leye County, northern Guangxi Province
The Lower Triassic mixed carbonate-siliciclastic series of the Luolou Formation in Guangxi overlies Late Permian skeletal reef limestones of the Wujiaping Formation (Ovtcharova et al. 2006; Galfetti et al. 2007, 2008). Latest Permian and the Griesbachian and lower Dienerian part of the Luolou Formation at Shaggan are well exposed, fossiliferous, and continuously exposed within a single fault bounded block. This section was studied by Brühwiler et al. (2008), who documented in detail the local ammonoid sequence. The studied gastropods come from a coquinoid lense intercalated within the uppermost portion of the basal microbial limestone (sample SHA 3, see Brühwiler et al. 2008). This coarse biocalcarenite is trapped between the uppermost microbial domes, and displays obvious signs of sorting, either by waves or by currents. This coquinoid lense contains abundant small bivalves, gastropods, brachiopods, and very abundant, small-sized specimens of Ophiceras sp. indet., which indicate a late Griesbachian age (Brühwiler et al. 2008). Most specimens are broken, but not abraded, which suggests that this material was not remobilized many times by waves or currents before being eventually deposited between the microbial domes. The limestone is hard and splintery. It has a beige yellow colour. The gastropod and ammonoid shells are calcitized.
Systematic palaeontology (by Andrzej Kaim & Alexander Nützel)
Institutional abbreviations: PIMUZ, Paläontologisches Institut und Museum der Universität Zürich, Switzerland.
Class Gastropoda Cuvier, 1798
Order Amphigastropoda Simroth, 1906
Superfamily Bellerophontoidea McCoy, 1852
Family Bellerophontidae McCoy, 1852
Genus Bellerophon Montfort, 1808
Type species: Bellerophon vasulites Montfort, 1808; Stringocephalen-Kalk, Middle Devonian, Germany.
Bellerophon abrekensis Kaim, 2009, Fig. 2

Bellerophon abrekensis Kaim, 2009 from Griesbachian (Early Triassic) sample SHA-3, Shanggan, Guangxi, South China. (a, d) PIMUZ 28178 in adapertural (a) and left lateral (d) views. (b, c, e, f) PIMUZ 28177 in ventral (b), ventro-abapertural (c), ventro-lateral (e), and right lateral (f) views. All scale bars 2 mm
1899 Bellerophon sp. indet.—Bittner: p. 28, pl. 4, figs. 26–28.
non 1936 Bellerophon asiaticus sp. nov.—Wirth: p. 441, fig 14: 7a, b.
1947 Bellerophon asiaticus Wirth—Kiparisova: p. 121, pl. 24, figs. 6, 7.
*2009 Bellerophon abrekensis sp. nov.—Kaim: p. 147, figs. 145–136.
Material: 8 specimens.
Measurements:
Specimen number | Length (mm) | Width (mm) |
|---|---|---|
PIMUZ 28176 | 9.6 | 7.8 |
PIMUZ 28178 | 12.0 | 8.6 |
PIMUZ 28179 | 9.8 | 7.8 |
Description: Shell globular, longer than wide; shell ornamented with collabral ribs bent posteriorly towards the selenizone. In adults, ribs are differentiated into sets of stronger and weaker ribs. No spiral ornamentation present; selenizone long and slightly elevated, slit short; pseudoumbilicus small.
Discussion: B. abrekensis has been described by Kaim (2009) based on well-preserved material from Late Griesbachian of South Primorye, Far East Russia. B. abrekensis has previously been known as “Bellerophon sp. indet.” of Bittner (1899) from the Lower Triassic of the Ussuri region, Far East Russia. It was misidentified as “Bellerophon asiaticus” by Kiparisova (1947). B. asiaticus Wirth, 1936 has been redefined by Yochelson and Hongfu (1985) and was assigned to the genus Retispira. B. asiaticus has also been reported from China by Yü et al. (1963). However, the description of the specimen from Sichuan Province is extremely brief and the illustration is very poor so that any taxonomic assignment of this material is speculative. Kaim (2009) compared B. abrekensis with Bellerophon panxianensis Yü in Wang and Xi (1980) and stated that the holotype of B. panxianensis is flattened laterally, crushed, and generally of poor preservation so that little taxonomic information can be inferred from it. Therefore, it is considered to represent a nomen dubium. Another Chinese bellerophontid is Stachella granaticarinata Wang in Wang & Xi, 1980. We examined a cast of the holotype of this species. It is an internal mould of a bellerophontid and therefore it also represents a nomen dubium. It may or may not be conspecific with B. abrekensis. The shells from Shanggan are not perfectly preserved but they display several characters identical to small-sized shells of B. abrekensis from its type locality. We could not observe any character differentiating them and we assign the specimens from Shanggan to B. abrekensis.
Order Vetigastropoda Salvini-Plawen, 1980
Superfamily Eotomarioidea Wenz, 1938
Family Eotomariidae Wenz, 1938
Subfamily Neilsoniinae Knight, 1956
Genus Wannerispira Kaim & Nützel nom. nov. pro Pagodina Wanner, 1941
Taxonomic remarks: Wanner (1941, p. 34) erected the genus Pagodina for a group of gastropods from the Permian of Timor. The name Pagodina, however, was previously used by Van Beneden (1853) for a crustacean. We therefore here propose the name Wannerispira new name as a replacement for Pagodina Wanner, 1941 non Van Beneden, 1853 nec Stabile, 1864. Wannerispira is named in honor of C. Wanner who originally described this genus. The type species of Wannerispira is Pagodina typus Wanner, 1941 by original designation.
Discussion: The Neilsoniinae unite small trochoid-shaped eotomariids. The included genera of the Neilsoniinae differ from each other only in minor details. The shells from Shanggan resemble Pagodina Wanner, 1941 (re-named as Wannerispira herein) and Apachella Winters, 1956. Apachella typically possesses an axial ornamentation above the periphery while shells having no axial ornament but usually some additional spiral ribs on its lateral flank are classified as Pagodina (=Wannerispira). Both genera, are distinguishable from the other neilsoniids (e.g. Neilsonia and Peruvispira) in having spiral ribs on the shell base. Wannerispirashangganensis sp. nov. has neither axial ornamentation nor additional spiral ribs on the whorl face.
Wannerispirashangganensis Kaim & Nützel sp. nov.
Fig. 3

Wannerispira shangganensis sp. nov. from Griesbachian (Early Triassic) sample SHA-3, Shanggan, Guangxi, South China. (a) PIMUZ 28180 in lateral view. (b, c, h) PIMUZ 28186 (holotype) in lateral (b), apertural (c), and apical (h) views. (d) PIMUZ 28287 in apertural view. (e) PIMUZ 28188 in lateral view. (f) PIMUZ 28181 in lateral view. (g) PIMUZ 28185 in lateral view. All scale bars 2 mm
Derivatio nominis: After the type locality, Shanggan.
Material: 19 specimens.
Holotype: PIMUZ 28186
Type locality: Shanggan, Guangxi Province, South China.
Type horizon: Layer SHA-3, Ophiceras beds, Griesbachian, Lower Triassic Luolou Formation.
Measurements:
Specimen number | No. of whorls | Height (mm) | Diameter (mm) |
|---|---|---|---|
PIMUZ 28180 | 4 | 5.5 | 4.7 |
PIMUZ 28181 | 4 | 5.35 | 4.7 |
PIMUZ 28182 | 5 | 10.03 | 9.8 |
PIMUZ 28183 | – | – | 8.8 |
PIMUZ 28184 | 3 | 4.4 | 4.1 |
PIMUZ 28185 | 3 | 4.1 | 3.4 |
PIMUZ 28186 | 5 | 5.8 | 4.3 |
Diagnosis: Species of the Neilsoniinae with no axial ornament and 9–10 spiral ribs on its base. Shell flanks more convex than in W. dongluoensis (Pan and Yu 1993).
Description: Shell trochoidal, relatively high spired; two earliest whorls rounded and smooth, then selenizone appears; selenizone concave between a pair of spiral ribs; lower rib located at shell periphery although it moves above the periphery at adolescent or adult whorls in some specimens; demarcation between lateral flank and base slightly angulated with weak spiral rib; base ornamented with 9–10 fine spiral ribs; growth lines visible only at their intersections with basal spiral ribs, otherwise no axial ornamentation observed; aperture D-shaped; peristome uninterrupted; small umbilical chink seems to be present.
Discussion: Neilsoniinae are represented mainly by Late Palaeozoic (mostly Permian) taxa. The attribution of the Norian (Late Triassic) monospecific genus Pareuryalox Haas, 1953 to this subfamily seem to be at least disputable as it has littorinform and non-carinate shells. If Pareuryalox is excluded there is no other published record of Neilsoniinae from the Triassic. It seems, however, that Ananias sp. illustrated but not formally described by Wheeley and Twitchett (2005) from the Griesbachian of Oman would be better classified as Wannerispira. Unfortunately, the poor preservation of the material from Oman precludes further comparisons with our species. W. shangganensis is very similar to W. dongluoensis (Pan and Yu 1993) reported from Upper Permian Changxing Formation, Dongluo Coal Mine, Guangxi Zhuang Region of South China by Pan and Yu (1993). Our species differs only in having more rounded shell flanks while in W. dongluoensis, the shell flanks between selenizone and suture are only slightly convex. Wannerispira shangganensis also resembles the Late Triassic Murchisonia subeuglypha Krumbeck, 1924 from Timor (Krumbeck 1924), which may also represent a species of Neilsoniinae. However, examination of type material is necessary to substantiate this possible affiliation. It is remarkable that the Triassic Murchisonia subeuglypha and the Permian type species of Wannerispira have both been described from Timor.
Order Neritimorpha Koken, 1896
Superfamily Neritoidea Rafinesque, 1815
Family Naticopsidae Waagen, 1880
Genus Naticopsis M’Coy, 1844
Type species: Naticopsis phillipsii M’Coy, 1844 from the Lower Carboniferous of Ireland
Naticopsis sp., Fig. 4a, b, d, e

Naticopsis sp. (a, b, d, e) and Palaeonarica guangxinensis sp. nov. (c, f) from Griesbachian (Early Triassic) sample SHA-3, Shanggan, Guangxi, South China. (a) PIMUZ 28189 in lateral view. (b) PIMUZ 28190 in apical view. (c, f) PIMUZ 28192 (holotype) in lateral (c) and apical (f) views. (d, e) PIMUZ 28191 in lateral (d) and apical (e) views. All scale bars 1 mm
Material: 6 specimens
Description: Shell globose with rounded whorls embracing majority of the preceding whorl; suture weakly incised; shell surface smooth.
Discussion: The shells under consideration are relatively poorly preserved and represent probably juveniles. The gross shell morphology is reminiscent of Naticopsis utahensis Batten & Stokes, 1986 from the Lower Triassic Sinbad Limestone of the USA (Batten and Stokes 1986). Neither aperture nor protoconch are preserved and therefore we leave the specimens at hand in open nomenclature.
Superfamily Neritopsoidea Gray, 1847
Family Palaeonaricidae Bandel, 2007
Genus Palaeonarica Kittl, 1892
Type species: Naticella pyrulaeformis Klipstein, 1843, Carnian (Late Triassic) Cassian Formation, Italy; subsequent designation by Cossmann (1915: 85).
Palaeonaricaguangxinensis Kaim & Nützel sp. nov.
Fig. 4c, f
Derivatio nominis: After Guangxi Province.
Material: 2 specimens
Holotype: PIMUZ 28192.
Type locality: Shanggan, Guangxi Province, South China.
Type horizon: Layer SHA-3, Ophiceras beds, Griesbachian, Lower Triassic Loulou Formation.
Measurements: The holotype (PIMUZ 28192) is 6.1 mm high and 7.2 mm in diameter.
Diagnosis: Neritoid shell, flat-topped, low-spired shell with strong spiral ribs, which are evenly distributed over the entire shell surface.
Description: Small, rapidly expanding neritoid shell; first two whorls rounded and smooth (protoconch?); next 1.25 whorls flat-topped and ornamented with strong spiral ribs; four ribs present on spire whorls; seven spiral ribs on last whorl; ribs regularly distributed over entire shell surface; aperture and umbilical region unknown.
Discussion: Palaeonarica guangxinensis is the first species of the genus described from Early Triassic. As stressed by Nützel and Erwin (2004) Palaeonarica is a gastropod typical for the Carnian and rare in the Norian. Palaeonaricaguangxinensis sp. nov. may belong to a different but similar and closely related genus. Early Triassic specimens with well-preserved aperture and/or protoconch are necessary to confirm our generic assignment. A shell similar to P. guangxinensis has been identified as Yunnania? sp. from the Griesbachian of Oman (Wheeley and Twitchett 2005) and might be conspecific. The trochoid genus Yunnania is more high-spired and has less rapidly increasing whorls. The shells of P. guangxinensis differ from the other species of Palaeonarica in having the spiral ribs evenly distributed on the shell flank. The other species classified to this genus have the spaces between adapical ribs are wider than the ones located abapically (see Bandel 2007). Pan (1977) described two species of Palaeonarica from the Carnian of Yunnan, China; however, they are more reminiscent of margaritid-like trochoids (rather than of Neritimorpha) in having an inflated trochoid shell, which is not flattened adapically.
Discussion
As mentioned above, the relatively good preservation of the studied gastropod fauna facilitates a reasonable taxonomic assignment, which is rarely possible for Griesbachian gastropods. The presence of the bellerophontid Bellerophon abrekensis underlines that bellerophontids were probably globally distributed during the early Triassic. They survived the end-Permian crisis with several genera and were reported from N America, Europe, Greenland, Asia (China, India, Pakistan, Siberia, and Primorye), Arabia, and Australia but became extinct within the Early Triassic (Yochelson and Hongfu 1985 and references therein). Thus Bellerophontoidea are considered as a typical example for the Dead Clade Walking phenomenon sensu Jablonski (2001). Species assignment to Bellerophon abrekensis from Far East Russia (where it is very abundant) shows that this species had a relatively wide geographic distribution. Wannerispira (alias Pagodina) represents another Palaeozoic holdover. It represents the only certain Triassic representative of the subfamily Neilsoniinae. Neilsoniinae comprise Late Palaeozoic (mostly Permian) taxa (e.g., Knight et al. 1960). The attribution of the Norian (Late Triassic) monospecific genus Pareuryalox Haas, 1953 to this subfamily is almost certainly wrong. Wheeley and Twitchett (2005) illustrated a similar, possibly conspecific Griesbachian specimen from Oman (as Ananias sp.).
The present Naticopsis species is a typical representative of this genus. Naticopsis is present in many Early Triassic gastropod faunas and plays an important role during the recovery (Nützel 2005). It is diverse in Late Palaeozoic faunas and seems to have a rich fossil record throughout the Triassic. The spirally ribbed neritimorph genus Palaeonarica is reported for the first time from the Early Triassic and this is the oldest known occurrence of this genus. Wheeley and Twitchett (2005) reported a similar shell from the Griesbachian of Oman (as Yunnania? sp.), which is congeneric and probably even conspecific. Thus two of the four species present in the studied fauna seem also to be present in the Griesbachian of Oman. This could suggest that the Griesbachian gastropod fauna is quite homogenous within the Tethys. The gastropods studied here have a size from about 5-10 mm. This is not unusual small for gastropods and seems to corroborate that the Lilliput Effect was not as pronounced as previously suggested (Brayard et al. 2010, in press; Nützel et al. 2010, in press). The fact that the studied material from Shanggan was transported and size sorted does not invalidate the statement that 10 mm is not unusually small for a gastropod. It may well be that larger gastropods were present in the source area prior to transport.
Conclusions
The present Early Triassic gastropod association from the Guangxi Province is remarkably well preserved when compared to other Griesbachian gastropod faunas. It is composed of four species: the bellerophontid Bellerophon abrekensis Kaim, 2009, the eotomariid Wannerispira shangganensis Kaim & Nützel sp. nov., the naticopsid Naticopsis sp., and the palaeonaricid Palaeonarica guangxinensis Kaim & Nützel sp. nov. Wannerispira shangganensis is the most abundant gastropod in this collection and represents the first known post-Permian species of the Neilsoniinae. Palaeonarica guangxinensis is the oldest known representative of the genus Palaeonarica and the family Palaeonaricidae, which previously have been known from the Late Triassic.
Bellerophon abrekensis occurs in Far East Russia, Primorye (Kaim 2009) and in the present collection. Wannerispira shangganensis and Palaeonarica guangxinensis closely resemble specimens, which have been reported from the Griesbachian of Oman. This suggests that Early Triassic gastropod faunas of the Tethys were quite homogenous although the data are admittedly still very scarce. Naticopsis is a cosmopolitan gastropod genus known from the majority of Early Triassic gastropod faunas. The fact that a small faunule comprising four species contains two new taxa suggests that the Griesbachian is still poorly sampled and that there is a need to sample more localities with well-preserved gastropods in order to understand the evolutionary fate of gastropods in the aftermath of the end-Permian mass extinction event.
References
Bandel, K. (2007). Description and classification of Late Triassic Neritimorpha (Gastropoda, Mollusca) from the St Cassian Formation, Italian Alps. Bulletin of Geosciences, 83, 215–274.
Batten, R. L., & Stokes, W. L. (1986). Early Triassic gastropods from the Sinbad Member of the Moenkopi Formation, San Rafael Swell, Utah. American Museum Novitates, 2864, 1–33.
Bittner, A. (1899). Versteinerungen aus den Trias-Ablagerungen des Süd-Ussuri-Gebietes in der Ostsibirischen Küstenprovinz. Mémoires du Comité Géologique, 7(4), 1–35.
Brayard, A., Bucher, H., Escarguel, G., Fluteau, F. & Bourquin, S. (2006). The Early Triassic ammonoid recovery: Paleoclimatic significance of diversity gradients. Palaeogeography, Palaeoclimatology, Palaeoecology, 239, 374–395.
Brayard, A., Nützel, A., Stephen, D. A., Bylund K. G., Jenks, J., & Bucher, H. (2010). Gastropod counter-evidences for the Early Triassic Lilliput effect. Geology (in press).
Brühwiler, T., Brayard, A., Bucher, H., & Guodun, K. (2008). Griesbachian and Dienerian (Early Triassic) ammonoid faunas from northwestern Guangxi and southern Guizhou (South China). Palaeontology, 51, 1151–1180.
Cossmann, M. (1915). Essais de Paléoconchologie comparée (292 pp). Livraison 10. Paris: Cossmann & Rudeval.
Cuvier, G. (1797). Tableau élémentaire de l’histoire naturelle des animaux (710 pp). Paris: Baudouin.
Erwin, D. H. & Signor, P. W. (1990). Extinction in an extinction-resistant clade: The evolutionary history of the Gastropoda. In E. C. Dudley (Ed.), The unity of evolutionary biology 1. Proceedings of the fourth international Congress of systematics and evolutionary biology (pp. 152–160). Portland: Dioscorides Press.
Galfetti, T., Bucher, H., Brayard, A., Hochuli, P. A., Weissert, H., Guodun, K., et al. (2007). Late Early Triassic climate change: insights from carbonate carbon isotopes, sedimentary evolution and ammonoid paleobiogeography. Palaeogeography, Palaeoclimatology, Palaeoecology, 243, 394–411.
Galfetti, T., Bucher, H., Martini, R., Hochuli, P. A., Weissert, H., Crasquin-Soleau, S., et al. (2008). Evolution of Early Triassic outer platform paleoenvironments in the Nanpanjiang Basin (South China) and their significance for the biotic recovery. Sedimentary Geology, 204, 36–60.
Gilder, S. A., Coe, R. S., Wu, H. R., Kunag, G. D., Zhao, X. X., & Wu, Q. (1995). Triassic paleomagnetic data from South China and their bearing on the tectonic evolution of the western circum-Pacific region. Earth and Planetary Science Letters, 131, 269–287.
Gray, J. E. (1847). A list of genera of recent Mollusca, their synonyma and types. Proceedings of the Zoological Society of London, 15, 129–182.
Haas, O. (1953). Mesozoic invertebrate faunas of Peru. Bulletin of the American Museum of Natural History, 101, 1–328.
Jablonski, D. (2001). Lessons from the past: Evolutionary impacts of mass extinctions. Proceedings of the National Academy of Sciences, 98, 5393–5398.
Kaim, A. (2009). Gastropods. In Y. Shigeta, et al. (Eds.), The Lower Triassic System in the Abrek Bay area, South Primorye, Russia. National Museum of Nature and Science Monographs, 38, 141–156.
Kiparisova, L. (1947). Klass Gastropoda. Briuhonogiye. In L. Kiparisova (Ed.), Atlas of the guide forms of the fossil faunas of the USSR (Vol. 7, pp. 120–124). Leningrad: State Editorial Office for Geological Literature USSR Ministry of Geology.
Kittl, E. (1892). Die Gastropoden der Schichten von St. Cassian der südalpinen Trias. II. Theil. Annalen des Kaiserlich-Königlichen Naturhistorischen Hofmuseums, 7, 35–97.
von Klipstein, A. (1843). Beiträge zur geologischen Kenntnis der östlichen Alpen (311 pp). Giessen: G.F. Heyer’s Verlag.
Knight, J. B. (1956). New families of Gastropoda. Journal of the Washington Academy of Sciences, 46, 41–42.
Knight, J. B., Cox, L. R., Keen, A. M., Batten, R. L., Yochelson, E. L., & Robertson, R. (1960). Systematic descriptions. In J. B. Knight, et al. (Eds.), Treatise on invertebrate paleontology. Part I: Mollusca 1 (pp. 169–351). Lawrence: Geological Society of America, Inc. and University of Kansas Press.
Koken, E. (1896). Die Gastropoden der Trias um Hallstadt. Jahrbuch der kaiserlich-königlichen geologischen Reichsanstalt, 46, 37–126.
Krumbeck, L. (1924). Die Brachiopoden, Lamellibranchiaten und Gastropoden der Trias von Timor II. Paläontologischer Teil. Paläontologie von Timor, XIII Lieferung, 22, 1–275.
McCoy, F. (1844). A synopsis of the characters of Carboniferous Limestone Fossils of Ireland (274 pp). Dublin: McGloshan and Gill.
McCoy, F. (1852). A synopsis of the classification of the of the British Palaeozoic rocks [By the Rev. Adam Sedgwick] with a systematic description of the British Palaeozoic fossils in the Geological Museum of the University of Cambridge [by Frederick McCoy] with figures of the new and imperfectly known species (661 pp.). London: Parker & Son.
de Montfort, P. D. (1808). Conchyliologie systématique et classification méthodique de coquilles; offrant leurs figures, leur arrangement générique, leurs descriptions caractéristiques, leurs noms; ainsi que leur synonymie en plusieurs langues. Coquilles univalves, cloisonnées. F. Schoell, Paris, 409 pp.
Nützel, A. (2005). Recovery of gastropods in the Early Triassic. In D. Bottjer & J.-C. Gall (Eds.), The biotic recovery from the end-Permian mass extinction. Comptes Rendus, Palevol, 4, 501–515.
Nützel, A., & Erwin, D. H. (2004). Late Triassic (Late Norian) gastropods from the Wallowa terrane (Idaho, USA). Paläontologische Zeitschrift, 78, 361–416.
Nützel, A., Mannani, M., Senowbari-Daryan, B., & Yazdi, M. (2010). Gastropods from the Late Triassic Nayband Formation (Iran), their relationships to other Tethyan faunas and remarks on the Triassic gastropod body size problem. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. doi:10.1127/0077-7749/2010/0049.
Ovtcharova, M., Bucher, H., Schaltegger, U., Galfettin, T., Brayard, A., & Guex, J. (2006). New Early to Middle Triassic U–Pb ages from South China: Calibration with ammonoid biochronozones and implications for the timing of the Triassic biotic recovery. Earth and Planetary Science Letters, 243, 463–475.
Pan, H. Z. (1977). Mesozoic and Cenozoic gastropods from Yunnan. In Mesozoic Fossils of Yunnan fasc. 2 (pp. 83–152). Peking: Science Press.
Pan, H. Z. (1982). Triassic marine fossil gastropods from SW China. Bulletin of Nanjing Institute of Geology and Palaeontology, Academia Sinica, 4, 153–188.
Pan, H. Z., Erwin, D. H., Nützel, A., & Zhu, X. S. (2003). Jiangxispira, a new gastropod genus from the Early Triassic of China with remarks on the phylogeny of the Heterostropha at the Permian/Triassic boundary. Journal of Paleontology, 77, 44–49.
Pan, Y. & Yu, W. (1993). Permian Gastropoda of China (68 pp). Beijing: Ocean Press.
Rafinesque, C. S. (1815). Analyse de la nature, ou tableau de l′univers et des corps organisées (223 pp). Palerme: L’auteur & Jean Barravecchia.
von Salvini-Plawen, L. (1980). A reconsideration of systematics in the Mollusca (phylogeny and higher classification). Malacologia, 19, 249–278.
Simroth, H. (1906). Versuch einer neuen Deutung der Bellerophontiden. Sitzungsberichte der naturforschenden Gesellschaft zu Leipzig, 1905, 3–8.
Stabile, G. (1864). Mollusques terrestres vivants du Piémont. Atti della Società di scienze naturali Milano, 7, 1–141.
Tong, J., & Erwin, D. H. (2001). Triassic gastropods of the Southern Qinling Mountains, China. Smithsonian Contributions to Paleobiology, 92, 1–47.
Van Beneden, P. J. (1853). Note sur un nouveau genre de Crustacé parasite (Pagodina). Bulletins de l’Académie royale des Sciences de Belgique, 20, 482.
Waagen, W. (1880). Salt-Range Fossils. I. Productus Limestone Fossils. 2. Pisces-Cephalopoda: Supplement Gasteropoda. Memoirs of the Geological Survey of India (Palaeontologia Indica), 13(1), 1–183.
Wang, H. & Xi, Y. (1980). Late Permian and Early Triassic gastropods of western Guizhou. In Nanking Institute of Geology and Palaeontology (Ed.), Stratigraphy and palaeontology of Upper Permian coal bearing Formation in Western Guizhou and Eastern Yunnan, China (pp. 195–232). Beijing: Academia Sinica (Science Press).
Wanner, C. (1941). Neue Beiträge zur Gastropodenfauna des Perm von Timor. Geological Expedition to the Lesser Sunda Islands, 4, 1–71.
Wenz, W. (1938–1944). Gastropoda. Teil I: Allgemeiner Teil und Prosobranchia. In O. H. Schindewolf (Ed.), Handbuch der Paläozoologie Band 6 (1639 pp). Berlin: Bornträger.
Wheeley, J. R., & Twitchett, R. J. (2005). Palaeoecological significance of a new Griesbachian (Early Triassic) gastropod assemblage from Oman. Lethaia, 38, 37–45.
Winters, S. S. (1956). New Permian gastropod genera from eastern Arizona. Journal of the Washington Academy of Sciences, 46, 44–45.
Wirth, E. (1936). Beiträge zur Kenntnis der Trias in Provinz Szechuan, West China. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie Beilage-Band, 75, 412–446.
Yochelson, E. L., & Hongfu, Y. (1985). Redescription of Bellerophon asiaticus Wirth (Early Triassic: Gastropoda) from China, and a survey of Triassic Bellerophontacea. Journal of Paleontology, 59, 1305–1319.
Yü, W., Wang, H. J., & Li, Z. S. (1963). Fossil Gastropoda from China (362 pp). Peking: Publishing House of Science.
Acknowledgments
Thomas Galfetti (Zürich) and Kuang Guodun (Nanning) are thanked for assistance in the field. Alex Cook (Brisbane) and Jiri Frýda (Prague) are acknowledged for helpful peer reviews. The research of A. Kaim has been supported by the Alexander von Humboldt Stiftung. A. Nützel acknowledges funding by the Deutsche Forschungsgemeinschaft (NU 96/6-1, 6-2). Fieldwork was supported by the Swiss National Science Foundation project 200020-113554 (to H. Bucher). This work is a contribution to IGCP project 572 “Permian–Triassic Ecosystems”.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial handling: D. Marty.
Rights and permissions
About this article
Cite this article
Kaim, A., Nützel, A., Bucher, H. et al. Early Triassic (Late Griesbachian) gastropods from South China (Shanggan, Guangxi). Swiss J Geosci 103, 121–128 (2010). https://doi.org/10.1007/s00015-010-0005-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00015-010-0005-5