
Summary
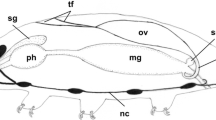
Oogenesis and cleavage ofPimpla turionellae were studied by conventional histological methods. — The development of the oocytes in the polytroph-meroistic ovariole is subdivided into 10 stages. In stage II, when the nutritive chamber is formed, the young oocyte contains achromidium in the ooplasm, which lies on the membrane of the nucleus, being situated in the center of the cell. In the following stages the chromidium is placed in the lower part of the oocyte. In all stages the nutritive cells are connected with the oocyte by a plasm continuation of the oocyte. The nucleus of the oocyte possesses a caryosphere and severalendobodies (“Binnenkörper”) but no nucleoli. In stage V the firstaccessory nuclei appear near the nucleus, which is now situated in the front part of the oocyte. In stage VIvitellogenesis begins along the periphery of the front part of the oocyte. The accessory nuclei are considerably multiplied in the anterior part of the oocyte.Oosome material accumulates around the chromidium, forming a cone-shaped region. In stage VIII the cytoplasm of the nutritive cells streams into the oocyte. The oocyte is filled up with yolk bodies. The oosome material is rounding off. The accessory nuclei dissolve. — The development of the deposited egg was examined until the formation of thepole cells. The oosome reaches the periplasm of the posterior pole, where it spreads. The forming pole cells take up the oosome material.
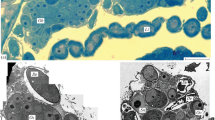
The distribution ofnucleic acids, polysaccharides, proteins andlipids in the cells of the ovariole and egg was analyzed with standard histochemical methods. — In all stages the polyploid nutritive cells deliver RNA substances to the oocyte by means of the plasm continuation. The endobodies of the oocyte nucleus are built up by proteins, RNA is absent. The yolk bodies contain proteins and polysaccharides. The accessory nuclei and the chromidium contain proteins; DNA and RNA could not be demonstrated. In all stages the oosome is rich in RNA. — The egg contains muchfat and glycogen yolk. In the growing oocyte the fat grana appear early. Large amounts of glycogen are deposited in the nearly grown up oocyte.
The problems ofgrowth andtectonics of the oocyte and hypotheses of the oosome function during the proliferative growth of the oocyte and later on during embryonic development as a factor predisposing the fate of pole cells are discussed.
Zusammenfassung
-
1.
Mit herkömmlicherhistologischer Technik ist die Oogenese und die Eurchung bis zur Sonderung der Polzellen vonPimpla turionellae untersucht worden. Die Oocytenentwicklung in der polytroph-meroistischen Ovariole ist in 10 Stadien eingeteilt (Abb. 1) und beschrieben worden (S. 167–174). Die Befunde über die Ei-, Nähr- und Follikelzellen und die besonderen Strukturelemente der Oocyte wie Dotterkugeln, akzessorische Kerne, Chromidium, Binnenkörper, Nährzellensekret, axiale Plasmasäule und Oosom beiPimpla werden mit denen bei anderen Insekten verglichen.
-
2.
Die Altersstadien des abgelegten Eies bis zum Präblastodermstadium mit gesonderten Polzellen (Abb. 6) werden gekennzeichnet. Besonders sind die Veränderungen am Eihinterpol während der Auflösung des Oosoms und die Bildung der Polzellen berücksichtigt worden. Die Beobachtungen an anderen Insekten werden diskutiert. DasOosom erreicht die Periplasmaschicht im Hinterpolbereich und breitet sich entlang dieser aus. Das Oosommaterial wird bei der Sonderung der Polzellen von diesen aufgenommen. Nach der Blastodermbildung sind mit der angewandten Technik nur noch wenige Oosomteilchen in den Polzellen zu entdecken.
-
3.
Mit bewährtenhistochemischen Methoden ist die Verteilung von Nucleinsäuren, Polysacchariden, Proteinen und Fetten innerhalb der Zellen einer Ovariole und im abgelegten Ei geprüft worden.
-
4.
Über den Nährfortsatz der Oocyte wird in allen Stadien bis zur Degeneration der Nährkammer (VIII)RNS-reiches Material von den polyploiden Nährzellen an dieOocyte abgegeben. Nach anfänglicher RNS-Zunahme (II–III) nimmt der RNS-Gehalt, relativ zum Riesenwachstum der Oocyte betrachtet, im Ooplasma stetig ab (IV–VII). Nach Einfließen von Nährzellmaterial in breiter Front (VIII) steigt der RNS-Spiegel im Ooplasma wieder an und bleibt bis nach der Eiablage und Polzellsonderung erhalten.
-
5.
DieDotterkugeln enthaltenProteine undPolysaccharide. Für dieakzessorischen Kerne ist nur derEiweißnachweis positiv verlaufen; DNS- und RNS-Nachweise sind negativ ausgefallen. In denBinnenkörpern des Oocytenkernes hatRNS nicht nachgewiesen werden können.Eiweißmaterial läßt sich dagegengut darstellen. Auch dasChromidium reagiert nur imProteinnachweis positiv. Das Grundplasma desOosoms, das eine rein euplasmatische Differenzierung ist, besteht ausEiweißstoffen. In allen Stadien enthält es aber auchRNS. Im Stadium VII mit ausgeprägtem Oosom-Vorfeld verteilen sich die RNS-Substanzen schüsseiförmig im unteren Oosomgebiet. Das sich zusammenlagernde Oosommaterial der folgenden Stadien reagiert in seiner ganzen Ausdehnung intensiv auf den RNS-Nachweis. Das Oosom bleibt auch während der Furchung RNS-reich.
-
6.
DasPimpla-Ei enthält vielFett- undGlykogendotter. Das Fettdottermaterial erscheint schon in frühen Stadien (III–IV) und erfüllt fortschreitend vom Hinterpol her den Oocytenraum. Das Glykogen tritt in älteren Stadien nach der Nährzelldegeneration plötzlich in hoher Konzentration innerhalb der Oocyte auf. Die Zwischenräume des Reticuloplasmas des abgelegten Eies sind dicht mit den Deutoplasmabestandteilen angefüllt.
-
7.
Die Probleme des Wachstums und der Tektonik der Oocyte werden aufgezeigt. Hypothesen zur Entstehung des Oosoms, zur Funktion des Oosoms in seiner 1. Phase beim Längenwachstum der Oocyte und zur Funktion des Oosoms in der 2. Phase als Substanzspeicher für Stoffwechselprozesse in der Keimbahn werden diskutiert und den Befunden in der Literatur gegenübergestellt.
Similar content being viewed by others

Literatur
Aggarwal, S. K.: Histochemical studies of the carbohydrates, proteins and nucleic acids in the oogenesis ofPeriplaneta americana andChrotogonus trachypterus. Res. Bull. Panjab Univ. N. S.11, 147–153 (1960).
Amy, R. L.: The embryology ofHabrobracon juglandis (Ash.). J. Morph.109, 199–218 (1961).
Bachmann, R., u.H. M. Seitz: Zur histochemischen Darstellung des Histidins mit Diazoniumsalzen. Z. Zellforsch.2, 307–310 (1961).
Beermann, W.: Der Nukleolus als lebenswichtiger Bestandteil des Zellkerns. Chromosoma (Berl.)11, 263–296 (1960).
Bier, K.: Beziehung zwischen Nährzellkerngröße und Ausbildung ribonucleinsäurehaltiger Strukturen in den Oocyten vonFormica rufa rufopratensis minor (Gössw.). Verh. Dtsch. Zool. Ges. Freiburg 1952,46, 369–374 (1952).
—: Über den Saisondimorphismus der Oogenese vonFormica rufa rufopratensis minor (Gössw.) und dessen Bedeutung für die Kastendetermination. Biol. Zbl.73, 170–190 (1954a).
—: Über Phasen gesteigerter Protein- und Kohlenhydrateinlagerung und die Fettverteilung im Hymenopterenovar. Verh. Dtsch. Zool. Ges. Tübingen 1954,48, 422–429 (1954b).
—: Synthese, interzellulärer Transport und Abbau von Ribonucleinsäure im Ovar der StubenfliegeMusca domestica. J. Cell Biol.16, 436–440 (1963a).
—: Die Kern-Plasma-Relation und das Riesenwachstum der Eizellen. Verh. Dtsch. Zool. Ges. München 1963,57, 84–91 (1963b).
—: Autoradiographische Untersuchungen über die Leistungen des Follikelepithels und der Nährzellen bei der Dotterbildung und Eiweißsynthese im Fliegenovar. Wilhelm Roux' Arch. Entwickl.-Mech. Org.154, 552–575 (1963c).
—: Gerichteter Ribonucleinsäuretransport durch das Cytoplasma. Naturwissenschaften51, 418 (1964).
—: Oogenese, das Wachstum von Riesenzellen. Naturwissenschaften54, 189–195 (1967).
— u.D. Ribbert: Struktur und Funktion der Oocytenchromosomen und Nukleolen sowie der Extra-DNS während der Oogenese panoistischer und meroistischer Insekten. Chromosoma (Berl.)23, 214–254 (1967).
Blochmann, F.: Über die Reifung der Eier bei Ameisen und Wespen. Festschrift zur Feier des 500jährigen Bestehens der Ruperto-Carola, dargebr. von d. Natur. med. Verein zu Heidelberg, 143–172 (1884).
Bonhag, P. F.: Histochemical studies of the ovarian nurse tissues and oocytes of the milkweed bug,Oncopeltus fasciatus (Dallas). I. Cytology, nucleic acids, and carbohydrates. J. Morph.96, 381–439 (1955).
—: The origin and distribution of periodic acid-Schiff-positive substances in the oocyte of the earwig,Anisolabis maritima. J. Morph.99, 433–464 (1956).
—: Ovarian structure and vitellogenesis in insects. Ann. Rev. Entomol.3, 137–160 (1958).
Bronskill, J. F.: Embryology ofPimpla turionellae (L.) (Hymenoptera, Ichneumonidae). Canad. J. Zool.37, 656–688 (1959).
Bronskill, J. F.: Embryogenesis ofMesoleius tenthredinis Morl. (Hymenoptera, Ichneumonidae). Canad. J. Zool.42, 439–453 (1964).
Brown, D. D., andE. Littna: Variations in the synthesis of stable RNA's during oogenesis and development ofXenopus laevis. J. molec. Biol.8, 688–695 (1964).
Bruiyan, N. I., andS. A. Shafiq: The differentiation of the posterior pole-plasm in the house flyMusca vicina (Mac.). Exp. Cell Res.16, 427–429 (1959).
Bucher, N.: Experimentelle Untersuchungen über die Beziehungen zwischen Keimzellen und somatischen Zellen im Ovar vonDrosophila melanogaster. Rev. suisse Zool.64, 91–188 (1957).
Buchner, P.: Vergleichende Eistudien. 1. Die akzessorischen Kerne des Hymenoptereneies. Arch. mikr. Anat.91, 1–202 (1918).
Counce, S. J.: Comparative ontogeny of the polar granules inDrosophila. Anat. Rec.134, 546–547 (1959).
—: Developmental morphology of polar granules inDrosophila melanogaster. J. Morph.112, 129–145 (1963).
—, andD. A. Ede: The effect on embryogenesis of a sexlinked female sterility factor inDrosophila melanogaster. J. Embryol. exp. Morph.5, 404–421 (1957).
Cruickshank, W. J.: Formation and possible function of the accessory yolk nuclei inAnagasta (=Ephestia) Kühniella. Nature (Lond.)201, 734–735 (1964).
Elsdale, T. R., M. Fischberg, andS. Smith: A mutation that reduces nucleolar number inXenopus laevis. Exp. Cell Res.14, 642–643 (1958).
Engels, W.: Der zeitliche Ablauf von Protein- und Kohlenhydratsynthesen in der Oogenese beiApis mellifica L. Zool. Anz., Suppl.29, 243–251 (1966).
—, u.K. Bier: Zur Glykogenspeicherung während der Oogenese und ihrer vorzeitigen Auslösung durch Blockierung der RNS-Versorgung. (Untersuchungen anMusca domestica L.) Wilhelm Roux' Arch. Entwiekl.-Mech. Org.158, 64–88 (1967).
—, u.W. Drescher: Einbau von H3-d-Glucose während der Oogenese beiApis mellifica L. Experientia (Basel)20, 445–447 (1964).
Gatenby, J. B.: Polyembryony in parasitic hymenoptera: a review. Quart. J. micr. Sci.63, 175–196 (1919).
—: The cytoplasmic inclusions of germ cells. Part VI. On the origin and probable constitution of the germ cell determinant ofApanteles glomeratus, with a note on the secondary nuclei. Quart. J. micr. Sci.64, 133–154 (1920).
Geigy, R.: Action de l'ultraviolet sur le pole germinal dans l'oeuf deDrosophila melanogaster (castration et mutabilite). Rev. suisse Zool.38, 187–288 (1931).
Geyer-Duszynska, I.: Experimental research on chromosome elimination in Cecidomyidae (Dipt.). J. exp. Zool.141, 391–448 (1959).
—: Spindle disappearance and chromosome behavior after partial-embryo irradiation in Cecidomyidae. Chromosoma (Berl.)12, 233–247 (1961).
—: Genetic factors in oogenesis and spermatogenesis in Cecidomyidae. Reprinted from Chromosomes Today1, 174–178 (1966).
Gill, K. S.: Developmental genetic studies on oogenesis inDrosophila melanogaster. J. exp. Zool.152, 251–277 (1963).
Hasper, M.: Zur Entwicklung der Geschlechtsorgane vonChironomus. Zool. Anz.35, 737–743 (1910).
—: Zur Entwicklung der Geschlechtsorgane vonChironomus. Zool. Jb., Abt. Anat. u. Ontog.31, 543–612 (1911).
Hegner, R. W.: Effects of removing the germ cell determinants from the eggs of some chrysomelid beetles. Biol. Bull.16, 19–26 (1909a).
—: The origin and early history of the germ cells in some chrysomelid beetles. J. Morph.20, 231–296 (1909b).
Hegner, R. W.: Experiments with chrysomelid beetles. 3. Effects of killing parts of the eggs ofLeptinotarsa decemlineata. Biol. Bull.20, 237–251 (1911).
Hogben, L.: Studies on synapsis. 1. Oogenesis in the Hymenoptera. Proc. roy. Soc. B91, 268–293 (1920).
Hopkins, C. R.: The histochemistry and fine structure of the accessory nuclei in the oocyte ofBombus terrestris. Quart. J. micr. Sci.105, 475–480 (1964).
Hotchkiss, R. D.: A microchemical reaction resulting in the staining of polysaccharide structures in fixed tissue preparations. Arch. Biochem.16, 131–141 (1948).
Ivanova-Kazas, O. M.: Adaptions to parasitism in the embryonic development of the ichneumon fly,Prestwichia aquatica (Hymenoptera). [In Russian.] Zool. Zhur.29, 530–544 (1950).
Jazdowska-Zagrodzinska, B.: Experimental studies on the role of “polar granules” in the segregation of pole cells inDrosophila melanogaster. J. Embryol. exp. Morph.16, 391–399 (1966).
Johannsen, O. A., andF. H. Butt: Embryology of insects and myriapods, 1. edit. New York and London: McGraw-Hill Book Co. 1941.
Jung, E.: Untersuchungen am Ei des SpeisebohnenkäfersBruchidius obtectus Say(Coleoptera). 1. Teil. Z. Morph. Ökol. Tiere56, 444–480 (1966).
Jura, C., A. Krzysztofowicz, andB. Weglarska: Histochemical investigations on the embryonic development ofMelasoma populi L. (Chrysomelidae) I. Zool. polon.8, 201–216 (1958).
—: Histochemieal investigations on the embryonic development ofMelasoma populi L. (Chrysomelidae, Coleoptera) II. Zool. polon.9, 1–15 (1959).
Kaspar, F.: Beiträge zur Laborbiologie vonPimpla turionellae (L.) undItoplectis maculator (Fabr.). Zulassungsarbeit für das Lehramt an den Gymnasien, Zool. Institut Würzburg (1962).
Katscher, H.: Bilaterale Bandbestrahlung des Eies der SchlupfwespePimpla turionellae in Blastodermstadien mit UV-Licht. Zulassungsarbeit für das Lehramt an den Mittelschulen, Zool. Institut Würzburg (1963).
Kaufmann, B. P., M. R. McDonald, M. H. Bernstein, R. C. v. Borstel, andN. K. Das: Patterns of organization of cellular materials. Carnegie Inst. of Washington, Year Book52, 238–248 (1953).
King, R. C.: Oogenesis in adultDrosophila melanogaster. IX. Growth24, 265–323 (1960).
—, andR. C. Devine: Oogenesis in adultDrosophila melanogaster. VII. Growth22, 299–326 (1958).
Kraft, A. v.: Entwicklungsgeschichtliche und histochemische Untersuchungen zur Oogenese vonTachycines (Insecta, Saltatoria). Zool. Jb., Abt. Anat. u. Ontog.78, 457–558 (1960).
Krainska, M.: A morphological and histochemical study in the gall-flyCynips folii. Quart. J. micr. Sci.102, 119–129 (1961).
Krause, G.: Einzelbeobachtungen und typische Gesamtbilder der Entwicklung von Blastoderm und Keimanlage im Ei der GewächshausschreckeTachycines asynamorus Adelung. Z. Morph. Ökol. Tiere34, 1–78 (1938).
—: Die Eitypen der Insekten. Biol. Zbl.59, 495–536 (1939).
—: Induktionssysteme in der Embryonalentwicklung von Insekten. Ergebn. Biol.20, 159–198 (1958).
- Preformed ooplasmic reaction systems in insect eggs. Symposium on germ cells and development, p. 302–337. Inst. Intern. d'Embryologie and Fondazione A. Baselli (1961).
—, u.J. Krause: Die Regulation der Embryonalanlage vonTachycines (Saltatoria) im Schnittversuch. Zool. Jb., Abt. Anat. u. Ontog.75, 481–550 (1957).
—, andK. Sander: Ooplasmatic reaction systems in insect embryogenesis. Advanc. Morphogenes.2, 259–299 (1962).
Mahowald, A. P.: Fine structure of pole cells and polar granules inDrosophila melanogaster. J. exp. Zool.151, 201–215 (1962).
Mahr, E.: Bewegungsvorgänge im Dotter-Entoplasmasystem des Insekteneies. Naturwissenschaften44, 226–227 (1957).
Martin, F.: Zur Entwicklungsgeschichte des polyembryonalen ChalcidiersAgeniaspis (Encyrtus) fuscicollis D. Z. wiss. Zool.110, 419–479 (1914).
Maul, V.: Dynamik und Erbverhalten plasnaatischer Eibereiche der Honigbiene. Zool. Jb., Abt. Anat. u. Ontog.84, 63–166 (1967).
Müller, H.: Die Entwicklungsleistungen abgeschnürter Eiteile bei der SchlupfwespePimpla turionellae L. Zulassungsarbeit für das Lehramt an den Gymnasien, Zool. Institut Würzburg (1963).
Mukerji, R. N.: The “nucleal-reaction” inApanteles sp., with special reference to the secondary nuclei and the germ-cell determinant of the egg. Proc. roy. Soc. B106, 131–139 (1930).
Nath, V., andD. R. Mehta: Studies in the origin of yolk. III. Oogenesis of the fireflyLuciola gorhami. Quart. J. micr. Sci.73, 7–24 (1930).
Netzel, H.: Die Ausprägung von Polarität und Bilateralsymmetrie in den Oocyten vonGryllus domesticus L. Wilhelm Roux' Arch. Entwickl.-Mech. Org.160, 119–166 (1968).
Oelhafen, F.: Zur Embryogenese vonCulex pipiens: Markierungen und Exstirpationen mit UV-Strahlenstich. Wilhelm Roux' Arch. Entwickl.-Mech. Org.153, 120–157 (1961).
Ovchnnikova, L. P., andG. V. Selivanova: Examination of applicability of gallocyaninchromalaun staining for the aims of cytophotometrical measurement of RNA quantity in the cytoplasm. Citologija (Moskau)6, 387–388 (1964).
Palm, N. B.: Normal and pathological histology of the ovaries inBombus (Latr.). Opusc. Entomol., Suppl.7, 1–101 (1948).
Patterson, J. T.: The development of Paracopidosomopsis. J. Morph.36, 1–69 (1921).
Peacock, A. D., andR. A. R. Gresson: The roles of the nurse-cells, oocytes and follicle cells in tenthredinid oogenesis. Quart. J. micr. Sci.71, 541–561 (1928).
Pearse, A. G. E.: Histochemistry, 2. edit. London: J. &. A. Churchill, Ltd. 1961.
Przelecka, A., andA. Dutkowski: Autoradiographic investigation of incorporation of fatty acids into the lipids of insect ovarioles. Bull. Acad. pol. Sci., Ser. Sci. biol.8, 573–575 (1965).
Rabes, J.: Untersuchungen über die weiblichen Geschlechtsorgane vonApanteles, mit besonderer Berücksichtigung der accessorischen Kerne. Z. Zellforsch.23, 701–745 (1935).
Ramamurty, P. S.: Über die Herkunft der RNS in den wachsenden Eizellen der SkorpionsfliegePanorpa communis. Naturwissenschaften50, 383–384 (1963).
Reith, F.: Versuche über die Determination der Keimesanlage beiCamponotus ligniperda. Z. wiss. Zool.139, 664–734 (1931).
Romeis, B.: Mikroskopische Technik, 15. Aufl. München: R. Oldenbourg 1948.
Sachtleben, H.: Über die Entwicklung der Geschlechtsorgane vonChironomus mit besonderer Berücksichtigung der die Keimbahn begleitenden Substanzen. Diss. München 1918.
Schnetter, W.: Experimente zur Analyse der morphogenetischen Funktion der Ooplasmabestandteile in der Embryonalentwicklung des Kartoffelkäfers (Leptinotarsa decemlineata). Wilhelm Roux' Arch. Entwickl.-Mech. Org.155, 637–692 (1965).
Schrader, F., andC. Leuchtenberger: The origin of certain nutritive substances in the eggs of Hemiptera. Exp. Cell Res.3, 136–146 (1952).
Seidel, F.: Die Geschlechtsorgane in der embryonalen Entwicklung vonPyrrhocoris apterus L. Z. Morph. Ökol. Tiere1, 429–506 (1924).
Silvestri, F.: Insect polyembryony and its general biological aspects. Bull. Mus. Comp. Zool.81, 469–498 (1937).
Spaknhof, L.: Einführung in die Praxis der Histochemie, 1. Aufl. Jena: VEB Gustav Fischer 1964.
Telfer, W. H.: The mechanism and control of yolk formation. Ann. Rev. Entomol.10, 161–184 (1965).
Ullmann, S. L.: Epsilon and polar granules inDrosophila pole cells and oocytes, p. 455–456. Prague: Publishing House of the Czechoslovak Academy of Sciences 1964.
Wolf, B. E.: Über die Entstehung von accessorischen Kernen im Ei vonPhryne fenestralis (Dipt.). Moderne Biologie, Festschrift zum 60. Geburtstag vonHans Nachtsheim, S. 251–274 (1950).
Wolf, R.: Der Eeinbau des Oosoms normaler und zentrifugierter Eier der GallmückeWachtliella persicariae L. (Diptera). Wilhelm Roux' Arch. Entwickl.-Mech. Org.158, 459–462 (1967).
Yao, T.: Cytochemical studies on the embryonic development ofDrosophila melanogaster. Quart. J. micr. Sci.90, 401–409 (1949).
Author information
Authors and Affiliations
Additional information
Meinem verehrten Lehrer, Herrn Prof. Dr.Gerhard Krause, danke ich herzlich für die Anregung und Leitung der Arbeit. Durchgeführt dank der Unterstützung der Deutschen Forschungsgemeinschaft durch Leihgaben.
Rights and permissions
About this article
Cite this article
Meng, C. Strukturwandel und histochemische Befunde insbesondere am Oosom während der Oogenese und nach der Ablage des Eies von Pimpla turionellae L. (Hymenoptera, Ichneumonidae). W. Roux' Archiv f. Entwicklungsmechanik 161, 162–208 (1968). https://doi.org/10.1007/BF00585970
Received:
Issue Date:
DOI: https://doi.org/10.1007/BF00585970