
Summary
-
1.
In the rotifer Brachionus calyciflorus a pair of posterolateral spines (PL) is subjected to extensive temporal variation. The length of these spines may vary from complete absence to more than lorica length. Production of spines is induced by starvation, low temperature, and a substance produced by the predatory rotifer Asplanchna. The aim of this study was, by comparing the effectiveness of these factors, to gain insight into the physiological mechanisms involved, and to assess their relative significance in natural populations.
-
2.
The other spines of the lorica vary in accordance with PL (Fig. 1), however, they never disappear. In the absence of PL, they continue to vary in the same sense (Table 2).
-
3.
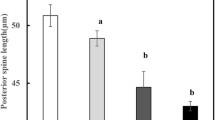
In the laboratory starvation and low temperature induce only short PL. To elicit long spines the Asplanchna-substance is always necessary (Fig. 2). The effects of the three factors are additive. High temperature seems to inhibit spine production. The type of the inducing factor has no recognizable influence on the variability of the reaction (Figs. 3 and 4); even under conditions where the reaction is maximal no reduction of the standard deviation is found.
-
4.
There is no clearly recognizable short-term inheritance of spine length: almost the full reaction to each of the inducing factors is realized in the first generation following induction (Fig. 5).
-
5.
Low temperature increases body size. Analysis shows that this temperature effect consists of two components, one acting on egg size, the other on the animals growth after hatching (Figs. 6 and 7).
-
6.
The final spine length is always determined before hatching (Fig. 5). The inducing factors determine the body proportions of the neonates; after hatching the spine length grows allometrically. The average growth rate of the PL relative to that of the body was k=0.54, i.e. the growth rate of the spines is smaller than that of the body (negative allometry). The value of k appears to be a constant, at least it is not influenced by the type of spine induction, the initial spine length, and the conditions after hatching (Table 4 and Fig. 9).
-
7.
Using the allometric relation, an index of spine length (I D ) was defined, which is independent of body size. ID permits the quantitative comparison of populations with different age structures, and of age (size) classes within the same sample. Differences between the I D -values of the age classes of a sample indicate fluctuations of spine inducing factors (Fig. 10). It is thus possible to estimate the strength of spine inducing factors during previous periods by studying a single plankton sample (Figs. 11 and 12).
-
8.
A reproducible bio-assay was developed to test the spine inducing activity of the medium. A relative unit of Asplanchna-substance (as) was defined. The quantity of substance produced per unit volume and per unit time is proportional to Asplanchna density and is influenced by temperature (within 10 to 25\dg C: Q 10=4). The amount of substance produced by 1 Asplanchna in 25 sec at 20\dg C in 1 ml medium will induce just recognizable spines in the experimental clone of Brachionus. Further data of substance production at 10, 15, and 25\dg C are given in Table 6. The substance decomposes exponentially (Fig. 14a). The velocity of this decomposition is influenced by temperature. Within the temperature range studied the half-life varies from 1 to 3 days (Fig. 14b). Theoretically, a specific equilibrium concentration of active substance corresponds to each population density of Asplanchna. This concentration can be computed using the data of production and decomposition (Table 6). However, in experimental populations as well as in the field, the actual concentrations were always considerably lower than expected on the basis of the Asplanchna density. This fact suggests that Brachionus incorporates the substance. Therefore, a pragmatic correction of the theoretical balance of the Asplanchna-substance was introduced by assuming that there is no accumulation of substance for more than 24 hours. With this correction, there is good agreement between the expected and the observed spine-indices in laboratory and field populations.
-
9.
A quantitative comparison of the effectiveness of the three spine inducing factors shows that under natural conditions, long spines (mean I D >0.3) cannot be induced by temperature and starvation, but must be due to Asplanchna-substance (Fig. 17).
-
10.
In the field the Asplanchna-substance is the most important agent. Bioassays show that water of all ponds containing Asplanchna possesses spine inducing activity (Table 7). Both Asplanchna sieboldi (s. 1.) and A. priodonta are effective. The spine-indices of neonate Brachionus show a good correlation with the prevailing Asplanchna density (Figs. 20\2-22). The correlations of the indices of adult Brachionus show time-lags which correspond to the age of the animals. After the disappearance of Asplanchna, the spine-indices decrease with age-dependent time-lag.
The spine-inducing influence of temperature is lower in the field than in the laboratory (Fig. 23). The possible effects of unknown factors in the field are discussed.
-
11.
The reaction norms of two stocks from different ponds were tested. They exhibited remarkable and significantly different I D -values at identical conditions of induction (Fig. 2), which occured at all three inducing factors.
-
12.
Hypothetical mechanisms of spine production are discussed; possible differences in the physiological mechanisms of the three spine-inducing factors are indicated. Some aspects of the adaptive value of spine length variation are discussed.
Zusammenfassung
-
1.
Die Temporalvariation von Brachionus calyciflorus betrifft vor allem ein Paar Posterolateraldornen (PL), die in ihrer Ausbildung von vollst\:andigem Fehlen bis zu K\:orperl\:ange variieren k\:onnen. Die Bildung dieser Dornen wird durch Hunger, tiefe Temperatur und einen Stoff hervorgerufen, den das r\:auberische R\:adertier Asplanchna ins Medium abgibt. Diese drei Faktoren wurden quantitativ vergleichend auf Unterschiede in der Art ihrer Wirksamkeit und auf ihre Bedeutung f\:ur die Variation im Freiland untersucht.
-
2.
Es besteht eine positive Korrelation zwischen der L\:ange der PL und derjenigen der anderen Dornen, die stets vorhanden sind (Abb. 1). Diese anderen Dornen variieren auch noch bei Fehlen der PL, und zwar in gleichem Sinne (Tabelle 2).
-
3.
Hunger und tiefe Temperatur k\:onnen nur kurze PL induzieren, zur Ausbildung langer PL ist der Asplanchna-Stoff notwendig (Abb. 2). Die Effekte der drei Faktoren k\:onnen sich \:uberlagern. Hohe Temperatur scheint dabei hemmend auf die Dornenbildung einzuwirken. Die Art der Inkubation hat keinen erkennbaren Einflu\sB auf die Variation der Reaktion (Abb. 3 und 4); auch bei maximaler Reaktion ist die Streubreite nicht verringert.
-
4.
Es gibt keine deutliche Tradierung des Variationstyps: bei allen drei Faktoren manifestiert sich die nahezu volle Reaktion auf einen induzierenden Reiz in der n\:achsten Generation (Abb. 5).
-
5.
Die Temperatur hat einen Einflu\sB auf die K\:orpergr\:o\sBe: je tiefer die Temperatur ist, desto gr\:o\sBer sind die Tiere. Dieser Temperatureinflu\sB besteht aus einer pr\:a- und einer postnatalen Komponente (Abb. 6 und 7).
-
6.
Die Determination der Dornenl\:ange erfolgt in allen F\:allen vor der Geburt (vgl. Abb. 5). Die dorneninduzierenden Faktoren bestimmen die Proportionen der Neonaten, nach der Geburt erfolgt das Dornenwachstum negativ allometrisch mit einer konstanten Allometrierate k=0,54, die von der Art der Dorneninduktion, der Gr\:o\sBe der Dornenanlage und den Aufzuchtbedingungen unabh\:angig ist (Tabelle 4 und Abb. 9).
-
7.
Mit Hilfe der Allometriebeziehung wird ein Dornenindex I D definiert, der von der K\:orpergr\:o\sBe der Tiere unabh\:angig ist. Mit ihm k\:onnen die Dornenausbildungen von Populationen verglichen werden, auch wenn diese unterschiedliche Altersstruktur besitzen. Ebenso sind auf diese Weise die verschiedenen Alters- (bzw. Gr\:o\sBen-) Klassen einer Stichprobe bez\:uglich ihrer Dornenausbildung miteinander vergleichbar. Unterschiede in den Dornenindices verschiedener Altersklassen einer Stichprobe weisen darauf hin, da\sB ein dorneninduzierender Faktor (wie etwa der Asplanchna-Stoff) Schwankungen unterworfen war (Abb. 10). Auf diese Weise kann man aufgrund einer einzigen Planktonprobe Aussagen \:uber die Vorgeschichte der dorneninduzierenden Faktoren in den letzten Tagen und Wochen machen (Abb. 11 und 12).
-
8.
Es wurde ein reproduzierbarer Test auf die dorneninduzierende Aktivit\:at eines Mediums ausgearbeitet. Mit Hilfe deses Testes lie\sB sich eine relative Einheit der Asplanchna-Stoffmenge (as) definieren. Die pro Volumen-und Zeiteinheit gebildete Stoffmenge ist der Asplanchnen-Dichte proportional und h\:angt von der Temperatur ab (im Bereich von 10\2-25\dg C: Q 10=4). Ein Asplanchna produziert z.B. bei 20\dg C in 25 sec die Stoffmenge, die in 1 ml gerade erkennbare Dornenbildung bewirkt. Entsprechende Werte werden f\:ur 10, 15 und 25\dg C angegeben (Tabelle 6). Der Stoff zerf\:allt jedoch exponentiell (Abb. 14a). Diese nat\:urliche Stoffzersetzung ist ebenfalls temperaturabh\:angig (Halbwertszeit je nach Temperatur 1\2-3 Tage, Abb. 14b). Jeder Asplanchnen-Dichte kann theoretisch eine Gleichgewichtskonzentration an Stoff zugeordnet werden, die sich aus Daten der Produktion und des Zerfalls berechnen l\:a\sBt (Tabelle 6). In Populationsversuchen wie im Freiland stellten sich jedoch die theoretisch errechneten Stoffkonzentrationen nicht ein, m\:oglicherweise weil die Brachionus den Stoff inkorporieren. Eine pragmatische Korrektur der theoretischen Asplanchnastoff-Bilanz (keine Akkumulation \:uber 24 Std hinaus) ergab gute \:Ubereinstimmung zwischen den erwarteten und den beobachteten Dornenindices (in Experimental-und Freilandpopulationen).
-
9.
Ein quantitativer Vergleich der Effektivit\:at der drei dorneninduzierenden Faktoren (Abb. 17) ergab, da\sB lange Dornen (mittl. I D >0,3) weder durch tiefe Temperatur noch durch Hunger induziert werden k\:onnen, sondern allein durch die Asplanchna-Substanz hervorgerufen werden.
-
10.
Auch im Freiland ist der Asplanchna-Stoff das wichtigste induzierende Agens. Labortests zeigen, da\sB Wasser aus Teichen, in denen Asplanchnen vorkommen, dorneninduzierende Aktivit\:at besitzen (Tabelle 7). Dies gilt f\:ur Asplanchna sieboldi (s. 1.) und f\:ur A. priodonta. Die Dornenindices der Neonaten zeigen in ihrem zeitlichen Verlauf eine gute Korrelation mit der Asplanchnen-Dichte (Abb. 20\2-22), die der Adulten zeigen eine Phasenverschiebung, die dem Alter der Tiere entspricht. Verschwinden die Asplanchnen aus dem Biotop, so gehen die Dornenindices mit einer zeitlichen Verz\:ogerung von einigen Tagen ebenfalls zur\:uck.
Der Einfluß der Temperatur ist im Freiland schwächer als im Labor (Abb. 23). Die mögliche Wirksamkeit unbekannter Faktoren im Freiland wird diskutiert.
-
11.
Mittels Labortests lie\sBen sich Unterschiede in der Reaktionsnorm zweier St\:amme von verschiedenen Fundorten nachweisen (Abb. 2). Diese betreffen (unabh\:angig von der Art des Induktionsfaktors) die mittleren Dornenindices, mit denen die Tiere auf bestimmte Induktionsreize reagieren.
-
12.
Es werden denkbare Mechanismen der Dornenbildung besprochen und m\:ogliche Unterschiede in der physiologischen Wirkungsweise der drei dorneninduzierenden Faktoren diskutiert. Einige Aspekte, die die Funktion der zyklomorphen Dornenbildung betreffen, werden angedeutet.
Similar content being viewed by others

Abbreviations
- a :
-
Ordinatenabschnitt der Allometriegeraden
- AL:
-
Anterolateraldornen
- AM:
-
Anteromediandornen
- as:
-
Asplanchnastoff-Einheit
- As:
-
Asplanchna
- As+ :
-
Asplanchnastoff-haltig
- As- :
-
Asplanchnastoff-frei
- Cn:
-
Brachionus calyciflorus, Stamm Cn
- D :
-
(Posterolateral-) Dornen-Länge
- E :
-
Längsdurchmesser der Subitaneier
- FD:
-
Futterdosis
- I D :
-
Dornenindex
- k :
-
Allometrierate
- k 1, k 2 :
-
unterer u. oberer Schwellenwert der As-Stoff-Konzentration
- K o , K m u. K n :
-
oberer, mittlerer u. unterer Konzentrationsbereich des As-Stoffes
- L :
-
Lorica-Länge
- L A :
-
Lorica-Länge der Adulten
- L N :
-
Lorica-Länge der Neonaten
- NB:
-
Normal-Versuchsbedingungen
- PL:
-
Posterolateraldornen
- PM:
-
Posteromediandornen
- T-3:
-
Brachionus calyciflorus, Stamm T-3
- T :
-
Halbwertszeit des Asplanchna-Stoffes
- z :
-
Zerfallskonstante des Asplanchna-Stoffes
Literatur
Ahlstrom, E. H.: A revision of the rotatorian genera Brachionus and Platyias with descriptions of one new species and two new varieties. Bull. Amer. Mus. Nat. Hist. 77, 143–184 (1940).
Arora, H. C.: Cyclomorphosis (form variation) in some species of Indian planktonic Rotatoria. Int. Rev. ges. Hydrobiol. 51, 623–632 (1966).
Amrén, H.: Temporal variation of the rotifer Keratella quadrata (Müll.) in some ponds of Spitsbergen. Zool. Bidr. Uppsala 36, 193–208 (1964).
Beauchamp, P. de: Coup d'oiel sur les recherches recentes relatives aux rotifères. Bull. biol. France Belg. 62, 51–125 (1928).
— Méthodes pour l'étude de la variabilité morphologique des rotifères. C. R. Soc. Biol. (Paris) 125, 445–447 (1937a).
— Quelques remarques sur la variation de Brachionus pala Ehrenberg. C. R. Soc. Biol. (Paris) 125, 448–450 (1937b).
— Un facteur de la variabilité chez les rotifères du genre Brachionus. C. R. Acad. Sci. (Paris) 234, 573–575 (1952a).
— Variation chez les rotifères du genre Brachionus. C. R. Acad. Sci. (Paris) 235, 1355–1356 (1952b).
Buchner, H., Mulzer, R., Rauh, F.: Untersuchungen über die Variabilität der Rädertiere. I. Problemstellung und vorläufige Mitteilung über die Ergebnisse. Biol. Zbl. 76, 289–315 (1957).
Dieffenbach, H.: Biologische Untersuchungen an Rädertieren in Teichgewässern. Int. Rev. ges. Hydrobiol., Biol., Suppl. 4, 1–42 (1911).
Erman, L. A.: Feeding habits of Rotifera from the quantitative aspect. Zool. Zh. 35, 965–971 (1956).
— Cyclomorphosis and feeding of Plankton Rotifera. Zool. Zh. 41, 998–1003 (1962a).
— On the quantitative aspects of feeding and selection of food in the planktonic rotifer Brachionus calyciflorus Pallas. Zool. Zh. 41, 34–47 (1962b).
Gilbert, J. J.: Rotifer ecology and embryological induction. Science 151, 1234–1237 (1966).
— Asplanchna and postero-lateral spine production in Brachionus calyciflorus. Arch. Hydrobiol. 64, 1–62 (1967).
— Waage, J. K.: Asplanchna, Asplanchna-substance, and postero-lateral spine length variation of the rotifer Brachionus calyciflorus in a natural environment. Ecology 48, 1027–1031 (1967).
Halbach, U.: Räuber und ihre Beute: Der Anpassungswert von Dornen bei Rädertieren. Naturwissenschaften 56, 142–143 (1969).
Halbach, U.: Das Zusammenwirken von Konkurrenz und Räuber-Beute-Beziehungen bei Rädertieren. Verh. dtsch. Zool. Ges. (1969) (im Druck).
Halbach, U.: Einfluß der Temperatur auf die Populationsdynamik des planktischen Rädertieres Brachionus calycifloris Pallas. Oecologia (Berl.) (1970) (im Druck).
Hutchinson, G. E.: A treatise on limnology. II. New York-London-Sydney: Wiley 1967.
Jacobs, J.: Cyclomorphosis in Daphnia galeata mendotae Birge, a case of environmentally controlled allometry. Arch. Hydrobiol. 58, 7–71 (1961).
Kikuchi, K.: Formation of the lateral spines in Brachionus pala. J. Fac. Sci. Tokyo Univ., sec. 4, Zool. 2, 163–169 (1931).
Magis, N.: Donées nouvelles sur la variabilité morphologique de Brachionus falcatus Zacharias. Ann. Soc. R. Zool. Belg. 97, 103–129 (1967).
Nayar, C. K. G.: Morphometric studies on the rotifer Brachionus calyciflorus Pallas. Curr. Sci. 33, 469–470 (1964).
— Cyclomorphosis of Brachionus calyciflorus Pallas. Hydrobiologia 25, 538–544 (1965).
Rauh, F.: Untersuchungen über die Variabilität der Rädertiere. III. Die experimentelle Beeinflussung der Variation von Brachionus calyciflorus und Brachionus capsuliflorus. Z. Morph. Ökol. Tiere 53, 61–106 (1963).
Sachse, R.: Beiträge zur Biologie litoraler Rädertiere. Int. Rev. ges. Hydrobiol., Biol. Suppl. 4, 43–93 (1911).
Schneider, P.: Sur la variabilité de Brachionus pala Ehrenberg dans les conditions expérimentales. C. R. Acad. Sci. (Paris) 125, 450–452 (1937).
Voigt, M.: Rotatoria. Die Rädertiere Mitteleuropas. Berlin: Borntraeger 1957.
Wesenberg-Lund, C.: Contributions to the biology of the Rotifera. II. The periodicity and sexual periods. K. danske vidensk. Selsk. 9, (2), 1–230 (1930).
Whitney, D. D.: The transformation of Brachionus pala into Brachionus amphiceros by sodium silicata. Biol. Bull., mar. biol. Lab., Woods Hole 31, 113–120 (1916).
Author information
Authors and Affiliations
Additional information
Diese Arbeit ist Teil einer Inaugural-Dissertation zur Erlangung der Doktorwürde der Naturwissenschaftlichen Fakultät der Ludwig Maximilians-Universität München.
Rights and permissions
About this article
Cite this article
Halbach, U. Die ursachen der Temporalvariation von Brachionus calyciflorus Pallas (Rotatoria). Oecologia 4, 262–318 (1970). https://doi.org/10.1007/BF00377250
Received:
Issue Date:
DOI: https://doi.org/10.1007/BF00377250